1. Introduction
Drug resistance of animal pathogens is a continuously growing problem, and immunosuppressive pathogens worsen mixed infections to animals, which is still a massive challenge for the prevention and control of animal infectious diseases. Moreover, antibiotics have been banned as growth promoters in 86 countries and regions around the world since 2018, and all varieties of growth-promoting antimicrobial additives will be prohibited by the end of 2020 in China. Therefore, there is a pressing need for the replacement of antimicrobial additives to prevent complicated infection of animals, as well as to improve their innate and acquired immunity, particularly to strengthen the animal mucosal immune defense responses and antibacterial ability. In order to decrease the occurrence of drug resistance, more effective roles of T helper cells in optimizing antibody immune response and effector CD4
+ or CD8
+ T cells in eliminating intracellular pathogens are greatly expected [
1].
Cytokines have been predicted to be much milder and far more efficient adjuvants [
2]. Relevant evaluations of the immune effect of cytokines as vaccine adjuvants have been conducted [
3,
4]. Interleukin-4 (IL-4) is an essential immunomodulator, which can stimulate the expression of immunoglobulins selectively. Interleukin-6 (IL-6) is one of the multiple functional cytokines with the ability to promote a variety of cell proliferation and differentiation. Both IL-4 and IL-6 are canonical type 2 cytokines, which are known to have the ability to significantly promote the proliferation of B lymphocyte cell and induce the type 2 immune response.
Antimicrobial peptides have been recognized as a vital component of the host due to their strong antibiotic activity in a wide range of microorganisms including bacteria, fungi, and enveloped viruses. Otherwise, they can also trigger a specific defense of the host [
5]. With the ability to modulate the innate immune response of the host, cathelicidin antimicrobial peptides (CAMPs) became a focus of adjuvants. Proline-arginine-rich 39-amino acid peptide (PR-39) is a proline-rich peptide, which carries out antibacterial function by terminating protein and DNA synthesis [
6]. Other biological functions such as prevention of inflammation of the tissue injury, enhancement of the wound repair ability, and regulation of angiogenesis have been reported [
7]. Tritrpticin, a member of cathelicidin family, has a broad spectrum of antibiotic activity. It acts through non-specific binding to the cytoplasmic membrane instead of binding to a specific protein receptor [
8]. Therefore, as more and more drug-resistant strains emerge, tritrpticin still shows a lower rate of inducing drug resistance. Porcine myeloid antimicrobial peptide-23 (PMAP-23) belongs to the amphipathic α-helical peptides group, which was reported to show potent antimicrobial activity against Gram-negative and Gram-positive bacteria without hemolytic activity [
9]. Protegrin-1(PG-1), a cysteine-rich β-hairpin antimicrobial peptide with 18 residues, was found out to be a potent antimicrobial agent with in vivo activity against clinically relevant, antibiotic-resistant bacteria [
10] (
Table 1).
Our previous studies demonstrated that IL-4, IL6, and fusion IL-4/6 genes can significantly enhance the immunity of mice or piglets [
11]. In order to verify the potential of fusion CAMPs and cytokines as immunomodulators, fusion gene of CAMPs and IL-4/IL-6 were constructed in this research to evaluate its effects on the immune response of mice. The chitosan was employed as the packing material due to its remarkable property of plasmid DNA delivery [
12,
13].
2. Materials and Methods
2.1. Construction of the Recombinant Eukaryotic Expression Plasmid
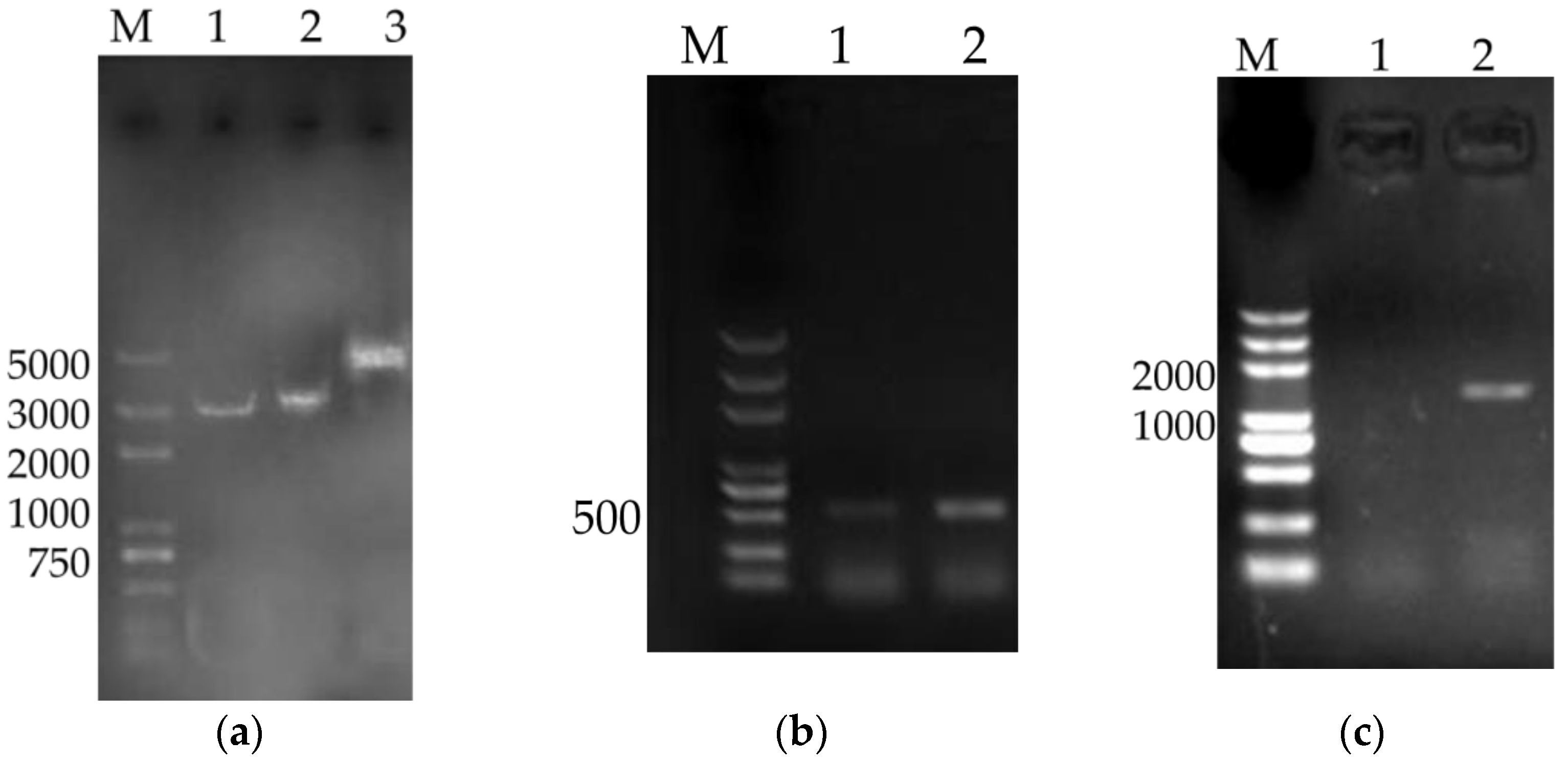
Plasmid pVAX1 (Invitrogen Company, Carlsbad, CA, USA) was used as the eukaryotic expression vector of DNA vaccine. Endo-Free Plasmid Giga Kit (OMEGA) was employed to extract plasmids among the experiment. The fusion gene of IL-4 and IL-6 (IL-4/6) was constructed previously in our lab by inserting the IL-4/6 fusion gene into the expression VR1020 plasmid (Genbank AY 294020 and AF 493992).
The recombinant gene (TPA-PR39-linker-Tritrpticin-linker-PAMP23-linker-Protegrin1-His-tag) of downstream tissue plasminogen activator (TPA) signal sequence and antimicrobial peptides fusion gene-CAMPs, which was connected by GSGDDDDK linker with each other, was amplified from pVR1020-CAMPs (VRP), which constructed in our lab previously. PCR primers for the TPA-CAMPs sequences were P1:5′-GTTTAAACTTAAGCTTATGGATGCAATGAAGAGAG-3′ and P2:5′-AAACGGGCCCTCTAGACTAATGGTGATGGTGATGATGCT-3′. Then, TPA-CAMPs were subcloned into pVAX1 to construct recombinant plasmid pVAX1-CAMPs (designated as VAP) using the In-fusion cloning technique. Furthermore, linkage of IL-4/6 and TPA-CAMPs was achieved by the use of the 63 bp 2A self-cleaving sequence (GGA AGC GGA GAG GGC AGG GGA AGT CTT CTA ACA TGC GGG GAC GTG GAG GAA AAT CCC GGG CCA) to make sure that the proteins are expressed singly. The fused fragment was subcloned into the eukaryotic expression plasmid pVAX1 (designated as VAP4/6).
2.2. Large-Scale Preparation of VAP, VAP4/6, and pVAX1
VAP, VAP4/6, and pVAX1 were transformed into
Escherichia coli (
E. coli) DH5α competent cells (Transgene Biotech, Beijing, China), respectively, and cultured in the Lurea Bertani (LB) medium with 100 µg/mL kanamycin (Kana) at 37 °C for 12 h. The positive clones were confirmed by colony PCR assay and inoculated in LB broth with kana (100 µg/mL) at 37 °C and shaken at 200 rpm for 15 h. Bacterial cells were pelleted by centrifugation and plasmid DNA were extracted by large-scale alkine lysis. Then spermine method was used to precipitate and purify extracted plasmids [
14]. Residual contamination by endogenous toxin of
E. coli was measured by Limulus amebocyte lysate test to be within 0.1 EU/mg [
15]. Subsequently, the plasmids were resuspended in sterile water and stored at −20 °C until use.
2.3. Preparation and Detection of Recombinant Plasmids Encapsulated in Chitosan Nanoparticles
Chitosan (CS) was provided by Sigma Aldrich (St. Louis, MO, USA). Its molecular weight (MW) was 150 kDa with 95% deacetylation. Plasmid VAP, VAP4/6, and pVAX1 entrapped in CS (designated as CS-VAP, CS-VAP4/6, and CS-pVAX1, respectively) were prepared by the ionotropic gelation method [
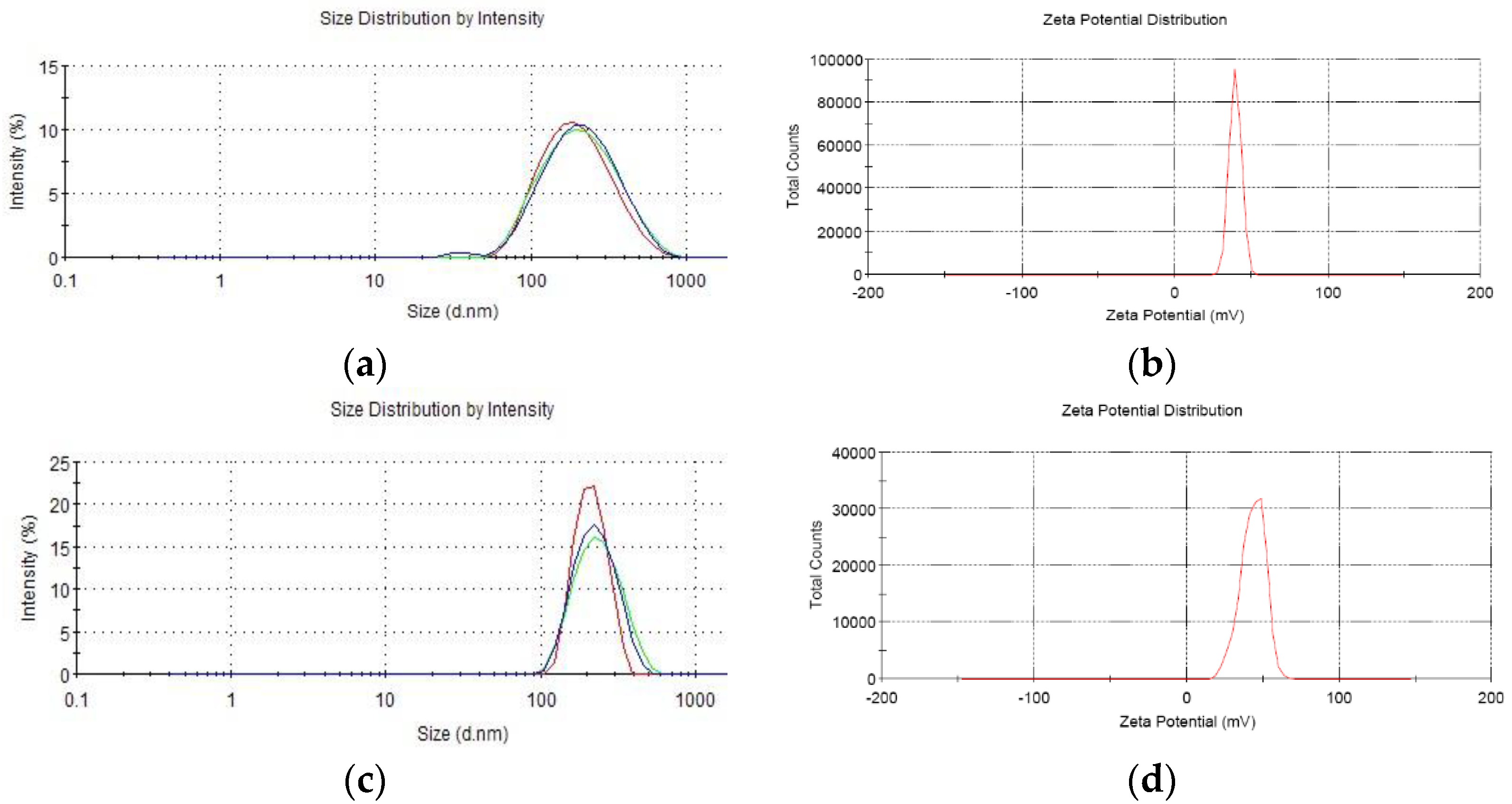
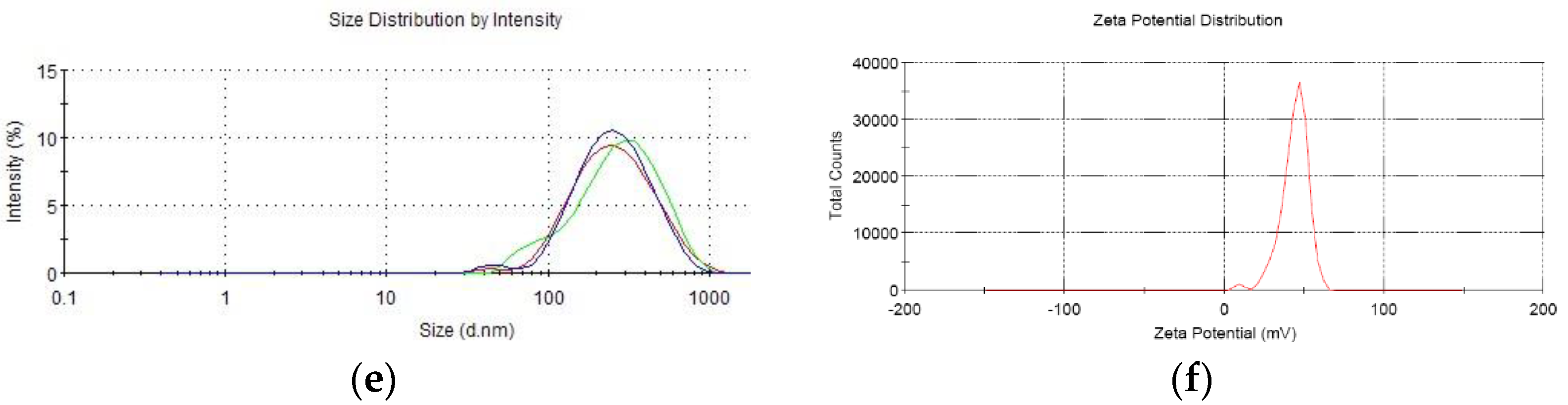
16]. Briefly, CS was dissolved in 1% glacial acetic acid buffer (pH 5.5) to a concentration of 2.4 mg/mL and filtered through 0.22 µm Millipore filter to remove bacteria. Then each recombinant plasmid was incubated in 55 °C for 20 min mixed with sodium polyphosphate solution. At last, each plasmid solution was gently dripped into CS solution until mass ratio (CS to plasmid) reached 30:1 in 50–55 °C water bath with magnetic stirring. Then, the solution was incubated for 10 min to form CS-recombinant plasmid. The granule diameter, dispersion rate, and zeta electronic potential were measured by a Zetasizer3000 HS/IHPL instrument (Malvern Instruments Ltd., Malvern, UK).
2.4. Biological Activity Assay of CS-VAP, CS-VAP4/6, and CS-pVAX1 in Pig Lymphocytes
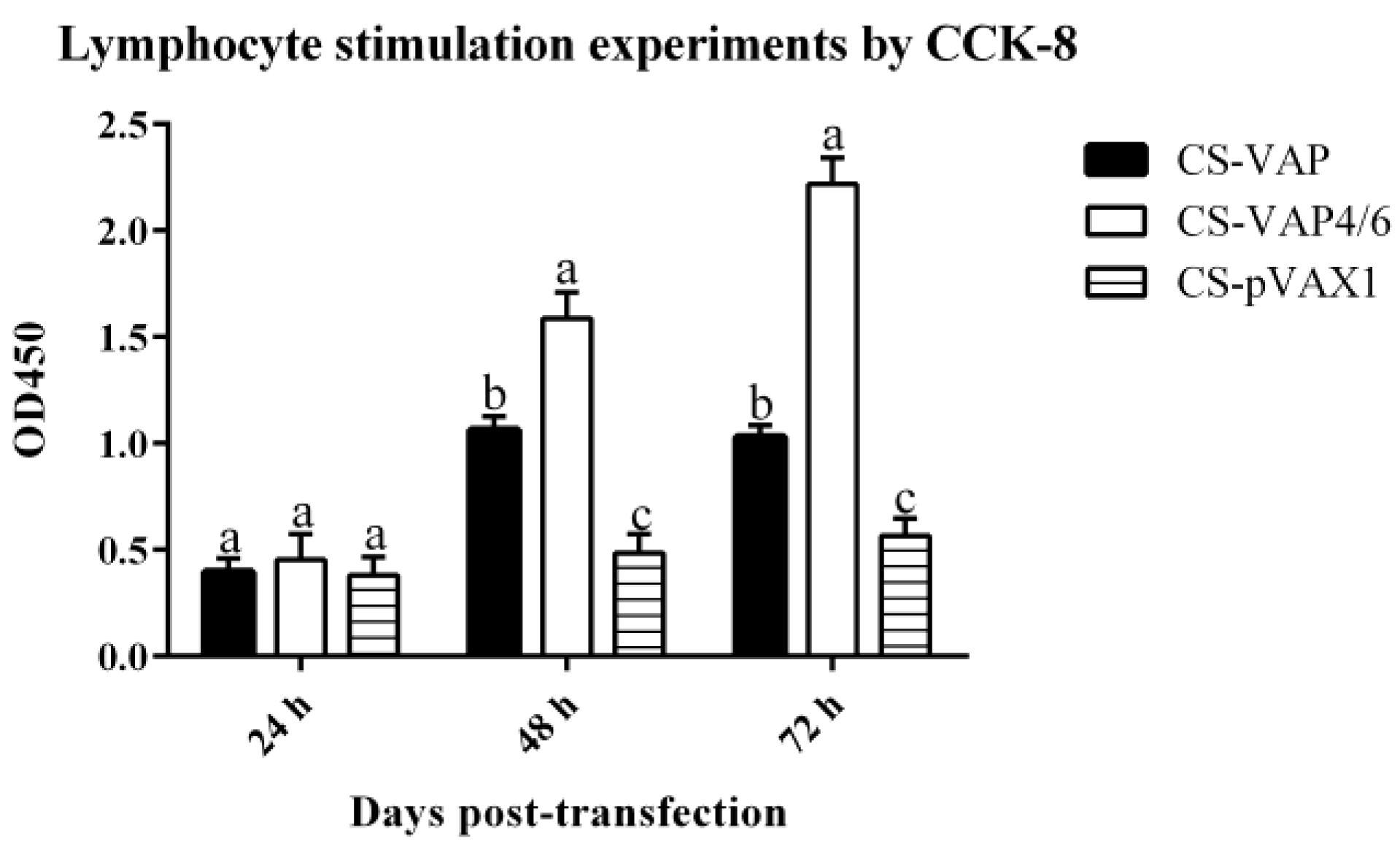
HEK 293 cells (1.0 × 105 cells/well) transfected with CS-VAP, CS-VAP4/6, and CS-pVAX1 (4 µg plasmid DNA/well), respectively, were to express CAMPs and IL-4/6 protein. Green fluorescent protein (GFP) plasmid was used as a positive control. The transfected cells were harvested at 24, 48, and 72 h, respectively. Reverse transcription polymerase chain reaction (RT-PCR) was employed to assess mRNA level. The cell supernatant from three time points was used to stimulate the lymphocyte proliferation through Counting Kit-8 (CCK8 Yiyuan biotechnology, Guangzhou, China). Pig peripheral blood mononuclear immune (PBMI) cells separated by Lymphocyte Separation Medium (LSM, ficoll 400) were diluted to 2 × 106 cells per milliliter and cultured in the dishes for 10 mL, then the pig lymphoblast was incubated for 24 h at 37 °C, and 5% CO2 stimulated with 5 µg/mL Con A (Sigma Chemical Co., St. Louis, MO, USA). The RPMI 1640 medium was used to modulate the cell concentration to 6 × 106 cells per milliliter, then 50 µL cells were incubated with 50 µL sample supernatants at 37 °C and 5% CO2. Each sample was divided into three duplicate wells, and RPMI 1640, PBS, and 100 µL pig lymphoblast cells were set as the control group. CCK8 (10 µL) was added to each well at 48 h. After incubating for another 2 h, the absorbances at 450 nm of each sample were determined, using a microplate reader 3550 (Bio-Rad, Hercules, CA, USA).
2.5. Animal Vaccination
Thirty healthy 4-week female Kunming mice (purchased from the Animal Center of West China Center of Medical Sciences, Sichuan University, Chengdu, China) were assigned to 3 groups (CS-VAP, CS-VAP4/6, and CS-pVAX1) randomly. Each group was injected intraperitoneally (IP) with 0.2 mL recombinant plasmid (0.5 mg/mL), respectively. Group CS-pVAX1 was set as a negative control. Peripheral blood samples were collected weekly from the tail vein of mice on weeks 0, 1, 2, 3, 4 after injection to evaluate the immunological changes. Finally, Staphylococcus aureus (ATCC 25923) and E. coli (ATCC 25922) stored in our laboratory were used to challenge the mice at 28 days. The bacteria were cultured in LB culture. Bacteria broth culture (0.2 mL) (109 CFU/mL) corresponding to 107 CFU/g of body weight was injected into the mice, respectively. Each bacterium was intraperitoneally injected into 5 mice. All mice were fed under the same conditions. The care and use of experimental animals complied with Chinese animal welfare laws, guidelines, and regulations (SYXK-Chuan-2018-185).
2.6. Immunological Assays In Vivo
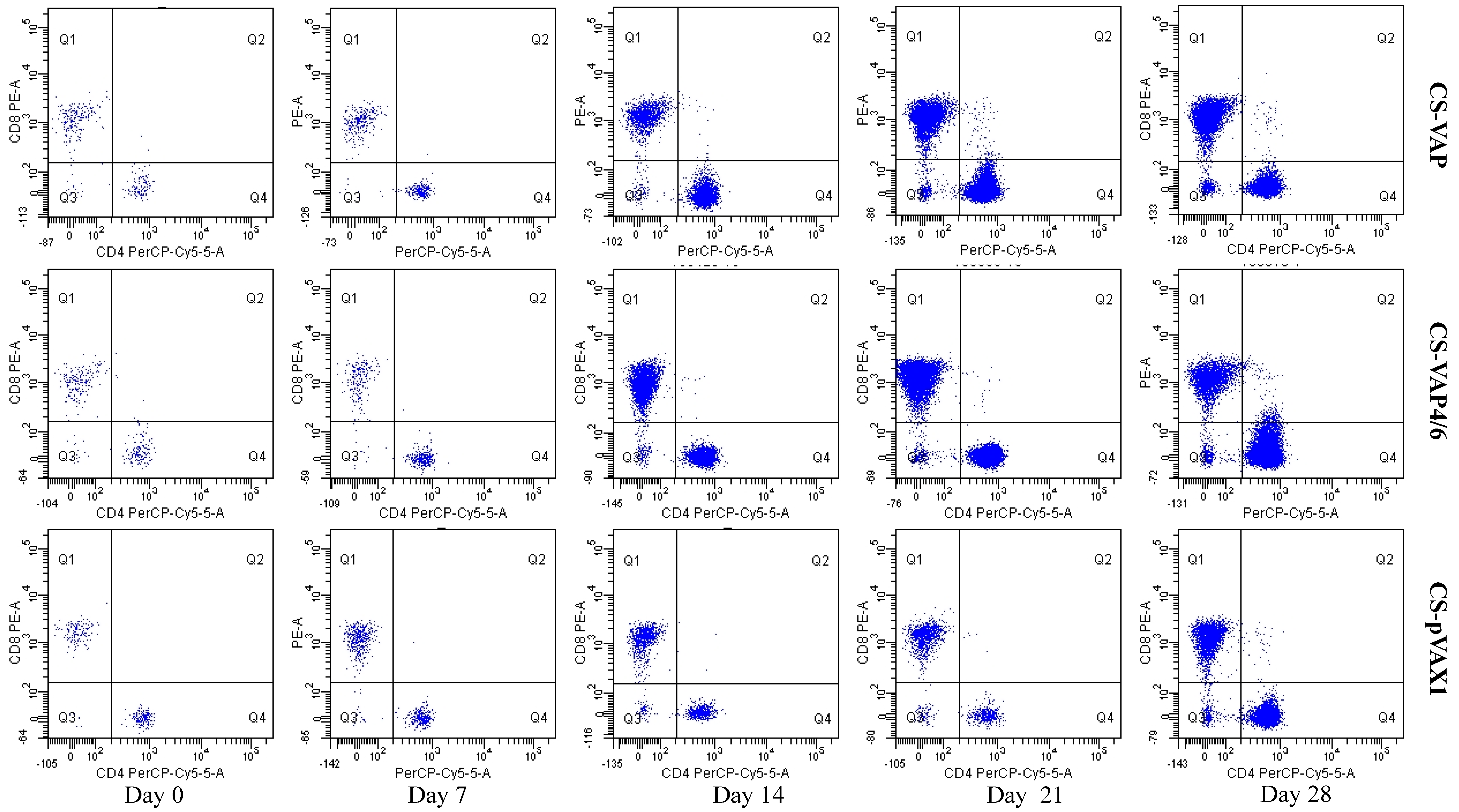
2.6.1. Assay of CD4 and CD8 Positive T Cells by Flow Cytometry
Anti-Mouse CD8a and Anti-Mouse CD4, labelled with phycoerythrin (PE) and PerCP-Cy5.5, respectively, were purchased from eBioscience (San Diego, CA, USA) Each test contained 50 µL of peripheral blood sample, and then incubated 0.25 µL PE labeled Anti-Mouse CD8a and 0.25 µL PerCP-Cy5.5 labeled Anti-Mouse CD4. After incubating for 20 min in the dark, 2 mL lysing solution (Becton Dickinson, Franklin Lakes, NJ, USA, 10%, v/v) was added to make the lysis of erythrocytes more complete and surviving cells were washed twice with PBS, with centrifugation between each step for 5 min at 500 g. FACScan flow cytometer was used for FCM analysis (BD Biosciences, San Jose, CA, USA). First, an electronic gate was set on lymphocytes and then gated CD4+ T cells in Y axis and CD8+ cells as figure. Dead cells were excluded according to the forward scatter and side scatter.
2.6.2. Assays of IgG, IgG1, IgG2 by Sandwich ELISA
Mouse Immunoglobin G (IgG), Immunoglobin G1 (IgG1), and Immunoglobin G2a (IgG2a) quantitation ELISA Kits (ebioscience, San Diego, CA, USA) were employed to detect the changes of immunoglobulins in sera. The sandwich ELISA was conducted according to the manufacturer’s protocols. In brief, adequate amounts of mouse serum samples were transferred to a 96-well plate coated with capture antibody. The plate was incubated for 1 h and applied with secondary antibody conjugated with HRP for more specific binding. After three times washing, tetramethyl benzidine (TMB) was added for target immunoglobin detection. Optical density (OD) were measured at 450 nm in microplate reader 3550 (Bia-rad, Hercules, CA, USA).
2.6.3. Analysis of Immune Gene Expression by Quantitative Real-Time Polymerase Chain Reaction (QRT-PCR)
RNAiso Plus (1 mL) (TaKaRa Bio, Kusatsu, Japan) was added into blood samples to get RNA extract. Total RNA extract was reverse-transcribed at 42 °C for 15 min using TransGen., TransScript All-in-one First-Strand cDNA Synthesis Super Mix for qPCR (One-step gDNA Removal) as described in the manufacturer’s guide. Primers were designed and synthesized according to the cDNA sequence of mouse β-actin, interleukin-1 (IL-1), interleukin-2 (IL-2), interleukin-4 (IL-4), interleukin-6 (IL-6), interleukin-7 (IL-7), interleukin-12 (IL-12), interleukin-15 (IL-15), interleukin-23 (IL-23), toll-like receptor 1 (TLR1), toll-like receptor 4 (TLR4), toll-like receptor 6 (TLR6), toll-like receptor 9 (TLR9), CAMP, interferon γ (IFN-γ), and tumor necrosis factor α (TNF-α), reported in GenBank (
Table 2).
Real-time PCR was performed on the iQ5 PCR detection system (Bio-Rad, Singapore) with an initial denaturation for 30 s at 95 °C, followed by 40 cycles of 5 s at 95 °C, 30 s at optimal annealing temperature, at the end of all cycles, a melt curve, from 65 °C to 95 °C per 6 s. Each RT-PCR reaction was carried out in a total volume of 15 µL, including 1 µL cDNA, 7.5 µL SsoAdvanceTM Universal SYBR Green Supermix (BioRad, Singapore), and 0.25 µL each of paired primers and 6 µL ddH
2O. Beta-actin gene was used as the reference and the mRNA levels of immunity-related genes were calculated by the geometric means method and the formula: relative level = 2
−ΔΔCt [
17].
2.7. Statistical Analysis
GraphPad Prism6 (Graphpad Software, San Diego, CA, USA) was used to manage and analyze the data. Comparisons between two groups were performed using the two-tailed Student’s t-test. p < 0.05 was set to be significant.
4. Discussion
The crisis of resistance is a continuously growing problem due to abusing antibiotics, so there is a pressing need for the replacement of antibiotics. AMPs, as essential components of a multicellular immune system, are currently used in development of an anti-infective drug. Studies show that AMPs exhibit pharmacodynamic properties, which will prevent the evolution of resistance in target microbes [
18]. With regard to animal immune modulator, cytokines are being intensively studied due to their complicated and vital effects on immune response [
19]; for instance, IL-4 and IL-6 can enhance the development of Th2 cells and antibody production, which are necessary for defense against extracellular pathogens [
20]. Combing the advantages of these, we wanted to explore whether AMPs and cytokines can synergistically enhance the immune response and anti-infection of mice.
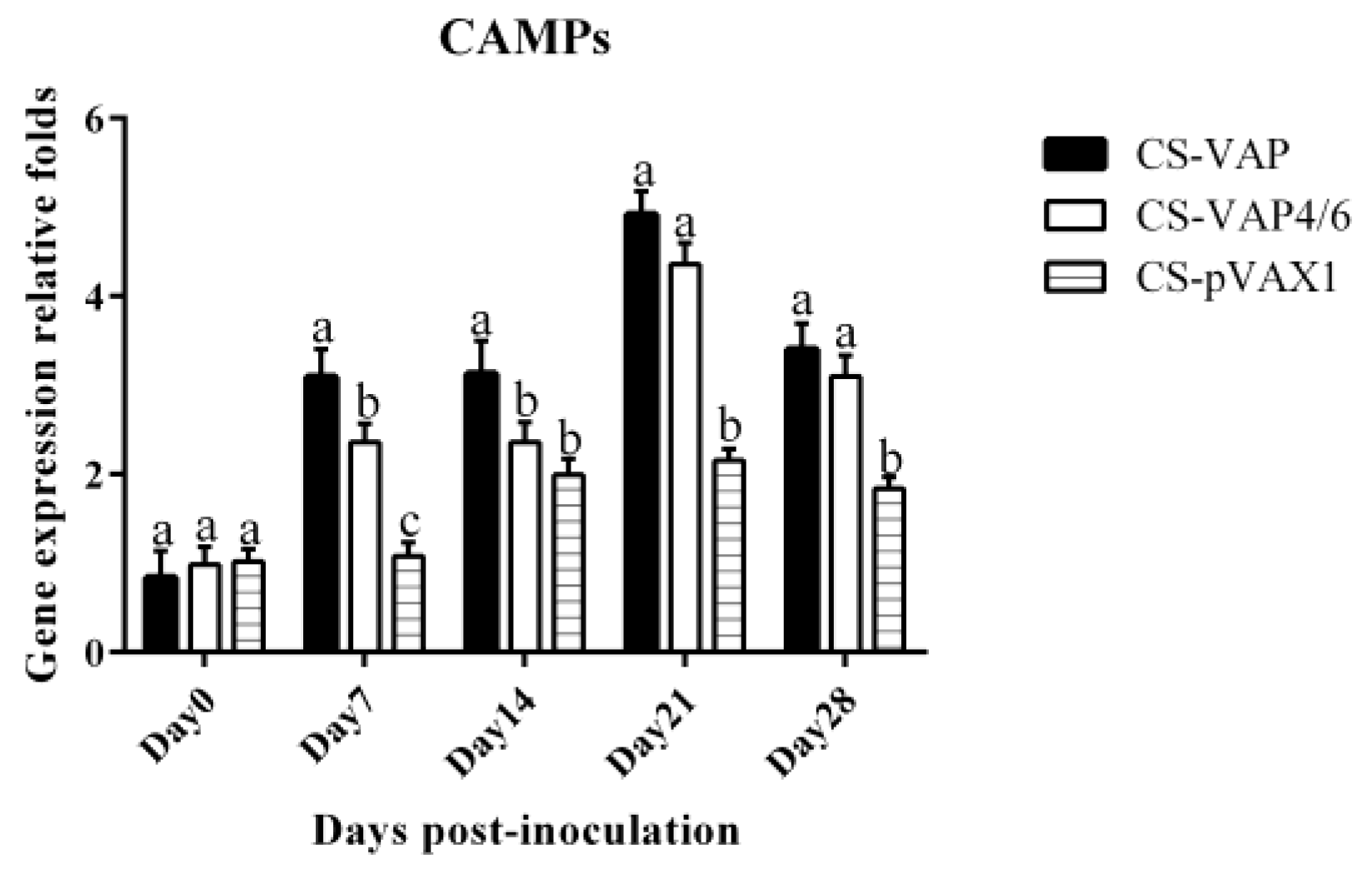
In this study, recombinant plasmids VAP and VAP4/6 were constructed for the first time and encapsulated with chitosan nanoparticles. We then assessed the immunological changes induced by the expression of VAP and VAP4/6 in vivo. In our experiment, the special 2A sequence was used to link CAMPs and IL-4/6 to produce two fusion proteins. This rational molecular design guarantees the bioactivity of CAMPs and IL-4/6, which was confirmed by the in vitro lymphocytes proliferation. There is no significant difference of CAMPs expression between CS-VAP and CS-VAP4/6 on 0, 21, and 28 days. However, on 7 and 14 days, CS-VAP had a higher expression level of CAMPs due to its possible more gene copies.
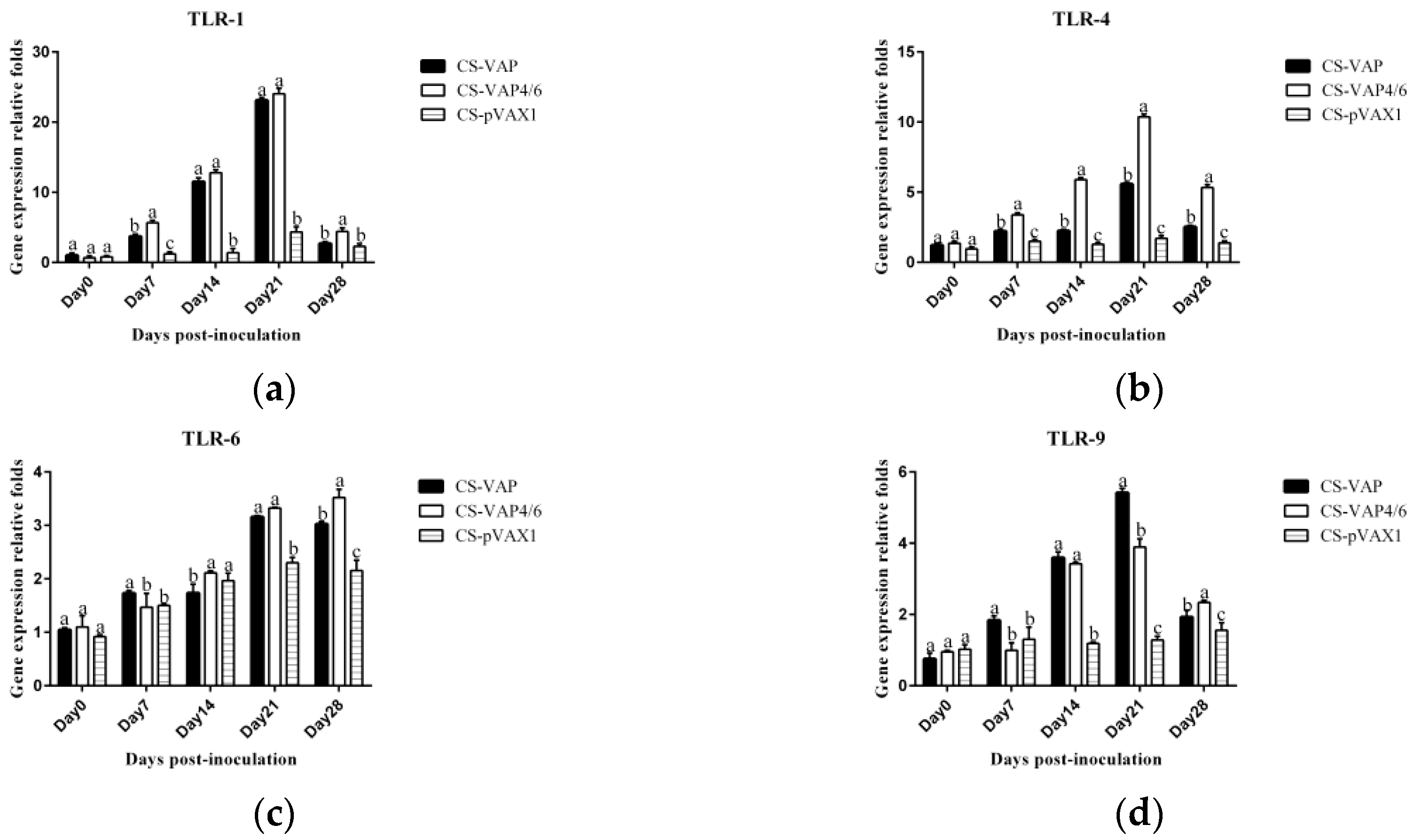
The TLRs, a family member of pattern-recognition receptors (PRRs), are transmembrane proteins that have specialized recognition of bacteria. Through pattern-recognition receptors (PRRs), innate immune cells are able to detect microbial pathogens. TLR1, TLR4, and TLR6 primarily recognize molecules unique to bacteria, and thus, allow for efficient discrimination from non-self [
21]. TLR9 is typically considered as a sensor for CpG DNA, which is relevant to viral and bacterial infections [
22]. Interestingly, a large number of AMPs have been found to induce TLR9 signaling through AMP-dsDNA complexes [
23,
24]. We observed up-regulations of TLR1, TLR4, TLR6, and TLR9 in CS-VAP and CS-VAP4/6 groups, implying the enhanced recognition of pathogens, which also coincided with increased IL-1, IL-2, IL-12, IFN-γ, and TNF-α. IL-1 is a pro-inflammatory factor that promotes antibody production and induces IL-2 production through T-cell helper cells. IL-2, IL-12, IFN-γ, and TNF-α can promote Th1 cell-mediated immune responses to target intracellular pathogens. The changes of cytokine levels are generally correlated with the increase of CD4+T and CD8+T cells. In
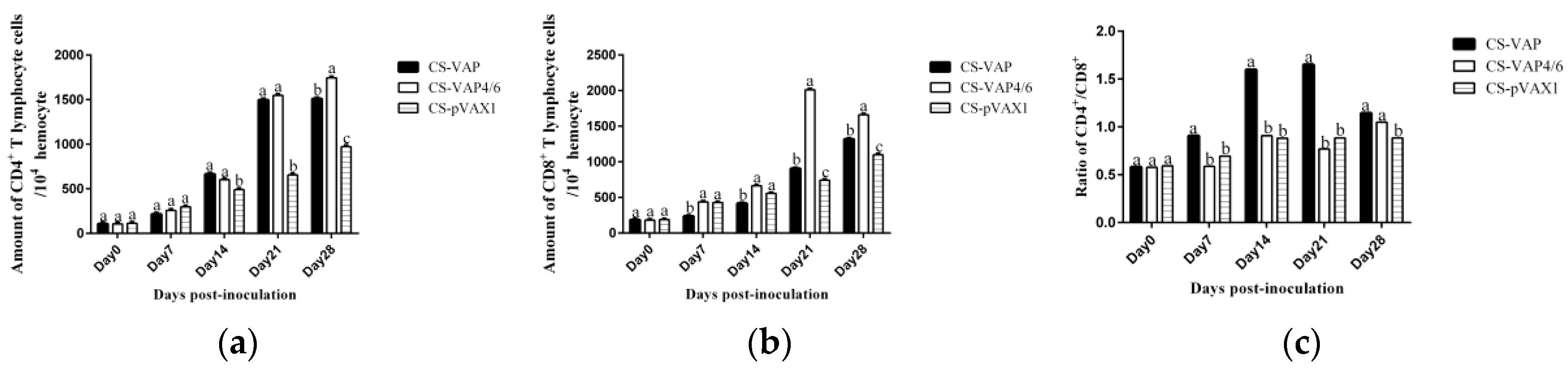
Figure 11, especially on 14, 21, and 28 days dpi, the increment of IL-1, IL-4, IL-6, IL-12, TNF-α, and IFN-γ ordinarily match up with the increase of CD4+ and CD8+ T lymphocytes in
Figure 6a,b, except for 7 dpi in
Figure 6c. The possible reason may be the diversified resource of cytokines from different immune cells. Other immune cells besides T lymphocytes, such as macrophages and dendritic cells, also produced cytokines into blood upon stimulation, which can affect the changes of cytokines in the sera.
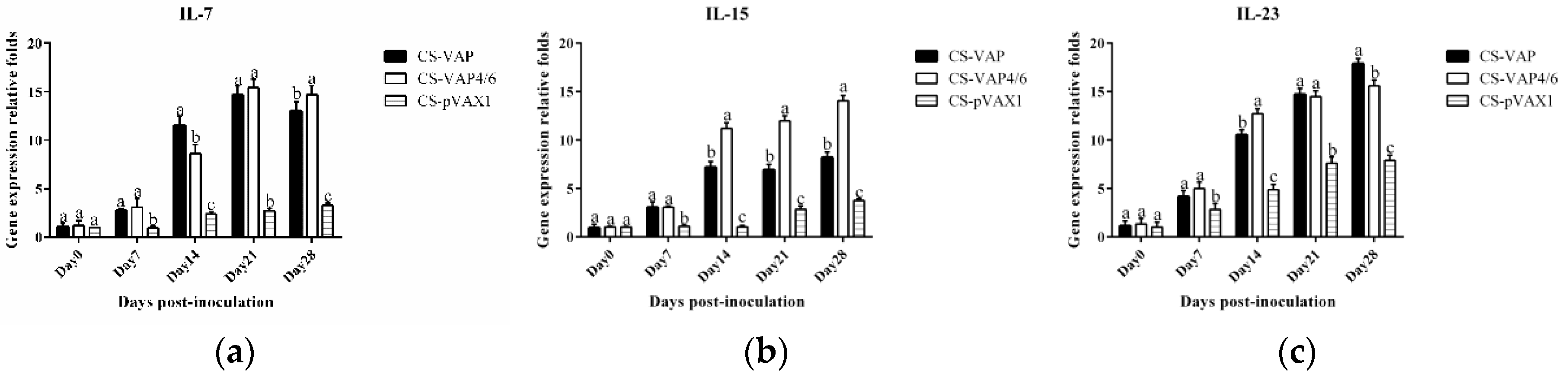
Moreover, cytokines IL-7 and IL-15 are able to regulate the pool of memory T cells corporately through low-level homeostatic division. IL-7 is primarily important for survival of memory CD8+ cells while IL-15 is vital for low-level proliferation to maintain the size of memory T-cells pool. IL-23 is best known for its ability to promote Th17 maturation. The raised expressions of IL-7, IL-15, and IL-23 in VAP and VAP4/6 mice indicated the up-regulation of the immune memory, which is consistent with the amplification of cellular immunity resulting from the addition of CD4+ and CD8+ T cells.
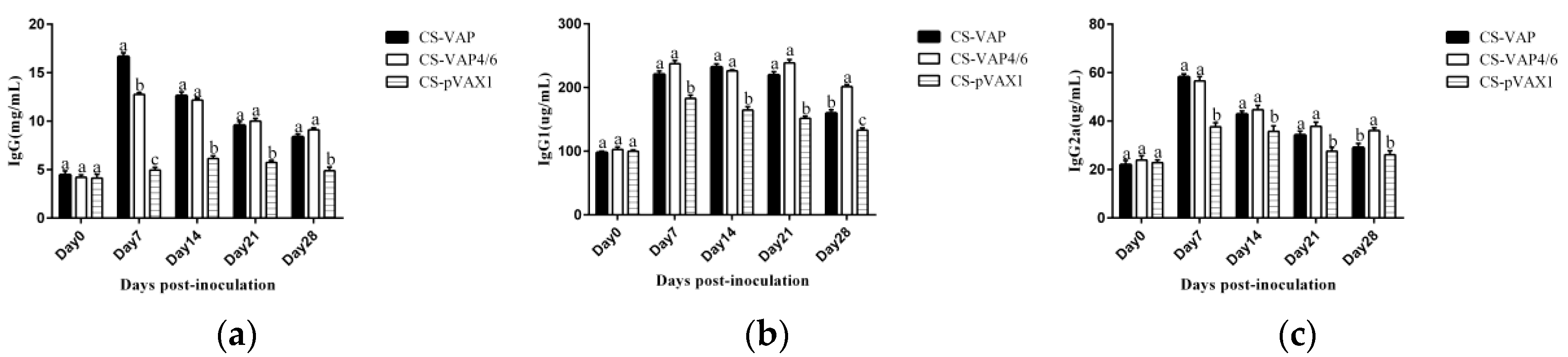
The result of flow cytometry showed that compared with the control mice, CD4+ and CD8+ T lymphocytes significantly increased in the mice treated with CS-VAP4/6 and CS-VAP, and the CS-VAP4/6-treated mice showed a higher increase than the CS-VAP group, indicating that the humoral immunity of treated mice was efficiently elevated, which was confirmed by the increased IgG, IgG1, and IgG2a in the sera. The increase of IgG was initially stimulated by the inoculation of recombinant plasmids and the expression of inserted immune genes, and it also possibly declined with the degrade of the recombinant plasmids in vivo after 7 dpi. To our knowledge, the mixed inoculation of CS-plasmid was responsible for the IGs increment in the control mice post-day 0, which probably resulted from the stimulation of specific immunity by CS-pVAX1 to the conditioned pathogen in the control mice. These results were in accordance with the reports that the expression of IL-4 gene significantly increased the quantity of IgG1- [
25,
26] and IL-6- stimulated CD4+ T cells to promote the production of IgG [
26].
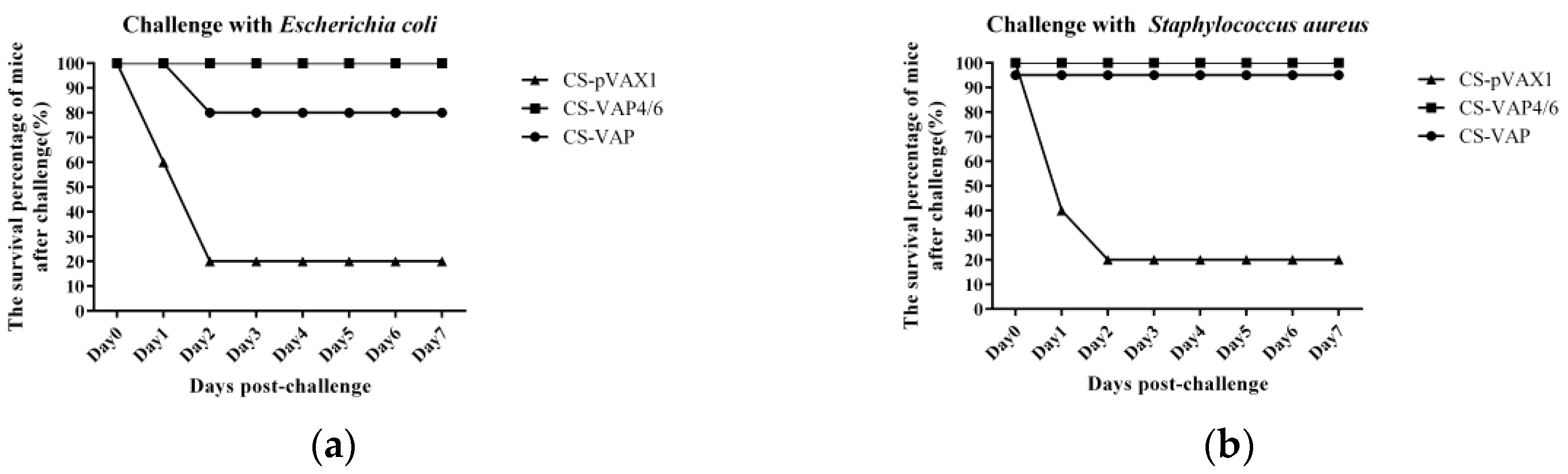
The challenge results of both E. coli and S. aureus separately showed the strong protective effect of the recombinant plasmid nanoparticles. Most of the treated mice survived and did not show any lesions or symptoms caused by the infection of bacteria. In contrast, the control mice were severely infected by the challenge bacteria, and most of them died of infection lesions and damage within 48 h. Additionally, there was no death in the next 5 days, so we stopped the observation on 7 days post-challenge due to limited time for experiment in our lab. Among the treated groups, CS-VAP4/6 manifested better improvement of innate and adaptive immunity (p < 0.05) and resulted in stronger resistance against challenge with bacteria, which is in accordance with the higher level of humoral and cellular immune responses. Similarly, the CS-VAP4/6 inoculation provoked more systemic immunity in the blood of mice compared with CS-VAP.
Chitosan was utilized as the DNA delivery material, not only for high transfection efficiency but also for its safety and compatibility in vivo [
27,
28]. So, we selected chitosan to entrap the recombinant plasmids and charge suitable nanoparticles with positive potentials, which can protect the plasmids from digestion. The properties of encapsulated nanoparticles showed that they can effectively transport the DNA to target cells. The proliferation of pig lymphocytes manifested the stimulatory effect of nanoparticles, indicating that the nanoparticles can express bioactive interleukins and CAMPs proteins, which can stimulate lymphocyte proliferation of pigs. It is noteworthy that the stimulatory effect of CS-VAP4/6 was stronger than those of CS-VAP and CS-pVAX1, demonstrating that the fusion gene has better immunological bioactivity. The enhanced expressions of cytokine genes demonstrated that the recombinant plasmid nanoparticles can not only improve the systemic immunity but also slows the process of releasing. The latter was predictable due to the chitosan function of long-term release [
29]. It was notable that the expression levels of most of the immune genes reached the highest point at 21 days post-inoculation, and it was significantly upgraded at day 28 post-inoculation, indicating that the nanoparticles have the effect of producing reliable releasing for nearly one month.
Here, the recombinant plasmid of co-expression of CAMPs and IL-4/6 was firstly constructed and entrapped with chitosan nanoparticles, which is used as a potential replacement of antibiot Many indicators consolidated that both of innate and adaptive immunity were enhanced, and the immunity against bacterial infection was also elevated for one month or so, which probably could not protect the mice in a medium- or long-term due to the limited existence of recombinant plasmid in vivo.

 ,
, 















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}