The Impact of the Antigenic Composition of Chimeric Proteins on Their Immunoprotective Activity against Chronic Toxoplasmosis in Mice

 ,
,  ,
,
Abstract
1. Introduction
2. Materials and Methods
2.1. Mice
2.2. Parasites
2.3. Production of Chimeric Recombinant Proteins
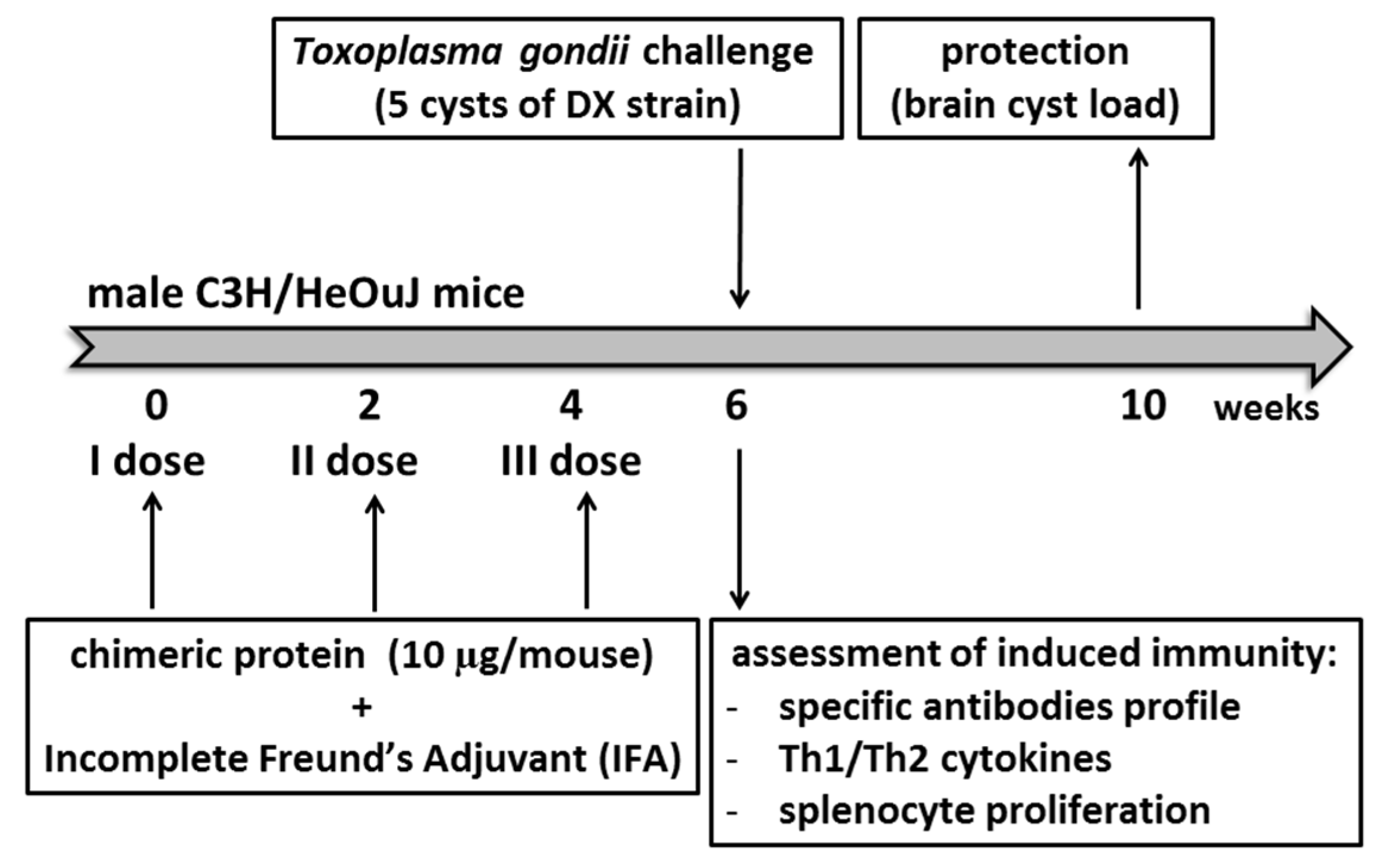
2.4. Mouse Immunization and Challenge
2.5. Brain Cyst Enumeration
2.6. Assessment of the IgG Response and Isotype Profile after Immunization
2.7. In Vitro Splenocyte Proliferation
2.8. Determination of Cytokine Production by Stimulated Splenocytes
2.9. Statistical Analysis
3. Results
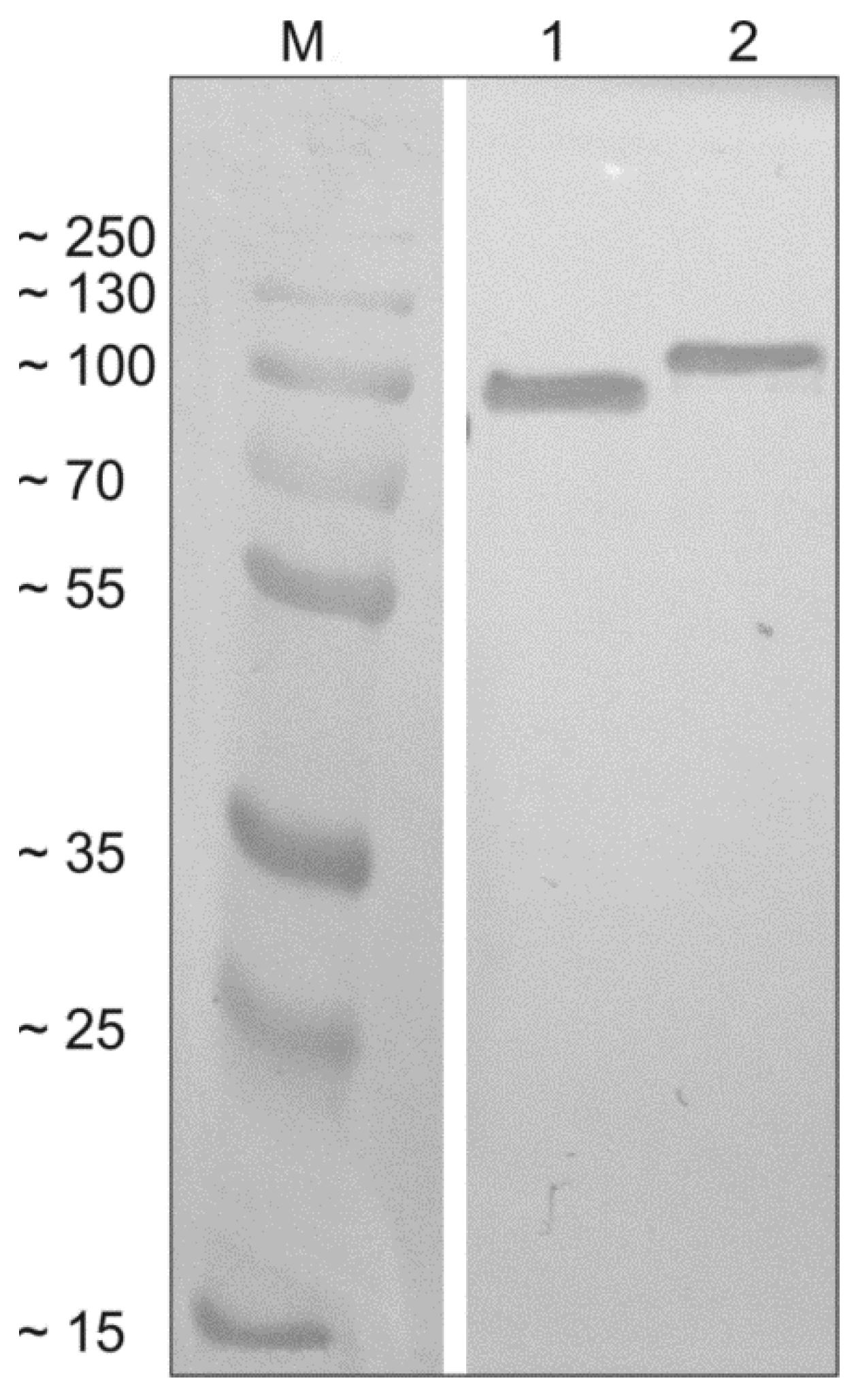
3.1. Recombinant Chimeric Proteins
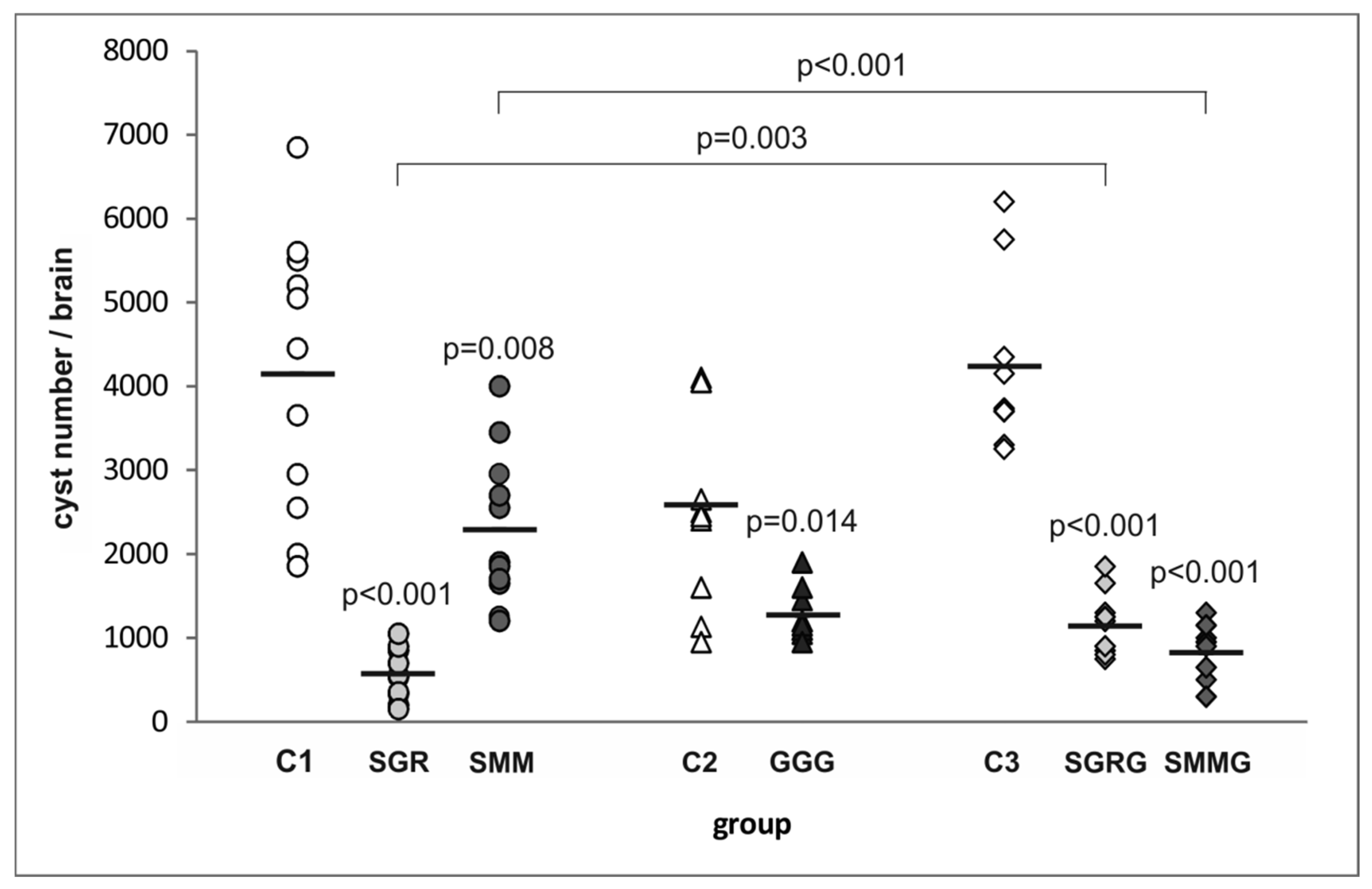
3.2. Protection Against Cyst Formation in Mice
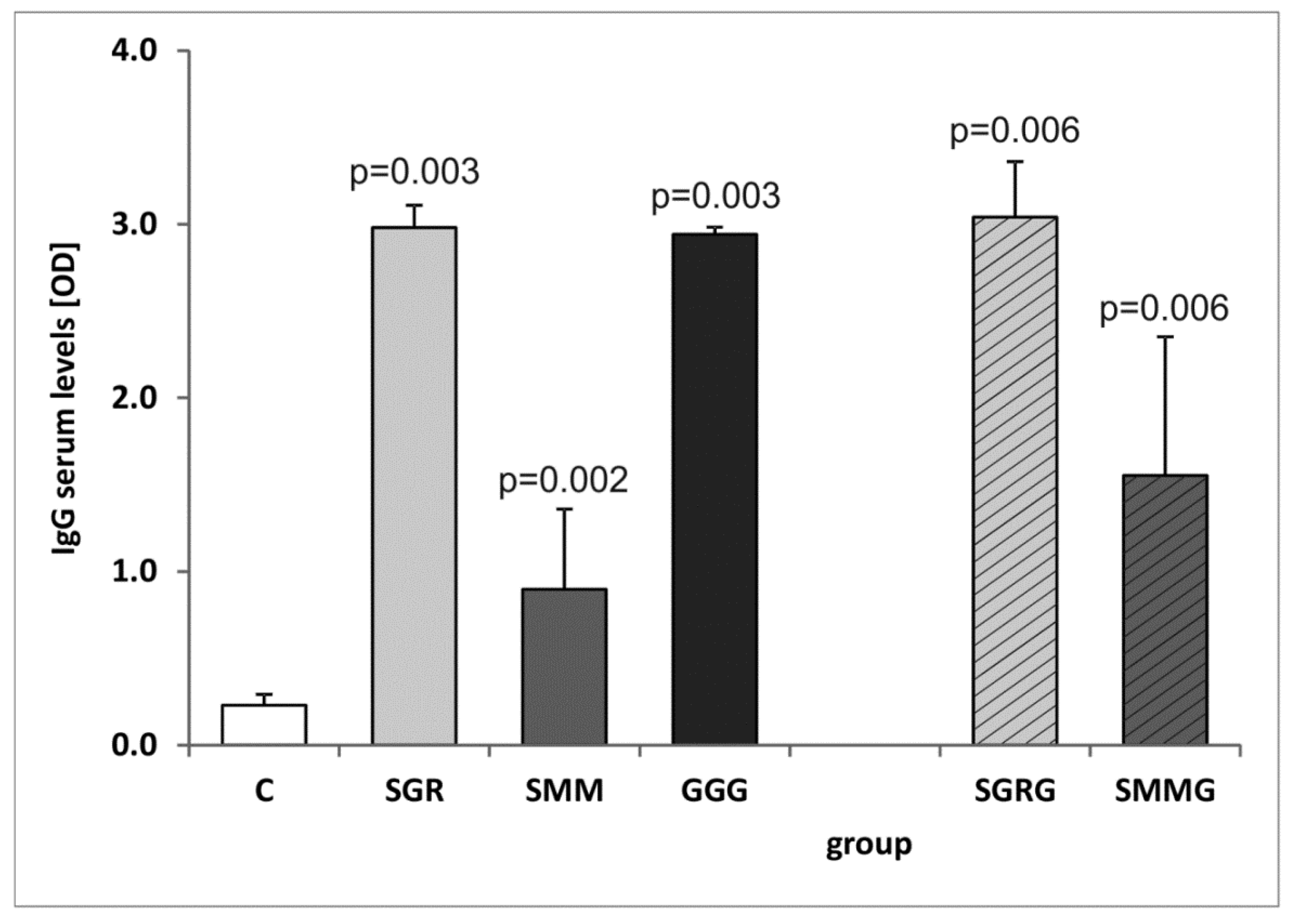
3.3. Analysis of the IgG Production and Isotype Profile
3.4. Cellular Response Induced by Vaccination
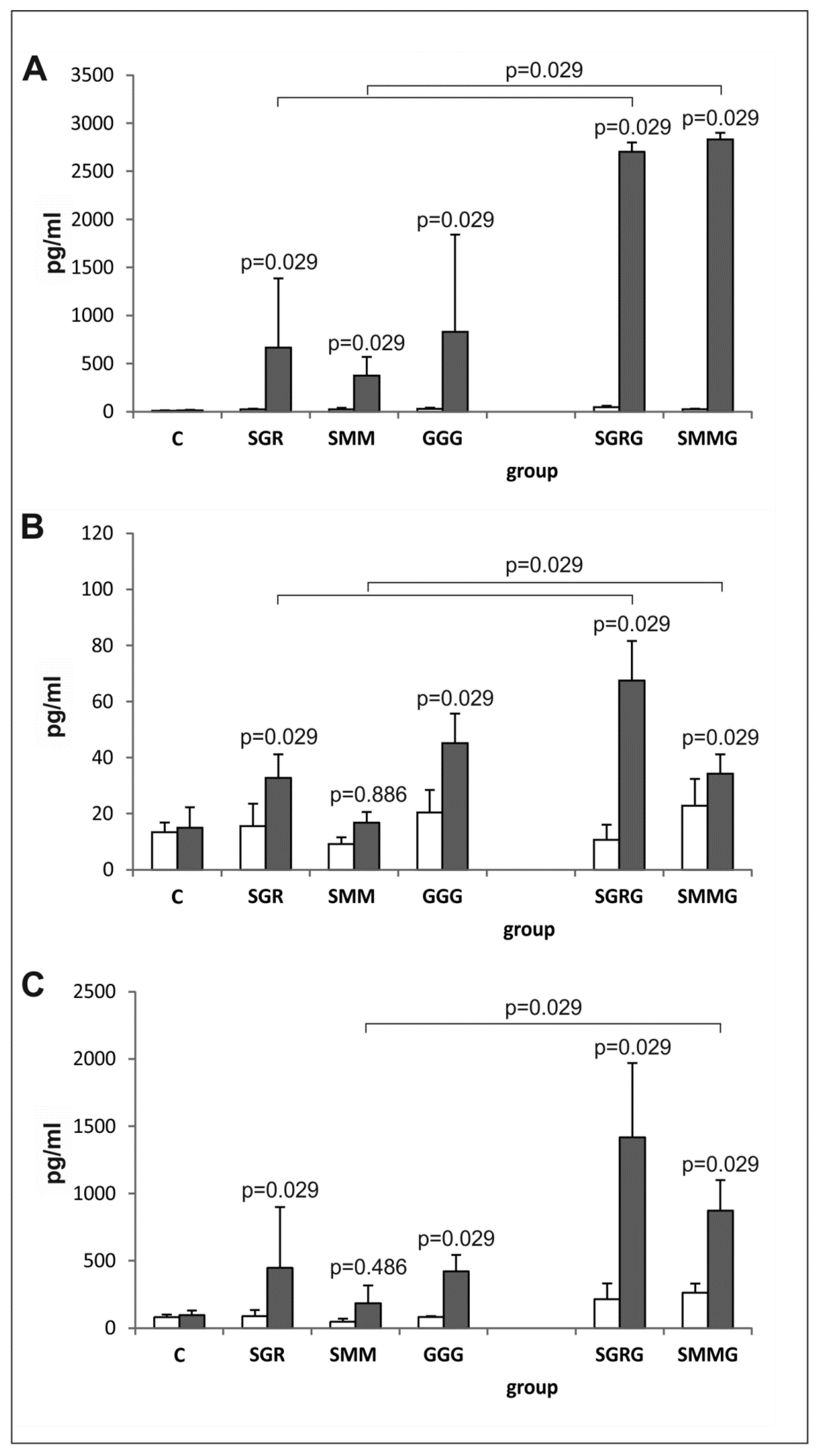
3.5. Determination of IL-2, IFN-γ and IL-10 in Vitro Synthesis
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Buxton, D. Toxoplasmosis: The first commercial vaccine. Parasitol Today 1993, 9, 335–337. [Google Scholar] [CrossRef]
- Buxton, D.; Thomson, K.M.; Maley, S.; Wright, S.; Bos, H.J. Experimental challenge of sheep 18 months after vaccination with a live (S48) Toxoplasma gondii vaccine. Vet. Rec. 1993, 133, 310–312. [Google Scholar] [CrossRef] [PubMed]
- Innes, E.A.; Bartley, P.M.; Maley, S.; Katzer, F.; Buxton, D. Veterinary vaccines against Toxoplasma gondii. Memórias Inst. Oswaldo Cruz 2009, 104, 246–251. [Google Scholar] [CrossRef] [PubMed]
- Jongert, E.; Roberts, C.W.; Gargano, N.; Förster-Waldl, E.; Petersen, E. Vaccines against Toxoplasma gondii: Challenges and opportunities. Memórias Inst. Oswaldo Cruz 2009, 104, 252–266. [Google Scholar] [CrossRef]
- Katzer, F.; Canton, G.; Burrells, A.; Palarea-Albaladejo, J.; Horton, B.; Bartley, P.M.; Pang, Y.; Chianini, F.; Innes, E.A.; Benavides, J. Immunization of lambs with the S48 strain of Toxoplasma gondii reduces tissue cyst burden following oral challenge with a complete strain of the parasite. Vet. Parasitol. 2014, 205, 46–56. [Google Scholar] [CrossRef]
- Flegr, J. How and why Toxoplasma makes us crazy. Trends Parasitol. 2013, 29, 156–163. [Google Scholar] [CrossRef]
- Hsu, P.C.; Groer, M.; Beckie, T. New findings: Depression, suicide, and Toxoplasma gondii infection. J. Am. Assoc. Nurse Pract. 2014, 26, 629–637. [Google Scholar] [CrossRef]
- Gulinello, M.; Acquarone, M.; Kim, J.H.; Spray, D.C.; Barbosa, H.S.; Sellers, R.; Tanowitz, H.B.; Weiss, L.M. Acquired infection with Toxoplasma gondii in adult mice results in sensorimotor deficits but normal cognitive behavior despite widespread brain pathology. Microbes Infect. 2010, 12, 528–537. [Google Scholar] [CrossRef]
- Kamerkar, S.; Davis, P.H. Toxoplasma on the brain: Understanding host-pathogen interactions in chronic CNS infection. J. Parasitol. Res. 2012, 589295. [Google Scholar]
- Flegr, J. Effects of Toxoplasma on human behavior. Schizophr. Bull. 2007, 33, 757–760. [Google Scholar] [CrossRef]
- Hill, D.; Dubey, J.P. Toxoplasma gondii: Transmission, diagnosis and prevention. Clin. Microbiol. Infect. 2002, 8, 634–640. [Google Scholar] [CrossRef] [PubMed]
- Innes, E.A. A brief history and overview of Toxoplasma gondii. Zoonoses Public Health 2010, 57, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Ferra, B.; Holec-Gąsior, L.; Kur, J. Serodiagnosis of Toxoplasma gondii infection in farm animals (horses, swine, and sheep) by enzyme-linked immunosorbent assay using chimeric antigens. Parasitol. Int. 2015, 64, 288–294. [Google Scholar] [CrossRef] [PubMed]
- Ferra, B.; Holec-Gąsior, L.; Kur, J. A new Toxoplasma gondii chimeric antigen containing fragments of SAG2, GRA1, and ROP1 proteins-impact of immunodominant sequences size on its diagnostic usefulness. Parasitol. Res. 2015, 114, 3291–3299. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Gatkowska, J.; Hiszczynska-Sawicka, E.; Kur, J.; Holec, L.; Dlugonska, H. Toxoplasma gondii: An evaluation of diagnostic value of recombinant antigens in a murine model. Exp. Parasitol. 2006, 114, 220–227. [Google Scholar] [CrossRef] [PubMed]
- Dziadek, B.; Gatkowska, J.; Brzostek, A.; Dziadek, J.; Dzitko, K.; Dlugonska, H. Toxoplasma gondii: The immunogenic and protective efficacy of recombinant ROP2 and ROP4 rhoptry proteins in murine experimental toxoplasmosis. Exp. Parasitol. 2009, 123, 81–89. [Google Scholar] [CrossRef]
- Holec-Gasior, L.; Kur, J.; Hiszczyńska-Sawicka, E. GRA2 and ROP1 recombinant antigens as potential markers for detection of Toxoplasma gondii-specific immunoglobulin G in humans with acute toxoplasmosis. Clin. Vaccine Immunol. 2009, 16, 510–514. [Google Scholar] [CrossRef]
- Holec-Gąsior, L.; Ferra, B.; Drapała, D.; Lautenbach, D.; Kur, J. A new MIC1-MAG1 recombinant chimeric antigen can be used instead of the Toxoplasma gondii lysate antigen in serodiagnosis of human toxoplasmosis. Clin. Vaccine Immunol. 2012, 19, 57–63. [Google Scholar] [CrossRef]
- Dziadek, B.; Gatkowska, J.; Brzostek, A.; Dziadek, J.; Dzitko, K.; Grzybowski, M.; Dlugonska, H. Evaluation of three recombinant multi-antigenic vaccines composed of surface and secretory antigens of Toxoplasma gondii in murine models of experimental toxoplasmosis. Vaccine 2011, 29, 821–830. [Google Scholar] [CrossRef]
- Gatkowska, J.; Wieczorek, M.; Dziadek, B.; Dzitko, K.; Dziadek, J.; Długońska, H. Assessment of the antigenic and neuroprotective activity of the subunit anti-Toxoplasma vaccine in T. gondii experimentally infected mice. Vet. Parasitol. 2018, 254, 82–94. [Google Scholar] [CrossRef]
- Foroutan, M.; Ghaffarifar, F.; Sharifi, Z.; Dalimi, A.; Jorjani, O. Rhoptry antigens as Toxoplasma gondii vaccine target. Clin. Exp. Vaccine Res. 2019, 8, 4–26. [Google Scholar] [CrossRef] [PubMed]
- Gatkowska, J.; Gasior, A.; Kur, J.; Dlugonska, H. Toxoplasma gondii: Chimeric Dr fimbriae as a recombinant vaccine against toxoplasmosis. Exp. Parasitol. 2008, 118, 266–270. [Google Scholar] [CrossRef] [PubMed]
- Costa-Silva, T.A.; Borges, M.M.; Galhardo, C.S.; Pereira-Chioccola, V.L. Immunization with excreted/secreted proteins in AS/n mice activating cellular and humoral response against Toxoplasma gondii infection. Acta Trop. 2012, 124, 203–209. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Zhang, D.; Wang, G.; Yin, H.; Wang, M. Immunization with excreted-secreted antigens reduces tissue cyst formation in pigs. Parasitol. Res. 2013, 112, 3835–3842. [Google Scholar] [CrossRef] [PubMed]
- Wagner, A.; Schabussova, I.; Ruttkowski, B.; Peschke, R.; Kur, J.; Kundi, M.; Joachim, A.; Wiedermann, U. Prime-boost vaccination with toxoplasma lysate antigen, but not with a mixture of recombinant protein antigens, leads to reduction of brain cyst formation in BALB/c mice. PLoS ONE 2015, 10, e0126334. [Google Scholar] [CrossRef]
- Allahyari, M.; Mohabati, R.; Amiri, S.; Esmaeili Rastaghi, A.R.; Babaie, J.; Mahdavi, M.; Vatanara, A.; Golkar, M. Synergistic effect of rSAG1 and rGRA2 antigens formulated in PLGA microspheres in eliciting immune protection against Toxoplasama gondii. Exp. Parasitol. 2016, 170, 236–246. [Google Scholar] [CrossRef]
- Wang, Y.; Yin, H. Research progress on surface antigen 1 (SAG1) of Toxoplasma gondii. Parasit. Vectors 2014, 7, 180. [Google Scholar] [CrossRef]
- Ferguson, D.J.; Parmley, S.F. Toxoplasma gondii MAG1 protein expression. Trends Parasitol. 2002, 18, 482. [Google Scholar] [CrossRef]
- Lourenco, E.V.; Bernardes, E.S.; Silva, N.M.; Mineo, J.R.; Panunto-Castelo, A.; Roque-Barreira, M.C. Immunization with MIC1 and MIC4 induces protective immunity against Toxoplasma gondii. Microbes Infect. 2006, 8, 1244–1251. [Google Scholar] [CrossRef]
- Pinzan, C.F.; Sardinha-Silva, A.; Almeida, F.; Lai, L.; Lopes, C.D.; Lourenço, E.V.; Panunto-Castelo, A.; Matthews, S.; Roque-Barreira, M.C. Vaccination with Recombinant Microneme Proteins Confers Protection against Experimental Toxoplasmosis in Mice. PLoS ONE 2015, 10, e0143087. [Google Scholar] [CrossRef]
- Scheiblhofer, S.; Laimer, J.; Machado, Y.; Weiss, R.; Thalhamer, J. Influence of protein fold stability on immunogenicity and its implications for vaccine design. Expert Rev. Vaccines 2017, 16, 479–489. [Google Scholar] [CrossRef] [PubMed]
- Letscher-Bru, V.; Villard, O.; Risse, B.; Zauke, M.; Klein, J.P.; Kien, T.T. Protective effect of vaccination with a combination of recombinant surface antigen 1 and interleukin-12 against toxoplasmosis in mice. Infect. Immun. 1998, 66, 4503–4506. [Google Scholar] [PubMed]
- Nielsen, H.V.; Di Cristina, M.; Beghetto, E.; Spadoni, A.; Petersen, E.; Gargano, N. Toxoplasma gondii: DNA vaccination with bradyzoite antigens induces protective immunity in mice against oral infection with parasite cysts. Exp. Parasitol. 2006, 112, 274–279. [Google Scholar] [CrossRef] [PubMed]
- Parmley, S.; Slifer, T.; Araujo, F. Protective effects of immunization with a recombinant cyst antigen in mouse models of infection with Toxoplasma gondii tissue cysts. J. Infect. Dis. 2002, 185, S90–S95. [Google Scholar] [CrossRef] [PubMed]
- Sonaimuthu, P.; Ching, X.T.; Fong, M.Y.; Kalyanasundaram, R.; Lau, Y.L. Induction of Protective Immunity against Toxoplasmosis in BALB/c Mice Vaccinated with Toxoplasma gondii Rhoptry-1. Front. Microbiol. 2016, 7, 808. [Google Scholar] [CrossRef]
- Beghetto, E.; Nielsen, H.V.; Del Porto, P.; Buffolano, W.; Guglietta, S.; Felici, F.; Petersen, E.; Gargano, N. A combination of antigenic regions of Toxoplasma gondii microneme proteins induces protective immunity against oral infection with parasite cysts. J. Infect. Dis. 2005, 191, 637–645. [Google Scholar] [CrossRef]
- Golkar, M.; Shokrgozar, M.A.; Rafati, S.; Musset, K.; Assmar, M.; Sadaie, R.; Cesbron-Delauw, M.F.; Mercier, C. Evaluation of protective effect of recombinant dense granule antigens GRA2 and GRA6 formulated in monophosphoryl lipid A (MPL) adjuvant against Toxoplasma chronic infection in mice. Vaccine 2007, 25, 4301–4311. [Google Scholar] [CrossRef]
- Bastos, L.M.; Macêdo, A.G., Jr.; Silva, M.V.; Santiago, F.M.; Ramos, E.L.; Santos, F.A.; Pirovani, C.P.; Goulart, L.R.; Mineo, T.W.; Mineo, J.R. Toxoplasma gondii-Derived Synthetic Peptides Containing B- and T-Cell Epitopes from GRA2 Protein Are Able to Enhance Mice Survival in a Model of Experimental Toxoplasmosis. Front. Cell. Infect. Microbiol. 2016, 6, 59. [Google Scholar] [CrossRef]
- Carruthers, V.B.; Suzuki, Y. Effects of Toxoplasma gondii infection on the brain. Schizophr. Bull. 2007, 33, 745–751. [Google Scholar] [CrossRef]
- Gazzinelli, R.T.; Wysocka, M.; Hieny, S.; Scharton-Kersten, T.; Cheever, A.; Kühn, R.; Müller, W.; Trinchieri, G.; Sher, A. In the absence of endogenous IL-10, mice acutely infected with Toxoplasma gondii succumb to a lethal immune response dependent on CD4+ T cells and accompanied by overproduction of IL-12, IFN-gamma and TNF-alpha. J. Immunol. 1996, 157, 798–805. [Google Scholar]
- Roers, A.; Siewe, L.; Strittmatter, E.; Deckert, M.; Schlüter, D.; Stenzel, W.; Gruber, A.D.; Krieg, T.; Rajewsky, K.; Müller, W. T cell-specific inactivation of the interleukin 10 gene in mice results in enhanced T cell responses but normal innate responses to lipopolysaccharide or skin irritation. J. Exp. Med. 2004, 200, 1289–1297. [Google Scholar] [CrossRef] [PubMed]
- Rosenberg, C.; De Craeye, S.; Jongert, E.; Gargano, N.; Beghetto, E.; Del Porto, P.; Vorup-Jensen, T.; Petersen, E. Induction of partial protection against infection with Toxoplasma gondii genotype II by DNA vaccination with recombinant chimeric tachyzoite antigens. Vaccine 2009, 27, 2489–2498. [Google Scholar] [CrossRef] [PubMed]
- Grzybowski, M.M.; Dziadek, B.; Gatkowska, J.M.; Dzitko, K.; Długońska, H. Towards vaccine against toxoplasmosis: Evaluation of the immunogenic and protective activity of recombinant ROP5 and ROP18 Toxoplasma gondii proteins. Parasitol. Res. 2015, 114, 4553–4563. [Google Scholar] [CrossRef] [PubMed]
- Yang, C.D.; Chang, G.N.; Chao, D. Protective immunity against Toxoplasma gondii in mice induced by a chimeric protein rSAG1/2. Parasitol. Res. 2004, 92, 58–64. [Google Scholar] [CrossRef] [PubMed]
- Chuang, S.C.; Chung, Y.C.; Yang, C.D. Protective immunity against toxoplasmosis in mice induced by single-dose immunization with rSAG1/2 protein released from poly(lactide-co-glycolide) microparticles. Parasite 2017, 24, 5. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.; Shi, L.; Cheng, Y.B.; Fan, G.X.; Ren, H.X.; Yuan, Y.K. Evaluation of protective effect of multi-epitope DNA vaccine encoding six antigen segments of Toxoplasma gondii in mice. Parasitol. Res. 2009, 105, 267–274. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Fragment | Primer Name | Primer Sequence | Corresponding Protein Residues |
|---|---|---|---|
| gra2 | SMMG-For | 5′-GGGATCTGGTAAGCTGGGAAAAGGTGAACATACACCACC-3′ | GRA2 51-185 (135 aa) |
| SMMG-Rev | 5′-GGTGGTGGTGCTCGATCTGCGAAAAGTCTG-3′ | (cloning to the pET30/SAG1-MIC1-MAG1) | |
| SGRG-For | 5′-TGGATCGCAAGCGATGGGAAAAGGTGAACATACACCACC-3′ | GRA2 51-185 (135 aa) | |
| SGRG-Rev | 5′-CAAGCTTGTCGACGGTCTGCGAAAAGTCTGGGACGGGCA-3′ | (cloning to the pET30/SAG2-GRA1-ROP1) |
| Trivalent Chimeric Proteins | Tetravalent Chimeric Proteins | |||||
|---|---|---|---|---|---|---|
| IgG1 | IgG2a | IgG1/IgG2a | IgG1 | IgG2a | IgG1/IgG2a | |
| SGR | SGRG | |||||
| mouse 1 | 51,200 | 3200 | 16/1 | >204,800 | 6400 | >32/1 |
| mouse 2 | >204,800 | 12,800 | >16/1 | 204,800 | 102,400 | >2/1 |
| mouse 3 | >204,800 | 102,400 | >2/1 | 204,800 | 12,800 | 16/1 |
| mouse 4 | 102,400 | 25,600 | 4/1 | 102,400 | 6400 | 16/1 |
| mouse 5 | 102,400 | 6400 | 16/1 | - | - | - |
| SMM | SMMG | |||||
| mouse 1 | >204,800 | 51,200 | 4/1 | >204,800 | 51,200 | >4/1 |
| mouse 2 | >204,800 | 204,800 | >1/1 | 102,400 | 3200 | 32/1 |
| mouse 3 | 102,400 | 12,800 | 8/1 | >204,800 | 25,600 | >8/1 |
| mouse 4 | >204,800 | 51,200 | >4/1 | 204,800 | 12,800 | >16/1 |
| mouse 5 | 102,400 | 1600 | 64/1 | - | - | - |
| mouse 6 | 204,800 | 6400 | 32/1 | - | - | - |
| GGG | ||||||
| mouse 1 | >204,800 | 51,200 | >4/1 | - | - | - |
| mouse 2 | >204,800 | 102,400 | >2/1 | - | - | - |
| mouse 3 | 204,800 | 25,600 | 8/1 | - | - | - |
| mouse 4 | >204,800 | 12,800 | >16/1 | - | - | - |
| mouse 5 | 204,800 | 6400 | >32/1 | - | - | - |
| Antigen/Group | Mean % in Proportion to TLA Stimulated Control Cells ± SD * | |||
|---|---|---|---|---|
| Control | SGR | SMM | GGG | |
| medium | 87.32 ± 17.72 | 87.02 ± 16.29 | 96.54 ± 14.31 | 106.21 ± 20.66 |
| TLA | 100.00 ± 17.34 | 99.33 ± 15.29 | 121.56 ± 26.19 * | 130.84 ± 19.63 * |
| SGRG | SMMG | - | ||
| medium | 76.22 ± 2.20 | 88.35 ± 20.29 | - | |
| TLA | 104.25 ± 2.62 | 109.94 ± 14.19 | - | |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gatkowska, J.; Dzitko, K.; Ferra, B.T.; Holec-Gąsior, L.; Kawka, M.; Dziadek, B. The Impact of the Antigenic Composition of Chimeric Proteins on Their Immunoprotective Activity against Chronic Toxoplasmosis in Mice. Vaccines 2019, 7, 154. https://doi.org/10.3390/vaccines7040154
Gatkowska J, Dzitko K, Ferra BT, Holec-Gąsior L, Kawka M, Dziadek B. The Impact of the Antigenic Composition of Chimeric Proteins on Their Immunoprotective Activity against Chronic Toxoplasmosis in Mice. Vaccines. 2019; 7(4):154. https://doi.org/10.3390/vaccines7040154
Chicago/Turabian StyleGatkowska, Justyna, Katarzyna Dzitko, Bartłomiej Tomasz Ferra, Lucyna Holec-Gąsior, Malwina Kawka, and Bożena Dziadek. 2019. "The Impact of the Antigenic Composition of Chimeric Proteins on Their Immunoprotective Activity against Chronic Toxoplasmosis in Mice" Vaccines 7, no. 4: 154. https://doi.org/10.3390/vaccines7040154
APA StyleGatkowska, J., Dzitko, K., Ferra, B. T., Holec-Gąsior, L., Kawka, M., & Dziadek, B. (2019). The Impact of the Antigenic Composition of Chimeric Proteins on Their Immunoprotective Activity against Chronic Toxoplasmosis in Mice. Vaccines, 7(4), 154. https://doi.org/10.3390/vaccines7040154