Differential Immune Transcriptome and Modulated Signalling Pathways in Rainbow Trout Infected with Viral Haemorrhagic Septicaemia Virus (VHSV) and Its Derivative Non-Virion (NV) Gene Deleted



Abstract
1. Introduction
1.1. Viral Haemorrhagic Septicaemia Viruses (VHSV)
1.2. Virus Dosages and Injection of Fingerling Rainbow Trout
1.3. RNA Extraction and cDNA Synthesis
1.4. Design of Oligo-Microarrays Enriched in Rainbow Trout Immune-Related Genes (Targeted Microarrays)
1.5. Hybridisation and Gene Expression Changes of Trout Transcripts to the Immune-Targeted Microarrays
1.6. Quantitative Estimation of Transcripts by Real-Time Quantitative PCR (RTqPCR)
1.7. Calculations used for Gene Set Enrichment Analysis (GSEA)
1.8. Ethics Statement
2. Results and Discussion
2.1. dNV- and wt-VHSV Dosages used for Microarray Analysis
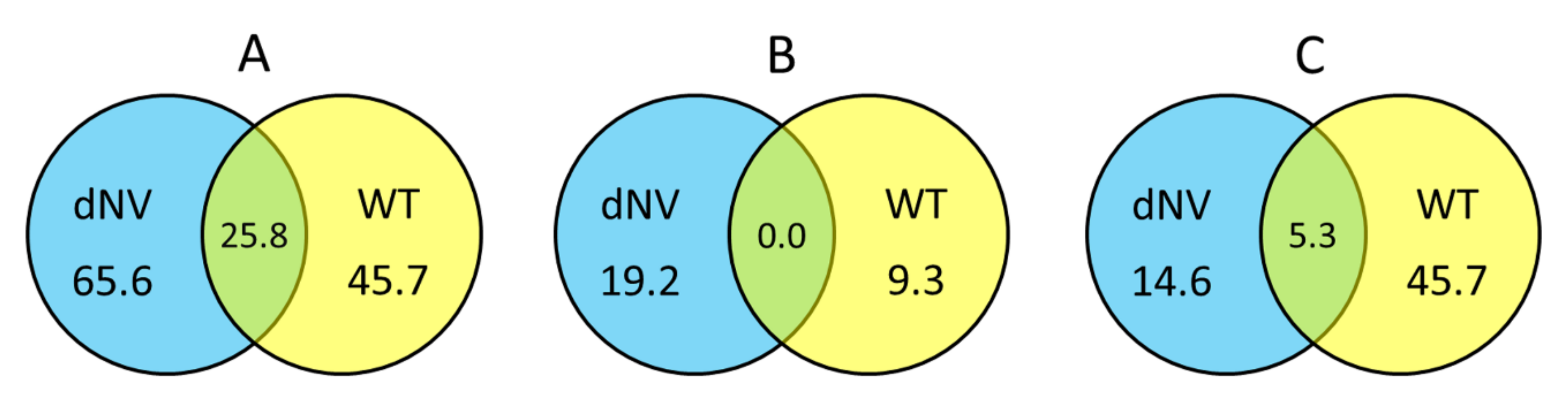
2.2. Overview of the Expression Profiles Obtained
2.3. dNV- and wt-VHSV Infection Effects on Trout Immune-Related Genes
2.3.1. Cytosolic Sensors
2.3.2. IFN System
2.3.3. TNF Superfamily and Caspases
2.3.4. Antigen Presentation
2.3.5. Cluster of Differentiation: B-Cell, T-Cell, and Cell-to-Cell Interactions
2.3.6. Cytokines: Chemokines and Interleukins
2.3.7. General Transcription Factors
2.3.8. Complement and VIG Genes
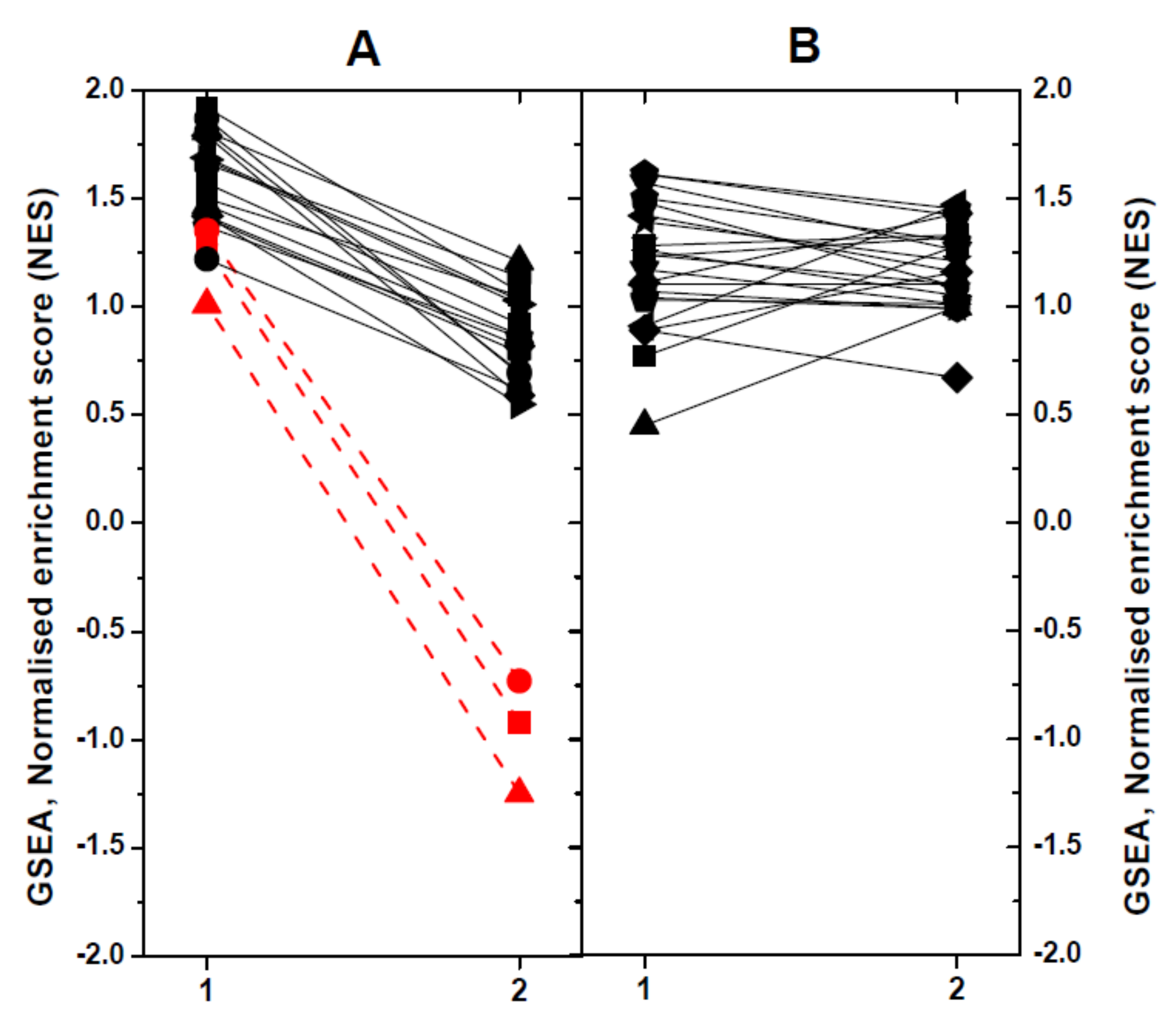
2.4. Modulated Pathways in dNV- and wt-VHSV-Injected Trout using GSEA
3. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Nishizawa, T.; Iida, H.; Takano, R.; Isshiki, T.; Nakajima, K.; Muroga, K. Genetic relatedness among Japanese, American and European isolates of viral hemorrhagic septicemia virus (VHSV) based on partial G and P genes. Dis. Aquat. Org. 2002, 48, 143–148. [Google Scholar] [CrossRef] [PubMed]
- Schutze, H.; Enzmann, P.J.; Kuchling, R.; Mundt, E.; Niemann, H.; Mettenleiter, T.C. Complete genomic sequence of the fish rhabdovirus infectious haematopoietic necrosis virus. J. Gen. Virol. 1995, 76, 2519–2527. [Google Scholar] [CrossRef] [PubMed]
- Schutze, H.; Mundt, E.; Mettenleiter, T.C. Complete genomic sequence of viral hemorrhagic septicemia virus, a fish rhabdovirus. Virus Genes 1999, 19, 59–65. [Google Scholar] [CrossRef] [PubMed]
- Skall, H.F.; Olesen, N.J.; Mellergaard, S. Viral haemorrhagic septicaemia virus in marine fish and its implications for fish farming - a review. J. Fish Dis. 2005, 28, 509–529. [Google Scholar] [CrossRef] [PubMed]
- Brudeseth, B.E.; Evensen, O. Occurrence of viral haemorrhagic septicaemia virus (VHSV) in wild marine fish species in the coastal regions of Norway. Dis. Aquat. Org. 2002, 52, 21–28. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Chinchilla, B.; Encinas, P.; Estepa, A.; Coll, J.M.; Gomez-Casado, E. Transcriptome analysis of rainbow trout in response to non-virion (NV) protein of viral haemorrhagic septicaemia virus (VHSV). Appl. Microbiol. Biotechnol. 2015, 99, 1827–1843. [Google Scholar] [CrossRef]
- Wahli, T.; Bergmann, S.M. Viral haemorrhagic septicaemia (VHS): Detection, distribution and combat. Cab. Rev: Persp. Agric. Vet. Sci. Nutr. Nat. Resour. 2011, 6, 1–10. [Google Scholar] [CrossRef]
- Kurath, G.; Leong, J.C. Characterization of infectious hematopoietic necrosis virus mRNA species reveals a nonvirion rhabdovirus protein. J. Virol. 1985, 53, 462–468. [Google Scholar] [CrossRef]
- Schutze, H.; Enzmann, P.J.; Mundt, E.; Mettenleiter, T.C. Identification of the non-virion (NV) protein of fish rhabdoviruses viral haemorrhagic septicaemia virus and infectious haematopoietic necrosis virus. J. Gen. Virol. 1996, 77, 1259–1263. [Google Scholar] [CrossRef]
- Einer-Jensen, K.; Ahrens, P.; Lorenzen, N. Parallel phylogenetic analyses using the N, G or Nv gene from a fixed group of VHSV isolates reveal the same overall genetic typing. Dis. Aquat. Org. 2005, 67, 39–45. [Google Scholar] [CrossRef][Green Version]
- Kurath, G.; Higman, K.H.; Björklund, H.V. Distribution and variation of NV genes in fish rhabdoviruses. J. Gen. Virol. 1997, 78, 113–117. [Google Scholar] [CrossRef] [PubMed]
- Biacchesi, S.; Thoulouze, M.I.; Bearzotti, M.; Yu, Y.X.; Bremont, M. Recovery of NV knockout infectious hematopoietic necrosis virus expressing foreign genes. J. Virol. 2000, 74, 11247–11253. [Google Scholar] [CrossRef] [PubMed]
- Biacchesi, S.; Lamoureux, A.; Merour, E.; Bernard, J.; Bremont, M. Limited interference at the early stage of infection between two recombinant novirhabdoviruses: Viral hemorrhagic septicemia virus and infectious hematopoietic necrosis virus. J. Virol. 2010, 84, 10038–10050. [Google Scholar] [CrossRef] [PubMed]
- Thoulouze, M.I.; Bouguyon, E.; Carpentier, C.; Bremont, M. Essential role of the NV protein of Novirhabdovirus for pathogenicity in rainbow trout. J. Virol. 2004, 78, 4098–4107. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.S.; Kim, K.H. Effects of NV gene knock-out recombinant viral hemorrhagic septicemia virus (VHSV) on Mx gene expression in Epithelioma papulosum cyprini (EPC) cells and olive flounder (Paralichthys olivaceus). Fish Shellfish Immunol. 2012, 32, 459–463. [Google Scholar] [CrossRef]
- Alonso, M.; Kim, C.H.; Johnson, M.C.; Pressley, M.; Leong, J.A. The NV gene of snakehead rhabdovirus (SHRV) is not required for pathogenesis, and a heterologous glycoprotein can be incorporated into the SHRV envelope. J. Virol. 2004, 78, 5875–5882. [Google Scholar] [CrossRef]
- Johnson, M.C.; Simon, B.E.; Kim, C.H.; Leong, J.A. Production of recombinant snakehead rhabdovirus: The NV protein is not required for viral replication. J. Virol. 2000, 74, 2343–2350. [Google Scholar] [CrossRef]
- Choi, M.K.; Moon, C.H.; Ko, M.S.; Lee, U.H.; Cho, W.J.; Cha, S.J.; Do, J.W.; Heo, G.J.; Jeong, S.G.; Hahm, Y.S.; et al. A nuclear localization of the infectious haematopoietic necrosis virus NV protein is necessary for optimal viral growth. PLoS ONE 2011, 6, e22362. [Google Scholar] [CrossRef]
- Ammayappan, A.; Vakharia, V.N. Nonvirion protein of novirhabdovirus suppresses apoptosis at the early stage of virus infection. J. Virol. 2011, 85, 8393–8402. [Google Scholar] [CrossRef]
- Cho, H.K.; Kim, J.; Moon, J.Y.; Nam, B.H.; Kim, Y.O.; Kim, W.J.; Park, J.Y.; An, C.M.; Cheong, J.; Kong, H.J. Microarray analysis of gene expression in olive flounder liver infected with viral haemorrhagic septicaemia virus (VHSV). Fish Shellfish Immunol. 2016, 49, 66–78. [Google Scholar] [CrossRef]
- Hwang, J.Y.; Kwon, M.G.; Seo, J.S.; Do, J.W.; Park, M.A.; Jung, S.H.; Ahn, S.J. Differentially expressed genes after viral haemorrhagic septicaemia virus infection in olive flounder (Paralichthys olivaceus). Vet. Microbiol. 2016, 193, 72–82. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.S.; Kim, K.H. Protection of olive flounder, Paralichthys olivaceus, against viral hemorrhagic septicemia virus (VHSV) by immunization with NV gene-knockout recombinant VHSV. Aquaculture 2011, 314, 39–43. [Google Scholar] [CrossRef]
- De Kinkelin, P.; LeBerre, M. Isolament d’un rhabdovirus pathogéne de la truite fario (Salmo trutta L.,1766). C R Acad. Sci. Hebd. Seances Acad. Sci. D 1977, 284, 101–104. (in French). [Google Scholar] [PubMed]
- Ballesteros, N.A.; Saint-Jean, S.S.; Encinas, P.A.; Perez-Prieto, S.I.; Coll, J.M. Oral immunization of rainbow trout to infectious pancreatic necrosis virus (IPNV) induces different immune gene expression profiles in head kidney and pyloric ceca. Fish Shellfish Immunol 2012, 33, 174–185. [Google Scholar] [CrossRef]
- Quesada-Garcia, A.; Encinas, P.; Valdehita, A.; Baumann, L.; Segner, H.; Coll, J.M.; Navas, J.M. Thyroid active agents T3 and PTU differentially affect immune gene transcripts in the head kidney of rainbow trout (Oncorhynchus mykiss). Aquat. Toxicol. 2016, 174, 159–168. [Google Scholar] [CrossRef]
- Salem, M.; Kenney, P.B.; Rexroad, C.E.; Yao, J. Development of a 37 k high-density oligonucleotide microarray: A new tool for functional genome research in rainbow trout. J. Fish Biol. 2008, 72, 2187–2206. [Google Scholar] [CrossRef]
- Gene Expression Omnibus. Available online: https://www.ncbi.nlm.nih.gov/geo/query/acc.cgi?acc=GSE37330 (accessed on 29 January 2020).
- Gene Expression Omnibus. Available online: http://www.ncbi.nlm.nih.gov/geo/query/acc.cgi?acc=GSE37797 (accessed on 29 January 2020).
- GSEA - Broad Institute. Available online: http://www.broad.mit.edu/GSEA (accessed on 29 January 2020).
- Subramanian, A.; Tamayo, P.; Mootha, V.K.; Mukherjee, S.; Ebert, B.L.; Gillette, M.A.; Paulovich, A.; Pomeroy, S.L.; Golub, T.R.; Lander, E.S.; et al. Gene set enrichment analysis: A knowledge-based approach for interpreting genome-wide expression profiles. Proc. Natl. Acad. Sci. USA 2005, 102, 15545–15550. [Google Scholar] [CrossRef]
- Subramanian, A.; Kuehn, H.; Gould, J.; Tamayo, P.; Mesirov, J.P. GSEA-P: A desktop application for Gene Set Enrichment Analysis. Bioinformatics 2007, 23, 3251–3253. [Google Scholar] [CrossRef]
- KEGG: Kyoto Encyclopedia of Genes and Genomes. Available online: http://www.kegg.jp (accessed on 29 January 2020).
- Glenney, G.W.; Wiens, G.D. Early diversification of the TNF superfamily in teleosts: Genomic characterization and expression analysis. J. Immunol. 2007, 178, 7955–7973. [Google Scholar] [CrossRef]
- Roth, W.; Wagenknecht, B.; Klumpp, A.; Naumann, U.; Hahne, M.; Tschopp, J.; Weller, M. APRIL, a new member of the tumor necrosis factor family, modulates death ligand-induced apoptosis. Cell Death Differ. 2001, 8, 403–410. [Google Scholar] [CrossRef]
- De Valck, D.; Jin, D.Y.; Heyninck, K.; Van de Craen, M.; Contreras, R.; Fiers, W.; Jeang, K.T.; Beyaert, R. The zinc finger protein A20 interacts with a novel anti-apoptotic protein which is cleaved by specific caspases. Oncogene 1999, 18, 4182–4190. [Google Scholar] [CrossRef] [PubMed]
- Kelly, C.; Shields, M.D.; Elborn, J.S.; Schock, B.C. A20 regulation of nuclear factor-kappaB: Perspectives for inflammatory lung disease. Am. J. Respir. Cell Mol. Biol. 2011, 44, 743–748. [Google Scholar] [CrossRef] [PubMed]
- Zelante, T.; Wong, A.Y.; Ping, T.J.; Chen, J.; Sumatoh, H.R.; Vigano, E.; Hong Bing, Y.; Lee, B.; Zolezzi, F.; Fric, J.; et al. CD103(+) Dendritic Cells Control Th17 Cell Function in the Lung. Cell Rep. 2015, 12, 1789–1801. [Google Scholar] [CrossRef] [PubMed]
- Chapoval, A.I.; Ni, J.; Lau, J.S.; Wilcox, R.A.; Flies, D.B.; Liu, D.; Dong, H.; Sica, G.L.; Zhu, G.; Tamada, K.; et al. B7-H3: A costimulatory molecule for T cell activation and IFN-gamma production. Nat. Immunol. 2001, 2, 269–274. [Google Scholar] [CrossRef]
- Dixon, B.; Shum, B.; Adams, E.J.; Magor, K.E.; Hedrick, R.P.; Muir, D.G.; Parham, P. CK-1, a putative chemokine of rainbow trout (Oncorhynchus mykiss). Immunol. Rev. 1998, 166, 341–348. [Google Scholar] [CrossRef]
- Montero, J.; Coll, J.; Sevilla, N.; Cuesta, A.; Bols, N.C.; Tafalla, C. Interleukin 8 and CK-6 chemokines specifically attract rainbow trout (Oncorhynchus mykiss) RTS11 monocyte-macrophage cells and have variable effects on their immune functions. Dev. Comp. Immunol. 2008, 32, 1374–1384. [Google Scholar] [CrossRef]
- Montero, J.; Ordas, M.C.; Alejo, A.; Gonzalez-Torres, L.; Sevilla, N.; Tafalla, C. CK12, a rainbow trout chemokine with lymphocyte chemo-attractant capacity associated to mucosal tissues. Mol. Immunol. 2011, 48, 1102–1113. [Google Scholar] [CrossRef]
- O’Farrell, C.; Vaghefi, N.; Cantonnet, M.; Buteau, B.; Boudinot, P.; Benmansour, A. Survey of transcript expression in rainbow trout leukocytes reveals a major contribution of interferon-responsive genes in the early response to a rhabdovirus infection. J. Virol. 2002, 76, 8040–8049. [Google Scholar] [CrossRef]
- Seo, J.; Yaneva, Y.R.; Cresswell, P. Viperin: A multifunctional, interferon-inducible protein that regulates virus replication. Cell Host Microbe 2011, 10, 534–539. [Google Scholar] [CrossRef]
- Boudinot, P.; Salhi, S.; Blanco, M.; Benmansour, A. Viral haemorrhagic septicaemia virus induces vig-2, a new interferon-responsive gene in rainbow trout. Fish Shellfish Immunol. 2001, 11, 383–397. [Google Scholar] [CrossRef]
- Najib, A.; Kim, M.S.; Choi, S.H.; Kang, Y.J.; Kim, K.H. Changes in microRNAs expression profile of olive flounder (Paralichthys olivaceus) in response to viral hemorrhagic septicemia virus (VHSV) infection. Fish Shellfish Immunol. 2016, 51, 384–391. [Google Scholar] [CrossRef] [PubMed]
- Laxman, N.; Rubin, C.J.; Mallmin, H.; Nilsson, O.; Pastinen, T.; Grundberg, E.; Kindmark, A. Global miRNA expression and correlation with mRNA levels in primary human bone cells. Rna 2015, 21, 1433–1443. [Google Scholar] [CrossRef] [PubMed]
- Estepa, A.; Coll, J. Innate Multigene Family Memories Are Implicated in the Viral-Survivor Zebrafish Phenotype. PLoS ONE 2015, 10, e0135483. [Google Scholar] [CrossRef] [PubMed]
- Encinas, P.; Garcia-Valtanen, P.; Chinchilla, B.; Gomez-Casado, E.; Estepa, A.; Coll, J. Identification of multipath genes differentially expressed in pathway-targeted microarrays in zebrafish infected and surviving spring viremia carp virus (SVCV) suggest preventive drug candidates. PLoS ONE 2013, 8, e73553. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| (a) Antigen processing and presentation | (q) Th1 and Th2 cell differentiation | |
| (b) Apoptosis | (r) Th17 cell differentiation | |
| (c) B-cell receptor signalling | (s) TLR (Toll-like receptor) signalling | |
| (d) Chemokine signalling | (t) TNF (Tumour necrosis factor) signalling | |
| (e) Complement and coagulation cascades | (u) Cytokine-cytokine receptor interaction | |
| (f) ErbB (Erb-B2 Receptor Tyrosine Kinase 2) signalling | ||
| (v) PI3-Akt (phosphatidylinositol 3-kinase and Protein kinase B ) signalling | ||
| (w) FoxO (Forkhead box O)signalling | ||
| (g) IL-17 signalling | ||
| (x) Natural killer cell-mediated cytotoxicity | ||
| (h) Jak–Stat signalling (Janus kinase-Signal transducer and activator of transcription) | (y) cAMP (Cyclic adenosine 3′,5′-monophosphate) signalling | |
| (i) MAPK (Mitogen-Activated Protein Kinase) signalling | (z) AMPK (AMP-activated protein kinase) signalling | |
| (j) Necroptosis signalling | (aa) Cell adhesion molecules | |
| (k) NF-kappa B signalling (Nuclear Factor kappa-light-chain-enhancer of activated B cells) | (bb) Proteasome | |
| (cc) Wnt (wingless-type MMTV integration site family) signalling | ||
| (l) NOD-like (nucleotide-binding oligomerization domain-like) receptor signalling | (dd) General transcription factors | |
| (ee) Hematopoietic cell lineage | ||
| (ff) TGFb (Transforming growth factor beta) signalling | ||
| (m) p53 signalling | (gg) mTOR (mammalian target of rapamycin) signalling | |
| (n) Protein processing in endoplasmic reticulum | ||
| (o) RIG-I-like receptor signalling | ||
| (p) T-cell receptor signalling |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chinchilla, B.; Encinas, P.; Coll, J.M.; Gomez-Casado, E. Differential Immune Transcriptome and Modulated Signalling Pathways in Rainbow Trout Infected with Viral Haemorrhagic Septicaemia Virus (VHSV) and Its Derivative Non-Virion (NV) Gene Deleted. Vaccines 2020, 8, 58. https://doi.org/10.3390/vaccines8010058
Chinchilla B, Encinas P, Coll JM, Gomez-Casado E. Differential Immune Transcriptome and Modulated Signalling Pathways in Rainbow Trout Infected with Viral Haemorrhagic Septicaemia Virus (VHSV) and Its Derivative Non-Virion (NV) Gene Deleted. Vaccines. 2020; 8(1):58. https://doi.org/10.3390/vaccines8010058
Chicago/Turabian StyleChinchilla, Blanca, Paloma Encinas, Julio M. Coll, and Eduardo Gomez-Casado. 2020. "Differential Immune Transcriptome and Modulated Signalling Pathways in Rainbow Trout Infected with Viral Haemorrhagic Septicaemia Virus (VHSV) and Its Derivative Non-Virion (NV) Gene Deleted" Vaccines 8, no. 1: 58. https://doi.org/10.3390/vaccines8010058
APA StyleChinchilla, B., Encinas, P., Coll, J. M., & Gomez-Casado, E. (2020). Differential Immune Transcriptome and Modulated Signalling Pathways in Rainbow Trout Infected with Viral Haemorrhagic Septicaemia Virus (VHSV) and Its Derivative Non-Virion (NV) Gene Deleted. Vaccines, 8(1), 58. https://doi.org/10.3390/vaccines8010058