Lagging Immune Response to Haemophilus influenzae Serotype b (Hib) Conjugate Vaccine after the Primary Vaccination with Hib of Infants in The Netherlands

 ,
,
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Population
2.2. Serology
2.3. Statistical Analysis
3. Results
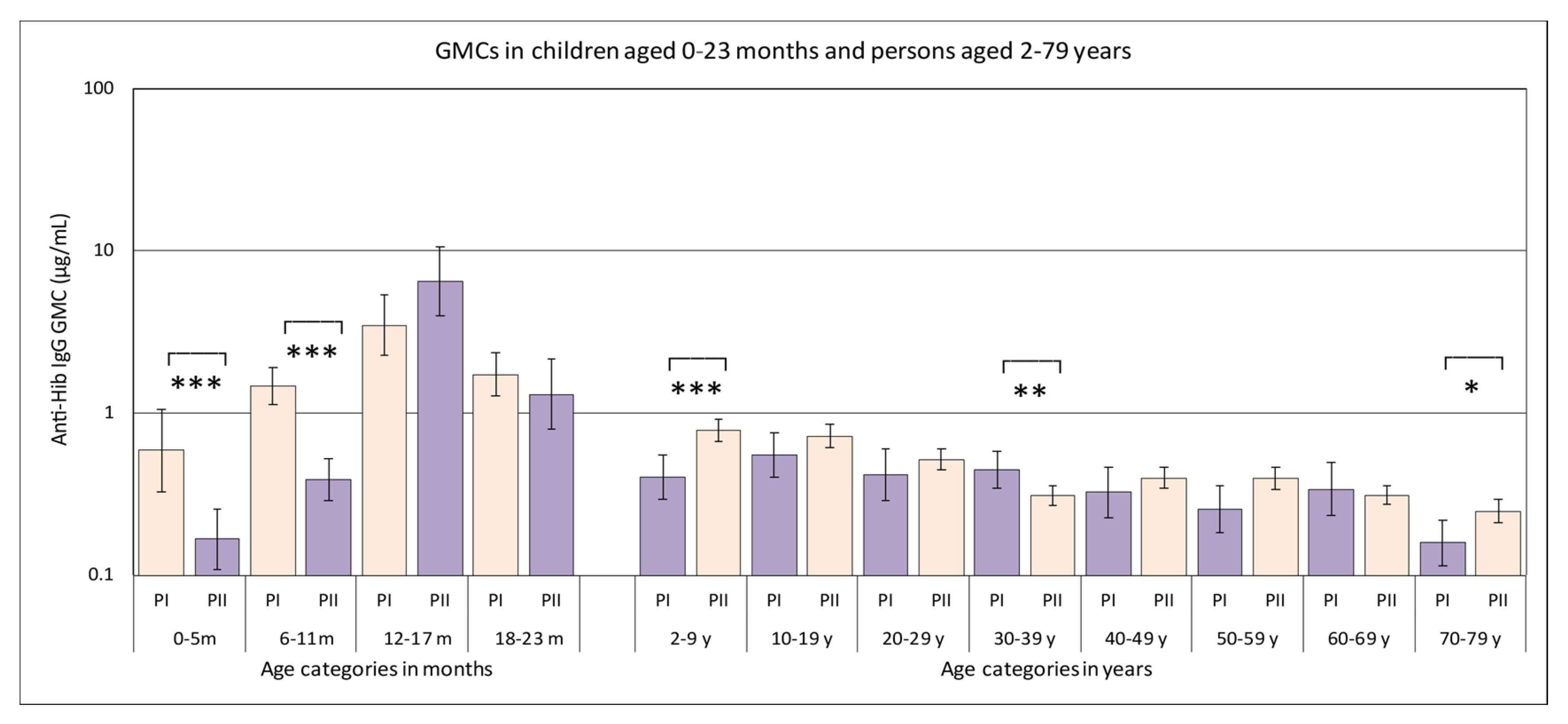
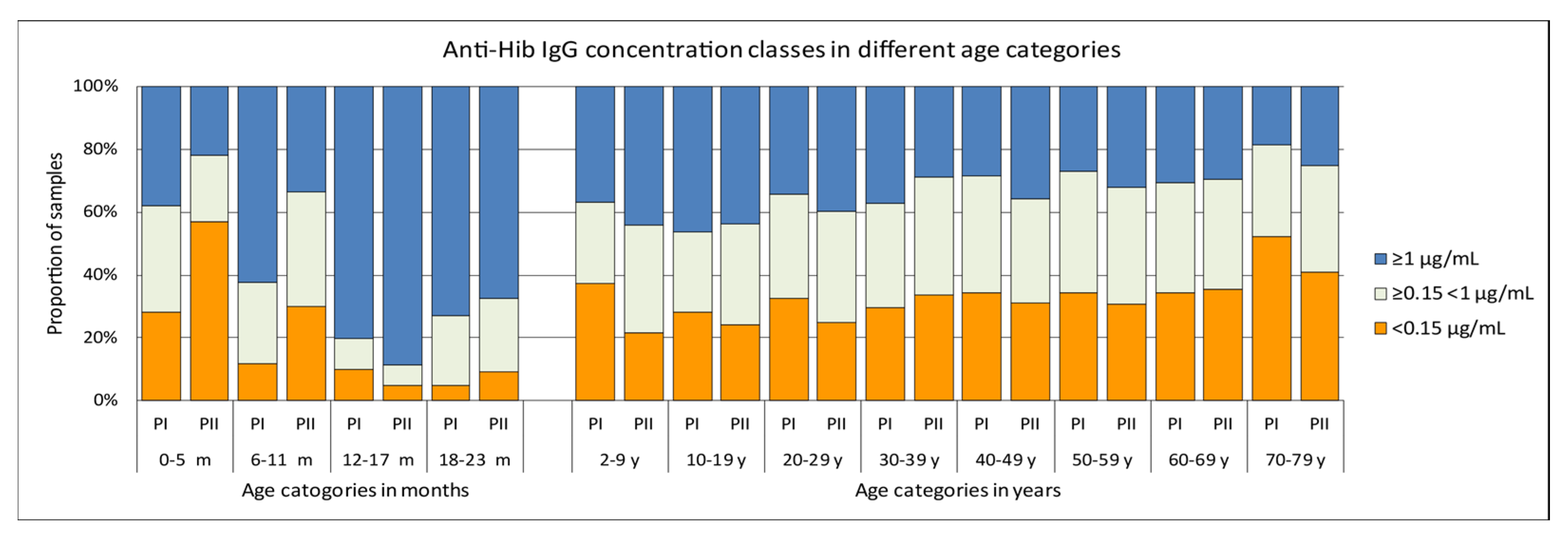
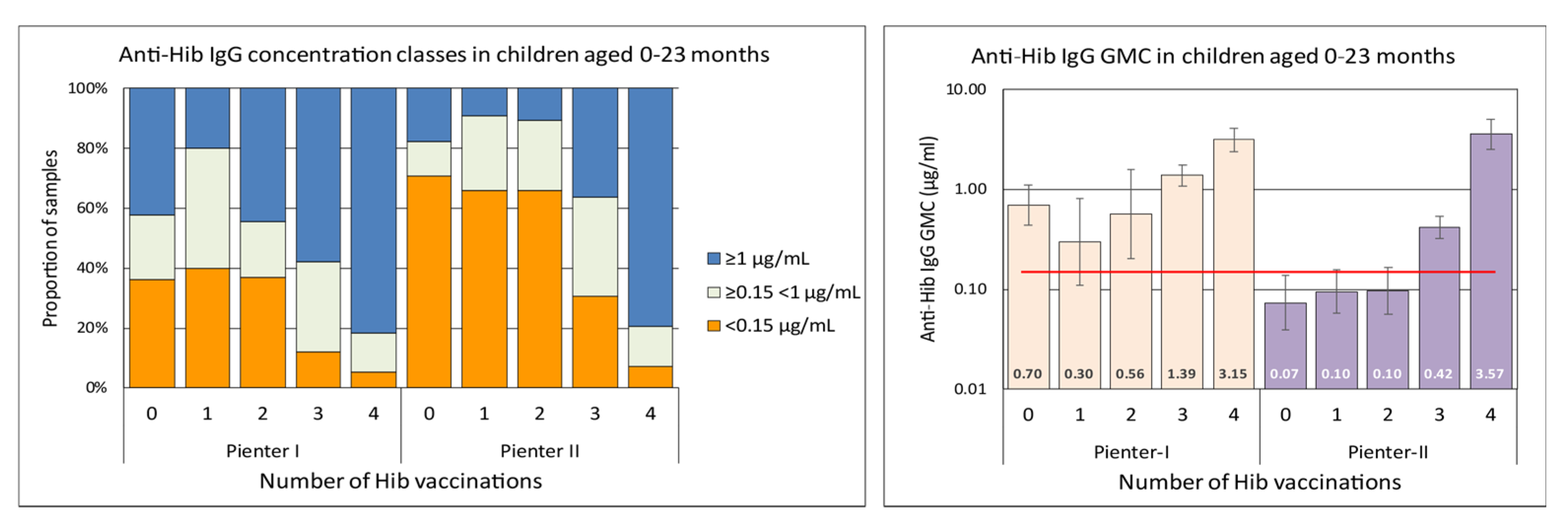
3.1. Hib-Specific IgG Levels in the Pienter-I and Pienter-II Study
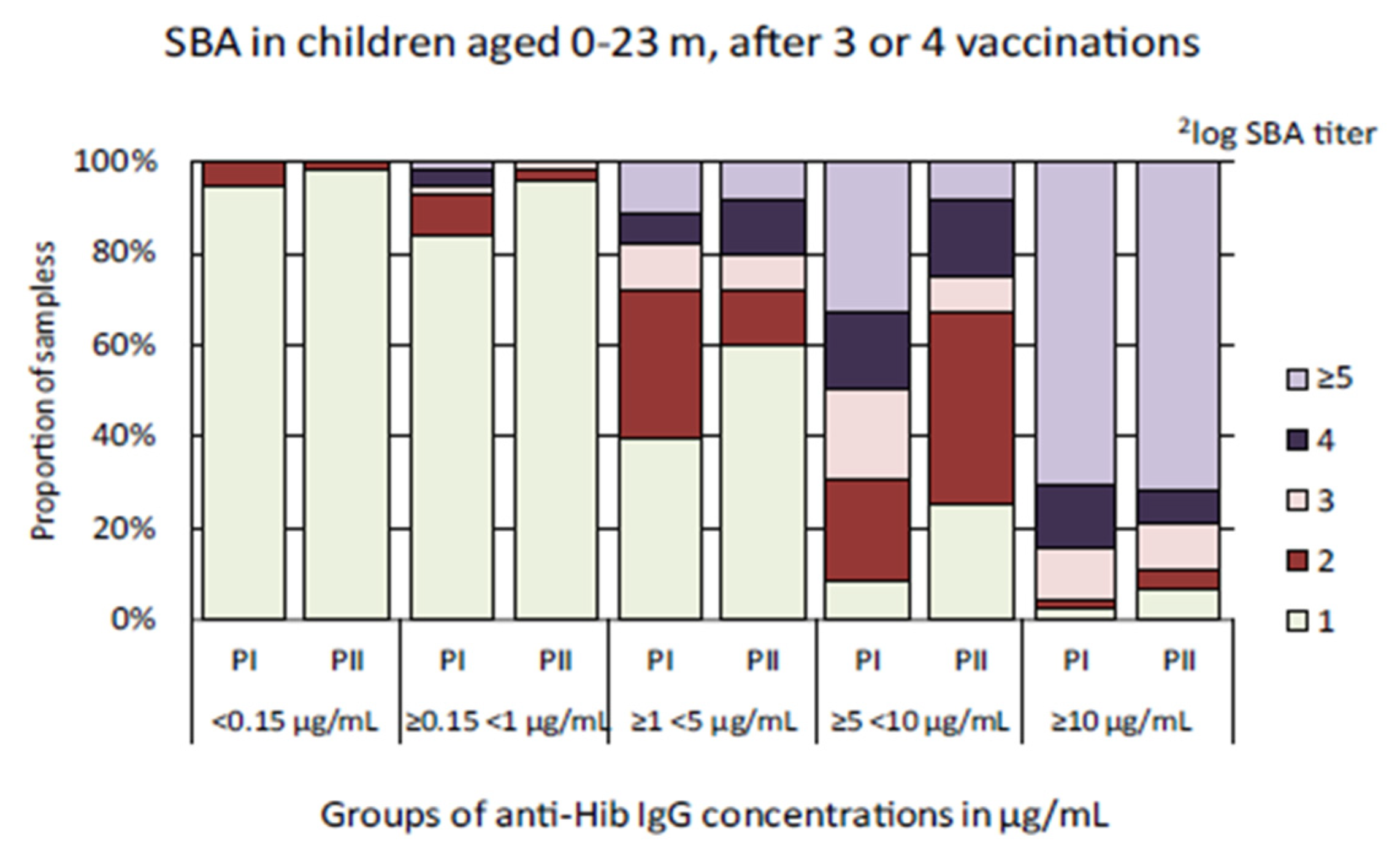
3.2. Hib-Specific Serum Bactericidal Assay
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Rijkers, G.T.; Vermeer-de Bondt, P.E.; Spanjaard, L.; Breukels, M.A.; Sanders, E.A. Return of Haemophilus influenzae type b infections. Lancet 2003, 361, 1563–1564. [Google Scholar] [CrossRef]
- Spanjaard, L.; van den Hof, S.; de Melker, H.E.; Vermeer-de Bondt, P.E.; van der Ende, A.; Rijkers, G.T. Increase in the number of invasive Haemophilus influenzae type b infections. Ned. Tijdschr. Geneeskd. 2005, 149, 2738–2742. [Google Scholar] [PubMed]
- van Den Hof, S.; de Melker, H.E.; Berbers, G.A.; van Der Kraak, P.H.; Spaendonck, M.A. Antibodies to Haemophilus influenzae serotype b in The Netherlands a few years after the introduction of routine vaccination. Clin. Infect. Dis. 2001, 32, 2–8. [Google Scholar] [CrossRef] [PubMed]
- van Lier, E.A.; Oomen, P.J.; Oostenbrug, M.W.; Zwakhals, S.L.; Drijfhout, I.H.; de Hoogh, P.A.; de Melker, H.E. High vaccination coverage of the National Immunization Programme in The Netherlands. Ned. Tijdschr. Geneeskd. 2009, 153, 950–957. [Google Scholar]
- De Melker, H.E.; Conyn-van Spaendonck, M.A. Immunosurveillance and the evaluation of national immunization programmes: A population-based approach. Epidemiol. Infect. 1998, 121, 637–643. [Google Scholar] [CrossRef]
- van der Klis, F.R.; Mollema, L.; Berbers, G.A.; de Melker, H.E.; Coutinho, R.A. Second national serum bank for population-based seroprevalence studies in The Netherlands. Neth. J. Med. 2009, 67, 301–308. [Google Scholar]
- Phipps, D.C.; West, J.; Eby, R.; Koster, M.; Madore, D.V.; Quataert, S.A. An ELISA employing a Haemophilus influenzae type b oligosaccharide-human serum albumin conjugate correlates with the radioantigen binding assay. J. Immunol. Methods 1990, 135, 121–128. [Google Scholar] [CrossRef]
- Mariani, M.; Luzzi, E.; Proietti, D.; Mancianti, S.; Casini, D.; Costantino, P.; van Gageldonk, P.; Berbers, G. A competitive enzyme-linked immunosorbent assay for measuring the levels of serum antibody to Haemophilus influenzae type b. Clin. Diagn. Lab. Immunol. 1998, 5, 667–674. [Google Scholar] [CrossRef]
- de Voer, R.M.; Schepp, R.M.; Versteegh, F.G.; van der Klis, F.R.; Berbers, G.A. Simultaneous detection of Haemophilus influenzae type b polysaccharide-specific antibodies and Neisseria meningitidis serogroup A, C, Y, and W-135 polysaccharide-specific antibodies in a fluorescent-bead-based multiplex immunoassay. Clin. Vaccine Immunol. 2009, 16, 433–436. [Google Scholar] [CrossRef]
- Schouls, L.; van der Heide, H.; Witteveen, S.; Zomer, B.; van der Ende, A.; Burger, M.; Schot, C. Two variants among Haemophilus influenzae serotype b strains with distinct bcs4, hcsA and hcsB genes display differences in expression of the polysaccharide capsule. BMC Microbiol. 2008, 8, 35. [Google Scholar] [CrossRef]
- WHO. Immunological Basis for Immunization: Haemophilus Influenzae Type B Vaccines; World Health Organization: Geneva, Switzerland, 2007. [Google Scholar]
- de Melker, H.E.; van den Hof, S.; Berbers, G.A.; Conyn-van Spaendonck, M.A. Evaluation of the national immunisation programme in The Netherlands: Immunity to diphtheria, tetanus, poliomyelitis, measles, mumps, rubella and Haemophilus influenzae type b. Vaccine 2003, 21, 716–720. [Google Scholar] [CrossRef]
- Eskola, J.; Kayhty, H. Early immunization with conjugate vaccines. Vaccine 1998, 16, 1433–1438. [Google Scholar] [CrossRef]
- Pollabauer, E.M.; Petermann, R.; Ehrlich, H.J. The influence of carrier protein on the immunogenicity of simultaneously administered conjugate vaccines in infants. Vaccine 2009, 27, 1674–1679. [Google Scholar] [CrossRef] [PubMed]
- Dagan, R.; Poolman, J.; Siegrist, C.A. Glycoconjugate vaccines and immune interference: A review. Vaccine 2010, 28, 5513–5523. [Google Scholar] [CrossRef]
- Carlsson, R.M.; Claesson, B.A.; Kayhty, H.; Selstam, U.; Iwarson, S. Studies on a Hib-tetanus toxoid conjugate vaccine: Effects of co-administered tetanus toxoid vaccine, of administration route and of combined administration with an inactivated polio vaccine. Vaccine 1999, 18, 468–478. [Google Scholar] [CrossRef]
- Rennels, M.B.; Englund, J.A.; Bernstein, D.I.; Losonsky, G.A.; Anderson, E.L.; Pichichero, M.E.; Munoz, F.M.; Wolff, M.C. Diminution of the anti-polyribosylribitol phosphate response to a combined diphtheria-tetanus-acellular pertussis/Haemophilus influenzae type b vaccine by concurrent inactivated poliovirus vaccination. Pediatr. Infect. Dis. J. 2000, 19, 417–423. [Google Scholar] [CrossRef] [PubMed]
- Eskola, J.; Ward, J.; Dagan, R.; Goldblatt, D.; Zepp, F.; Siegrist, C.A. Combined vaccination of Haemophilus influenzae type b conjugate and diphtheria-tetanus-pertussis containing acellular pertussis. Lancet 1999, 354, 2063–2068. [Google Scholar] [CrossRef]
- Daum, R.S.; Zenko, C.E.; Given, G.Z.; Ballanco, G.A.; Parikh, H.; Germino, K. Magnitude of interference after diphtheria-tetanus toxoids-acellular pertussis/Haemophilus influenzae type b capsular polysaccharide-tetanus vaccination is related to the number of doses administered. J. Infect. Dis. 2001, 184, 1293–1299. [Google Scholar] [CrossRef] [PubMed]
- McVernon, J.; Andrews, N.; Slack, M.P.; Ramsay, M.E. Risk of vaccine failure after Haemophilus influenzae type b (Hib) combination vaccines with acellular pertussis. Lancet 2003, 361, 1521–1523. [Google Scholar] [CrossRef]
- Southern, J.; McVernon, J.; Gelb, D.; Andrews, N.; Morris, R.; Crowley-Luke, A.; Goldblatt, D.; Miller, E. Immunogenicity of a fourth dose of Haemophilus influenzae type b (Hib) conjugate vaccine and antibody persistence in young children from the United Kingdom who were primed with acellular or whole-cell pertussis component-containing Hib combinations in infancy. Clin. Vaccine Immunol. 2007, 14, 1328–1333. [Google Scholar]
- Collins, S.; Litt, D.; Almond, R.; Findlow, J.; Linley, E.; Ramsay, M.; Borrow, R.; Ladhani, S. Haemophilus influenzae type b (Hib) seroprevalence and current epidemiology in England and Wales. J. Infect. 2018, 76, 335–341. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Age Category | Pienter I | Pienter II | ||||
|---|---|---|---|---|---|---|
| n | GMC | 95% CI | n | GMC | 95% CI | |
| 0–5 month *** | 50 | 0.59 | 0.33–1.05 | 114 | 0.17 | 0.11–0.25 |
| 6–11 month *** | 201 | 1.48 | 1.14–1.92 | 181 | 0.39 | 0.29–0.52 |
| 12–17 month | 112 | 3.48 | 2.26–5.36 | 61 | 6.51 | 4.01–10.56 |
| 18–23 month | 85 | 1.73 | 1.28–2.34 | 43 | 1.31 | 0.80–2.14 |
| 2–9 year *** | 212 | 0.40 | 0.30–0.55 | 680 | 0.78 | 0.67–0.92 |
| 10–19 year | 225 | 0.55 | 0.40–0.75 | 694 | 0.72 | 0.61–0.85 |
| 20–29 year | 129 | 0.42 | 0.29–0.60 | 693 | 0.52 | 0.45–0.60 |
| 30–39 year ** | 224 | 0.44 | 0.34–0.58 | 705 | 0.31 | 0.27–0.36 |
| 40–49 year | 168 | 0.32 | 0.23–0.46 | 630 | 0.40 | 0.34–0.46 |
| 50–59 year | 171 | 0.26 | 0.18–0.36 | 628 | 0.39 | 0.34–0.46 |
| 60–69 year | 131 | 0.34 | 0.23–0.49 | 721 | 0.31 | 0.27–0.36 |
| 70–79 year * | 144 | 0.16 | 0.11–0.22 | 548 | 0.25 | 0.21–0.29 |
| Anti-Hib IgG Concentration Range | Study | n | GMC | 95% CI | GMT | 95% CI |
|---|---|---|---|---|---|---|
| <0.15 µg/mL | Pienter-I | 18 | 0.05 | 0.04–0.08 | 1.04 | 0.96–1.13 |
| Pienter-II | 48 | 0.04 | 0.03–0.06 | 1.02 | 0.99–1.04 | |
| ≥0.15 < 1 µg/mL * | Pienter-I | 56 | 0.38 | 0.33–0.44 | 1.17 | 1.06–1.30 |
| Pienter-II | 65 | 0.39 | 0.34–0.45 | 1.04 | 0.99–1.09 | |
| ≥1 < 5 µg/mL | Pienter-I | 78 | 2.28 | 2.08–2.51 | 1.87 | 1.63–2.14 |
| Pienter-II | 50 | 2.19 | 1.90–2.51 | 1.63 | 1.35–1.97 | |
| ≥5 <10 µg/mL * | Pienter-I | 36 | 7.08 | 6.64–7.54 | 3.25 | 2.70–3.92 |
| Pienter-II | 12 | 6.83 | 5.90–7.93 | 2.19 | 1.47–3.27 | |
| ≥10 µg/mL | Pienter-I | 44 | 18.81 | 15.65–22.61 | 5.21 | 4.56–5.96 |
| Pienter-II | 28 | 23.34 | 18.58–29.32 | 4.66 | 3.78–5.75 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Schouls, L.; Schot, C.; de Voer, R.M.; van der Klis, F.; Knol, M.; Tcherniaeva, I.; Berbers, G. Lagging Immune Response to Haemophilus influenzae Serotype b (Hib) Conjugate Vaccine after the Primary Vaccination with Hib of Infants in The Netherlands. Vaccines 2020, 8, 347. https://doi.org/10.3390/vaccines8030347
Schouls L, Schot C, de Voer RM, van der Klis F, Knol M, Tcherniaeva I, Berbers G. Lagging Immune Response to Haemophilus influenzae Serotype b (Hib) Conjugate Vaccine after the Primary Vaccination with Hib of Infants in The Netherlands. Vaccines. 2020; 8(3):347. https://doi.org/10.3390/vaccines8030347
Chicago/Turabian StyleSchouls, Leo, Corrie Schot, Richarda M. de Voer, Fiona van der Klis, Mirjam Knol, Irina Tcherniaeva, and Guy Berbers. 2020. "Lagging Immune Response to Haemophilus influenzae Serotype b (Hib) Conjugate Vaccine after the Primary Vaccination with Hib of Infants in The Netherlands" Vaccines 8, no. 3: 347. https://doi.org/10.3390/vaccines8030347
APA StyleSchouls, L., Schot, C., de Voer, R. M., van der Klis, F., Knol, M., Tcherniaeva, I., & Berbers, G. (2020). Lagging Immune Response to Haemophilus influenzae Serotype b (Hib) Conjugate Vaccine after the Primary Vaccination with Hib of Infants in The Netherlands. Vaccines, 8(3), 347. https://doi.org/10.3390/vaccines8030347