Recent Progress on the Versatility of Virus-Like Particles

 , and
, and
Abstract
1. Introduction
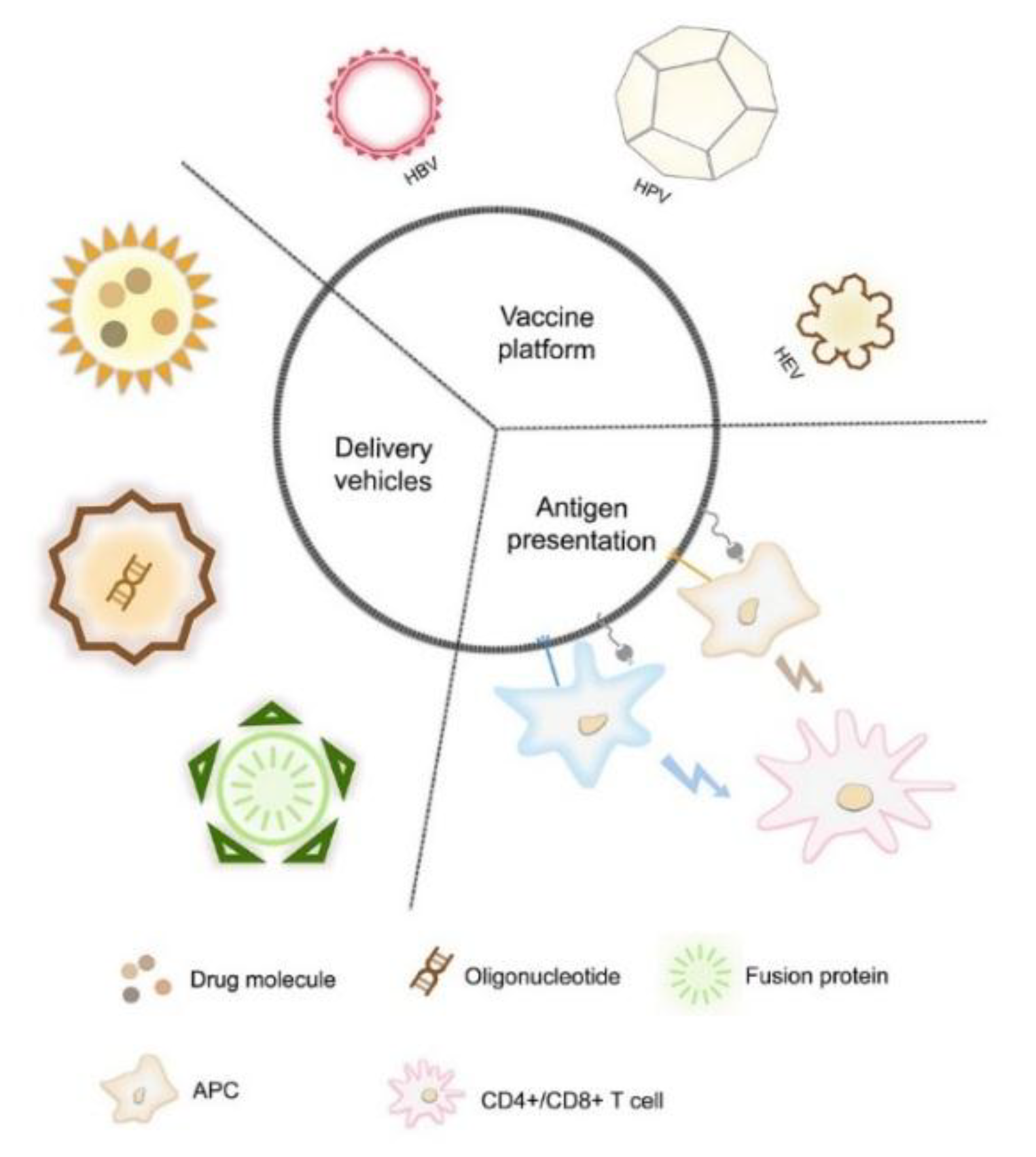
2. VLP Technology Applied in Basic Research
2.1. VLP Immunogens
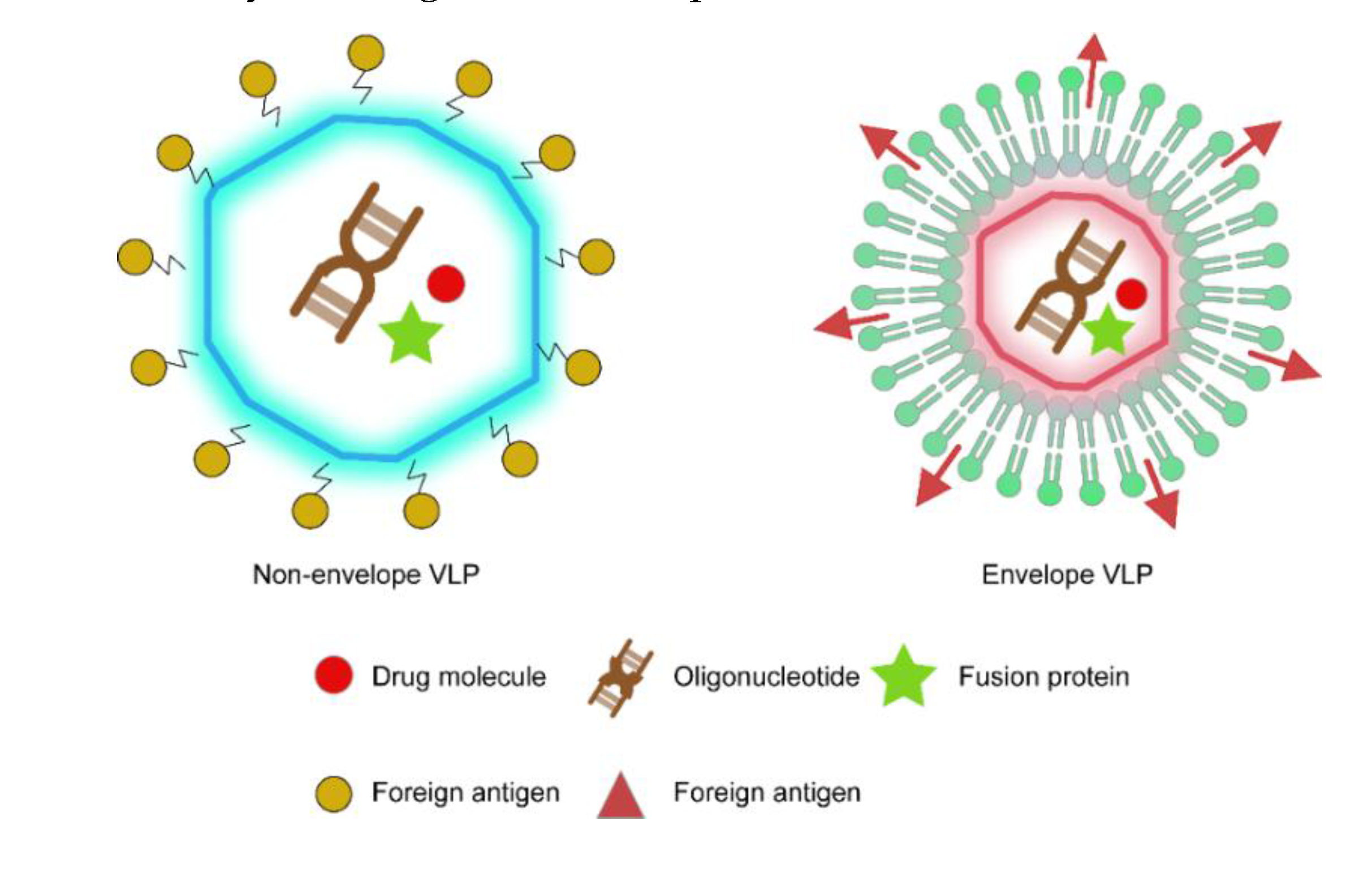
2.2. Epitope-Displaying VLP Nanocarriers
2.3. VLP Delivery Platform
3. VLP-Based Vaccines
3.1. Recombinant HBV Vaccine
3.2. HPV VLP Vaccines
3.3. Recombinant HEV Vaccine
4. Research Progress on VLP-Based Vaccines in Development
5. Other VLP-Related Information
5.1. The Administration Route of VLP-Based Vaccine
5.2. VLP-Based Vaccine versus Virosomes-Based Vaccine
6. Discussion and Conclusion
Author Contributions
Funding
Conflicts of Interest
References
- Standaert, B.; Rappuoli, R. How comprehensive can we be in the economic assessment of vaccines? J. Mark. Access Health Policy 2017, 5, 1336044. [Google Scholar] [CrossRef]
- Vetter, V.; Denizer, G.; Friedland, L.R. Understanding modern-day vaccines: What you need to know. Ann. Med. 2018, 50, 110–120. [Google Scholar] [CrossRef]
- Tretyakova, I.; Plante, K.S.; Rossi, S.L. Venezuelan equine encephalitis vaccine with rearranged genome resists reversion and protects non-human primates from viremia after aerosol challenge. Vaccine 2020. [Google Scholar] [CrossRef]
- Tebbens, R.J.; Pallansch, M.A.; Kew, O.M. Risks of paralytic disease due to wild or vaccine-derived poliovirus after eradication. Risk Anal. 2006, 26, 1471–1505. [Google Scholar] [CrossRef]
- Metz, S.W.; Thomas, A.; White, L. Dengue virus-like particles mimic the antigenic properties of the infectious dengue virus envelope. Virol. J. 2018, 15, 60. [Google Scholar] [CrossRef]
- Deschuyteneer, M.; Elouahabi, A.; Plainchamp, D. Molecular and structural characterization of the L1 virus-like particles that are used as vaccine antigens in Cervarix, the AS04-adjuvanted HPV-16 and -18 cervical cancer vaccine. Hum. Vaccines 2010, 6, 407–419. [Google Scholar] [CrossRef] [PubMed]
- Zabel, F.; Kundig, T.M.; Bachmann, M.F. Virus-induced humoral immunity: On how B cell responses are initiated. Curr. Opin. Virol. 2013, 3, 357–362. [Google Scholar] [CrossRef] [PubMed]
- Phelps, D.K.; Speelman, B.; Post, C.B. Theoretical studies of viral capsid proteins. Curr. Opin. Struct. Biol. 2000, 10, 170–173. [Google Scholar] [CrossRef]
- Sasagawa, T.; Pushko, P.; Steers, G. Synthesis and assembly of virus-like particles of human papillomaviruses type 6 and type 16 in fission yeast Schizosaccharomyces pombe. Virology 1995, 206, 126–135. [Google Scholar] [CrossRef]
- Chen, B.J.; Leser, G.P.; Morita, E. Influenza virus hemagglutinin and neuraminidase, but not the matrix protein, are required for assembly and budding of plasmid-derived virus-like particles. J. Virol. 2007, 81, 7111–7123. [Google Scholar] [CrossRef] [PubMed]
- Haynes, J.R. Influenza virus-like particle vaccines. Expert Rev. Vaccines 2009, 8, 435–445. [Google Scholar] [CrossRef]
- Mohsen, M.O.; Gomes, A.C.; Vogel, M. Interaction of Viral Capsid-Derived Virus-Like Particles (VLPs) with the Innate Immune System. Vaccines 2018, 6, 37. [Google Scholar] [CrossRef] [PubMed]
- Cinamon, G.; Zachariah, M.A.; Lam, O.M. Follicular shuttling of marginal zone B cells facilitates antigen transport. Nat. Immunol. 2008, 9, 54–62. [Google Scholar] [CrossRef] [PubMed]
- Zabel, F.; Mohanan, D.; Bessa, J. Viral particles drive rapid differentiation of memory B cells into secondary plasma cells producing increased levels of antibodies. J. Immunol. 2014, 192, 5499–5508. [Google Scholar] [CrossRef] [PubMed]
- Cimica, V.; Galarza, J.M. Adjuvant formulations for virus-like particle (VLP) based vaccines. Clin. Immunol. 2017, 183, 99–108. [Google Scholar] [CrossRef] [PubMed]
- Hong, S.; Zhang, Z.; Liu, H. B Cells Are the Dominant Antigen-Presenting Cells that Activate Naive CD4 (+) T Cells upon Immunization with a Virus-Derived Nanoparticle Antigen. Immunity 2018, 49, 695–708. [Google Scholar] [CrossRef]
- Storni, T.; Ruedl, C.; Schwarz, K. Nonmethylated CG Motifs Packaged into Virus-Like Particles Induce Protective Cytotoxic T Cell Responses in the Absence of Systemic Side Effects. J. Immunol. 2004, 172, 1777–1785. [Google Scholar] [CrossRef]
- Gatto, D.; Ruedl, C.; Odermatt, B. Rapid Response of Marginal Zone B Cells to Viral Particles. J. Immunol. 2004, 173, 4308–4316. [Google Scholar] [CrossRef]
- Peacey, M.; Wilson, S.; Baird, M.A. Versatile RHDV virus-like particles: Incorporation of antigens by genetic modification and chemical conjugation. Biotechnol. Bioeng. 2007, 98, 968–977. [Google Scholar] [CrossRef]
- Billaud, J.N.; Peterson, D.; Barr, M. Combinatorial approach to hepadnavirus-like particle vaccine design. J. Virol. 2005, 79, 13656–13666. [Google Scholar] [CrossRef]
- Yong, C.Y.; Yeap, S.K.; Goh, Z.H. Induction of humoral and cell-mediated immune responses by hepatitis B virus epitope displayed on the virus-like particles of prawn nodavirus. Appl. Environ. Microbiol. 2015, 81, 882–889. [Google Scholar] [CrossRef] [PubMed]
- Biabanikhankahdani, R.; Alitheen, N.B.M.; Ho, K.L. pH-responsive Virus-like Nanoparticles with Enhanced Tumour-targeting Ligands for Cancer Drug Delivery. Sci. Rep. 2016, 6, 37891. [Google Scholar] [CrossRef] [PubMed]
- Goldinger, S.M.; Dummer, R.; Baumgaertner, P. Nano-particle vaccination combined with TLR-7 and -9 ligands triggers memory and effector CD8(+) T-cell responses in melanoma patients. Eur. Immunol. 2012, 42, 3049–3061. [Google Scholar] [CrossRef] [PubMed]
- Frietze, K.M.; Peabody, D.S.; Chackerian, B. Engineering virus-like particles as vaccine platforms. Curr. Opin. Virol. 2016, 18, 44–49. [Google Scholar] [CrossRef] [PubMed]
- Pyrski, M.; Rugowska, A.; Wierzbinski, K.R. HBcAg produced in transgenic tobacco triggers Th1 and Th2 response when intramuscularly delivered. Vaccine 2017, 35, 5714–5721. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.T.S.; Zhou, S. Effect of surface antigen density of hepatitis B virus core protein virus-like particles on antibody response level. J. Microbiol. 2019, 1–12. [Google Scholar] [CrossRef]
- Ramirez, A.; Morris, S.; Maucourant, S. A virus-like particle vaccine candidate for influenza A virus based on multiple conserved antigens presented on hepatitis B tandem core particles. Vaccine 2018, 36, 873–880. [Google Scholar] [CrossRef]
- Janitzek, C.M.P.J.; Thrane, S. A proof-of-concept study for the design of a VLP-based combinatorial HPV and placental malaria vaccine. Sci. Rep. 2019, 9, 5260. [Google Scholar] [CrossRef]
- Steinmetz, N.F. Viral nanoparticles as platforms for next-generation therapeutics and imaging devices. Nanomedicine 2010, 6, 634–641. [Google Scholar] [CrossRef]
- Seow, Y.; Wood, M.J. Biological gene delivery vehicles: Beyond viral vectors. Mol. Ther. 2009, 17, 767–777. [Google Scholar] [CrossRef]
- Brown, W.L.; Mastico, R.A.; Wu, M. RNA bacteriophage capsid-mediated drug delivery and epitope presentation. Intervirology 2002, 45, 371–380. [Google Scholar] [CrossRef] [PubMed]
- Kovacs, E.W.; Hooker, J.M.; Romanini, D.W. Dual-surface-modified bacteriophage MS2 as an ideal scaffold for a viral capsid-based drug delivery system. Bioconjug. Chem. 2007, 18, 1140–1147. [Google Scholar] [CrossRef] [PubMed]
- Destito, G.; Yeh, R.; Rae, C.S. Folic acid-mediated targeting of cowpea mosaic virus particles to tumor cells. Chem. Biol. 2007, 14, 1152–1162. [Google Scholar] [CrossRef] [PubMed]
- Steinmetz, N.F.; Hong, V.; Spoerke, E.D. Buckyballs meet viral nanoparticles: Candidates for biomedicine. J. Am. Chem. Soc. 2009, 131, 17093–17095. [Google Scholar] [CrossRef]
- Wold, W.S.; Toth, K. Adenovirus vectors for gene therapy, vaccination and cancer gene therapy. Curr. Gene Ther. 2013, 13, 421–433. [Google Scholar] [CrossRef]
- Zochowska, M.; Piguet, A.C.; Jemielity, J. Virus-like particle-mediated intracellular delivery of mRNA cap analog with in vivo activity against hepatocellular carcinoma. Nanomedicine 2015, 11, 67–76. [Google Scholar] [CrossRef]
- Zochowska, M.; Paca, A.; Schoehn, G. Adenovirus dodecahedron, as a drug delivery vector. PLoS ONE 2009, 4, e5569. [Google Scholar] [CrossRef]
- Chroboczek, J.; Szurgot, I.; Szolajska, E. Virus-like particles as vaccine. Acta Biochim. Pol. 2014, 61, 531–539. [Google Scholar] [CrossRef]
- Gerety, R.J.; Tabor, E. Newly licensed hepatitis B vaccine. Known safety and unknown risks. JAMA 1983, 249, 745–746. [Google Scholar] [CrossRef]
- Szmuness, W.; Stevens, C.E.; Harley, E.J. Hepatitis B vaccine: Demonstration of efficacy in a controlled clinical trial in a high-risk population in the United States. N. Engl. J. Med. 1980, 303, 833–841. [Google Scholar] [CrossRef]
- Hieu, N.T.; Kim, K.H.; Janowicz, Z. Comparative efficacy, safety and immunogenicity of Hepavax-Gene and Engerix-B, recombinant hepatitis B vaccines, in infants born to HBsAg and HBeAg positive mothers in Vietnam: An assessment at 2 years. Vaccine 2002, 20, 1803–1808. [Google Scholar] [CrossRef]
- Stephenne, J. Production in yeast versus mammalian cells of the first recombinant DNA human vaccine and its proved safety, efficacy, and economy: Hepatitis B vaccine. Adv. Biotechnol. Process. 1990, 14, 279–299. [Google Scholar]
- Lacson, E.; Teng, M.; Ong, J. Antibody response to Engerix-B and Recombivax-HB hepatitis B vaccination in end-stage renal disease. Hemodial. Int. 2005, 9, 367–375. [Google Scholar] [CrossRef]
- Stevens, C.E.; Taylor, P.E.; Tong, M.J. Yeast-recombinant hepatitis B vaccine. Efficacy with hepatitis B immune globulin in prevention of perinatal hepatitis B virus transmission. JAMA 1987, 257, 2612–2616. [Google Scholar] [CrossRef] [PubMed]
- Zuckerman, J.N.; Zuckerman, A.J.; Symington, I. Evaluation of a new hepatitis B triple-antigen vaccine in inadequate responders to current vaccines. Hepatology 2001, 34, 798–802. [Google Scholar] [CrossRef] [PubMed]
- Shouval, D.; Ilan, Y.; Adler, R. Improved immunogenicity in mice of a mammalian cell-derived recombinant hepatitis B vaccine containing pre-S1 and pre-S2 antigens as compared with conventional yeast-derived vaccines. Vaccine 1994, 12, 1453–1459. [Google Scholar] [CrossRef]
- Qawasmi, M.; Samuh, M.; Glebe, D. Age-dependent decrease of anti-HBs titers and effect of booster doses using 2 different vaccines in Palestinian children vaccinated in early childhood. Hum. Vaccines Immunother. 2015, 11, 1717–1724. [Google Scholar] [CrossRef] [PubMed]
- Raz, R.; Koren, R.; Bass, D. Safety and immunogenicity of a new mammalian cell-derived recombinant hepatitis B vaccine containing Pre-S1 and Pre-S2 antigens in adults. Isr. Med. Assoc. J. 2001, 3, 328–332. [Google Scholar]
- Jama, N.J. A Two-Dose Hepatitis B Vaccine for Adults (Heplisav-B). JAMA 2018, 319, 822–823. [Google Scholar] [CrossRef]
- Halperin, S.A.; Ward, B.; Cooper, C. Comparison of safety and immunogenicity of two doses of investigational hepatitis B virus surface antigen co-administered with an immunostimulatory phosphorothioate oligodeoxyribonucleotide and three doses of a licensed hepatitis B vaccine in healthy adults 18–55 years of age. Vaccine 2012, 30, 2556–2563. [Google Scholar] [CrossRef]
- Tong, N.K.; Beran, J.; Kee, S.A. Immunogenicity and safety of an adjuvanted hepatitis B vaccine in pre-hemodialysis and hemodialysis patients. Kidney Int. 2005, 68, 2298–2303. [Google Scholar] [CrossRef] [PubMed]
- Rebedea, I.; Diaconescu, I.G.; Bach, D. Comparison of thiomersal-free and thiomersal-containing formulations of a recombinant hepatitis B vaccine (Hepavax-Gene) in healthy adults. Vaccine 2006, 24, 5320–5326. [Google Scholar] [CrossRef] [PubMed]
- Garland, S.M.; Hernandez-Avila, M.; Wheeler, C.M. Quadrivalent vaccine against human papillomavirus to prevent anogenital diseases. N. Engl. J. Med. 2007, 356, 1928–1943. [Google Scholar] [CrossRef] [PubMed]
- Quadrivalent vaccine against human papillomavirus to prevent high-grade cervical lesions. N. Engl. J. Med. 2007, 356, 1915–1927. [CrossRef] [PubMed]
- Paavonen, J.; Jenkins, D.; Bosch, F.X. Efficacy of a prophylactic adjuvanted bivalent L1 virus-like-particle vaccine against infection with human papillomavirus types 16 and 18 in young women: An interim analysis of a phase III double-blind, randomised controlled trial. Lancet 2007, 369, 2161–2170. [Google Scholar] [CrossRef]
- Herrero, R.; Hildesheim, A.; Rodriguez, A.C. Rationale and design of a community-based double-blind randomized clinical trial of an HPV 16 and 18 vaccine in Guanacaste, Costa Rica. Vaccine 2008, 26, 4795–4808. [Google Scholar] [CrossRef]
- Iversen, O.E.; Miranda, M.J.; Ulied, A. Immunogenicity of the 9-Valent HPV Vaccine Using 2-Dose Regimens in Girls and Boys vs a 3-Dose Regimen in Women. JAMA 2016, 316, 2411–2421. [Google Scholar] [CrossRef]
- Gu, Y.; Wei, M.; Wang, D. Characterization of an Escherichia coli-derived human papillomavirus type 16 and 18 bivalent vaccine. Vaccine 2017, 35, 4637–4645. [Google Scholar] [CrossRef]
- Qiao, Y.L.; Wu, T.; Li, R.C. Efficacy, safety, and immunogenicity of an Escherichia coli-produced bivalent human papillomavirus vaccine: An interim analysis of a randomized clinical trial. J. Natl. Cancer Inst. 2020, 112, 145–153. [Google Scholar] [CrossRef]
- Wei, M.; Wang, D.; Li, Z. N-terminal truncations on L1 proteins of human papillomaviruses promote their soluble expression in Escherichia coli and self-assembly in vitro. Emerg. Microbes Infect. 2018, 7, 160. [Google Scholar] [CrossRef]
- Li, S.W.; Zhang, J.; Li, Y.M. A bacterially expressed particulate hepatitis E vaccine: Antigenicity, immunogenicity and protectivity on primates. Vaccine 2005, 23, 2893–2901. [Google Scholar] [CrossRef] [PubMed]
- Zhu, F.C.; Zhang, J.; Zhang, X.F. Efficacy and safety of a recombinant hepatitis E vaccine in healthy adults: A large-scale, randomised, double-blind placebo-controlled, phase 3 trial. Lancet 2010, 376, 895–902. [Google Scholar] [CrossRef]
- Shrestha, M.P.; Scott, R.M.; Joshi, D.M. Safety and efficacy of a recombinant hepatitis E vaccine. N. Engl. J. Med. 2007, 356, 895–903. [Google Scholar] [CrossRef] [PubMed]
- Cao, Y.F.; Tao, H.; Hu, Y.M. A phase 1 randomized open-label clinical study to evaluate the safety and tolerability of a novel recombinant hepatitis E vaccine. Vaccine 2017, 35, 5073–5080. [Google Scholar] [CrossRef] [PubMed]
- Shinde, V.; Fries, L.; Wu, Y. Improved Titers against Influenza Drift Variants with a Nanoparticle Vaccine. N. Engl. J. Med. 2018, 378, 2346–2348. [Google Scholar] [CrossRef] [PubMed]
- Portnoff, A.D.; Patel, N.; Massare, M.J. Influenza Hemagglutinin Nanoparticle Vaccine Elicits Broadly Neutralizing Antibodies against Structurally Distinct Domains of H3N2 HA. Vaccines 2020, 8, 99. [Google Scholar] [CrossRef] [PubMed]
- Low, J.G.; Lee, L.S.; Ooi, E.E. Safety and immunogenicity of a virus-like particle pandemic influenza A (H1N1) 2009 vaccine: Results from a double-blinded, randomized Phase I clinical trial in healthy Asian volunteers. Vaccine 2014, 32, 5041–5048. [Google Scholar] [CrossRef][Green Version]
- Pillet, S.; Aubin, E.; Trepanier, S. Humoral and cell-mediated immune responses to H5N1 plant-made virus-like particle vaccine are differentially impacted by alum and GLA-SE adjuvants in a Phase 2 clinical trial. NPJ Vaccines 2018, 3, 3. [Google Scholar] [CrossRef]
- von Seidlein, L.; Hanboonkunupakarn, B.; Jittamala, P. Combining antimalarial drugs and vaccine for malaria elimination campaigns: A randomized safety and immunogenicity trial of RTS,S/AS01 administered with dihydroartemisinin, piperaquine, and primaquine in healthy Thai adult volunteers. Hum. Vaccines Immunother. 2020, 16, 33–41. [Google Scholar] [CrossRef]
- Collins, K.A.; Snaith, R.; Cottingham, M.G. Enhancing protective immunity to malaria with a highly immunogenic virus-like particle vaccine. Sci. Rep. 2017, 7, 46621. [Google Scholar] [CrossRef]
- Blazevic, V.; Lappalainen, S.; Nurminen, K. Norovirus VLPs and rotavirus VP6 protein as combined vaccine for childhood gastroenteritis. Vaccine 2011, 29, 8126–8133. [Google Scholar] [CrossRef]
- Malm, M.; Heinimaki, S.; Vesikari, T. Rotavirus capsid VP6 tubular and spherical nanostructures act as local adjuvants when co-delivered with norovirus VLPs. Clin. Exp. Immunol. 2017, 189, 331–341. [Google Scholar] [CrossRef] [PubMed]
- Groome, M.J.; Koen, A.; Fix, A. Safety and immunogenicity of a parenteral P2-VP8-P [8] subunit rotavirus vaccine in toddlers and infants in South Africa: A randomised, double-blind, placebo-controlled trial. Lancet Infect. Dis. 2017, 17, 843–853. [Google Scholar] [CrossRef]
- Coste, A.; Sirard, J.C.; Johansen, K. Nasal immunization of mice with virus-like particles protects offspring against rotavirus diarrhea. J. Virol. 2000, 74, 8966–8971. [Google Scholar] [CrossRef] [PubMed]
- Leroux-Roels, G.; Cramer, J.P.; Mendelman, P.M. Safety and Immunogenicity of Different Formulations of Norovirus Vaccine Candidate in Healthy Adults: A Randomized, Controlled, Double-Blind Clinical Trial. J. Infect. Dis. 2018, 217, 597–607. [Google Scholar] [CrossRef]
- Kim, H.J.; Son, H.S.; Lee, S.W. Efficient expression of enterovirus 71 based on virus-like particles vaccine. PLoS ONE 2019, 14, e0210477. [Google Scholar] [CrossRef]
- Zhang, W.; Dai, W.; Zhang, C. A virus-like particle-based tetravalent vaccine for hand, foot, and mouth disease elicits broad and balanced protective immunity. Emerg. Microbes Infect. 2018, 7, 94. [Google Scholar] [CrossRef]
- Fries, L.; Shinde, V.; Stoddard, J. Immunogenicity and safety of a respiratory syncytial virus fusion protein (RSV F) nanoparticle vaccine in older adults. Immun. Ageing 2017, 14, 8. [Google Scholar] [CrossRef]
- Glenn, G.M.; Smith, G.; Fries, L. Safety and immunogenicity of a Sf9 insect cell-derived respiratory syncytial virus fusion protein nanoparticle vaccine. Vaccine 2013, 31, 524–532. [Google Scholar] [CrossRef]
- Yusibov, V.; Hooper, D.C.; Spitsin, S.V. Expression in plants and immunogenicity of plant virus-based experimental rabies vaccine. Vaccine 2002, 20, 3155–3164. [Google Scholar] [CrossRef]
- Karayiannis, P. Hepatitis B virus: Virology, molecular biology, life cycle and intrahepatic spread. Hepatol. Int. 2017, 11, 500–508. [Google Scholar] [CrossRef] [PubMed]
- Splawn, L.M.; Bailey, C.A.; Medina, J.P. Heplisav-B vaccination for the prevention of hepatitis B virus infection in adults in the United States. Drugs Today 2018, 54, 399–405. [Google Scholar] [CrossRef] [PubMed]
- Netter, H.J.; Chang, S.F.; Bruns, M.J.F.V. Host-range and pathogenicity of hepatitis B viruses. Future Med. 2008, 3, 83–94. [Google Scholar] [CrossRef]
- Zhao, Q.; Li, S.; Yu, H. Virus-like particle-based human vaccines: Quality assessment based on structural and functional properties. Trends Biotechnol. 2013, 31, 654–663. [Google Scholar] [CrossRef] [PubMed]
- Zhang, T.Y.; Guo, X.R.; Wu, Y.T.; Kang, X.-Z.; Zheng, Q.-B.; Qi, R.-Y.; Chen, B.-B.; Lan, Y.; Wei, M.; Wang, S.-J; et al. A unique B cell epitope-based particulate vaccine shows effective suppression of hepatitis B surface antigen in mice. Gut 2019, 69, 343–354. [Google Scholar] [CrossRef] [PubMed]
- Wei, M.; Wang, D.; Li, Z. N-terminal truncations on L1 proteins of human papillomaviruses promote their soluble expression in Escherichia coli and self-assembly in vitro. Emerg. Microbes Infect. 2018, 7, 160. [Google Scholar] [CrossRef]
- Kondo, K. Development of an HPV vaccine--remaining issues and perspective. Nihon Rinsho 2009, 67, 62–68. [Google Scholar]
- Mohsen, M.O.; Zha, L.; Cabral-Miranda, G. Major findings and recent advances in virus-like particle (VLP)-based vaccines. Semin. Immunol. 2017, 34, 123–132. [Google Scholar] [CrossRef]
- Chabeda, A.; van Zyl, A.R.; Rybicki, E.P. Substitution of Human Papillomavirus Type 16 L2 Neutralizing Epitopes Into L1 Surface Loops: The Effect on Virus-Like Particle Assembly and Immunogenicity. Front. Plant Sci. 2019, 10, 779. [Google Scholar] [CrossRef]
- Li, Z.; Wang, D.; Gu, Y. Crystal Structures of Two Immune Complexes Identify Determinants for Viral Infectivity and Type-Specific Neutralization of Human Papillomavirus. MBio 2017, 8. [Google Scholar] [CrossRef]
- Beran, J. Safety and immunogenicity of a new hepatitis B vaccine for the protection of patients with renal insufficiency including pre-haemodialysis and haemodialysis patients. Expert Opin. Biol. Ther. 2008, 8, 235–247. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Song, S.; He, M. Rational design of a triple-type human papillomavirus vaccine by compromising viral-type specificity. Nat. Commun. 2018, 9, 5390. [Google Scholar] [CrossRef] [PubMed]
- Wu, T.; Li, S.W.; Zhang, J. Hepatitis E vaccine development: A 14 year odyssey. Hum. Vaccines Immunother. 2012, 8, 823–827. [Google Scholar] [CrossRef] [PubMed]
- Cai, W.; Tang, Z.M.; Wen, G.P. A high-throughput neutralizing assay for antibodies and sera against hepatitis E virus. Sci. Rep. 2016, 6, 25141. [Google Scholar] [CrossRef]
- Li, S.W.; Zhao, Q.; Wu, T. The development of a recombinant hepatitis E vaccine HEV 239. Hum. Vaccines Immunother. 2015, 11, 908–914. [Google Scholar] [CrossRef]
- Moris, P.; Jongert, E.; van der Most, R.G. Characterization of T-cell immune responses in clinical trials of the candidate RTS, S malaria vaccine. Hum. Vaccines Immunother. 2018, 14, 17–27. [Google Scholar] [CrossRef]
- Agnandji, S.T.; Lell, B.; Fernandes, J.F. A phase 3 trial of RTS, S/AS01 malaria vaccine in African infants. N. Engl. J. Med. 2012, 367, 2284–2295. [Google Scholar] [CrossRef]
- Parra, G.I.; Bok, K.; Taylor, R. Immunogenicity and specificity of norovirus Consensus GII.4 virus-like particles in monovalent and bivalent vaccine formulations. Vaccine 2012, 30, 3580–3586. [Google Scholar] [CrossRef]
- Atmar, R.L.; Baehner, F.; Cramer, J.P. Rapid Responses to 2 Virus-Like Particle Norovirus Vaccine Candidate Formulations in Healthy Adults: A Randomized Controlled Trial. J. Infect. Dis. 2016, 214, 845–853. [Google Scholar] [CrossRef]
- Nunez, I.A.; Ross, T.M. Human COBRA 2 vaccine contains two major epitopes that are responsible for eliciting neutralizing antibody responses against heterologous clades of viruses. Vaccine 2020, 38, 830–839. [Google Scholar] [CrossRef]
- Burke, R.M.; Tate, J.E.; Kirkwood, C.D. Current and new rotavirus vaccines. Curr. Opin. Infect. Dis. 2019, 32, 435–444. [Google Scholar] [CrossRef] [PubMed]
- Mattison, C.P.; Cardemil, C.V.; Hall, A. Progress on norovirus vaccine research: Public health considerations and future directions. Expert Rev. Vaccines 2018, 17, 773–784. [Google Scholar] [CrossRef]
- Atmar, R.L.; Bernstein, D.I.; Harro, C.D. Norovirus vaccine against experimental human Norwalk Virus illness. N. Engl. J. Med. 2011, 365, 2178–2187. [Google Scholar] [CrossRef] [PubMed]
- Bernstein, D.I.; Atmar, R.L.; Lyon, G.M. Norovirus vaccine against experimental human GII.4 virus illness: A challenge study in healthy adults. J. Infect. Dis. 2015, 211, 870–878. [Google Scholar] [CrossRef] [PubMed]
- Kim, L.; Liebowitz, D.; Lin, K. Safety and immunogenicity of an oral tablet norovirus vaccine, a phase I randomized, placebo-controlled trial. JCI Insight 2018, 3. [Google Scholar] [CrossRef] [PubMed]
- De Lalla, F.; Rinaldi, E.; Santoro, D. Immune response to hepatitis B vaccine given at different injection sites and by different routes: A controlled randomized study. Eur. J. Epidemiol. 1988, 4, 256–258. [Google Scholar] [CrossRef]
- Lemon, S.M.; Weber, D. Immunogenicity of plasma-derived hepatitis B vaccine: Relationship to site of injection and obesity. J. Gen. Intern. Med. 1986, 1, 199–201. [Google Scholar] [CrossRef][Green Version]
- Arnou, R.; Eavis, P.; Pardo, J.R. Immunogenicity, large scale safety and lot consistency of an intradermal influenza vaccine in adults aged 18–60 years: Randomized, controlled, phase III trial. Hum. Vaccines 2010, 6, 346–354. [Google Scholar] [CrossRef]
- Clarke, P.D.; Adams, P.; Ibanez, R. Rate, intensity, and duration of local reactions to a virosome-adjuvanted vs. an aluminium-adsorbed hepatitis A vaccine in UK travellers. Travel Med. Infect. Dis. 2006, 4, 313–318. [Google Scholar] [CrossRef]
- Zurbriggen, R.; Novak-Hofer, I.; Seelig, A. IRIV-adjuvanted hepatitis A vaccine: In vivo absorption and biophysical characterization. Prog. Lipid Res. 2000, 39, 3–18. [Google Scholar] [CrossRef]
- Fehr, T.; Bachmann, M.F.; Bucher, E. Role of repetitive antigen patterns for induction of antibodies against antibodies. J. Exp. Med. 1997, 185, 1785–1792. [Google Scholar] [CrossRef]
- Bovier, P.A. Epaxal: A virosomal vaccine to prevent hepatitis A infection. Expert Rev. Vaccines 2008, 7, 1141–1150. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Vaccine | Expression System | Adjuvant | VLP Platform | Antigen | Administration Route | Development Stage | References |
|---|---|---|---|---|---|---|---|
| Heptavax-B (Merck & Co.) | None (Plasma derived) | Aluminum hydroxide | HBsAg | SHBs | IM | Licensed | [39,40] |
| Engerix-B® (GSK, Belgium) | Yeast (S. cerevisiae) | Aluminum hydroxide | HBsAg | SHBs | IM | Licensed | [41,42,43] |
| Recombivax HB (H-B-Vax®II) (Merck & Co., USA) | Yeast (S. cerevisiae) | Aluminum sulfate | HBsAg | SHBs | IM | Licensed | [43,44] |
| Sci-B-Vac® (Bio-Hep-B®) (SciGen, Israel) | Mammalian (CHO cells) | Aluminum hydroxide | HBsAg | SHBs, MHBs, LHBs | IM | Licensed | [45,46,47,48] |
| Heplisav-B (Dynavax) | Yeast (H. polymorpha) | 1018 ISS | HBsAg | SHBs | IM | Licensed | [49,50] |
| Fendrix® (GSK, Belgium) | Yeast (S. cerevisiae) | AS04 (Aluminum hydroxide & MPL) | HBsAg | SHBs | IM | Licensed | [51] |
| Hepavax-Gene® (Crucell) | Yeast (H. polymorpha) | Aluminum hydroxide | HBsAg | SHBs, MHBs | IM | Licensed | [41,52] |
| Gardasil® (Merck & Co.) | Yeast (S. cerevisiae) | Aluminum hydroxyphosphate sulphate | HPV | HPV6/11/16/18 VLP | IM | Licensed | [53,54] |
| Cervarix® (GSK) | Insect cells (High FiveTM cells) | AS04 (Aluminum hydroxide & MPL) | HPV | HPV16/18 VLP | IM | Licensed | [55,56] |
| Gardasil-9® (Merck & Co.) | Yeast (S. cerevisiae) | Aluminum hydroxyphosphate sulfate | HPV | HPV6/11/16/18/31/33/45/52/58 VLP | IM | Licensed | [57] |
| Cecolin® (Innovax) | Bacteria (E. coli) | Aluminum hydroxide | HPV | HPV16/18 VLP | IM | Licensed | [58,59] |
| Cecolin-9® (Innovax) | Bacteria (E. coli) | Aluminum hydroxide | HPV | HPV6/11/16/18/31/33/45/52/58 VLP | IM | Phase 2 | [60] |
| Hecolin® (Innovax) | Bacteria (E. coli) | Aluminum hydroxide | HEV | HEV p239 (aa 439–617) | IM | Licensed | [61,62] |
| rHEV (GSK) | Insect cells | Aluminum hydroxide | HEV | HEV (aa 112–607) | IM | Phase 2 | [63] |
| p179 (Changchun Institute of Biological Products Co. Ltd.) | Bacteria (E. coli) | Aluminum hydroxide | HEV | HEV (aa 439–617) | IM | Phase 1 | [64] |
| tNIV (Novavax) | Insect cells (Sf-9 cells) | Matrix-M | Influenza virus | A/Hong Kong/4801/2014(H3N2) HA | IM | Phase 1/2a | [65,66] |
| gH1-Qbeta | Bacteria (E. coli) | None | Qβ-VLPs | A/California/07/2009(H1N1) HA(gH1 domain) | IM | Phase 1 | [67] |
| H5VLP+GLA vaccine | Plant | GLA-SE | Medicago VLP | A/Indonesia/05/2005(H5N1) H5 | IM | Phase 2 | [68] |
| RTS,S/AS01 (GSK) | Yeast (S. cerevisiae) | AS01 | HBsAg | P.f.CSP | IM | Phase 3 | [69] |
| R21 | Yeast (Pichia pastoris yeast strain) | Abisco-100 and Matrix-M | HBsAg | P.f.CSP | IM | Phase 1/2a | [70] |
| Norovirus VLPs and rotavirus VP6 combined vaccine | Insect cells | None | Norovirus GII-4 VLP | NoV GII-4 VLP, rVP6 | IM/ID | Pre-clinical | [71,72] |
| P2-VP8-P [8] vaccine (Walter Reed Army Institute of Research) | Bacteria (E. coli) | Aluminum hydroxide | Wa strain (G1P [8]) of human rotavirus | VP8 subunit (aa 64–223) & P2 epitope from tetanus toxin | IM | Phase 1 | [73] |
| Rotaviruses VP 2/6/7 and VP 2/6 (Baylor College of Medicine, USA) | Insect cells | None or cholera toxin | Rotavirus VLPs | VP 2/6/7 and VP 2/6 | Nasally immunized | Pre-clinical | [74] |
| Bivalent GI.1/GII.4 vaccine (Takeda Pharmaceutical Company Limited) | Insect cells (Sf-9 cells) | Aluminum hydroxide & MPL | Norovirus VLPs | GI.1VLP, GII.4 VLP | IM | Phase 2b | [75] |
| EV71 VLP C4a | Insect cells (High FiveTM cells) | Aluminum hydroxide | EV 71 VLP | EV71 C4a-gp41 | IM | Pre-clinical | [76] |
| HFMD tetravalent vaccine | Insect cells | Alhydrogel adjuvant | EV71, CVA6, CVA10, CVA16 VLP | EV71, CVA6, CVA10, CVA16 VLP | Intraperitoneally | Pre-clinical | [77] |
| RSV F (Novavax) | Insect (Sf-9 cells) | Aluminum phosphate | RSV | RSV-F | IM | Phase 1 | [78,79] |
| Rabies (TJU) | Plant (Transgenic spinach) | None | AlMV | Rabies GP/NP | Oral | Phase 1 | [80] |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Qian, C.; Liu, X.; Xu, Q.; Wang, Z.; Chen, J.; Li, T.; Zheng, Q.; Yu, H.; Gu, Y.; Li, S.; et al. Recent Progress on the Versatility of Virus-Like Particles. Vaccines 2020, 8, 139. https://doi.org/10.3390/vaccines8010139
Qian C, Liu X, Xu Q, Wang Z, Chen J, Li T, Zheng Q, Yu H, Gu Y, Li S, et al. Recent Progress on the Versatility of Virus-Like Particles. Vaccines. 2020; 8(1):139. https://doi.org/10.3390/vaccines8010139
Chicago/Turabian StyleQian, Ciying, Xinlin Liu, Qin Xu, Zhiping Wang, Jie Chen, Tingting Li, Qingbing Zheng, Hai Yu, Ying Gu, Shaowei Li, and et al. 2020. "Recent Progress on the Versatility of Virus-Like Particles" Vaccines 8, no. 1: 139. https://doi.org/10.3390/vaccines8010139
APA StyleQian, C., Liu, X., Xu, Q., Wang, Z., Chen, J., Li, T., Zheng, Q., Yu, H., Gu, Y., Li, S., & Xia, N. (2020). Recent Progress on the Versatility of Virus-Like Particles. Vaccines, 8(1), 139. https://doi.org/10.3390/vaccines8010139