Intradermal Delivery of Dendritic Cell-Targeting Chimeric mAbs Genetically Fused to Type 2 Dengue Virus Nonstructural Protein 1

 ,
,  , , , , , and
, , , , , and 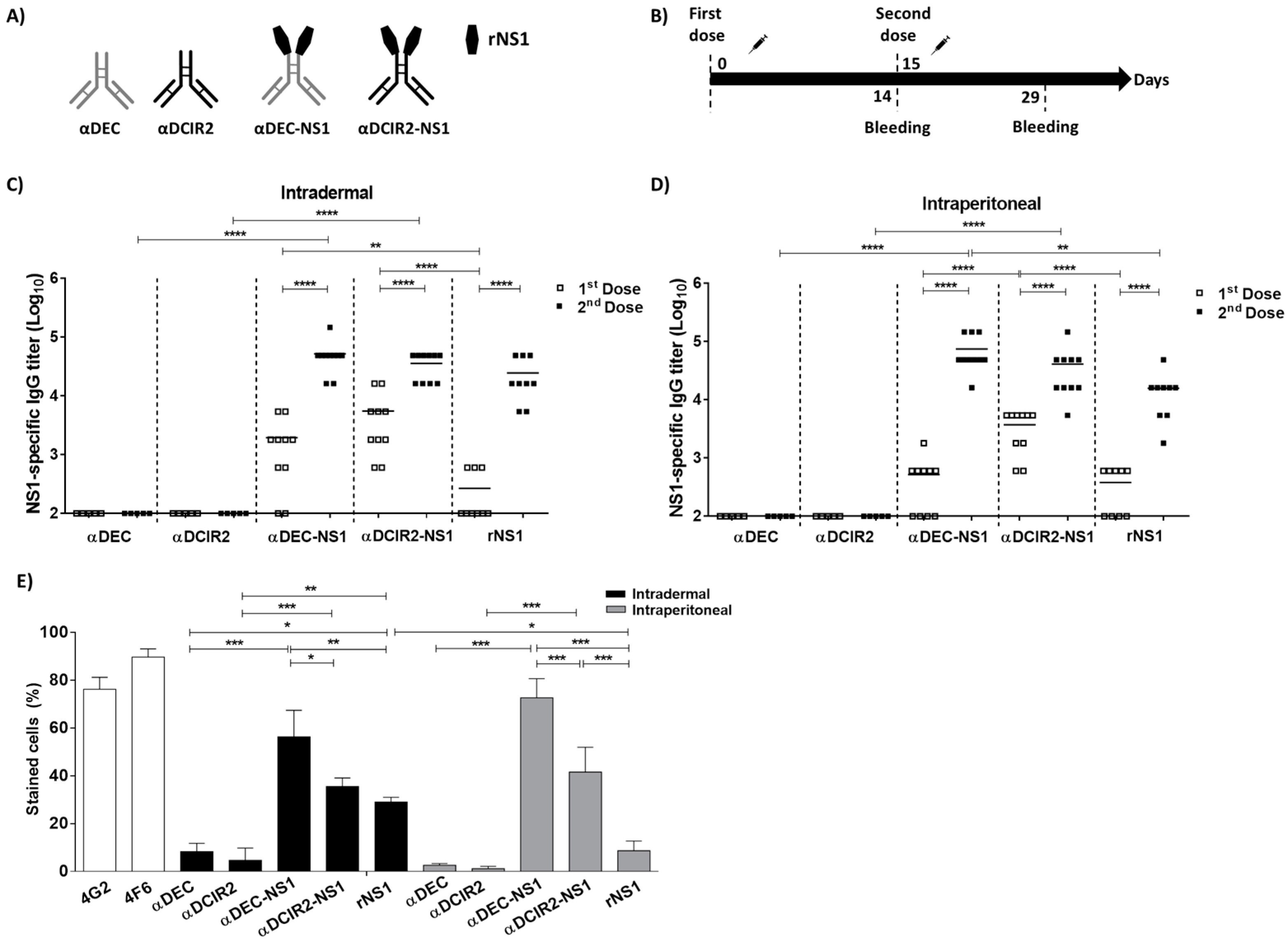{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Ethics Statement
2.2. Virus and Cell Lines
2.3. Expression and Purification of the Recombinant mAbs and DENV2 NS1
2.4. Immunization Regimens
2.5. ELISA
2.6. Antibody Avidity Determination
2.7. Flow Cytometry Analysis
2.7.1. Recognition of the Native DENV2 NS1 Protein by the Immune Serum Samples
2.7.2. Binding of Mouse Serum to Platelets
2.8. Inhibition of Platelet Aggregation
2.9. Transendothelial Electrical Resistance (TEER) Assay
2.10. Statistical Analyses
3. Results
3.1. Intradermal Delivery of the NS1-Based DC-Targeting mAbs
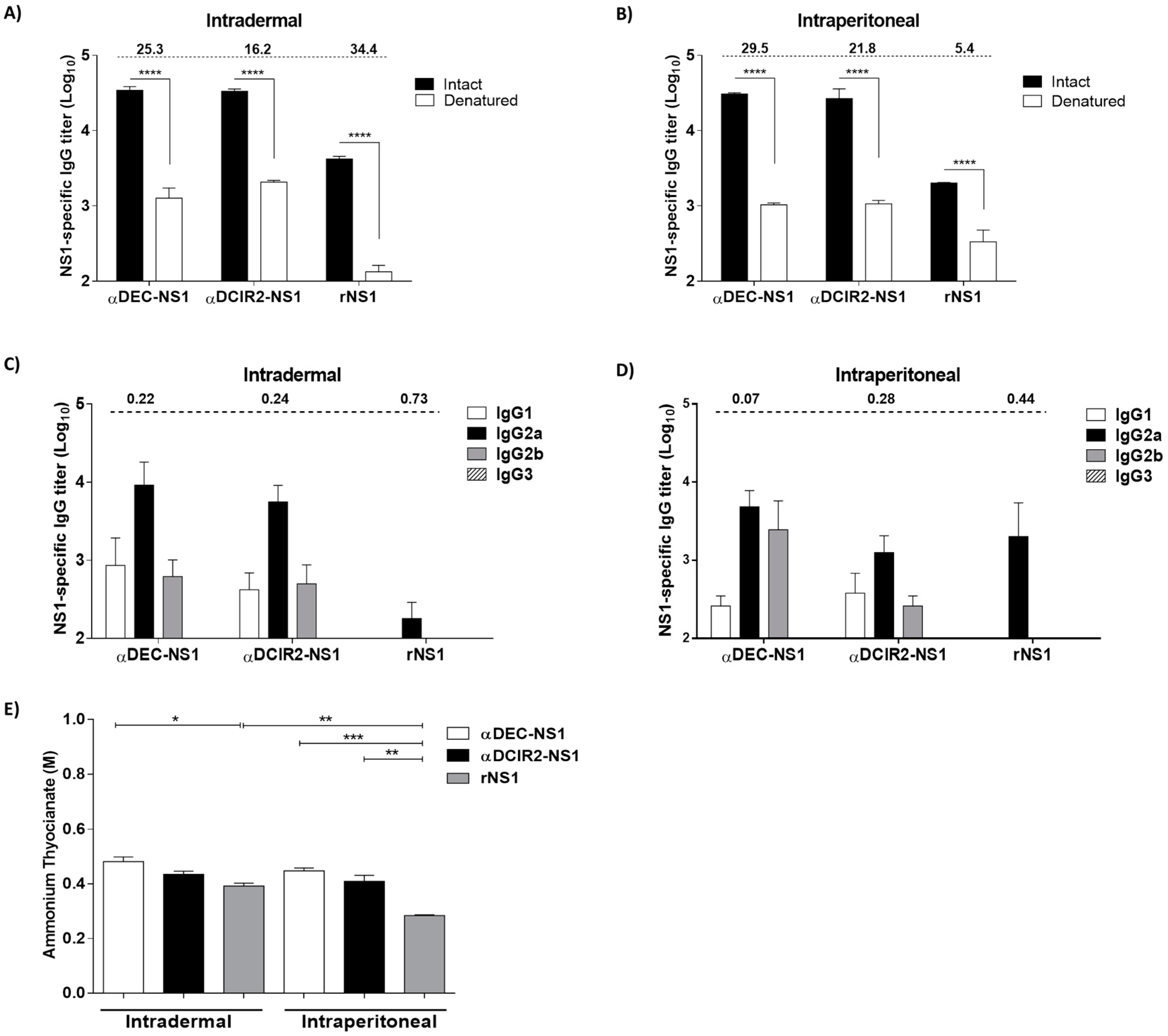
3.2. Epitope Specificity, IgG Subclass, and Antigen Avidity of Anti-NS1 Antibodies Raised in the Immunized Mice
3.3. Longevity and Memory Antibody Responses in the Mice Immunized with Chimeric DC-Targeting mAbs
3.4. Safety of αDEC-NS1 and αDCIR2-NS1 Immunization via the i.d. and i.p. Delivery Routes
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Tesfaye, D.Y.; Gudjonsson, A.; Bogen, B.; Fossum, E. Targeting Conventional Dendritic Cells to Fine-Tune Antibody Responses. Front. Immunol. 2019, 10. [Google Scholar] [CrossRef]
- Reuter, A.; Panozza, S.E.; Macri, C.; Dumont, C.; Li, J.; Liu, H.; Segura, E.; Vega-Ramos, J.; Gupta, N.; Caminschi, I.; et al. Criteria for Dendritic Cell Receptor Selection for Efficient Antibody-Targeted Vaccination. J. Immunol. 2015, 194, 2696–2705. [Google Scholar] [CrossRef]
- Lehmann, C.; Heger, L.; Heidkamp, G.; Baranska, A.; Lühr, J.; Hoffmann, A.; Dudziak, D. Direct Delivery of Antigens to Dendritic Cells via Antibodies Specific for Endocytic Receptors as a Promising Strategy for Future Therapies. Vaccines 2016, 4, 8. [Google Scholar] [CrossRef] [PubMed]
- Jiang, W.; Swiggard, W.J.; Heufler, C.; Peng, M.; Mirza, A.; Steinman, R.M.; Nussenzweig, M.C. The receptor DEC-205 expressed by dendritic cells and thymic epithelial cells is involved in antigen processing. Nature 1995, 375, 151–155. [Google Scholar] [CrossRef] [PubMed]
- Mahnke, K.; Guo, M.; Lee, S.; Sepulveda, H.; Swain, S.L.; Nussenzweig, M.; Steinman, R.M. The Dendritic Cell Receptor for Endocytosis, Dec-205, Can Recycle and Enhance Antigen Presentation via Major Histocompatibility Complex Class II–Positive Lysosomal Compartments. J. Cell Biol. 2000, 151, 673–684. [Google Scholar] [CrossRef] [PubMed]
- Dudziak, D.; Kamphorst, A.O.; Heidkamp, G.F.; Buchholz, V.R.; Trumpfheller, C.; Yamazaki, S.; Cheong, C.; Liu, K.; Lee, H.-W.; Park, C.G.; et al. Differential antigen processing by dendritic cell subsets in vivo. Science 2007, 315, 107–111. [Google Scholar] [CrossRef]
- Boscardin, S.B.; Hafalla, J.C.R.; Masilamani, R.F.; Kamphorst, A.O.; Zebroski, H.A.; Rai, U.; Morrot, A.; Zavala, F.; Steinman, R.M.; Nussenzweig, R.S.; et al. Antigen targeting to dendritic cells elicits long-lived T cell help for antibody responses. J. Exp. Med. Artic. JEM 2006, 00, 599–606. [Google Scholar] [CrossRef]
- Antonialli, R.; Sulczewski, F.B.; da Silva Amorim, K.N.; da Silva Almeida, B.; Ferreira, N.S.; Yamamoto, M.M.; Soares, I.S.; de Souza Ferreira, L.C.; Rosa, D.S.; Boscardin, S.B. CpG Oligodeoxinucleotides and Flagellin Modulate the Immune Response to Antigens Targeted to CD8α+ and CD8α− Conventional Dendritic Cell Subsets. Front. Immunol. 2017, 8. [Google Scholar] [CrossRef]
- Soares, H.; Waechter, H.; Glaichenhaus, N.; Mougneau, E.; Yagita, H.; Mizenina, O.; Dudziak, D.; Nussenzweig, M.C.; Steinman, R.M. A subset of dendritic cells induces CD4+ T cells to produce IFN-γ by an IL-12–independent but CD70-dependent mechanism in vivo. J. Exp. Med. 2007, 204, 1095–1106. [Google Scholar] [CrossRef]
- Trumpfheller, C.; Finke, J.S.; López, C.B.; Moran, T.M.; Moltedo, B.; Soares, H.; Huang, Y.; Schlesinger, S.J.; Park, C.G.; Nussenzweig, M.C.; et al. Intensified and protective CD4+ T cell immunity in mice with anti–dendritic cell HIV gag fusion antibody vaccine. J. Exp. Med. 2006, 203, 607–617. [Google Scholar] [CrossRef]
- Wang, B.; Zaidi, N.; He, L.-Z.; Zhang, L.; Kuroiwa, J.M.; Keler, T.; Steinman, R.M. Targeting of the non-mutated tumor antigen HER2/neu to mature dendritic cells induces an integrated immune response that protects against breast cancer in mice. Breast Cancer Res. 2012, 14, R39. [Google Scholar] [CrossRef] [PubMed]
- Gurer, C.; Strowig, T.; Brilot, F.; Pack, M.; Trumpfheller, C.; Arrey, F.; Park, C.G.; Steinman, R.M.; Münz, C. Targeting the nuclear antigen 1 of Epstein-Barr virus to the human endocytic receptor DEC-205 stimulates protective T-cell responses. Blood 2008, 112, 1231–1239. [Google Scholar] [CrossRef] [PubMed]
- Do, Y.; Koh, H.; Park, C.G.; Dudziak, D.; Seo, P.; Mehandru, S.; Choi, J.H.; Cheong, C.; Park, S.; Perlin, D.S.; et al. Targeting of LcrV virulence protein from Yersinia pestis to dendritic cells protects mice against pneumonic plague. Eur. J. Immunol. 2010, 40, 2791–2796. [Google Scholar] [CrossRef]
- Flynn, B.J.; Kastenmuller, K.; Wille-Reece, U.; Tomaras, G.D.; Alam, M.; Lindsay, R.W.; Salazar, A.M.; Perdiguero, B.; Gomez, C.E.; Wagner, R.; et al. Immunization with HIV Gag targeted to dendritic cells followed by recombinant New York vaccinia virus induces robust T-cell immunity in nonhuman primates. Proc. Natl. Acad. Sci. USA 2011, 108, 7131–7136. [Google Scholar] [CrossRef]
- Coconi-Linares, N.; Ortega-Dávila, E.; López-González, M.; García-Machorro, J.; García-Cordero, J.; Steinman, R.M.; Cedillo-Barrón, L.; Gómez-Lim, M.A. Targeting of envelope domain III protein of DENV type 2 to DEC-205 receptor elicits neutralizing antibodies in mice. Vaccine 2013, 31, 2366–2371. [Google Scholar] [CrossRef] [PubMed]
- Ngu, L.N.; Nji, N.N.; Ambada, G.E.; Sagnia, B.; Sake, C.N.; Tchadji, J.C.; Njambe Priso, G.D.; Lissom, A.; Tchouangueu, T.F.; Manga Tebit, D.; et al. In vivo targeting of protein antigens to dendritic cells using anti-DEC-205 single chain antibody improves HIV Gag specific CD4 + T cell responses protecting from airway challenge with recombinant vaccinia-gag virus. Immunity Inflamm. Dis. 2019, 7, 55–67. [Google Scholar] [CrossRef] [PubMed]
- Lakhrif, Z.; Moreau, A.; Hérault, B.; Di-Tommaso, A.; Juste, M.; Moiré, N.; Dimier-Poisson, I.; Mévélec, M.-N.; Aubrey, N. Targeted Delivery of Toxoplasma gondii Antigens to Dendritic Cells Promote Immunogenicity and Protective Efficiency against Toxoplasmosis. Front. Immunol. 2018, 9. [Google Scholar] [CrossRef] [PubMed]
- Dhodapkar, M.V.; Sznol, M.; Zhao, B.; Wang, D.; Carvajal, R.D.; Keohan, M.L.; Chuang, E.; Sanborn, R.E.; Lutzky, J.; Powderly, J.; et al. Induction of Antigen-Specific Immunity with a Vaccine Targeting NY-ESO-1 to the Dendritic Cell Receptor DEC-205. Sci. Transl. Med. 2014, 6, 232ra51. [Google Scholar] [CrossRef]
- Zhou, S.; Ren, T.; Gu, H.; Wang, C.; Li, M.; Zhao, Z.; Xing, L.; Zhang, L.; Sun, Y.; Yang, P.; et al. Intradermal delivery of a fractional dose of influenza H7N9 split vaccine elicits protective immunity in mice and rats. Hum. Vaccin. Immunother. 2018, 14, 623–629. [Google Scholar] [CrossRef]
- Counoupas, C.; Pinto, R.; Nagalingam, G.; Britton, W.J.; Triccas, J.A. Protective efficacy of recombinant BCG over-expressing protective, stage-specific antigens of Mycobacterium tuberculosis. Vaccine 2018, 36, 2619–2629. [Google Scholar] [CrossRef]
- Jackson, L.A.; Rupp, R.; Papadimitriou, A.; Wallace, D.; Raanan, M.; Moss, K.J. A phase 1 study of safety and immunogenicity following intradermal administration of a tetravalent dengue vaccine candidate. Vaccine 2018, 36, 3976–3983. [Google Scholar] [CrossRef]
- Hegazy-Hassan, W.; Zepeda-Escobar, J.A.; Ochoa-García, L.; Contreras-Ortíz, J.M.E.; Tenorio-Borroto, E.; Barbabosa-Pliego, A.; Aparicio-Burgos, J.E.; Oros-Pantoja, R.; Rivas-Santiago, B.; Díaz-Albiter, H.; et al. TcVac1 vaccine delivery by intradermal electroporation enhances vaccine induced immune protection against Trypanosoma cruzi infection in mice. Vaccine 2019, 37, 248–257. [Google Scholar] [CrossRef] [PubMed]
- Osorio, J.E.; Velez, I.D.; Thomson, C.; Lopez, L.; Jimenez, A.; Haller, A.A.; Silengo, S.; Scott, J.; Boroughs, K.L.; Stovall, J.L.; et al. Safety and immunogenicity of a recombinant live attenuated tetravalent dengue vaccine (DENVax) in flavivirus-naive healthy adults in Colombia: A randomised, placebo-controlled, phase 1 study. Lancet Infect. Dis. 2014, 14, 830–838. [Google Scholar] [CrossRef]
- Taweechaisupapong, S.; Sriurairatana, S.; Angsubhakorn, S.; Yoksan, S.; Khin, M.M.; Sahaphong, S.; Bhamarapravati, N. Langerhans cell density and serological changes following intradermal immunisation of mice with dengue 2 virus. J. Med. Microbiol. 1996, 45, 138–145. [Google Scholar] [CrossRef]
- Turvey, M.E.; Uppu, D.S.S.M.; Mohamed Sharif, A.R.; Bidet, K.; Alonso, S.; Ooi, E.E.; Hammond, P.T. Microneedle-based intradermal delivery of stabilized dengue virus. Bioeng. Transl. Med. 2019, 4. [Google Scholar] [CrossRef] [PubMed]
- Poggianella, M.; Slon Campos, J.L.; Chan, K.R.; Tan, H.C.; Bestagno, M.; Ooi, E.E.; Burrone, O.R. Dengue E Protein Domain III-Based DNA Immunisation Induces Strong Antibody Responses to All Four Viral Serotypes. PLoS Negl. Trop. Dis. 2015, 9, e0003947. [Google Scholar] [CrossRef]
- Williams, M.; Ewing, D.; Blevins, M.; Sun, P.; Sundaram, A.K.; Raviprakash, K.S.; Porter, K.R.; Sanders, J.W. Enhanced immunogenicity and protective efficacy of a tetravalent dengue DNA vaccine using electroporation and intradermal delivery. Vaccine 2019, 37, 4444–4453. [Google Scholar] [CrossRef]
- Andreata-Santos, R.; Dos Santos Alves, R.P.; Pereira, S.A.; Pereira, L.R.; de Freitas, C.L.; Pereira, S.S.; Venceslau-Carvalho, A.A.; Castro-Amarante, M.F.; Favaro, M.T.P.; Mathias-Santos, C.; et al. Transcutaneous Administration of Dengue Vaccines. Viruses 2020, 12, 514. [Google Scholar] [CrossRef]
- Bragazzi, N.L.; Orsi, A.; Ansaldi, F.; Gasparini, R.; Icardi, G. Fluzone® intra-dermal (Intanza®/Istivac® Intra-dermal): An updated overview. Hum. Vaccin. Immunother. 2016, 12, 2616–2627. [Google Scholar] [CrossRef]
- Stoitzner, P.; Schaffenrath, S.; Tripp, C.H.; Reider, D.; Komenda, K.; Del Frari, B.; Djedovic, G.; Ebner, S.; Romani, N. Human skin dendritic cells can be targeted in situ by intradermal injection of antibodies against lectin receptors. Exp. Dermatol. 2014, 23, 909–915. [Google Scholar] [CrossRef]
- Ebner, S.; Ehammer, Z.; Holzmann, S.; Schwingshackl, P.; Forstner, M.; Stoitzner, P.; Huemer, G.M.; Fritsch, P.; Romani, N. Expression of C-type lectin receptors by subsets of dendritic cells in human skin. Int. Immunol. 2004, 16, 877–887. [Google Scholar] [CrossRef] [PubMed]
- Flacher, V.; Tripp, C.H.; Haid, B.; Kissenpfennig, A.; Malissen, B.; Stoitzner, P.; Idoyaga, J.; Romani, N. Skin Langerin+ Dendritic Cells Transport Intradermally Injected Anti-DEC-205 Antibodies but Are Not Essential for Subsequent Cytotoxic CD8+ T Cell Responses. J. Immunol. 2012, 188, 2146–2155. [Google Scholar] [CrossRef] [PubMed]
- Combadiere, B.; Liard, C. Transcutaneous and intradermal vaccination. Hum. Vaccin. 2011, 7, 811–827. [Google Scholar] [CrossRef] [PubMed]
- Lenz, A.; Heine, M.; Schuler, G.; Romani, N. Human and murine dermis contain dendritic cells: Isolation by means of a novel method and phenotypical and functional characterization. J. Clin. Investig. 1993, 92, 2587–2596. [Google Scholar] [CrossRef] [PubMed]
- Meunier, L.; Gonzalez-Ramos, A.; Cooper, K.D. Heterogeneous populations of class II MHC+ cells in human dermal cell suspensions. Identification of a small subset responsible for potent dermal antigen-presenting cell activity with features analogous to Langerhans cells. J. Immunol. 1993, 151, 4067–4080. [Google Scholar]
- Halstead, S.B. In Vivo Enhancement of Dengue Virus Infection in Rhesus Monkeys by Passively Transferred Antibody. J. Infect. Dis. 1979, 140, 527–533. [Google Scholar] [CrossRef]
- Flipse, J.; Smit, J.M. The Complexity of a Dengue Vaccine: A Review of the Human Antibody Response. PLoS Negl. Trop. Dis. 2015, 9, e0003749. [Google Scholar] [CrossRef]
- Balsitis, S.J.; Williams, K.L.; Lachica, R.; Flores, D.; Kyle, J.L.; Mehlhop, E.; Johnson, S.; Diamond, M.S.; Beatty, P.R.; Harris, E. Lethal antibody enhancement of dengue disease in mice is prevented by Fc modification. PLoS Pathog. 2010, 6. [Google Scholar] [CrossRef]
- Amorim, J.H.; Diniz, M.O.; Cariri, F.A.M.O.; Rodrigues, J.F.; Bizerra, R.S.P.; Gonçalves, A.J.S.; De Barcelos Alves, A.M.; De Souza Ferreira, L.C. Protective immunity to DENV2 after immunization with a recombinant NS1 protein using a genetically detoxified heat-labile toxin as an adjuvant. Vaccine 2012, 30, 837–845. [Google Scholar] [CrossRef]
- Costa, S.M.; Azevedo, A.S.; Paes, M.V.; Sarges, F.S.; Freire, M.S.; Alves, A.M.B. DNA vaccines against dengue virus based on the ns1 gene: The influence of different signal sequences on the protein expression and its correlation to the immune response elicited in mice. Virology 2007, 358, 413–423. [Google Scholar] [CrossRef]
- Costa, S.M.; Freire, M.S.; Alves, A.M.B. DNA vaccine against the non-structural 1 protein (NS1) of dengue 2 virus. Vaccine 2006, 24, 4562–4564. [Google Scholar] [CrossRef] [PubMed]
- Amorim, J.H.; Alves, R.P.D.S.; Boscardin, S.B.; Ferreira, L.C.D.S. The dengue virus non-structural 1 protein: Risks and benefits. Virus Res. 2014, 181, 53–60. [Google Scholar] [CrossRef] [PubMed]
- Muller, D.A.; Young, P.R. The flavivirus NS1 protein: Molecular and structural biology, immunology, role in pathogenesis and application as a diagnostic biomarker. Antivir. Res. 2013, 98, 192–208. [Google Scholar] [CrossRef] [PubMed]
- Edeling, M.A.; Diamond, M.S.; Fremont, D.H. Structural basis of Flavivirus NS1 assembly and antibody recognition. Proc. Natl. Acad. Sci. USA 2014, 111, 4285–4290. [Google Scholar] [CrossRef]
- Gutsche, I.; Coulibaly, F.; Voss, J.E.; Salmon, J.; d’Alayer, J.; Ermonval, M.; Larquet, E.; Charneau, P.; Krey, T.; Mégret, F.; et al. Secreted dengue virus nonstructural protein NS1 is an atypical barrel-shaped high-density lipoprotein. Proc. Natl. Acad. Sci. USA 2011, 108, 8003–8008. [Google Scholar] [CrossRef]
- Gao, G.; Wang, Q.; Dai, Z.; Calcedo, R.; Sun, X.; Li, G.; Wilson, J.M. Adenovirus-based vaccines generate cytotoxic T lymphocytes to epitopes of NS1 from dengue virus that are present in all major serotypes. Hum. Gene Ther. 2008, 19, 927–936. [Google Scholar] [CrossRef]
- Henriques, H.R.; Rampazo, E.V.; Gonçalves, A.J.S.; Vicentin, E.C.M.; Amorim, J.H.; Panatieri, R.H.; Amorim, K.N.S.; Yamamoto, M.M.; Ferreira, L.C.S.; Alves, A.M.B.; et al. Targeting the non-structural protein 1 from dengue virus to a dendritic cell population confers protective immunity to lethal virus challenge. PLoS Negl. Trop. Dis. 2013, 7, e2330. [Google Scholar] [CrossRef]
- Amorim, J.H.; Porchia, B.F.M.M.; Balan, A.; Cavalcante, R.C.M.; da Costa, S.M.; de Barcelos Alves, A.M.; de Souza Ferreira, L.C. Refolded dengue virus type 2 NS1 protein expressed in Escherichia coli preserves structural and immunological properties of the native protein. J. Virol. Methods 2010, 167, 186–192. [Google Scholar] [CrossRef]
- Maeda, D.L.N.F.; Batista, M.T.; Pereira, L.R.; de Jesus Cintra, M.; Amorim, J.H.; Mathias-Santos, C.; Pereira, S.A.; Boscardin, S.B.; dos Ramos Silva, S.; Faquim-Mauro, E.L.; et al. Adjuvant-Mediated Epitope Specificity and Enhanced Neutralizing Activity of Antibodies Targeting Dengue Virus Envelope Protein. Front. Immunol. 2017, 8. [Google Scholar] [CrossRef]
- Rocha, L.; Alves, R.; Caetano, B.; Pereira, L.; Mitsunari, T.; Amorim, J.; Polatto, J.; Botosso, V.; Gallina, N.; Palacios, R.; et al. Epitope Sequences in Dengue Virus NS1 Protein Identified by Monoclonal Antibodies. Antibodies 2017, 6, 14. [Google Scholar] [CrossRef]
- Tanjoni, I.; Evangelista, K.; Della-Casa, M.S.; Butera, D.; Magalhães, G.S.; Baldo, C.; Clissa, P.B.; Fernandes, I.; Eble, J.; Moura-da-Silva, A.M. Different regions of the class P-III snake venom metalloproteinase jararhagin are involved in binding to α2β1 integrin and collagen. Toxicon 2010, 55, 1093–1099. [Google Scholar] [CrossRef] [PubMed]
- Cheong, C.; Choi, J.H.; Vitale, L.; He, L.Z.; Trumpfheller, C.; Bozzacco, L.; Do, Y.; Nchinda, G.; Park, S.H.; Dandamudi, D.B.; et al. Improved cellular and humoral immune responses in vivo following targeting of HIV Gag to dendritic cells within human anti-human DEC205 monoclonal antibody. Blood 2010, 116, 3828–3838. [Google Scholar] [CrossRef] [PubMed]
- Beatty, P.R.; Puerta-Guardo, H.; Killingbeck, S.S.; Glasner, D.R.; Hopkins, K.; Harris, E. Dengue virus NS1 triggers endothelial permeability and vascular leak that is prevented by NS1 vaccination. Sci. Transl. Med. 2015, 7, 304ra141. [Google Scholar] [CrossRef] [PubMed]
- Niu, L.; Chu, L.Y.; Burton, S.A.; Hansen, K.J.; Panyam, J. Intradermal delivery of vaccine nanoparticles using hollow microneedle array generates enhanced and balanced immune response. J. Control. Release 2019, 294, 268–278. [Google Scholar] [CrossRef]
- Yasuda, T.; Ura, T.; Taniguchi, M.; Yoshida, H. Intradermal Delivery of Antigens Enhances Specific IgG and Diminishes IgE Production: Potential Use for Vaccination and Allergy Immunotherapy. PLoS ONE 2016, 11, e0167952. [Google Scholar] [CrossRef]
- Collins, A.M. IgG subclass co-expression brings harmony to the quartet model of murine IgG function. Immunol. Cell Biol. 2016, 94, 949–954. [Google Scholar] [CrossRef]
- Coutelier, J.P.; van der Logt, J.T.; Heessen, F.W.; Vink, A.; van Snick, J. Virally induced modulation of murine IgG antibody subclasses. J. Exp. Med. 1988, 168, 2373–2378. [Google Scholar] [CrossRef]
- Germann, T.; Bongartz, M.; Dlugonska, H.; Hess, H.; Schmitt, E.; Kolbe, L.; Kölsch, E.; Podlaski, F.J.; Gately, M.K.; Rüde, E. Interleukin-12 profoundly up-regulates the synthesis of antigen-specific complement-fixing IgG2a, IgG2b and IgG3 antibody subclassesin vivo. Eur. J. Immunol. 1995, 25, 823–829. [Google Scholar] [CrossRef]
- Nimmerjahn, F.; Lux, A.; Albert, H.; Woigk, M.; Lehmann, C.; Dudziak, D.; Smith, P.; Ravetch, J.V. Fc RIV deletion reveals its central role for IgG2a and IgG2b activity in vivo. Proc. Natl. Acad. Sci. USA 2010, 107, 19396–19401. [Google Scholar] [CrossRef]
- Bailey, M.J.; Broecker, F.; Duehr, J.; Arumemi, F.; Krammer, F.; Palese, P.; Tan, G.S. Antibodies Elicited by an NS1-Based Vaccine Protect Mice against Zika Virus. MBio 2019, 10. [Google Scholar] [CrossRef]
- Le Moignic, A.; Malard, V.; Benvegnu, T.; Lemiègre, L.; Berchel, M.; Jaffrès, P.-A.; Baillou, C.; Delost, M.; Macedo, R.; Rochefort, J.; et al. Preclinical evaluation of mRNA trimannosylated lipopolyplexes as therapeutic cancer vaccines targeting dendritic cells. J. Control. Release 2018, 278, 110–121. [Google Scholar] [CrossRef]
- Zaneti, A.B.; Yamamoto, M.M.; Sulczewski, F.B.; da Silva Almeida, B.; Souza, H.F.S.; Ferreira, N.S.; Maeda, D.L.N.F.; Sales, N.S.; Rosa, D.S.; de Souza Ferreira, L.C.; et al. Dendritic Cell Targeting Using a DNA Vaccine Induces Specific Antibodies and CD4+ T Cells to the Dengue Virus Envelope Protein Domain III. Front. Immunol. 2019, 10. [Google Scholar] [CrossRef]
- Shin, C.; Han, J.-A.; Koh, H.; Choi, B.; Cho, Y.; Jeong, H.; Ra, J.-S.; Sung, P.S.; Shin, E.-C.; Ryu, S.; et al. CD8α− Dendritic Cells Induce Antigen-Specific T Follicular Helper Cells Generating Efficient Humoral Immune Responses. Cell Rep. 2015, 11, 1929–1940. [Google Scholar] [CrossRef]
- Sulczewski, F.B.; Martino, L.A.; da Silva Almeida, B.; Zaneti, A.B.; Ferreira, N.S.; da Silva Amorim, K.N.; Yamamoto, M.M.; de Souza Apostolico, J.; Rosa, D.S.; Boscardin, S.B. Conventional type 1 dendritic cells induce T H 1, T H 1-like follicular helper T cells and regulatory T cells after antigen boost via DEC205 receptor. Eur. J. Immunol. 2020, eji.202048694. [Google Scholar] [CrossRef] [PubMed]
- Bailey, M.J.; Duehr, J.; Dulin, H.; Broecker, F.; Brown, J.A.; Arumemi, F.O.; Bermúdez González, M.C.; Leyva-Grado, V.H.; Evans, M.J.; Simon, V.; et al. Human antibodies targeting Zika virus NS1 provide protection against disease in a mouse model. Nat. Commun. 2018, 9, 4560. [Google Scholar] [CrossRef] [PubMed]
- Lai, Y.-C.; Chuang, Y.-C.; Liu, C.-C.; Ho, T.-S.; Lin, Y.-S.; Anderson, R.; Yeh, T.-M. Antibodies Against Modified NS1 Wing Domain Peptide Protect Against Dengue Virus Infection. Sci. Rep. 2017, 7, 6975. [Google Scholar] [CrossRef] [PubMed]
- Wan, S.W.; Lu, Y.T.; Huang, C.H.; Lin, C.F.; Anderson, R.; Liu, H.S.; Yeh, T.M.; Yen, Y.T.; Wu-Hsieh, B.A.; Lin, Y.S. Protection against dengue virus infection in mice by administration of antibodies against modified nonstructural protein 1. PLoS ONE 2014, 9, e0092495. [Google Scholar] [CrossRef] [PubMed]
- Modhiran, N.; Watterson, D.; Muller, D.A.; Panetta, A.K.; Sester, D.P.; Liu, L.; Hume, D.A.; Stacey, K.J.; Young, P.R. Dengue virus NS1 protein activates cells via Toll-like receptor 4 and disrupts endothelial cell monolayer integrity. Sci. Transl. Med. 2015, 7, 304ra142. [Google Scholar] [CrossRef]
- Avirutnan, P.; Hauhart, R.E.; Somnuke, P.; Blom, A.M.; Diamond, M.S.; Atkinson, J.P. Binding of flavivirus nonstructural protein NS1 to C4b binding protein modulates complement activation. J. Immunol. 2011, 187, 424–433. [Google Scholar] [CrossRef] [PubMed]
- Lin, S.-W.; Chuang, Y.-C.; Lin, Y.-S.; Lei, H.-Y.; Liu, H.-S.; Yeh, T.-M. Dengue virus nonstructural protein NS1 binds to prothrombin/thrombin and inhibits prothrombin activation. J. Infect. 2012, 64, 325–334. [Google Scholar] [CrossRef] [PubMed]
- Lin, C.-F.; Wan, S.-W.; Chen, M.-C.; Lin, S.-C.; Cheng, C.-C.; Chiu, S.-C.; Hsiao, Y.-L.; Lei, H.-Y.; Liu, H.-S.; Yeh, T.-M.; et al. Liver injury caused by antibodies against dengue virus nonstructural protein 1 in a murine model. Lab. Investig. 2008, 88, 1079–1089. [Google Scholar] [CrossRef] [PubMed]
- Wan, S.-W.; Lin, C.-F.; Chen, M.-C.; Lei, H.-Y.; Liu, H.-S.; Yeh, T.-M.; Liu, C.-C.; Lin, Y.-S. C-Terminal Region of Dengue Virus Nonstructural Protein 1 Is Involved in Endothelial Cell Cross-Reactivity via Molecular Mimicry. Am. J. Infect. Dis. 2008, 4, 85–91. [Google Scholar] [CrossRef]
- Lin, C.-F.; Chiu, S.; Hsiao, Y.-L.; Wan, S.; Lei, H.; Shiau, A.-L.; Liu, H.-S.; Yeh, T.; Chen, S.-H.; Liu, C.-C.; et al. Expression of cytokine, chemokine, and adhesion molecules during endothelial cell activation induced by antibodies against dengue virus nonstructural protein 1. J. Immunol. 2005, 174, 395–403. [Google Scholar] [CrossRef] [PubMed]
- Lin, C.-F.; Lei, H.-Y.; Shiau, a.-L.; Liu, H.-S.; Yeh, T.-M.; Chen, S.-H.; Liu, C.-C.; Chiu, S.-C.; Lin, Y.-S. Endothelial cell apoptosis induced by antibodies against dengue virus nonstructural protein 1 via production of nitric oxide. J. Immunol. 2002, 169, 657–664. [Google Scholar] [CrossRef] [PubMed]
- Libraty, D.H.; Young, P.R.; Pickering, D.; Endy, T.P.; Kalayanarooj, S.; Green, S.; Vaughn, D.W.; Nisalak, A.; Ennis, F.A.; Rothman, A.L. High Circulating Levels of the Dengue Virus Nonstructural Protein NS1 Early in Dengue Illness Correlate with the Development of Dengue Hemorrhagic Fever. J. Infect. Dis. 2002, 186, 1165–1168. [Google Scholar] [CrossRef] [PubMed]
- Young, P.R.; Hilditch, P.A.; Bletchly, C.; Halloran, W. An antigen capture enzyme-linked immunosorbent assay reveals high levels of the dengue virus protein NS1 in the sera of infected patients. J. Clin. Microbiol. 2000, 38, 1053–1057. [Google Scholar] [CrossRef] [PubMed]
- Chuang, Y.-C.; Wang, S.-Y.; Lin, Y.-S.; Chen, H.-R.; Yeh, T.-M. Re-evaluation of the pathogenic roles of nonstructural protein 1 and its antibodies during dengue virus infection. J. Biomed. Sci. 2013, 20, 42. [Google Scholar] [CrossRef] [PubMed]
- Chen, M.-C.; Lin, C.-F.; Lei, H.-Y.; Lin, S.-C.; Liu, H.-S.; Yeh, T.-M.; Anderson, R.; Lin, Y.-S. Deletion of the C-Terminal Region of Dengue Virus Nonstructural Protein 1 (NS1) Abolishes Anti-NS1-Mediated Platelet Dysfunction and Bleeding Tendency. J. Immunol. 2009, 183, 1797–1803. [Google Scholar] [CrossRef] [PubMed]
- Cheng, H.J.; Lei, H.Y.; Lin, C.F.; Luo, Y.H.; Wan, S.W.; Liu, H.S.; Yeh, T.M.; Lin, Y.S. Anti-dengue virus nonstructural protein 1 antibodies recognize protein disulfide isomerase on platelets and inhibit platelet aggregation. Mol. Immunol. 2009, 47, 398–406. [Google Scholar] [CrossRef] [PubMed]




© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pereira, L.R.; Vicentin, E.C.M.; Pereira, S.A.; Maeda, D.L.N.F.; Alves, R.P.d.S.; Andreata-Santos, R.; Sousa, F.T.G.d.; Yamamoto, M.M.; Castro-Amarante, M.F.; Favaro, M.T.d.P.; et al. Intradermal Delivery of Dendritic Cell-Targeting Chimeric mAbs Genetically Fused to Type 2 Dengue Virus Nonstructural Protein 1. Vaccines 2020, 8, 565. https://doi.org/10.3390/vaccines8040565
Pereira LR, Vicentin ECM, Pereira SA, Maeda DLNF, Alves RPdS, Andreata-Santos R, Sousa FTGd, Yamamoto MM, Castro-Amarante MF, Favaro MTdP, et al. Intradermal Delivery of Dendritic Cell-Targeting Chimeric mAbs Genetically Fused to Type 2 Dengue Virus Nonstructural Protein 1. Vaccines. 2020; 8(4):565. https://doi.org/10.3390/vaccines8040565
Chicago/Turabian StylePereira, Lennon Ramos, Elaine Cristina Matos Vicentin, Sara Araujo Pereira, Denicar Lina Nascimento Fabris Maeda, Rúbens Prince dos Santos Alves, Robert Andreata-Santos, Francielle Tramontini Gomes de Sousa, Marcio Massao Yamamoto, Maria Fernanda Castro-Amarante, Marianna Teixeira de Pinho Favaro, and et al. 2020. "Intradermal Delivery of Dendritic Cell-Targeting Chimeric mAbs Genetically Fused to Type 2 Dengue Virus Nonstructural Protein 1" Vaccines 8, no. 4: 565. https://doi.org/10.3390/vaccines8040565
APA StylePereira, L. R., Vicentin, E. C. M., Pereira, S. A., Maeda, D. L. N. F., Alves, R. P. d. S., Andreata-Santos, R., Sousa, F. T. G. d., Yamamoto, M. M., Castro-Amarante, M. F., Favaro, M. T. d. P., Romano, C. M., Sabino, E. C., Boscardin, S. B., & Ferreira, L. C. d. S. (2020). Intradermal Delivery of Dendritic Cell-Targeting Chimeric mAbs Genetically Fused to Type 2 Dengue Virus Nonstructural Protein 1. Vaccines, 8(4), 565. https://doi.org/10.3390/vaccines8040565