Pandemic Influenza Vaccines: What did We Learn from the 2009 Pandemic and are We Better Prepared Now?


Abstract
1. Introduction
2. Global Capacity of Influenza Vaccine Production
3. Currently Available Influenza Vaccines
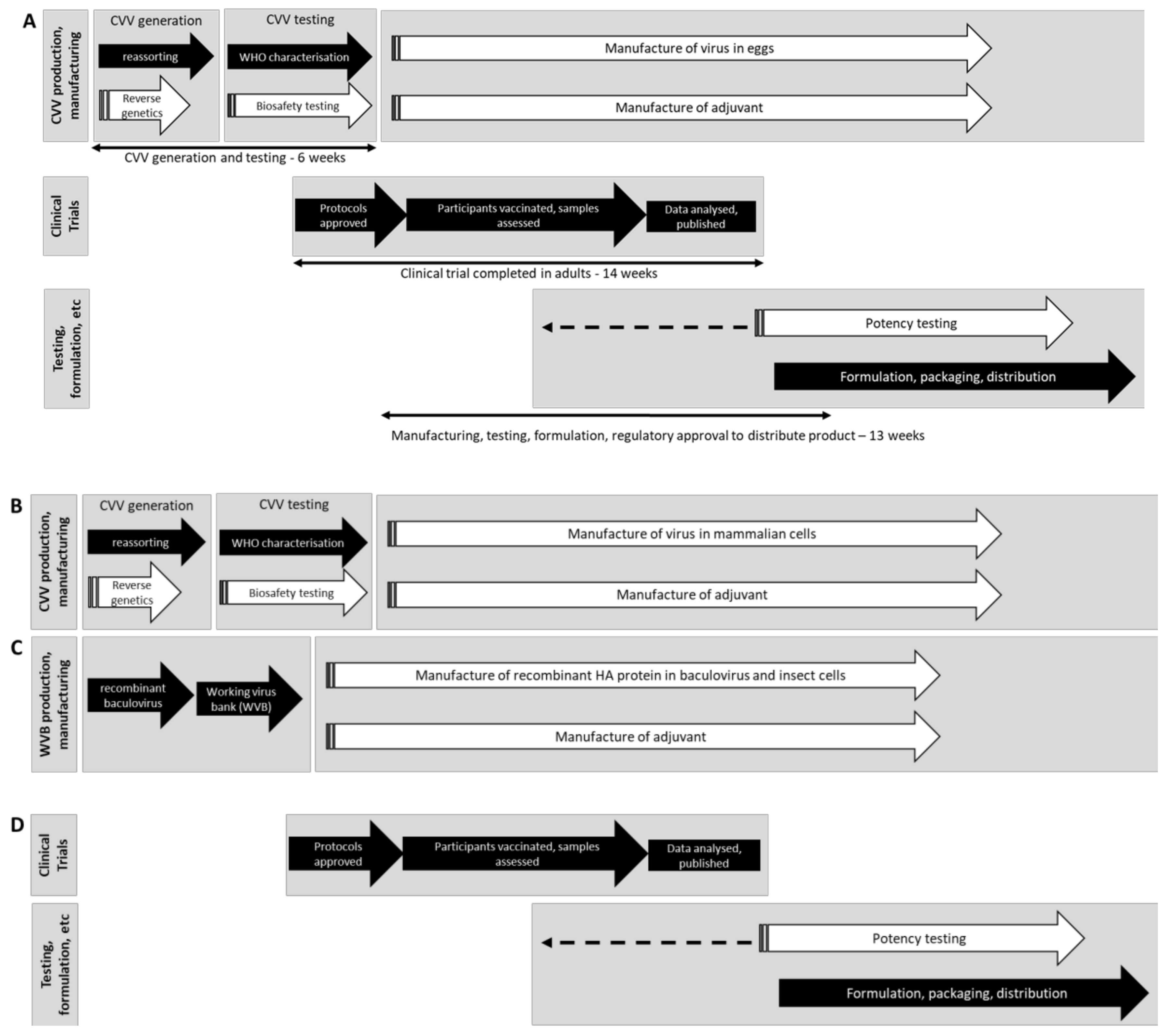
4. Challenges That Influenza Vaccine Manufacturers Encountered in 2009 and the Key Changes That have Been Made Since
4.1. Biocontainment
4.2. Development of RG and Cell Culture CVVs
4.3. Availability of Pre-Pandemic Stockpiles and Pre-Pandemic CVVs
4.4. Adjuvants
4.5. Standardisation and Release of Vaccines
5. What Challenges Remain for the Next Influenza Pandemic Vaccine?
5.1. Future Pandemic Vaccine Immunogenicity Remains an Unknown
5.2. Switching Vaccine Manufacturing from Seasonal Vaccines to Pandemic Vaccine
5.3. Pandemic Surveillance
5.4. The Nagoya Protocol and WHO Pandemic Influenza Preparedness Framework
6. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Taubenberger, J.K.; Kash, J.C.; Morens, D.M. The 1918 influenza pandemic: 100 years of questions answered and unanswered. Sci. Transl. Med. 2019, 11, eaau5485. [Google Scholar] [CrossRef] [PubMed]
- Rajagopal, S.; Treanor, J. Pandemic (avian) influenza. Semin. Respir. Crit. Care Med. 2007, 28, 159–170. [Google Scholar] [CrossRef] [PubMed]
- Minor, P.D. Assaying the potency of influenza vaccines. Vaccines 2015, 3, 90–104. [Google Scholar] [CrossRef] [PubMed]
- First Global Estimates of 2009 H1N1 Pandemic Mortality Released by CDC-Led Collaboration. Available online: https://www.cdc.gov/flu/spotlights/pandemic-global-estimates.htm (accessed on 25 October 2019).
- Clark, N.M.; Lynch, J.P. Influenza: Epidemiology, clinical features, therapy, and prevention. Semin. Respir. Crit. Care Med. 2011, 32, 373–392. [Google Scholar] [CrossRef]
- Collin, N.; de Radiguès, X. WHO H1N1 vaccine task force, vaccine production capacity for seasonal and pandemic (H1N1) 2009 influenza. Vaccine 2009, 27, 5184–5186. [Google Scholar] [CrossRef]
- Population Estimates: Year One through 2050. A.D. Available online: https://www.ecology.com/population-estimates-year-2050/ (accessed on 20 October 2019).
- McLean, K.A.; Goldin, S.; Nannei, C.; Sparrow, E.; Torelli, G. The 2015 global production capacity of seasonal and pandemic influenza vaccine. Vaccine 2016, 34, 5410–5413. [Google Scholar] [CrossRef]
- Update on Production Capacity for Seasonal and Pandemic Influenza GAP III 2006–2016. Available online: https://www.who.int/influenza_vaccines_plan/GAPIII_Session2_Friede.pdf (accessed on 1 October 2019).
- Global Action Plan for Influenza Vaccines. Available online: https://www.who.int/influenza_vaccines_plan/en/ (accessed on 1 October 2019).
- Girard, M.P.; Tam, J.S.; Assossou, O.M.; Kieny, M.P. The 2009 A (H1N1) influenza virus pandemic: A review. Vaccine 2010, 28, 4895–4902. [Google Scholar] [CrossRef]
- PIP Contributions. Available online: http://open.who.int/2018–19/pip (accessed on 17 January 2020).
- Developing Countries Vaccine Manufacturers Network. Available online: https://www.dcvmn.org/- (accessed on 18 April 2020).
- International Federation of Pharmaceutical Manufacturers & Associations. Influenza Vaccine Supply (IVS) International Task Force. Available online: https://www.ifpma.org/subtopics/influenza-vaccine/ (accessed on 17 January 2020).
- Wong, S.S.; Webby, R.J. Traditional and new influenza vaccines. Clin. Microbiol. Rev. 2013, 26, 476–492. [Google Scholar] [CrossRef]
- Yusibov, V.; Kushnir, N.; Streatfield, S.J. Advances and challenges in the development and production of effective plant-based influenza vaccines. Expert Rev. Vaccines 2015, 14, 519–535. [Google Scholar]
- Shinde, V.; Fries, L.; Wu, Y.; Agrawal, S.; Cho, I.; Thomas, D.N.; Spindler, M.; Lindner, E.; Hahn, T.; Plested, J.; et al. Improved titers against influenza drift variants with a nanoparticle vaccine. N. Engl. J. Med. 2018, 378, 2346–2348. [Google Scholar] [CrossRef]
- Portnoff, A.D.N.; Massare, M.J.; Zhou, H.; Tian, J.-H.; Zhou, B.; Shinde, V.; Glenn, G.M.; Smith, G. Influenza hemagglutinin nanoparticle vaccine elicits broadly neutralizing antibodies against structurally distinct domains of H3N2 HA. Vaccines 2020, 8, 99. [Google Scholar] [CrossRef] [PubMed]
- Bernstein, D.I.; Guptill, J.; Naficy, A.; Nachbagauer, R.; Berlanda-Scorza, F.; Feser, J.; Wilson, P.C.; Solórzano, A.; Van der Wielen, M.; Walter, E.B.; et al. Immunogenicity of chimeric haemagglutinin-based, universal influenza virus vaccine candidates: Interim results of a randomised, placebo-controlled, phase 1 clinical trial. Lancet Infect. Dis. 2020, 20, 80–91. [Google Scholar] [CrossRef]
- Neumann, G.; Watanabe, T.; Ito, H.; Watanabe, S.; Goto, H.; Gao, P.; Hughes, M.; Perez, D.R.; Donis, R.; Hoffmann, E.; et al. Generation of influenza A viruses entirely from cloned cDNAs. Proc. Natl. Acad. Sci. USA 1999, 96, 9345–9350. [Google Scholar] [CrossRef] [PubMed]
- Barr, I.G.; Donis, R.O.; Katz, J.M.; McCauley, J.W.; Odagiri, T.; Trusheim, H.; Tsai, T.F.; Wentworth, D.E. Cell culture-derived influenza vaccines in the severe 2017–2018 epidemic season: A step towards improved influenza vaccine effectiveness. npj Vaccines 2018, 3, 44. [Google Scholar] [CrossRef] [PubMed]
- Buckland, B.; Boulanger, R.; Fino, M.; Srivastava, I.; Holtz, K.; Khramtsov, N.; McPherson, C.; Meghrous, J.; Kubera, P.; Cox, M.M.J.; et al. Technology transfer and scale-up of the Flublok® recombinant hemagglutinin (HA) influenza vaccine manufacturing process. Vaccine 2014, 32, 5496–5502. [Google Scholar] [CrossRef]
- Rockman, S.; Schoofs, P.; Greenberg, M. Development and testing of the Australian pandemic influenza vaccine—A timely response. Microbiol. Aust. 2011, 32, 11–14. [Google Scholar] [CrossRef]
- Executive Order on Modernizing Influenza Vaccines in the United States to Promote National Security and Public Health. Available online: https://www.whitehouse.gov/presidential-actions/executive-order-modernizing-influenza-vaccines-united-states-promote-national-security-public-health/ (accessed on 19 September 2019).
- Update of WHO Biosafety Risk Assessment and Guidelines for the Production and Quality Control of Human Influenza Pandemic Vaccines, 28 May 2009. Available online: https://www.who.int/biologicals/publications/trs/areas/vaccines/influenza/H1N1_vaccine_production_biosafety_SHOC.27May2009.pdf (accessed on 25 October 2019).
- Donis, R.O.; Davis, C.T.; Foust, A.; Hossain, M.J.; Johnson, A.; Klimov, A.; Loughlin, R.; Xu, X.; Tsai, T.; et al.; Influenza Cell Culture Working Group Performance characteristics of qualified cell lines for isolation and propagation of influenza viruses for vaccine manufacturing. Vaccine 2014, 32, 6583–6590. [Google Scholar] [CrossRef]
- WHO. Working Group Meeting on Revision of WHO TRS 941, Annex 5: WHO Biosafety Risk Assessment and Guidelines for the Production and Quality Control of Human Influenza Pandemic Vaccines. Available online: https://www.who.int/biologicals/areas/vaccines/INFLUENZA_WG_mtg_TRS941_Annex_5_12_Oct_2017_for_web.pdf?ua=1 (accessed on 18 April 2020).
- Guidelines for the Safe Development and Production of Vaccines to Human Pandemic Influenza Viruses and Viruses with Pandemic Potential. Revision of Annex 5 of WHO Technical Report Series WHO TRS No. 941, Annex 5; H1N1 Specific Update 2009; and H7N9 Update 2013. Available online: https://www.who.int/biologicals/expert_committee/POST_ECBS_2018_Pandemic_influenza_12_Nov_2018.pdf (accessed on 18 April 2020).
- Webby, R.J.; Perez, D.R.; Coleman, J.S.; Guan, Y.; Knight, J.H.; Govorkova, E.A.; McClain-Moss, L.R.; Peiris, J.S.; Rehg, J.E.; Tuomanen, E.I.; et al. Responsiveness to a pandemic alert: Use of reverse genetics for rapid development of influenza vaccines. Lancet 2004, 363, 1099–1103. [Google Scholar] [CrossRef]
- Tripp, R.A.; Tompkins, S.M. Virus-vectored influenza virus vaccines. Viruses 2011, 6, 3055–3079. [Google Scholar] [CrossRef]
- Production of Pilot Lots of Inactivated Influenza Vaccines from Reassortants Derived from Avian Influenza Viruses. Interim Biosafety Risk Assessment. Available online: https://www.who.int/csr/resources/publications/influenza/WHO_CDS_CSR_RMD_2003_5/en/ (accessed on 18 April 2020).
- Verity, E.E.; Camuglia, S.; Agius, C.T.; Ong, C.; Shaw, R.; Barr, I.; Middleton, D.; Rockman, S. Rapid generation of pandemic influenza virus vaccine candidate strains using synthetic DNA. Influenza Other Respir. Viruses 2012, 6, 101–109. [Google Scholar] [CrossRef]
- Dormitzer, P.R.; Suphaphiphat, P.; Gibson, D.G.; Wentworth, D.E.; Stockwell, T.B.; Algire, M.A.; Alperovich, N.; Barro, M.; Brown, D.M.; Craig, S.; et al. Synthetic generation of influenza vaccine viruses for rapid response to pandemics. Sci. Transl. Med. 2013, 5, 185ra68. [Google Scholar] [CrossRef] [PubMed]
- Gambaryan, A.S.; Marinina, V.P.; Tuzikov, A.B.; Bovin, N.V.; Rudneva, I.A.; Sinitsyn, B.V.; Shilov, A.A.; Matrosovich, M.N. Effects of host-dependent glycosylation of hemagglutinin on receptor-binding properties on H1N1 human influenza A virus grown in MDCK cells and in embryonated eggs. Virology 1998, 247, 170–177. [Google Scholar] [CrossRef]
- Schild, G.C.; Oxford, J.S.; de Jong, J.C.; Webster, R.G. Evidence for host-cell selection of influenza virus antigenic variants. Nature 1983, 303, 706–709. [Google Scholar] [CrossRef] [PubMed]
- Australian Health Management Plan for Pandemic Influenza. Available online: https://www1.health.gov.au/internet/main/publishing.nsf/Content/519F9392797E2DDCCA257D47001B9948/$File/w-AHMPPI-2019.PDF (accessed on 18 April 2020).
- Pandemic Influenza Plan. 2017 Update. Available online: https://www.cdc.gov/flu/pandemic-resources/pdf/pan-flu-report-2017v2.pdf (accessed on 18 April 2020).
- Review of Australia’s Health Sector Response to Pandemic A(H1N1) 2009—Lessons Identified. Available online: https://www1.health.gov.au/internet/publications/publishing.nsf/Content/review-2011-l/$File/lessons%20identified-oct11.pdf (accessed on 24 January 2020).
- McBride, W.J.H.; Abhayaratna, W.P.; Barr, I.; Booy, R.; Carapetis, J.; Carson, S.; De Looze, F.; Ellis-Pegler, R.; Heron, L.; Karrasch, J.; et al. Efficacy of a trivalent influenza vaccine against seasonal strains and against 2009 pandemic H1N1: A randomized, placebo-controlled trial. Vaccine 2016, 34, 4991–4997. [Google Scholar] [CrossRef]
- Antigenic and Genetic Characteristics of Zoonotic Influenza Viruses and Candidate Vaccine Viruses Developed for Potential Use in Human Vaccines. Available online: https://www.who.int/influenza/vaccines/virus/characteristics_virus_vaccines/en/ (accessed on 18 January 2020).
- Cox, N.J.; Trock, S.C.; Burke, S.A. Pandemic preparedness and the Influenza Risk Assessment Tool (IRAT). Curr. Top. Microbiol. Immunol. 2014, 385, 119–136. [Google Scholar] [PubMed]
- Tool for Influenza Pandemic Risk Assessment (TIPRA). Available online: https://www.who.int/influenza/publications/TIPRA_manual_v1/en/ (accessed on 18 February 2020).
- Tregoning, J.S.; Russell, R.F.; Kinnear, E. Adjuvanted influenza vaccines. Hum. Vaccines Immunother. 2018, 14, 550–564. [Google Scholar] [CrossRef] [PubMed]
- Apostólico Jde, S.; Lunardelli, V.A.; Coirada, F.C.; Boscardin, S.B.; Santoro Rosa, D. Adjuvants: Classification, modus operandi, and licensing. J. Immunol. Res. 2016, 2016, 1459394. [Google Scholar] [CrossRef]
- Health Canada Approves Pandemic H1N1 Flu Vaccine for Canadians. Available online: https://www.eesti.ca/health-canada-approves-pandemic-h1n1-flu-vaccine-for-canadians/article25655 (accessed on 18 April 2020).
- Johansen, K.; Nicoll, A.; Ciancio, B.C.; Kramarz, P. Pandemic influenza A(H1N1) 2009 vaccines in the European Union. Eurosurveillance 2009, 14, 19361. [Google Scholar]
- Broadbent, A.J.; Subbarao, K. Influenza virus vaccines: Lessons from the 2009 H1N1 pandemic. Curr. Opin. Virol. 2011, 1, 254–262. [Google Scholar] [CrossRef]
- Lambert, L.C.; Fauci, A.S. Influenza vaccines for the future. N. Engl. J. Med. 2010, 363, 2036–2044. [Google Scholar] [CrossRef]
- Nolan, T.; McVernon, J.; Skeljo, M.; Richmond, P.; Wadia, U.; Lambert, S.; Nissen, M.; Marshall, H.; Booy, R.; Heron, L.; et al. Greenberg, Immunogenicity of a monovalent 2009 influenza A(H1N1) vaccine in infants and children: A randomized trial. JAMA 2010, 303, 37–46. [Google Scholar] [CrossRef] [PubMed]
- Greenberg, M.E.; Lai, M.H.; Hartel, G.F.; Wichems, C.H.; Gittleson, C.; Bennet, J.; Dawson, G.; Hu, W.; Leggio, C.; Washington, D.; et al. Response to a monovalent 2009 influenza A (H1N1) vaccine. N. Engl. J. Med. 2009, 361, 2405–2413. [Google Scholar] [CrossRef] [PubMed]
- Nolan, T.C.; Richmond, N.T.; Formica, K.; Hoschler, M.V.; Skeljo, T.; Stoney, J.; McVernon, G.; Hartel, D.C.; Sawlwin, J. Zambon, Safety and immunogenicity of a prototype adjuvanted inactivated split-virus influenza A (H5N1) vaccine in infants and children. Vaccine 2008, 26, 6383–6391. [Google Scholar] [CrossRef] [PubMed]
- Baz, M.; Luke, C.J.; Cheng, X.; Jin, H.; Subbarao, K. H5N1 vaccines in humans. Virus Res. 2013, 187, 78–98. [Google Scholar] [CrossRef]
- Dodd, C.N.; de Ridder, M.; Huang, W.T.; Weibel, D.; Giner-Soriano, M.; Perez-Vilar, S.; Diez-Domingo, J.; Svenson, L.W.; Mahmud, S.M.; Carleton, B.; et al. Incidence rates of narcolepsy diagnoses in Taiwan, Canada, and Europe: The use of statistical simulation to evaluate methods for the rapid assessment of potential safety issues on a population level in the SOMNIA study. PLoS ONE 2018, 13, e0204799. [Google Scholar] [CrossRef]
- Sarkanen, T.; Alakuijala, A.; Julkunen, I.; Partinen, M. Narcolepsy associated with pandemrix vaccine. Curr. Neurol. Neurosci. Rep. 2018, 18, 43. [Google Scholar] [CrossRef]
- Chada, K.E.; Forshee, R.; Golding, H.; Anderson, S.; Yang, H. A systematic review and meta-analysis of cross-reactivity of antibodies induced by oil-in-water emulsion adjuvanted influenza H5N1 virus monovalent vaccines. Vaccine 2017, 35, 3162–3170. [Google Scholar] [CrossRef]
- Sun, X.; Belser, J.A.; Pulit-Penaloza, J.A.; Creager, H.M.; Guo, Z.; Jefferson, S.N.; Liu, F.; York, I.A.; Stevens, J.; Maines, T.R.; et al. Stockpiled pre-pandemic H5N1 influenza virus vaccines with AS03 adjuvant provide cross-protection from H5N2 clade 2.3.4.4 virus challenge in ferrets. Virology 2017, 508, 164–169. [Google Scholar] [CrossRef]
- Oshansky, C.M.; Zhou, J.; Gao, Y.; Schweinle, J.E.; Biscardi, K.; DeBeauchamp, J.; Pavetto, C.; Wollish, A.; Webby, R.; BRITE Study Coordination Team; et al. Safety and immunogenicity of influenza A(H5N1) vaccine stored up to twelve years in the National Pre-Pandemic Influenza Vaccine Stockpile (NPIVS). Vaccine 2019, 37, 435–443. [Google Scholar] [CrossRef]
- Wood, J.M.; Weir, J.P. Standardisation of inactivated influenza vaccines—Learning from history. Influenza Other Respir. Viruses 2018, 12, 195–201. [Google Scholar] [CrossRef]
- WHO. Generic Protocol for the Calibration of Seasonal and Pandemic Influenza Antigen Working Reagents by WHO Essential Regulatory Laboratories. Sixty-Second Report of the Expert Committee on Biological Standardization, Annex 5; WHO Technical Report Series: Geneva, Switzerland, 2013; Volume 979. [Google Scholar]
- Nunnally, B.K.; Turula, V.E.; Sitrin, R.D. Vaccine Analysis: Strategies, Principles, and Control; Springer: Berlin/Heidelberg, Germany, 2015. [Google Scholar]
- McElhaney, J.E. Influenza vaccine responses in older adults. Ageing Res. Rev. 2011, 10, 379–388. [Google Scholar] [CrossRef] [PubMed]
- Xu, R.; Ekiert, D.C.; Krause, J.C.; Hai, R.; Crowe, J.E., Jr.; Wilson, I.A. Structural basis of preexisting immunity to the 2009 H1N1 pandemic influenza virus. Science 2010, 328, 357–360. [Google Scholar] [CrossRef] [PubMed]
- Kelly, H.; Peck, H.A.; Laurie, K.L.; Wu, P.; Nishiura, H.; Cowling, B.J. The age-specific cumulative incidence of infection with pandemic influenza H1N1 2009 was similar in various countries prior to vaccination. PLoS ONE 2011, 6, e21828. [Google Scholar] [CrossRef] [PubMed]
- Bernstein, D.I.; Edwards, K.M.; Dekker, C.L.; Belshe, R.; Talbot, H.K.; Graham, I.L.; Noah, D.L.; He, F.; Hill, H. Effects of adjuvants on the safety and immunogenicity of an avian influenza H5N1 vaccine in adults. J. Infect. Dis. 2008, 197, 667–675. [Google Scholar] [CrossRef] [PubMed]
- World Now at Start of 2009 Influenza Pandemic. Available online: https://www.who.int/mediacentre/news/statements/2009/h1n1_pandemic_phase6_20090611/en/ (accessed on 21 January 2020).
- 2009 H1N1 Pandemic Timeline. Available online: https://www.cdc.gov/flu/pandemic-resources/2009-pandemic-timeline.html (accessed on 1 February 2020).
- Swine Influenza. Available online: https://www.who.int/mediacentre/news/statements/2009/h1n1_20090425/en/ (accessed on 21 January 2020).
- Zhang, W. “SWITCH” from seasonal to pandemic vaccine production at the start of an influenza pandemic. In Proceedings of the 29th Meeting between WHO ERLs, CCs and Influenza Vaccine Manufacturers, London, UK, 16 January 2020. [Google Scholar]
- Ampofo, W.K.; Baylor, N.; Cobey, S.; Cox, N.J.; Daves, S.; Edwards, S.; Ferguson, N.; Grohmann, G.; Hay, A.; et al.; WHO Improving influenza vaccine virus selection: Report of a WHO informal consultation held at WHO headquarters, Geneva, Switzerland, 14–16 June 2010. Influenza Other Respir. Viruses 2012, 6, 142–152. [Google Scholar] [PubMed]
- Influenza—Global Influenza Surveillance and Response System (GISRS). Available online: https://www.who.int/influenza/gisrs_laboratory/en/ (accessed on 24 January 2020).
- One Health. Available online: https://www.who.int/features/qa/one-health/en/ (accessed on 18 February 2020).
- Nagoya Protocol and Public Health. Available online: https://www.who.int/influenza/pip/NagoyaProtocolandPH_EN_20Mar2018.pdf (accessed on 29 October 2019).
- Parties to the Nagoya Protocol. Available online: https://www.cbd.int/abs/nagoya-protocol/signatories/ (accessed on 24 January 2020).
- Pandemic Influenza Preparedness (PIP) Framework. Available online: https://www.who.int/influenza/pip/en/ (accessed on 28 January 2020).
- Berera, D.; Zambon, M. Antivirals in the 2009 pandemic—Lessons and implications for future strategies. Influenza Other Respir. Viruses 2013, 7 (Suppl. 3), 72–79. [Google Scholar] [CrossRef]
- Non-Pharmaceutical Public Health Measures for Mitigating the Risk and Impact of Epidemic and Pandemic Influenza. Available online: https://extranet.who.int/sph/docs/file/3848 (accessed on 1 February 2020).



{kind=link}
{kind=link}
| Issue | Situation in 2009 | Situation in 2019 |
|---|---|---|
| Vaccines | All vaccines (live and inactivated) prepared in embryonated hen’s eggs; only 7 manufacturers producing the majority of vaccines [11] | Expanded manufacturing platforms; embryonated hen’s eggs, mammalian cells, baculovirus, recombinant proteins; number and geographical dispersion of manufacturers increased [11] |
| Biocontainment | Strict biocontainment requirements for handling of wildtype virus under BSL-3 conditions by WHO | Biocontainment requirement determined locally by activity-based risk assessment [28] |
| CVV generated by reassorting limited to BSL-3 laboratories | Biocontainment level required for CVV generation by reassortment determined by activity-based risk assessment [28] | |
| CVV generated by reverse genetics in BSL-3 laboratories, by cloning from wildtype virus | CVV generated by reverse genetics in BSL-2 laboratories using synthetic genes with any potentially pathogenic determinants (e.g., polybasic site) easily removed. | |
| All CVVs must undergo safety testing (including ferret testing) to demonstrate attenuation. All safety testing must be completed prior to distribution | CVV must undergo safety testing to demonstrate attenuation. Risk assessment performed to determine testing requirements prior to distribution | |
| Results of safety test can be used for a further CVV with identical HA/NA sequence as previously generated and tested CVV | ||
| Manufacture can only be performed at BSL-3 with wildtype virus or BSL-2 with attenuated CVV once all safety testing is complete | Manufacture with wildtype or attenuated CVV assessed by activity-based risk assessment | |
| Host passage adaptations | A/California/4/2009 CVV unable to be rescued in time for a potential CVV | Potential for rescue of CVV in mammalian cells as well as eggs; egg adaption sites better understood |
| CVVs | Emerging pre-pandemic strains under constant surveillance by WHO and panel of pre-pandemic CVVs prepared | Increased surveillance by WHO to identify emerging pre-pandemic strains, the panel of pre-pandemic CVVs is constantly updated;IRAT [41] and TIPRA [42] risk assessment tools now available to help with pandemic vaccine stockpile prioritisation |
| Genetic sequence of influenza viruses limited with few specialist databases/tools and reliance on GenBank | Next generation sequencers and additional specialist influenza databases available (GISAID, IRD) and online analysis tools (Nextstrain, IRD, GISAID, FluSurver) | |
| Nagoya Protocol requires bilateral agreements if using virus from a signatory country | ||
| Adjuvants | MF59 and AS03 licensed, approved and used in pandemic vaccines | MF59, Alum (Al(OH)3 and AlPO4), AS03 and virosomes licensed, approved and used in seasonal vaccines. Adverse incidents to be considered before use in future pandemic |
| Assessment of vaccine potency and immunogenicity | SRID assay | Alternate potency assays under development |
| Immunogenicity determined by clinical trial | Pre-pandemic clinical trials of novel subtypes would give insight as to appropriate dose and vaccination regime; potential for use of government stockpiles with cross-clade vaccines | |
| ‘Switching’ Manufacturing | WHO declared a pandemic on June 11 2009, resulting in ‘switch’ in manufacturing from seasonal to pandemic vaccines | Despite three “WHO Switch meetings” it is still unclear as to how a switch from seasonal to pandemic vaccines would occur; potential for ‘switch’ to be initiated by individual governments |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rockman, S.; Laurie, K.; Barr, I. Pandemic Influenza Vaccines: What did We Learn from the 2009 Pandemic and are We Better Prepared Now? Vaccines 2020, 8, 211. https://doi.org/10.3390/vaccines8020211
Rockman S, Laurie K, Barr I. Pandemic Influenza Vaccines: What did We Learn from the 2009 Pandemic and are We Better Prepared Now? Vaccines. 2020; 8(2):211. https://doi.org/10.3390/vaccines8020211
Chicago/Turabian StyleRockman, Steven, Karen Laurie, and Ian Barr. 2020. "Pandemic Influenza Vaccines: What did We Learn from the 2009 Pandemic and are We Better Prepared Now?" Vaccines 8, no. 2: 211. https://doi.org/10.3390/vaccines8020211
APA StyleRockman, S., Laurie, K., & Barr, I. (2020). Pandemic Influenza Vaccines: What did We Learn from the 2009 Pandemic and are We Better Prepared Now? Vaccines, 8(2), 211. https://doi.org/10.3390/vaccines8020211