Role of Antisperm Antibodies in Infertility, Pregnancy, and Potential for Contraceptive and Antifertility Vaccine Designs: Research Progress and Pioneering Vision


 ,
,  ,
,
Abstract
1. Introduction
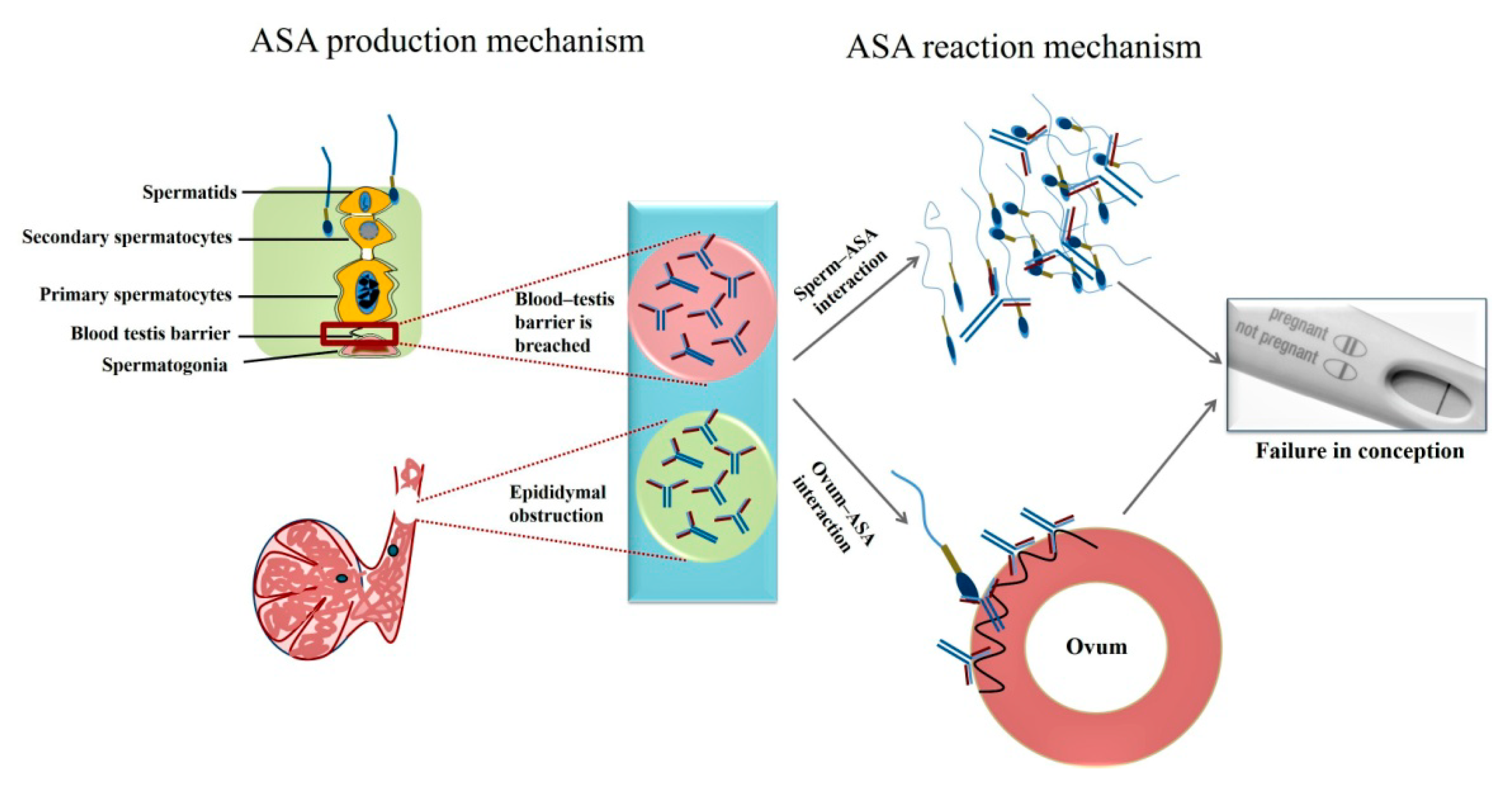
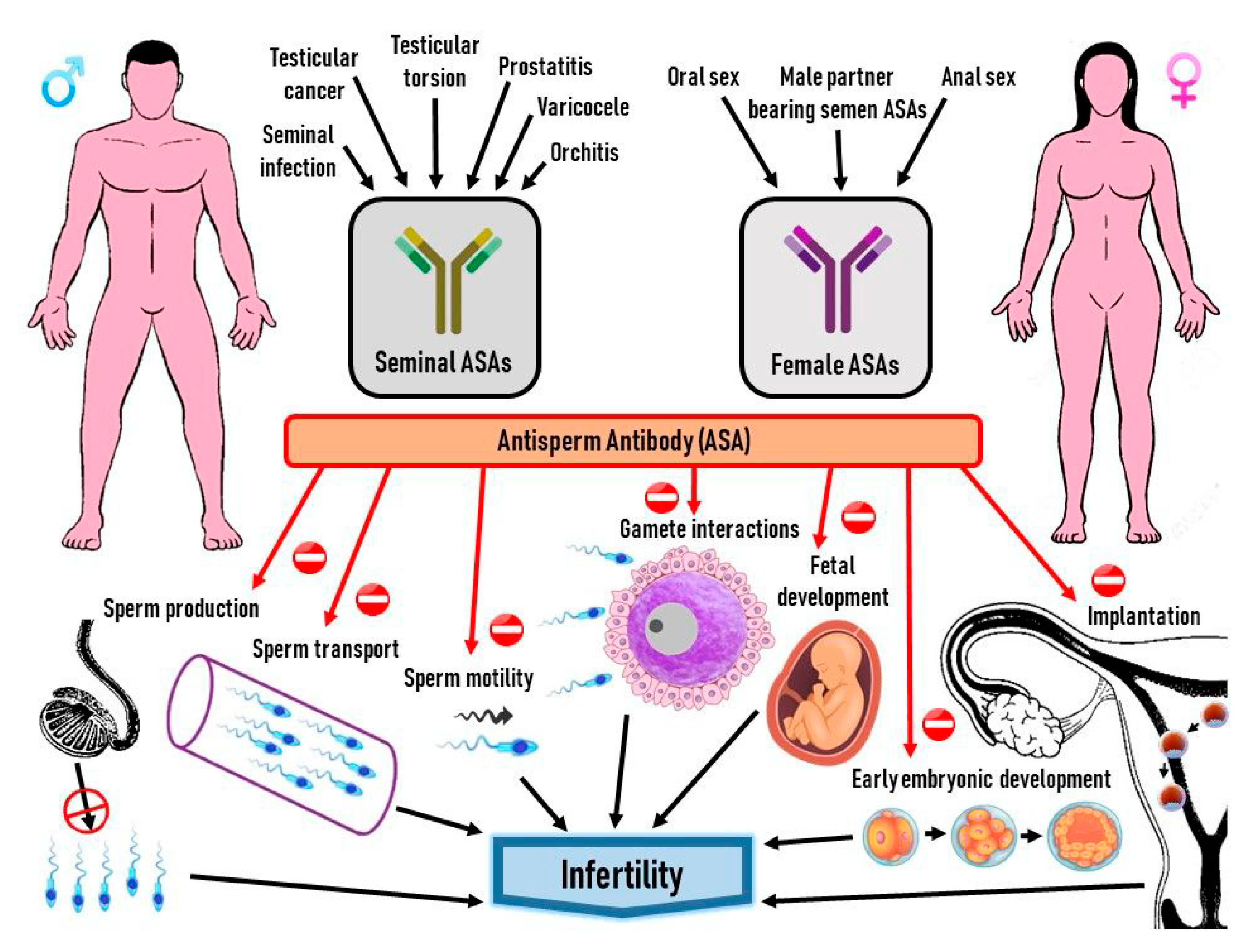
2. Role of Antisperm Antibodies in Infertility
2.1. Detection Methods for Antisperm Antibodies
2.2. Diagnosis of Obstructive Azoospermia UsingSerum ASA Values
2.3. ASA and Salpingitis
2.4. ASA and Oligoasthenoteratozoospermia (OAT)
2.5. ASA and Normospermia and Vasectomy Reversal
2.6. ASA in Cryptorchidism
2.7. ASA Influence on Semen Parameter
2.8. VaricocelesAssociated with ASAs
2.9. Circulating ASAs in Polycystic Ovarian Syndrome (PCOS)
2.10. ASAs and TheirInfluence on the Sperm Acrosome Reaction
2.11. Influence of ASAs on Sperm Transport
2.12. ASAs in the Complement System
3. Role of Antisperm Antibodies in Pregnancy
ASAs and Missed Abortions
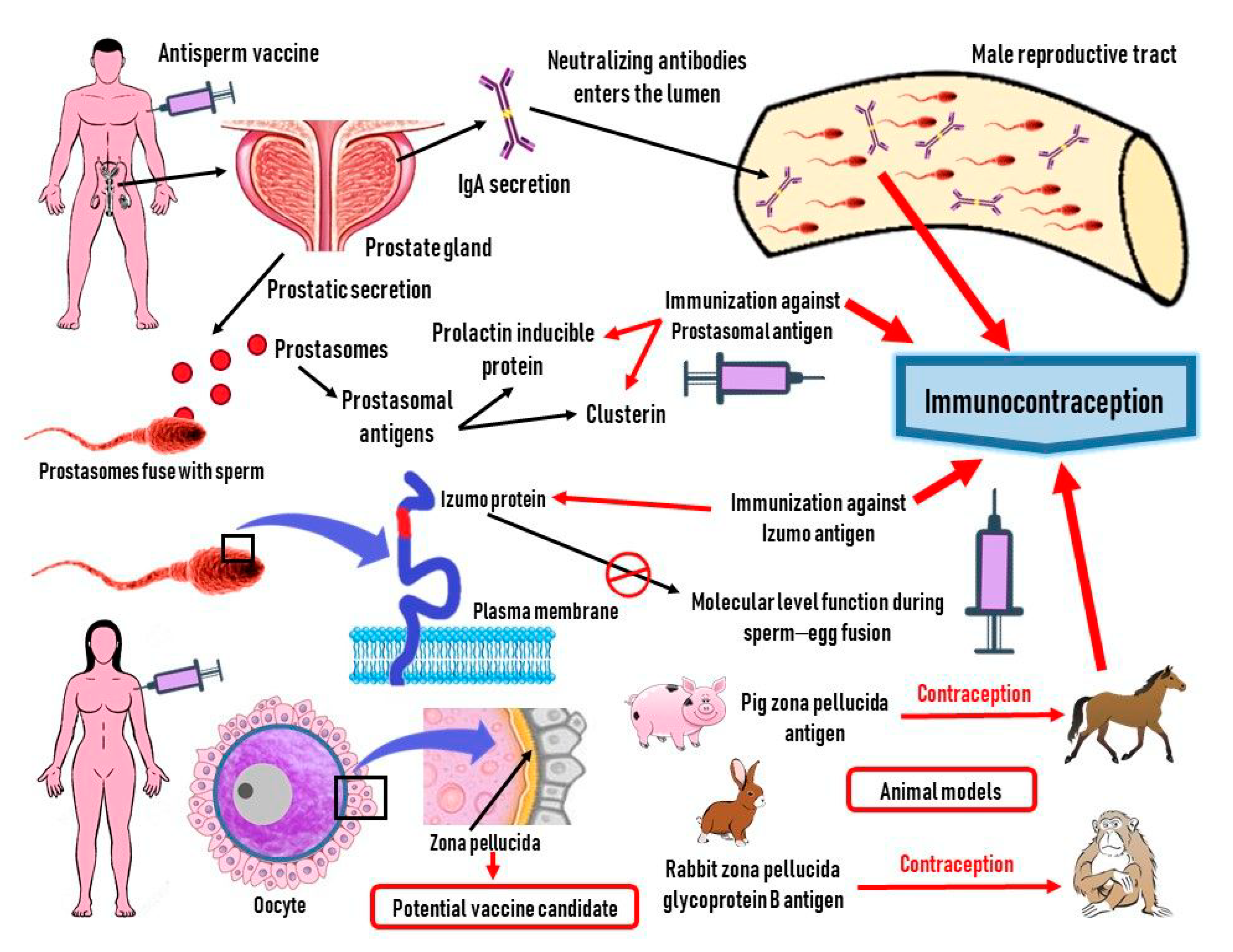
4. Role of ASAs in Designing Effective Contraception
4.1. Antifertility Vaccines
4.2. ASAs in Contraception
Sperm and Oocyte Proteome and Immunological Infertility
4.3. Oocyte Antigens in Contraception
4.4. ASA-Derived Infertility and Their Treatment
5. Other Methods and Modes of Contraceptives
6. Patents Related to ASAs and Their Application as Contraceptives
7. Concluding Remarks and Future Perspectives
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Ronquist, G. Prostasomes: Their characterisation: Implications for human reproduction. In The Male Role in Pregnancy Loss and Embryo Implantation Failure; Springer International Publishing: Berlin/Heidelberg, Germany, 2015; pp. 191–209. [Google Scholar]
- Vazquez-Levin, M.H.; Marin-Briggiler, C.I.; Veaute, C. Antisperm antibodies: Invaluable tools toward the identification of sperm proteins involved in fertilization. Am. J. Reprod. Immunol. 2014, 72, 206–218. [Google Scholar] [CrossRef] [PubMed]
- Bozhedomov, V.A.; Nikolaeva, M.A.; Ushakova, I.V.; Lipatova, N.A.; Bozhedomova, G.E.; Sukhikh, G.T. Functional deficit of sperm and fertility impairment in men with antisperm antibodies. J. Reprod. Immunol. 2015, 112, 95–101. [Google Scholar] [CrossRef] [PubMed]
- Vivas, A.G.; Lozano, H.J.; Velasco, J. Immune-testicular regulation and cytokines. Investig. Clin. 2007, 48, 107–121. [Google Scholar]
- Check, J.H. Antisperm antibodies and human reproduction. Clin. Exp. Obstet. Gynecol. 2010, 37, 169–174. [Google Scholar] [PubMed]
- Yousif, S.; Thamer, F.; Shihab, N.; Latif, M.O.; Abdulhadi, B. Measurement of antisperm antibodies (ASA) type IgG in Iraqi infertile male. J. Gene C Environ. Resour. Conserv. 2015, 3, 202–204. [Google Scholar]
- Cui, D.; Han, G.; Shang, Y.; Liu, C.; Xia, L.; Li, L.; Yi, S. Antisperm antibodies in infertile men and their effect on semen parameters: A systematic review and meta-analysis. Clin. Chim. Acta 2015, 444, 29–36. [Google Scholar] [CrossRef] [PubMed]
- Rumke, P. The presence of sperm antibodies in the serum of two patients with oligospermia. Vox. Sang 1954, 4, 135–140. [Google Scholar]
- Wilson, L. Sperm agglutinins in human semen and blood. Proc. Soc. Exp. Biol. Med. 1954, 85, 652–655. [Google Scholar] [CrossRef]
- Naz, R.K. Antisperm Contraceptive Vaccine. In Immune Infertility; Springer International Publishing: Berlin/Heidelberg, Germany, 2017; pp. 249–261. [Google Scholar]
- Thaper, D.; Vander, H.; Prabha, V. Association of antisperm antibodies with bacterial infection: An Insight to Infertility. Androl. Gynecol. 2014, 2. [Google Scholar] [CrossRef]
- Thaper, D.; Prabha, V. Molecular mimicry: An explanation for autoimmune diseases and infertility. Scand. J. Immunol. 2018, 23, e12697. [Google Scholar] [CrossRef]
- Restrepo, B.; Cardona-Maya, W. Antisperm antibodies and fertility association. Acta Urol. Espanol. 2013, 37, 571–578. [Google Scholar] [CrossRef] [PubMed]
- Selvaraj, K.; Selvaraj, P. Chapter 24: Immunology in Infertility. In Principles & Practice of Assisted Reproductive Technology; Rao, K., Carp, H., Fischer, R., Eds.; JP Medical Ltd.: London, UK, 2014. [Google Scholar]
- Jensen, C.F.S.; Ostergren, P.; Dupree, J.M.; Ohl, D.A.; Sonksen, J.; Fode, M. Varicocele and male infertility. Nat. Rev. Urol. 2017, 14, 523–533. [Google Scholar] [CrossRef] [PubMed]
- Hertl, M.; Krause, W.K. Sperm-Specific T Lymphocytes. In Immune Infertility; Springer International Publishing: Berlin/Heidelberg, Germany, 2017; pp. 125–132. [Google Scholar]
- Sinisi, A.A.; Finizio, B.; Pasquali, D.; Scurini, C.; D’apuzzo, A.; Bellastella, A. Prevalence of antisperm antibodies by SpermMARtest in subjects undergoing a routine sperm analysis for infertility. Int. J. Androl. 1993, 16, 311–314. [Google Scholar] [CrossRef] [PubMed]
- Marconi, M.; Pilatz, A.; Wagenlehner, F.; Diemer, T.; Weidner, W. Are antisperm antibodies really associated with proven chronic inflammatory and infectious diseases of the male reproductive tract? Eur. Urol. 2009, 56, 708–715. [Google Scholar] [CrossRef] [PubMed]
- Mazumdar, S.; Levine, A.S. Antisperm antibodies: Etiology, pathogenesis, diagnosis, and treatment. Fertil. Steril. 1998, 70, 799–810. [Google Scholar] [CrossRef]
- Eggert-Kruse, W.; Rohr, G.; Probst, S.; Rusu, R.; Hund, M.; Demirakca, T.; Aufenanger, J.; Runnebaum, B.; Petzoldt, D. Antisperm antibodies and microorganisms in genital secretions-a clinically significant relationship? Andrologia 1998, 30, 61–71. [Google Scholar] [CrossRef] [PubMed]
- Eggert-Kruse, W.; Buhlinger-Göpfarth, N.; Rohr, G.; Probst, S.; Aufenanger, J.; Näher, H.; Runnebaum, B. Immunology: Antibodies to Chlamydia trachomatis in semen and relationship with parameters of male fertility. Hum. Reprod. 1996, 11, 1408–1417. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Kortebani, G.; Gonzales, G.F.; Barrera, C.; Mazzolli, A.B. Leucocyte populations in semen and male accessory gland function: Relationship with antisperm antibodies and seminal quality. Andrologia 1992, 24, 197–204. [Google Scholar] [CrossRef] [PubMed]
- Komori, K.; Tsujimura, A.; Miura, H.; Shin, M.; Takada, T.; Honda, M.; Matsumiya, K.; Fujioka, H. Serial follow-up study of serum testosterone and antisperm antibodies in patients with non-obstructive azoospermia after conventional or microdissection testicular sperm extraction. Int. J. Androl. 2004, 27, 32–36. [Google Scholar] [CrossRef] [PubMed]
- Kendirci, M.; Hellstrom, W.J. Antisperm antibodies and varicocele. South. Med. J. 2006, 99, 13–15. [Google Scholar] [CrossRef] [PubMed]
- Munuce, M.J.; Berta, C.L.; Pauluzzi, F.; Caille, A.M. Relationship between antisperm antibodies, sperm movement, and semen quality. Urol. Int. 2000, 65, 200–203. [Google Scholar] [CrossRef] [PubMed]
- Fijak, M.; Pilatz, A.; Hedger, M.P.; Nicolas, N.; Bhushan, S.; Michel, V.; Tung, K.S.K.; Schuppe, H.C.; Meinhardt, A. Infectious, inflammatory and ‘autoimmune’ male factor infertility: How do rodent models inform clinical practice? Hum. Reprod. Update 2018, 24, 416–441. [Google Scholar] [CrossRef] [PubMed]
- Lee, R.; Goldstein, M.; Ullery, B.W.; Ehrlich, J.; Soares, M.; Razzano, R.A.; Herman, M.P.; Callahan, M.A.; Li, P.S.; Schlegel, P.N. Value of serum antisperm antibodies in diagnosing obstructive azoospermia. J. Urol. 2009, 181, 264–269. [Google Scholar] [CrossRef] [PubMed]
- Esteves, S.C.; Schneider, D.T.; Verza, S., Jr. Influence of antisperm antibodies in the semen of intracytoplasmic sperm injection outcome. Int. Braz. J. Urol. 2007, 33, 795–802. [Google Scholar] [CrossRef] [PubMed]
- Wallach, E.E.; Bronson, R.; Cooper, G.; Rosenfeld, D. Sperm antibodies: Their role in infertility. Fertil. Steril. 1984, 42, 171–183. [Google Scholar] [CrossRef]
- Chamley, L.W.; Clarke, G.N. Antisperm antibodies and conception. In Seminars in Immunopathol; Springer: Berlin/Heidelberg, Germany, 2007; Volume 29, pp. 169–184. [Google Scholar]
- Zini, A.; Fahmy, N.; Belzile, E.; Ciampi, A.; Al-Hathal, N.; Kotb, A. Antisperm antibodies are not associated with pregnancy rates after IVF and ICSI: Systematic review and meta-analysis. Hum. Reprod. 2011, 26, 1288–1295. [Google Scholar] [CrossRef]
- Zini, A.; Lefebvre, J.; Kornitzer, G.; Bissonnette, F.; Kadoch, I.J.; Dean, N.; Phillips, S. Anti-sperm antibody levels are not related to fertilization or pregnancy rates after IVF or IVF/ICSI. J. Reprod. Immunol. 2011, 88, 80–84. [Google Scholar] [CrossRef] [PubMed]
- Clarke, G.N. Etiology of sperm immunity in women. Fertil. Steril. 2009, 91, 639–643. [Google Scholar] [CrossRef] [PubMed]
- Naz, R.K.; Menge, A.C. Antisperm antibodies: Origin, regulation, and sperm reactivity in human infertility. Fertil. Steril. 1994, 61, 1001–1013. [Google Scholar] [CrossRef]
- Kremer, J.; Jager, S. OPINION: The significance of antisperm antibodies for sperm-cervical mucus interaction. Hum. Reprod. 1992, 7, 781–784. [Google Scholar] [CrossRef]
- Wallach, E.; Diller, L.; Hembree, W. Male contraception and family planning: A social and historical review. Fertil. Steril. 1977, 28, 1271–1279. [Google Scholar] [CrossRef]
- Cheng, C.Y.; Mruk, D.D. The blood-testis barrier and its implications for male contraception. Pharmacol. Rev. 2012, 64, 16–64. [Google Scholar] [CrossRef] [PubMed]
- Delves, P.J.; Lund, T.; Roitt, I.M. Antifertility vaccines. Trends Immunol. 2002, 23, 213–219. [Google Scholar] [CrossRef]
- Primakoff, P.; Lathrop, W.; Woolman, L.; Cowan, A.; Myles, D. Fully effective contraception in male and female guinea pigs immunized with the sperm protein PH-20. Nature 1988, 335, 543–546. [Google Scholar] [CrossRef] [PubMed]
- Pujianto, D.A.; Hajizah, H.; Mansur, I.G.; Amarudin, A. Antisperm antibodies disrupt plasma membrane integrity and inhibit tyrosine phosphorylation in human spermatozoa. Med. J. Indones. 2018, 27, 3. [Google Scholar] [CrossRef]
- Shibahara, H.; Koriyama, J. Methods for direct and indirect antisperm antibody testing. In Spermatogenesis; Humana Press: Totowa, NJ, USA, 2013; pp. 51–60. [Google Scholar]
- Al-Fahham, A.A. Etiology of antisperm antibodies in the serum of virgins. Open J. Obstet. Gynecol. 2018, 8, 236–245. [Google Scholar] [CrossRef]
- Clarke, G.N.; Elliott, P.J.; Smaila, C. Detection of sperm antibodies in semen using the immunobead test: A survey of 813 consecutive patients. Am. J. Reprod. Immunol. Microbiol. 1985, 7, 118–123. [Google Scholar] [CrossRef]
- Wakimoto, Y.; Fukui, A.; Kojima, T.; Hasegawa, A.; Shigeta, M.; Shibahara, H. Application of computer-aided sperm analysis (CASA) for detecting sperm-immobilizing antibody. Am. J. Reprod. Immunol. 2018, 79, e12814. [Google Scholar] [CrossRef]
- Wigby, S.; Suarez, S.S.; Lazzaro, B.P.; Pizzari, T.; Wolfner, M.F. Sperm success and immunity. Cur. Top. Dev. Biol. 2019, 135, 287–313. [Google Scholar]
- Dondero, F.; Gandini, L.; Lombardo, F.; Salacone, P.; Caponecchia, L.; Lenzi, A. Antisperm antibody detection: 1. Methods and standard protocol. Am. J. Reprod. Immunol. 1997, 38, 218–223. [Google Scholar] [CrossRef]
- Dondero, F.; Lenzi, A.; Gandini, L.; Lombardo, F.; Culasso, F. A comparison of the direct immunobead test and other tests for sperm antibodies detection. J. Endocrinol. Investig. 1991, 14, 443–449. [Google Scholar] [CrossRef] [PubMed]
- Lenzi, A.; Gandini, L.; Lombardo, F.; Rago, R.; Paoli, D.; Dondero, F. Antisperm antibody detection: 2. Clinical, biological, and statistical correlation between methods. Am. J. Reprod. Immunol. 1997, 38, 224–230. [Google Scholar] [CrossRef] [PubMed]
- Hendry, W.F.; Parslow, J.M.; Stedronska, J. Exploratory scrototomy in 168 azoospermic males. BJU Int. 1983, 55, 785–791. [Google Scholar] [CrossRef] [PubMed]
- Miller, S.; Couture, S.; James, G.; Plourde, S.; Rioux, J.; Labrecque, M. Unilateral absence of vas deferens: Prevalence among 23,013 men seeking vasectomy. Int. Braz. J. Urol. 2016, 42, 1010–1017. [Google Scholar] [CrossRef] [PubMed]
- Çek, M.; Sturdza, L.; Pilatz, A. Acute and chronic epididymitis. Eur. Urol. Suppl. 2017, 16, 124–131. [Google Scholar] [CrossRef]
- Al-Salem, A.H. Acute Scrotum. In An Illustrated Guide to Pediatric Urology; Springer International Publishing: Berlin/Heidelberg, Germany, 2017; pp. 601–617. [Google Scholar]
- Chua, M.E.; Mendoza, J.S.; Gaston, M.J.; Luna, S.L.; Morales, M.L. Hormonal therapy using gonadotropin releasing hormone for improvement of fertility index among children with cryptorchidism: A meta-analysis and systematic review. J. Pediatr. Surg. 2014, 49, 1659–1667. [Google Scholar] [CrossRef] [PubMed]
- Cobellis, G.; Noviello, C.; Nino, F.; Romano, M.; Mariscoli, F.; Martino, A.; Parmeggiani, P.; Papparella, A. Spermatogenesis and cryptorchidism. Front. Endocrinol. 2014, 5, 63. [Google Scholar] [CrossRef] [PubMed]
- Marconi, M.; Weidner, W. Site and risk factors of antisperm antibodies production in the male population. In Immune Infertility; Springer International Publishing: Berlin/Heidelberg, Germany, 2017; pp. 133–147. [Google Scholar]
- Marconi, M.; Weidner, W. Re: Value of serum antisperm antibodies in diagnosing obstructive azoospermia. Eur. Urol. 2009, 56, 215–216. [Google Scholar] [CrossRef]
- Gubin, D.A.; Dmochowski, R.; Kutteh, W.H. Multivariant analysis of men from infertile couples with and without antisperm antibodies. Am. J. Reprod. Immunol. 1998, 39, 157–160. [Google Scholar] [CrossRef]
- World Health Organization. Manual for the Standardized Investigation and Diagnosis of the Infertile Couple; Cambridge University Press: Cambridge, UK, 1993. [Google Scholar]
- World Health Organization. Challenges in Reproductive Health Research: Biennial Report 1992–1993; World Health Organization: Geneva, Switzerland, 1994. [Google Scholar]
- Bukar, M.; Mustapha, Z.; Takai, U.I.; Tahir, A. Hysterosalpingographic findings in infertile women: A seven year review. Nigeria J. Clin. Pract. 2011, 14, 168–170. [Google Scholar] [CrossRef]
- Taylor, B.D.; Darville, T.; Haggerty, C.L. Does bacterial vaginosis cause pelvic inflammatory disease? Sex. Transm. Dis. 2013, 40, 117–122. [Google Scholar] [CrossRef] [PubMed]
- Chhetri, A.; Biswas, S.C.; Gupta, K. Infection and Infertility. In Practical Guide in Infertility; Jaypee Brothers Medical Publishers: Delhi, India, 2018; p. 107. [Google Scholar]
- Spencer, T.H.; Umeh, P.O.; Irokanulo, E.; Baba, M.M.; Spencer, B.B.; Umar, A.I.; Ardzard, S.A.; Oderinde, S.; Onoja, O. Bacterial isolates associated with pelvic inflammatory disease among female patients attending some hospitals in Abuja, Nigeria. Afr. J. Infect. Dis. 2014, 8, 9–13. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Taylor-Robinson, D.; Jensen, J.S.; Svenstrup, H.; Stacey, C.M. Difficulties experienced in defining the microbial cause of pelvic inflammatory disease. Int. J. STD AIDS 2012, 23, 18–24. [Google Scholar] [CrossRef]
- Romosan, G.; Bjartling, C.; Skoog, L.; Valentin, L. Ultrasound for diagnosing acute salpingitis: A prospective observational diagnostic study. Hum. Reprod. 2013, 28, 1569–1579. [Google Scholar] [CrossRef] [PubMed]
- Rajaratnam, A.; D’Cunha, P.; Furtado, Z.; Fernandes, H. Tuberculous salpingitis: A case report. J. Clin. Diag. Res. JCDR 2013, 7, 1186–1188. [Google Scholar] [CrossRef] [PubMed]
- Liu, Q.Z.; Zhu, W.J.; Liu, H.P.; Jiang, H. Evaluation on antisperm antibody in infertile women with chronic salpingitis. J. Reprod. Contracept. 2014, 25, 159–163. [Google Scholar]
- Koide, S.S.; Wang, L.; Kamada, M. Antisperm antibodies associated with infertility: Properties and encoding genes of target antigens. Proc. Soc. Exp. Biol. Med. 2000, 224, 123–132. [Google Scholar] [CrossRef]
- Mugo, N.R.; Kiehlbauch, J.; Kiviat, N.; Nguti, R.; Gichuhi, J.W.; Stamm, W.E.; Cohen, C.R. Endometrial histopathology in patients with laparoscopic proven salpingitis and HIV-1 infection. Infect. Dis. Obstet. Gynecol. 2011, 2011, 407057. [Google Scholar] [CrossRef] [PubMed]
- Taylor, B.D.; Haggerty, C.L. Management of Chlamydia trachomatis genital tract infection: Screening and treatment challenges. Infect. Drug. Resist. 2011, 4, 19–29. [Google Scholar] [CrossRef]
- Malhotra, M.; Sood, S.; Mukherjee, A.; Muralidhar, S.; Bala, M. Genital Chlamydia trachomatis: An update. Ind. J. Med. Res. 2013, 138, 303–316. [Google Scholar]
- Baccetti, B.; Capitani, S.; Collodel, G.; Strehler, E.; Piomboni, P. Recent advances in human sperm pathology. Contraception 2002, 65, 283–287. [Google Scholar] [CrossRef]
- Cavallini, G. Male idiopathic oligoasthenoteratozoospermia. As. J. Androl. 2006, 8, 143–157. [Google Scholar] [CrossRef] [PubMed]
- Aydos, S.E.; Tukun, A. Infertility in a man with oligoasthenoteratozoospermia associated with nonrobertsonian translocation t (9; 15)(p10; q10). Fertil. Steril. 2006, 86, 1001.e7–e9. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.C.; Jing, L.I.; Liang, W.B.; Zhu, W.J. Evaluation on antisperm antibody in infertile men with oligoasthenoteratozoospermia. J. Reprod. Contracept. 2014, 25, 49–53. [Google Scholar]
- Guven, E.S.; Dilbaz, S.; Ceylaner, S.; Acar, H.; Cinar, O.; Ozdegirmenci, O.; Karcaaltincaba, D. An uncommon complementary isochromosome of 46, XY, i (9)(p10), i (9)(q10) in an infertile oligoasthenoteratozoospermic man. Fertil. Steril. 2011, 95, 290.e5–e8. [Google Scholar] [CrossRef] [PubMed]
- Miyazaki, T.; Mori, M.; Yoshida, C.A.; Ito, C.; Yamatoya, K.; Moriishi, T.; Kawai, Y.; Komori, H.; Kawane, T.; Izumi, S.; et al. Galnt3 deficiency disrupts acrosome formation and leads to oligoasthenoteratozoospermia. Histochem. Cell Biol. 2013, 139, 339–354. [Google Scholar] [CrossRef] [PubMed]
- Pelliccione, F.; d’Angeli, A.; Cinque, B.; Falone, S.; Micillo, A.; Francavilla, F.; Amicarelli, F.; Gandini, L.; Francavilla, S. Activation of the immune system and sperm DNA fragmentation are associated with idiopathic oligoasthenoteratospermia in men with couple subfertility. Fertil. Steril. 2011, 95, 2676–2679. [Google Scholar] [CrossRef] [PubMed]
- Flickinger, C.J.; Howards, S.S.; Herr, J.C. Effects of vasectomy on the epididymis. Microsc. Res. Technol. 1995, 30, 82–100. [Google Scholar] [CrossRef] [PubMed]
- Newton, R.A. IgG antisperm antibodies attached to sperm do not correlate with infertility following vasovasostomy. Microsurgery 1988, 9, 278–280. [Google Scholar] [CrossRef] [PubMed]
- Royle, M.G.; Parslo, J.; Kingscott, W.; Wallace, D.M.; Hendry, W.F. Reversal of vasectomy: The effects of sperm antibodies on subsequent fertility. BJU Int. 1981, 53, 654–659. [Google Scholar] [CrossRef] [PubMed]
- Farber, N.J.; Flannigan, R.; Li, P.; Li, P.S.; Goldstein, M. The kinetics of sperm return and late failure following vasovasostomy or vasoepididymostomy: A systematic review. J. Urol. 2019, 201, 241–250. [Google Scholar] [CrossRef] [PubMed]
- Nduwayo, L.; Barthelemy, C.; Lansac, J.; Tharanne, M.J.; Lecomte, P. Management of necrospermia. Contracept. Fertil. Sex. 1992, 23, 682–685. [Google Scholar]
- Mallidis, C.; Lim, T.C.; Hill, S.T.; Skinner, D.J.; Brown, D.J.; Johnston, W.I.; Baker, H.W. Necrospermia and chronic spinal cord injury. Fertil. Steril. 2000, 74, 221–227. [Google Scholar] [CrossRef]
- Coutinho, E.M.; Melo, J.F.; Barbosa, I.; Segal, S.J. Antispermatogenic action of gossypol in men. Fertil. Steril. 1984, 42, 424–430. [Google Scholar] [CrossRef]
- El-Gothamy, Z.; El-Samahy, M. Ultrastructure sperm defects in addicts. Fertil. Steril. 1992, 57, 699–702. [Google Scholar] [CrossRef]
- Chavez-Badiola, A.; Drakeley, A.J.; Finney, V.; Sajjad, Y.; Lewis-Jones, D.I. Necrospermia, antisperm antibodies, and vasectomy. Fertil. Steril. 2008, 89, 723.e5–e7. [Google Scholar] [CrossRef] [PubMed]
- Hauser, R.; Yogev, L.; Paz, G.; Yavetz, H.; Azem, F.; Lessing, J.B.; Botchan, A. Comparison of efficacy of two techniques for testicular sperm retrieval in nonobstructive azoospermia: Multifocal testicular sperm extraction versus multifocal testicular sperm aspiration. J. Androl. 2006, 27, 28–33. [Google Scholar] [CrossRef] [PubMed]
- Urry, R.L.; Carrell, D.T.; Starr, N.T.; Snow, B.W.; Middleton, R.G. The incidence of antisperm antibodies in infertility patients with a history of cryptorchidism. J. Urol. 1994, 151, 381–383. [Google Scholar] [CrossRef]
- Jiang, H.; Zhu, W.J. Cryptorchidism is not a risk factor for antisperm antibody production in post-orchidopexy males with infertility. Urol. Int. 2013, 90, 470–474. [Google Scholar] [CrossRef]
- Barthold, J.S.; Gonzalez, R. The epidemiology of congenital cryptorchidism, testicular ascent and orchiopexy. J. Urol. 2003, 170, 2396–2401. [Google Scholar] [CrossRef]
- Walsh, T.J.; Dall’Era, M.A.; Croughan, M.S.; Carroll, P.R.; Turek, P.J. Prepubertal orchiopexy for cryptorchidism may be associated with lower risk of testicular cancer. J. Urol. 2007, 178, 1440–1446. [Google Scholar] [CrossRef] [PubMed]
- Kuiri-Hänninen, T.; Koskenniemi, J.; Dunkel, L.; Toppari, J.; Sankilampi, U. Postnatal testicular activity in healthy boys and boys with cryptorchidism. Front. Endocrin. 2019, 10, 489. [Google Scholar] [CrossRef] [PubMed]
- Patel, R.P.; Kolon, T.F.; Huff, D.S.; Carr, M.C.; Zderic, S.A.; Canning, D.A.; Snyder, H.M., 3rd. Testicular microlithiasis and antisperm antibodies following testicular biopsy in boys with cryptorchidism. J. Urol. 2005, 174, 2008–2010. [Google Scholar] [CrossRef] [PubMed]
- Miniberg, D.T.; Chen, M.E.; Witkin, S.S. Antisperm antibodies in cryptorchid boys. Eur. J. Pediatr. 1993, 52 (Suppl. 2), S23–S24. [Google Scholar] [CrossRef] [PubMed]
- Sinisi, A.A.; Pasquali, D.; Papparella, A.; Valente, A.; Orio, F.; Esposito, E.; Cobellis, G.; Cuomo, A.; Angelone, G.; Martone, A.; et al. Antisperm antibodies in cryptorchidism before and after surgery. J. Urol. 1998, 160, 1834–1837. [Google Scholar] [CrossRef]
- Zini, A.; Phillips, S.; Lefebvre, J.; Baazeem, A.; Bissonnette, F.; Kadoch, I.J.; Gabriel, M.S. Anti-sperm antibodies are not associated with sperm DNA damage: A prospective study of infertile men. J. Reprod. Immunol. 2010, 85, 205–208. [Google Scholar] [CrossRef]
- Matsuda, Y.; Oshio, S.; Yazaki, T.; Umeda, T.; Akihama, S. The effect of some proteinase inhibitors on liquefaction of human semen. Hum. Reprod. 1994, 9, 664–668. [Google Scholar] [CrossRef] [PubMed]
- Check, J.H.; Aly, J. Sperm antibodies and assisted reproduction. In Immune Infertility; Springer: Cham, Switzerland, 2017; pp. 223–234. [Google Scholar]
- Garcia Patricia, C.; Eliana, M.R.; Oduvaldo, C.M.P. Antisperm antibodies in infertilemen and their correlation with seminal parameters. Reprod. Med. Biol. 2007, 6, 33–38. [Google Scholar] [CrossRef] [PubMed]
- Lynch Dona, M.; Howe Stephen, E. Comparison of a direct and indirect ELISA for quantitating antisperm antibody in semen. J. Androl. 1987, 8, 215–220. [Google Scholar] [CrossRef]
- Chao, D. Discuss the seminal plasma antisperm antibody, and the association between semen parameters and sperm morphology. Asia Pac. Tradit. Med. 2011, 7, 104–105. [Google Scholar]
- Chu, Y.; Lin, Y. Antisperm antibody in sperm surface, and clinical analysis of semen parameters in infertility male. J. Fam. Plan. 2004, 10, 162–164. [Google Scholar]
- Rossato, M.; Galeazzi, C.; Ferigo, M.; Foresta, C. Antisperm antibodies modify plasma membrane functional integrity and inhibit osmosensitive calcium influx in human sperm. Hum. Reprod. 2004, 19, 1816–1820. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Yeh, W.R.; Acosta, A.A.; Seltman, H.J.; Doncel, G. Impact of immunoglobulin isotype and sperm surface location of antisperm antibodies on fertilization in vitro in the human. Fertil. Steril. 1995, 63, 1287–1292. [Google Scholar] [CrossRef]
- Oliveros, S.Y.; Sanchez, L.M.; Noda, M.F.; Curbelo, A.J. Anticuerpos antiespermatozoides en parejas con infertilidad de causa no explicada. Rev. Cubana Endocrinol. 2000, 11, 11–17. [Google Scholar]
- Zini, A.; Dohle, G. Are varicoceles associated with increased deoxyribonucleic acid fragmentation? Fertil. Steril. 2011, 96, 1283–1287. [Google Scholar] [CrossRef]
- Takihara, H.; Sakatoku, J.; Cockett, A.T.K. The pathophysiology of varicocele in male infertility. Fertil. Steril. 1991, 55, 861–868. [Google Scholar]
- Will, M.A.; Swain, J.; Fode, M.; Sonksen, J.; Christman, G.M.; Ohl, D. The great debate. Varicocele treatment and impact on fertility. Fertil. Steril. 2011, 95, 841–852. [Google Scholar] [CrossRef]
- Mehrsai, A.; Valojerdi, M.R.; Djaladat, H.; Poormand, G. Evaluation of antisperm antibodies in infertile men associated with varicocele. Pre and post varicocelectomy. Saudi Med. J. 2005, 26, 1479–1481. [Google Scholar]
- Golomb, J.; Vardinon, N.; Homonnai, Z.T.; Braf, Z.; Yust, I. Demonstration of antispermatozoal antibodies in varicocele-related infertility with an enzyme-linked immunosorbent assay (ELISA). Fertil. Steril. 1986, 45, 397–402. [Google Scholar] [CrossRef]
- Bozhedomov, V.A.; Lipatova, N.A.; Alexeev, R.A.; Alexandrova, L.M.; Nikolaeva, M.A.; Sukhikh, G.T. The role of the antisperm antibodies in male infertility assessment after microsurgical varicelectomy. Andrologia 2014, 2, 847–855. [Google Scholar] [CrossRef]
- Bozhedomov, V.A.; Lipatova, N.A.; Rokhlikov, I.M.; Alexeev, R.A.; Ushakova, I.V.; Sukhikh, G.T. Male fertility and varicocoele: Role of immune factors. Andrologia 2014, 2, 51–58. [Google Scholar] [CrossRef] [PubMed]
- Palacio, J.R.; Iborra, A.; Ulcova-Gallova, Z.; Badia, R.; Martinez, P. The presence of antibodies to oxidative modified proteins in serum from polycystic ovary syndrome patients. Clin. Exp. Immunol. 2006, 144, 217–222. [Google Scholar] [CrossRef] [PubMed]
- Haller-Kikkatalo, K.; Salumets, A.; Uibo, R. Review on autoimmune reactions in female infertility: Antibodies to follicle stimulating hormone. Clin. Dev. Immunol. 2012, 2012, 762541. [Google Scholar] [CrossRef] [PubMed]
- Alborzi, S.; Tavazoo, F.; Dehaghani, A.S.; Ghaderi, A.; Alborzi, S.; Alborzi, M. Determination of antiovarian antibodies after laparoscopic ovarian electrocauterization in patients with polycystic ovary syndrome. Fertil. Steril. 2009, 91, 1159–1163. [Google Scholar] [CrossRef] [PubMed]
- Adams, J.M.; Taylor, A.E.; Crowley, W.F., Jr.; Hall, J.E. Polycystic ovarian morphology with regular ovulatory cycles: Insights into the pathophysiology of polycystic ovarian syndrome. J. Clin. Endocrinol. Metabol. 2004, 89, 4343–4350. [Google Scholar] [CrossRef] [PubMed]
- Morin-Papunen, L.C.; Duleba, A.J.; Bloigu, A.; Järvelin, M.R.; Saikku, P.; Pouta, A. Chlamydia antibodies and self-reported symptoms of oligo-amenorrhea and hirsutism: A new etiologic factor in polycystic ovary syndrome? Fertil. Steril. 2010, 94, 1799–1804. [Google Scholar] [CrossRef] [PubMed]
- Lazarov, V.V.; Trifonova, N.L.; Dimitrova-Dikanarova, D.K. Antibodies against immunologically sequestered antigens in patients with polycystic ovary syndrome (PCOS). Akush. Ginekol. (Sofiia) 2016, 55, 35–39. [Google Scholar] [PubMed]
- Chernyshov, V.P.; Radysh, T.V.; Gura, I.V.; Tatarchuk, T.P.; Khominskaya, Z.B. Immune disorders in women with premature ovarian failure in initial period. Am. J. Reprod. Immunol. 2001, 46, 220–225. [Google Scholar] [CrossRef]
- Singla, R.; Gupta, Y.; Khemani, M.; Aggarwal, S. Thyroid disorders and polycystic ovary syndrome: An emerging relationship. Ind. J. Endocrinol. Metabol. 2015, 19, 25–29. [Google Scholar] [CrossRef]
- Malinowski, A.; Dyński, M.A.; Głowacka, E.; Nowak, M.; Wilczyński, J.R.; Kolasa, D.; Tchórzewski, H.; Szpakowski, M. [Antiphospholipid autoantibodies in women treated for infertility]. Ginekol. Pol. 2000, 71, 1011–1016. [Google Scholar]
- Yang, X.Y.; Zhu, W.J.; Jing, L.I. Evaluation of Circulating Antisperm Antibody in Infertile Women with Polycystic Ovary Syndrome. J. Reprod. Contracept. 2012, 23, 25–28. [Google Scholar] [CrossRef]
- Myogo, K.; Yamano, S.; Nakagawa, K.; Kamada, M.; Maegawa, M.; Irahara, M.; Aono, T. Sperm-immobilizing antibodies block capacitation in human spermatozoa. Arch. Androl. 2001, 47, 135–142. [Google Scholar] [CrossRef]
- Bandoh, R.; Yamano, S.; Kamada, M.; Daitoh, T.; Aono, T. Effect of sperm-immobilizing antibodies on the acrosome reaction of human spermatozoa. Fertil. Steril. 1992, 57, 387–392. [Google Scholar] [CrossRef]
- Ziegler, A.; Hirsch, T.; Krause, W.; Neumann, K.; Schieferstein, G.; Dohr, G.; Kohlstädt, S.; Uchanska-Ziegler, B. Monoclonal antibodies against antigens expressed by human sperm: Monoklonale Antikörper mit Spezifität für humane Spermatozoenantigene. Andrologia 1990, 22, 101–109. [Google Scholar] [CrossRef]
- Francavilla, F.; Romano, R.; Santucci, R.; Marrone, V.; Properzi, G.; Ruvolo, G. Interference of antisperm antibodies with the induction of the acrosome reaction by zona pellucida (ZP) and its relationship with the inhibition of ZP binding. Fertil. Steril. 1997, 67, 1128–1133. [Google Scholar] [CrossRef]
- Schwarzinger, F.; Schüller, T.; Krause, W. Flow cytometric determination of binding of monoclonal antibodies to human spermatozoa. Andrologia 1997, 29, 163–168. [Google Scholar] [CrossRef] [PubMed]
- Fenichel, P.; Hsi, B.L.; Farahifar, D.; Donzeaut, M.; Barrier-Delpech, D.; Yeh, C.J. Evaluation of the human sperm acrosome reaction using a monoclonal antibody, GB24, and fluorescence-activated cell sorter. J. Reprod. Fertil. 1989, 87, 699–706. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Uhler, M.L.; Leung, A.; Chan, S.Y.; Schmid, I.; Wang, C. Assessment of human sperm acrosome reaction by flow cytometry: Validation and evaluation of the method by fluorescence-activated cell sorting. Fertil. Steril. 1993, 60, 1076–1081. [Google Scholar] [CrossRef]
- Bohring, C.; Skrzypek, J.; Krause, W. Influence of antisperm antibodies on the acrosome reaction as determined by flow cytometry. Fertil. Steril. 2001, 76, 275–280. [Google Scholar] [CrossRef]
- Tamburrino, L.; Marchiani, S.; Minetti, F.; Forti, G.; Muratori, M.; Baldi, E. The CatSper calcium channel in human sperm: Relation with motility and involvement in progesterone-induced acrosome reaction. Hum. Reprod. 2014, 29, 418–428. [Google Scholar] [CrossRef] [PubMed]
- Baron, L.; Fara, K.; Zapata-Carmona, H.; Zuniga, L.; Kong, M.; Signorelli, J.; Diaz, E.S.; Morales, P. Participation of protein kinases and phosphatases in the progesterone-induced acrosome reaction and calcium influx in human spermatozoa. Andrologia 2016, 4, 1073–1083. [Google Scholar] [CrossRef] [PubMed]
- Sebkova, N.; Ded, L.; Vesela, K.; Dvorakova-Hortova, K. Progress of sperm IZUMO1 relocation during spontaneous acrosome reaction. Reproduction 2014, 147, 231–240. [Google Scholar] [CrossRef] [PubMed]
- Xuan, X.J.; Xu, C.; Zhao, Y.R.; Wu, K.L.; Chen, T.; Zhang, H.B.; Li, X.; Su, S.Z.; Ma, G.; Tang, R.; et al. Application of spontaneous acrosome reaction of sperm in prediction of outcome of in-vitro fertilization and embryo transfer. Zhonghua Yi Xue Za Zhi 2016, 96, 1285–1288. [Google Scholar] [PubMed]
- Godinez-Solis, Y.; Martinez-Aguilar, G.; Solis-Heredia, M.J.; Quintanilla-Vega, B. Spontaneous sperm acrosome reaction is altered by subchronic lead exposure in mice. Toxicol. Lett. 2016, 259, S234. [Google Scholar] [CrossRef]
- Ferrer, M.S.; Klabnik-Bradford, J.; Anderson, D.E.; Bullington, A.C.; Palomares, R.A.; Miller, L.M.J.; Stawicki, R.; Miesner, M. Sperm-bound antisperm antibodies prevent capacitation of bovine spermatozoa. Theriogenology 2017, 89, 58–67. [Google Scholar] [CrossRef] [PubMed]
- Chiu, W.W.; Chamley, L.W. Clinical associations and mechanisms of action of antisperm antibodies. Fertil. Steril. 2004, 82, 529–535. [Google Scholar] [CrossRef]
- Jager, S.; Kremer, J.; van Slochteren-Draaisma, T. A simple method of screening for antisperm antibodies in the human male. Detection of spermatozoal surface IgG with the direct mixed antiglobulin reaction carried out on untreated fresh human semen. Int. J. Fertil. 1978, 23, 12–21. [Google Scholar] [PubMed]
- Haas, G.G.; Schreiber, A.D.; Blasco, L. The incidence of sperm-associated immunoglobulin and C3, the third component of complement, in infertile men. Fertil. Steril. 1983, 39, 542–547. [Google Scholar] [CrossRef]
- Menge, A.C.; Beitner, O. Interrelationships among semen characteristics, antisperm antibodies, and cervical mucus penetration assays in infertile human couples. Fertil. Steril. 1989, 51, 486–492. [Google Scholar] [CrossRef]
- Wang, C.; Baker, H.G.; Jennings, M.G.; Burger, H.G.; Lutjen, P. Interaction between human cervical mucus and sperm surface antibodies. Fertil. Steril. 1985, 44, 484–488. [Google Scholar] [CrossRef]
- Mortimer, D.; Pandya, I.J.; Sawers, R.S. Relationship between human sperm motility characteristics and sperm penetration into human cervical mucus in vitro. J. Reprod. Fertil. 1986, 78, 93–102. [Google Scholar] [CrossRef] [PubMed]
- Kremer, J.; Jager, S. Characteristics of anti-spermatozoal antibodies responsible for the shaking phenomenon with special regard to immunoglobulin class and antigen-reactive sites. Int. J. Androl. 1980, 3, 143–152. [Google Scholar] [CrossRef] [PubMed]
- Han, L.; Taub, R.; Jensen, J.T. Cervical mucus and contraception: What we know and what we don’t. Contraception 2017, 96, 310–321. [Google Scholar] [CrossRef] [PubMed]
- D’Cruz, O.J.; Haas, G.G.; Wang, B.L.; DeBault, L.E. Activation of human complement by IgG antisperm antibody and the demonstration of C3 and C5b-9-mediated immune injury to human sperm. J. Immunol. 1991, 146, 611–620. [Google Scholar] [PubMed]
- D’Cruz, O.J.; Wang, B.L.; Haas, G.G., Jr. Phagocytosis of immunoglobulin G and C3-bound human sperm by human polymorphonuclear leukocytes is not associated with the release of oxidative radicals. Biol. Reprod. 1992, 46, 721–732. [Google Scholar] [CrossRef] [PubMed]
- D’Cruz, O.J.; Haas, G.G.; Lambert, H.O. Evaluation of antisperm complement-dependent immune mediators in human ovarian follicular fluid. J. Immunol. 1990, 144, 3841–3848. [Google Scholar] [PubMed]
- Price, R.J.; Boettcher, B. The presence of complement in human cervical mucus and its possible relevance to infertility in women with complement-dependent sperm-immobilizing antibodies. Fertil. Steril. 1979, 32, 61–66. [Google Scholar] [CrossRef]
- Simopoulou, M.; Sfakianoudis, K.; Maziotis, E.; Grigoriadis, S.; Giannelou, P.; Rapani, A.; Pantos, K. The impact of autoantibodies on IVF treatment and outcome: A systematic review. Int. J. Mol. Sci. 2019, 20, 892. [Google Scholar] [CrossRef] [PubMed]
- Heng, B.C.; Huang, W.; Zhong, X.; Yin, P.; Tong, G.Q. Roles of antiphospholipid antibodies, antithyroid antibodies and antisperm antibodies in female reproductive health. Integr. Med. Int. 2015, 2, 21–31. [Google Scholar] [CrossRef]
- Robertson, S.A.; Sharkey, D.J. Seminal fluid and fertility in women. Fertil. Steril. 2016, 106, 511–519. [Google Scholar] [CrossRef]
- Berger, G.K.; Smith-Harrison, L.I.; Sandlow, J.I. Sperm agglutination: Prevalence and contributory factors. Andrologia 2019, 51, e13254. [Google Scholar] [CrossRef] [PubMed]
- Bronson, R.A. Antisperm antibodies: A critical evaluation and clinical guidelines. J. Reprod. Immunol. 2000, 45, 159–183. [Google Scholar] [CrossRef]
- Nip, M.M.; Taylor, P.V.; Rutherford, A.J.; Hancock, K.W. Autoantibodies and antisperm antibodies in sera and follicular fluids of infertile patients; relation to reproductive outcome after in-vitro fertilization. Hum. Reprod. 1995, 10, 2564–2569. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.X.; Zhu, W.J. Circulating antisperm antibody (ASA) in women is not associated with missed abortions at the first-trimester pregnancy. J. Reprod. Contracept. 2011, 22, 139–143. [Google Scholar] [CrossRef]
- Griebel, C.P.; Halvorsen, J.; Golemon, T.B.; Day, A.A. Management of spontaneous abortion. Am. Fam. Physician 2005, 72, 1243–1250. [Google Scholar] [PubMed]
- Clifford, K.; Rai, R.; Watson, H.; Regan, L. Pregnancy: An informative protocol for the investigation of recurrent miscarriage: Preliminary experience of 500 consecutive cases. Hum. Reprod. 1994, 9, 1328–1332. [Google Scholar] [CrossRef] [PubMed]
- Mestecky, J.; Russell, M.W. Induction of mucosal immune responses in the human genital tract. Pathog. Dis. 2000, 27, 351–355. [Google Scholar] [CrossRef]
- Kashir, J.; Buntwal, L.; Nomikos, M.; Calver, B.L.; Stamatiadis, P.; Ashley, P.; Vassilakopoulou, V.; Sanders, D.; Knaggs, P.; Livaniou, E.; et al. Antigen unmasking enhances visualization efficacy of the oocyte activation factor, phospholipase C zeta, in mammalian sperm. MHR Basic Sci. Reprod. Med. 2017, 23, 54–67. [Google Scholar] [CrossRef]
- Giefing-Kröll, C.; Berger, P.; Lepperdinger, G.; Grubeck-Loebenstein, B. How sex and age affect immune responses, susceptibility to infections, and response to vaccination. Aging Cell 2015, 14, 309–321. [Google Scholar] [CrossRef]
- Shetty, J.; Naaby-Hansen, S.; Shibahara, H.; Bronson, R.; Flickinger, C.J.; Herr, J.C. Human sperm proteome: Immunodominant sperm surface antigens identified with sera from infertile men and women. Biol. Reprod. 1999, 61, 61–69. [Google Scholar] [CrossRef]
- Hansen, L.K.; Becher, N.; Bastholm, S.; Glavind, J.; Ramsing, M.; Kim, C.J.; Romero, R.; Jensen, J.S.; Uldbjerg, N. The cervical mucus plug inhibits, but does not block, the passage of ascending bacteria from the vagina during pregnancy. Acta. Obstet. Gynecol. Scand. 2014, 93, 102–108. [Google Scholar] [CrossRef] [PubMed]
- Drab, T.; Ren, S.; Manaskova-Postlerova, P.; Ticha, M.; Jonákova, V.; Liberda, J. Glycosidases in porcine follicular fluid and their effect on zona pellucida-AWN 1 spermadhesin interaction. Theriogenology 2017, 100, 80–87. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.; Shen, Z.; Li, J.; He, W.; Yang, Y.; Liang, Z. Nasal immunization using a mimovirus vaccine based on the Eppin B-cell epitope induced suppressed fertility in mice. Hum. Vacc. Immunotherapeut. 2014, 10, 2227–2234. [Google Scholar] [CrossRef] [PubMed]
- Said, T.M.; Agarwal, A. Tests for Sperm Antibodies. In Immune Infertility; Springer International Publishing: Berlin/Heidelberg, Germany, 2017; pp. 197–207. [Google Scholar]
- Hafez, E.S.; Hafez, B. (Eds.) Reproduction in Farm Animals; John Wiley & Sons: Hoboken, NJ, USA, 2013. [Google Scholar]
- Yu, Q.; Mei, X.Q.; Ding, X.F.; Dong, T.T.; Dong, W.W.; Li, H.G. Construction of a catsper1 DNA vaccine and its antifertility effect on male mice. PLoS ONE 2015, 10, e0127508. [Google Scholar] [CrossRef] [PubMed]
- Naz, R.K. Recent progress toward development of vaccines against conception. Expert Rev. Vacc. 2014, 13, 145–154. [Google Scholar] [CrossRef] [PubMed]
- Baskin, M.J. Temporary sterilization by the injection of human spermatozoa. A preliminary report. Am. J. Obstetr. Gynecol. 1932, 24, 892–897. [Google Scholar] [CrossRef]
- Diekman, A.B.; Herr, J.C. Sperm antigens and their use in the development of an immunocontraceptive. Am. J. Reprod. Immunol. 1997, 37, 111–117. [Google Scholar] [CrossRef]
- Ellerman, D.A.; Pei, J.; Gupta, S.; Snell, W.J.; Myles, D.; Primakoff, P. Izumo is part of a multiprotein family whose members form large complexes on mammalian sperm. Mol. Reprod. Dev. 2009, 76, 1188–1199. [Google Scholar] [CrossRef] [PubMed]
- Inoue, N.; Ikawa, M.; Isotani, A.; Okabe, M. The immunoglobulin superfamily protein Izumo is required for sperm to fuse with eggs. Nature 2005, 434, 234–238. [Google Scholar] [CrossRef]
- Naz, R.K. Vaccine for human contraception targeting sperm Izumo protein and YLP12 dodecamer peptide. Protein Sci. 2014, 23, 857–868. [Google Scholar] [CrossRef]
- Hao, Z.; Wolkowicz, M.J.; Shetty, J.; Klotz, K.; Bolling, L.; Sen, B.; Westbrook, V.A.; Coonrod, S.; Flickinger, C.J.; Herr, J.C. SAMP32, a testis-specific, isoantigenic sperm acrosomal membrane-associated protein. Biol. Reprod. 2002, 66, 735–744. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.G.; Huang, T.H.; Xie, Q.D.; An, G. Investigation of recombinant mouse sperm protein izumo as a potential immunocontraceptive antigen. Am. J. Reprod. Immunol. 2008, 59, 225–234. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Lv, Z.; Shi, J.; Hu, Y.; Xu, C. Immunocontraceptive potential of the Ig-like domain of Izumo. Mol. Reprod. Dev. 2009, 76, 794–801. [Google Scholar] [CrossRef] [PubMed]
- Carlsson, L.; Ronquist, G.; Nilsson, B.; Larsson, A. Dominant prostasome immunogens for sperm-agglutinating autoantibodies of infertile men. J. Androl. 2004, 25, 699–705. [Google Scholar] [CrossRef] [PubMed]
- Naz, R.K. Treatment Modalities for Antisperm Antibodies-Mediated immune infertility. In Immune Infertility; Springer International Publishing: Berlin/Heidelberg, Germany, 2017; pp. 235–245. [Google Scholar]
- Eswaran, N.; Sundaram, V.A.; Rao, K.A.; Balasundaram, S.T. Simple isolation and characterization of seminal plasma extracellular vesicle and its total RNA in an academic lab. 3 Biotech 2018, 8, 139. [Google Scholar] [CrossRef] [PubMed]
- Weissenbacher, E.R.; Wirth, M.; Mylonas, I.; Ledger, W.J.; Witkin, S.S. Immunology in Reproductive Medicine. In Immunology of the Female Genital Tract; Springer: Berlin/Heidelberg, Germany, 2014; pp. 163–249. [Google Scholar]
- Kadam, A.L.; Fateh, M.; Naz, R.K. Fertilization antigen (FA-1) completely blocks human sperm binding to human zona pellucida: FA-1 antigen may be a sperm receptor for zona pellucida in humans. J. Reprod. Immunol. 1995, 29, 19–30. [Google Scholar] [CrossRef]
- Naz, R.K.; Wolf, D.P. Antibodies to sperm-specific human FA-1 inhibiti in vitro fertilization in rhesus monkeys: Development of a simian model for testing of anti-FA-1 contraceptive vaccine. J. Reprod. Immunol. 1994, 27, 111–121. [Google Scholar] [CrossRef]
- Naz, R.K. Effect of fertilization antigen (FA-1) DNA vaccine on fertility of female mice. Mol. Reprod. Dev. 2006, 73, 1473–1479. [Google Scholar] [CrossRef]
- Naz, R.K. Effect of sperm DNA vaccine on fertility of female mice. Mol. Reprod. Dev. 2006, 73, 918–928. [Google Scholar] [CrossRef]
- Wallace, E.M.; Gow, S.M.; Wu, F.C. Comparison between testosterone enanthate-induced azoospermia and oligozoospermia in a male contraceptive study. I: Plasma luteinizing hormone, follicle stimulating hormone, testosterone, estradiol, and inhibin concentrations. J. Clin. Endocrinol. Metabol. 1993, 77, 290–293. [Google Scholar]
- Chiriva-Internati, M.; Wang, Z.; Salati, E.; Bumm, K.; Barlogie, B.; Lim, S.H. Sperm protein 17 (Sp17) is a suitable target for immunotherapy of multiple myeloma. Blood 2002, 100, 961–965. [Google Scholar] [CrossRef] [PubMed]
- Diekman, A.B.; Orton, E.J.; Westbrook, V.; Klotz, K.L.; Naaby-Hansen, S.O.; Herr, J.C. Anti-sperm antibodies from infertile patients and their cognate sperm antigens: A review. Identity between SAGA-1, the H6-3C4 antigen, and CD52. Am. J. Reprod. Immunol. 2000, 43, 134–143. [Google Scholar] [CrossRef] [PubMed]
- Parry, S.; Wong, N.K.; Easton, R.L.; Panico, M.; Haslam, S.M.; Morris, H.R.; Anderson, P.; Klotz, K.L.; Herr, J.C.; Diekman, A.B.; et al. The sperm agglutination antigen-1 (SAGA-1) glycoforms of CD52 are O-glycosylated. Glycobiology 2007, 17, 1120–1126. [Google Scholar] [CrossRef] [PubMed]
- Diekman, A.B.; Westbrook-Case, V.A.; Naaby-Hansen, S.; Klotz, K.L.; Flickinger, C.J.; Herr, J.C. Biochemical characterization of sperm agglutination antigen-1, a human sperm surface antigen implicated in gamete interactions. Biol. Reprod. 1997, 57, 1136–1144. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Hale, G.; Jacobs, P.; Wood, L.; Fibbe, W.E.; Barge, R.; Novitzky, N.; Toit, C.D.; Abrahams, L.; Thomas, V.; Bunjes, D.; et al. CD52 antibodies for prevention of graft-versus-host disease and graft rejection following transplantation of allogeneic peripheral blood stem cells. Bone Marrow Transplant. 2000, 26, 69–76. [Google Scholar] [CrossRef] [PubMed]
- Albitar, M.; Do, K.A.; Johnson, M.M.; Giles, F.J.; Jilani, I.; O’Brien, S.; Cortes, J.; Thomas, D.; Rassenti, L.Z.; Kipps, T.J.; et al. Free circulating soluble CD52 as a tumor marker in chronic lymphocytic leukemia and its implication in therapy with anti-CD52 antibodies. Cancer 2004, 101, 999–1008. [Google Scholar] [CrossRef] [PubMed]
- Rowan, W.C.; Hale, G.; Tite, J.P.; Brett, S.J. Cross-linking of the CAMPATH-1 antigen (CD52) triggers activation of normal human T lymphocytes. Int. Immunol. 1995, 7, 69–77. [Google Scholar] [CrossRef]
- Naz, R.K.; Chauhan, S.C.; Trivedi, R.N. Monoclonal antibody against human sperm-specific YLP 12 peptide sequence involved in oocyte binding. Arch. Androl. 2002, 48, 169–175. [Google Scholar] [CrossRef]
- Wheat, T.E.; Goldberg, E. Sperm-specific lactate dehydrogenase C4: Antigenic structure and immunosuppression of fertility. Isozymes Curr. Top. Biol. Med. Res. 1983, 7, 113–130. [Google Scholar]
- Beyler, S.A.; Wheat, T.E.; Goldberg, E. Binding of antibodies against antigenic domains of murine lactate dehydrogenase-C4 to human and mouse spermatozoa. Biol. Reprod. 1985, 32, 1201–1210. [Google Scholar] [CrossRef]
- Burgos, C.; Maldonado, C.; de Burgos, N.M.G.; Aoki, A.; Blanco, A. Intracellular localization of the testicular and sperm-specific lactate dehydrogenase isozyme C4 in mice. Biol. Reprod. 1995, 53, 84–92. [Google Scholar] [CrossRef] [PubMed]
- O’Hern, P.A.; Bambra, C.S.; Isahakia, M.; Goldberg, E. Reversible contraception in female baboons immunized with a synthetic epitope of sperm-specific lactate dehydrogenase. Biol. Reprod. 1995, 52, 331–339. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Wheat, T.E.; Goldberg, E. Antigenic domains of the sperm-specific lactate dehydrogenase C4, isozyme. Mol. Immunol. 1985, 22, 643–649. [Google Scholar] [CrossRef]
- Mahi-Brown, C.A.; Vandevoort, C.A.; Mcguinness, R.P.; Overstreet, J.W.; O’hern, P.A.; Goldberg, E. Immunization of male but not female mice with the sperm-specific isozyme of lactate dehydrogenase (LDH-C4) impairs fertilization in vivo. Am. J. Reprod. Immunol. 1990, 24, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Kille, J.W.; Goldberg, E. Inhibition of oviducal sperm transport in rabbits immunized against sperm-specific lactate dehydrogenase (LDH-C4). J. Reprod. Immunol. 1980, 2, 15–21. [Google Scholar] [CrossRef]
- Li, S.S.; Feldmann, R.J.; Okabe, M.; Pan, Y.C. Molecular features and immunological properties of lactate dehydrogenase C4 isozymes from mouse and rat testes. J. Biol. Chem. 1983, 258, 7017–7028. [Google Scholar] [PubMed]
- Goldberg, E.; Wheat, T.E.; Powell, J.E.; Stevens, V.C. Reduction of fertility in female baboons immunized with lactate dehydrogenase C 4. Fertil. Steril. 1981, 35, 214–217. [Google Scholar] [CrossRef]
- Ohl, D.A.; Naz, R.K. Infertility due to antisperm antibodies. Urology 1995, 46, 591–601. [Google Scholar] [CrossRef]
- Millar, S.E.; Chamow, S.M.; Baur, A.W.; Oliver, C.; Robey, F.; Dean, J. Vaccination with a synthetic zona pellucida peptide produces long-term contraception in female mice. Science 1989, 246, 935–939. [Google Scholar] [CrossRef] [PubMed]
- Cohen, J.; Malter, H.; Fehilly, C.; Wright, G.; Elsner, C.; Kort, H.; Massey, J. Implantation of embryos after partial opening of oocyte zona pellucida to facilitate sperm penetration. Lancet 1988, 332, 162. [Google Scholar] [CrossRef]
- Mask, T.A.; Schoenecker, K.A.; Kane, A.J.; Ransom, J.L.; Bruemmer, J.E. Serum antibody immunoreactivity to equine zona protein after SpayVac vaccination. Theriogenology 2015, 84, 261–267. [Google Scholar] [CrossRef] [PubMed]
- Kirkpatrick, J.F.; Turner, J.W., Jr.; Liu, I.K.; Fayrer-Hosken, R. Applications of pig zona pellucida immunocontraception to wildlife fertility control. J. Reprod. Fertil. Suppl. 1996, 50, 183–189. [Google Scholar] [PubMed]
- Lefievre, L.; Conner, S.J.; Salpekar, A.; Olufowobi, O.; Ashton, P.; Pavlovic, B.; Lenton, W.; Afnan, M.; Brewis, I.A.; Monk, M.; et al. Four zona pellucida glycoproteins are expressed in the human. Hum. Reprod. 2004, 19, 1580–1586. [Google Scholar] [CrossRef] [PubMed]
- Prasad, S.V.; Wilkins, B.; Skinner, S.M.; Dunbar, B.S. Evaluating zona pellucida structure and function using antibodies to rabbit 55 kDa ZP protein expressed in baculovirus expression system. Mol. Reprod. Dev. 1996, 43, 519–529. [Google Scholar] [CrossRef]
- Chakravarty, S.; Suraj, K.; Gupta, S.K. Baculovirus-expressed recombinant human zona pellucida glycoprotein-B induces acrosomal exocytosis in capacitated spermatozoa in addition to zona pellucida glycoprotein-C. Mol. Hum. Reprod. 2005, 11, 365–372. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Wang, Y.; Zhang, B.; Li, J.; Aipire, A.; Li, Y.; Zhang, F. Enhanced contraception of canine zona pellucida 3 DNA vaccine via targeting DEC-205 in mice. Theriogenology 2018, 113, 56–62. [Google Scholar] [CrossRef] [PubMed]
- Joonè, C.J.; Schulman, M.L.; Bertschinger, H.J. Ovarian dysfunction associated with zona pellucida-based immunocontraceptive vaccines. Theriogenology 2017, 89, 329–337. [Google Scholar] [CrossRef] [PubMed]
- Gupta, S.K.; Srivastava, N.; Choudhury, S.; Rath, A.; Sivapurapu, N.; Gahlay, G.K.; Batra, D. Update on zona pellucida glycoproteins based contraceptive vaccine. J. Reprod. Immunol. 2004, 62, 79–89. [Google Scholar] [CrossRef] [PubMed]
- Bronson, R.; Cooper, G.; Rosenfeld, D.; Witkin, S.S. Detection of spontaneously occurring sperm-directed antibodies in infertile couples by immunobead binding and enzyme-linked immunosorbent assay. Ann. N. Y. Acad. Sci. 1984, 438, 504–507. [Google Scholar] [CrossRef] [PubMed]
- Rose, N.R.; Hjort, T.; Rumke, P.; Harper, M.J.; Vyazov, O. Techniques for detection of iso-and auto-antibodies to human spermatozoa. Clin. Expt. Immunol. 1976, 23, 175–199. [Google Scholar]
- Shulman, S.; Harlin, B.; Davis, P.; Reyniak, J.V. Immune infertility and new approaches to treatment. Fertil. Steril. 1978, 29, 309–313. [Google Scholar] [CrossRef]
- Ulcova-Gallova, Z.; Mraz, L.; Planickova, E.; Macku, F.; Ulc, I. Three years experience with local hydrocortisone treatment in women with immunological cause of infertility. Zentralblatt fur Gynakologie 1990, 12, 867–872. [Google Scholar]
- Aghamir, S.M.; Salavati, A.; Yousefie, R.; Tootian, Z.; Ghazaleh, N.; Jamali, M.; Azimi, P. Does bone marrow-derived mesenchymal stem cell transfusion prevent antisperm antibody production after traumatic testis rupture? Urology 2014, 84, 82–86. [Google Scholar] [CrossRef] [PubMed]
- Krapez, J.A.; Hayden, C.J.; Rutherford, A.J.; Balen, A.H. Survey of the diagnosis and management of antisperm antibodies. Hum. Reprod. (Oxf. Engl.) 1998, 13, 3363–3367. [Google Scholar] [CrossRef] [PubMed]
- Galle, P.C.; McRae, M.A.; Colliver, J.A.; Alexander, J.S. Sperm washing and intrauterine insemination for cervical factor, oligospermia, immunologic infertility and unexplained infertility. J. Reprod. Med. 1990, 35, 116–122. [Google Scholar] [PubMed]
- Agarwal, A. Treatment of immunological infertility by sperm washing and intrauterine insemination. Arch. Androl. 1992, 29, 207–213. [Google Scholar] [CrossRef] [PubMed]
- Clarke, G.N.; Bourne, H.; Baker, H.G. Intracytoplasmic sperm injection for treating infertility associated with sperm autoimmunity. Fertil. Steril. 1997, 68, 112–117. [Google Scholar] [CrossRef]
- Olson, C.K.; Keppler-Noreuil, K.M.; Romitti, P.A.; Budelier, W.T.; Ryan, G.; Sparks, A.E.; Van Voorhis, B.J. In vitro fertilization is associated with an increase in major birth defects. Fertil. Steril. 2005, 84, 1308–1315. [Google Scholar] [CrossRef] [PubMed]
- Georgiou, I.; Syrrou, M.; Pardalidis, N.; Karakitsios, K.; Mantzavinos, T.; Giotitsas, N.; Loutradis, D.; Dimitriadis, F.; Saito, M.; Miyagawa, I.; et al. Genetic and epigenetic risks of intracytoplasmic sperm injection method. Asian J. Androl. 2006, 8, 643–673. [Google Scholar] [CrossRef]
- Comhaire, F.H. Male contraception: Hormonal, mechanical and other. Hum. Reprod. 1994, 9, 586–590. [Google Scholar] [CrossRef]
- O’Donnell, L.; Narula, A.; Balourdos, G.; Gu, Y.Q.; Wreford, N.G.; Robertson, D.M.; Bremner, W.J.; McLachlan, R.I. Impairment of spermatogonial development and spermiation after testosterone-induced gonadotropin suppression in adult monkeys (Macaca fascicularis). J. Clin. Endocrinol. Metab. 2001, 86, 1814–1822. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Wang, C.; Swerdloff, R.S. Male hormonal contraception. Am. J. Obstet. Gynaecol. 2004, 190, S60–S68. [Google Scholar] [CrossRef] [PubMed]
- Nieschlag, E. Clinical trials in male hormonal contraception. Contraception 2010, 82, 457–470. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Amory, J.K.; Page, S.T.; Bremner, W.J. Drug insight: Recent advances in male hormonal contraception. Nat. Rev. Endocrinol. 2006, 2, 32–41. [Google Scholar] [CrossRef]
- Talwar, G.P.; Singh, O.M.; Gupta, S.K.; Hasnain, S.E.; Pal, R.; Majumdar, S.S.; Vrati, S.; Mukhopadhay, A.; Srinivasan, J.; Deshmukh, U.; et al. The HSD-hCG vaccine prevents pregnancy in women: Feasibility study of a reversible safe contraceptive vaccine. Am. J. Reprod. Immunol. 1997, 37, 153–160. [Google Scholar] [CrossRef] [PubMed]
- O’Rand, M.G.; Wedgren, E.E. Sperm Antigen Corresponding to a Sperm Autoantigenic Epitope and Methods of Using the Same. Patent US5175148, 29 December 1992. [Google Scholar]
- Kitto, B.G.; Hirschhorn, D.C. DNA Immunocontraceptive Vaccines and Uses Thereof. Patent WO2005093043A1, 1 September 2005. [Google Scholar]
- O’Rand, M.G.; Wedgren, E.E.; Richardson, R.T.; Lea, A.I. Sperm Antigen Corresponding to a Sperm Zona Binding Protein Autoantigenic Epitope. Patent US5480799 A, 2 January 1996. [Google Scholar]
- O’Neill, C. Fertility Control. Patent US4879285, 7 November 1989. [Google Scholar]
- Donahoe, P. Use of Mullerian Inhibiting Substance (MIS) as a Contraceptive Agent. Patent US4753794, 28 June 1988. [Google Scholar]
- Stevens, V.C. Antigenic Modification of Polypeptides. Patent US4713366A, 15 December 1987. [Google Scholar]
- John, C.H.; Friederike, J.; Arabinda, M.; Jagathpala, S.; Michael, J.W. Human Sperm Specific Lysozyme-Like Proteins. Patent US7125550B2, 24 October 2006. [Google Scholar]
- Robert, B.; Bill, P.; Janet, H.; McLaren, L. Compositions and Methods for Reducing or Preventing Fertilization in Fish and Birds. Patent US6790457B1, 14 September 2004. [Google Scholar]
- Richard, A.C.; Kevin, J.W. Topical Application of Antibodies for Contraception and for Prophylaxis Against Sexually Transmitted Diseases. Patent US6355235B1, 12 March 2002. [Google Scholar]
- Robert, B.; Michael, M.; Bill, P.; Warwick, C.K. Method to Prevent Fertilization in Mammals by Administering a Single Dose of Zona Pellucida Derived Antigens, Liposome and Freund’s Adjuvant. Patent US5736141A, 7 April 1998. [Google Scholar]
- Jerry, J.R.; Kevin, P.B.; Yuzhi, Z. Chimeric Contraceptive Vaccines. Patent US6045799A, 11 January 2000. [Google Scholar]
- Robert, G.B.; Marc, M.; Bill, P. Antigens for Immunocontraception. Patent US7056515B2, 6 June 2006. [Google Scholar]
- Wu, X.; Rupprecht, C. Rabies Virus-Based Recombinant Immunocontraceptive Compositions and Methods of Use. Patent US8524247B2, 3 September 2013. [Google Scholar]
- Kirk, D.; Mason, H.; Walmsley, A.; Arntzen, C. Methods and Compositions for Stable Transgenic Plant Pharmaceuticals and Their Use as Contraceptives. Patent CA 2441699, 9 September 2004. [Google Scholar]
- Saenziturriaga, L.E. Protein for the Immunocastration for Mammals. U.S. Patent US 8940693, 27 January 2015. [Google Scholar]
- Babiak, I.; Hole, R. Methods of Inhibiting Gonad Maturation. Patent US 2015/0104474 A1, 3 April 2018. [Google Scholar]
- McNamara, M.K. Immunogenic LHRH Compositions and Methods Relating Thereto. U.S. Patent US8741303, 3 June 2014. [Google Scholar]
- Min, W. Intra Testicular Injection of Immunogens. Patent US 2014/0271716 A1, 18 September 2014. [Google Scholar]
- Hay, B.A.; Li, J. Antibody-Mediated Immunocontraception. Patent US20140223591A1, 7 August 2014. [Google Scholar]
- Myroslavivna, H.A.; Volodymyrivna, C.V.; Andriiovych, N.Y.; Yosypivna, K.L.; Maciej, K. Method for the Prediction of Immune Dependent Male Infertility Development in Patients after Varicocelectomy. Patent UA102086U, 6 June 2015. [Google Scholar]
- Qingdao Huaren Tech Incubator. Medicine for Treating Immune Male Infertility. Patent CN105079573A, 19 June 2015.
- Hong, C. Pharmaceutical Composition for Curing Immune Infertility. Patent CN105560668 (A), 30 August 2016. [Google Scholar]
- Jun, F.; Wei, S.; Linfang, W.; Shiying, Y.; Yong, W.; Xiaodong, Z.; Rongyan, Y.; Yanyun, L. Detection Methods and Kits of Anti-ACTL7a Antibody and Anti-GAPDH-2 Antibody, and Uses of Kits. Patent CN106153938A, 8 July 2016. [Google Scholar]
- Yue, C. Albizzia julibrissin and Rhizoma cyperi pill. Patent CN107050378(A), 25 August 2017. [Google Scholar]
- Juzuo, Z.; Xiujun, F.; Jian, Z.; Zhilong, C.; Baozhen, W.; Chen, H.; Jie, C.; Mengxia, L. Placental Chondroitin Sulphate an Immunogenic Composition and Application. Patent WO2019075893A1, 22 September 2019. [Google Scholar]
- Jinming, W. Multi-Functional Beverage Containing Peptides. Patent CN109363037 (A), 30 June 2019. [Google Scholar]
- Zhang, Y.; Zhangyun Wei, L.; Yongtang, Z.; Wenhui, L. Macroweb Bombinator Antisperm Polypeptide and Its Preparation Process and Use in Medicine. Patent CN1142182C, 3 July 2004. [Google Scholar]
- Teitelbaum, R. Biological Female Contraceptives. Patent ES2671345T3, 6 June 2018. [Google Scholar]
- Rachel, T. Biological Female Contraceptives. Patent JP2019017389A, 10 September 2019. [Google Scholar]
- Tetelbaum, R. Biological Female Contraceptives. Patent KR101927747B1, 4 May 2013. [Google Scholar]
- Bradner, J.E.; Matzuk, M.; Qi, J. Male Contraceptive Compositions and Methods of Use. Patent JP6022442B2, 5 April 2016. [Google Scholar]
- Hamilton, D.W.; Roberts, K.P.; Ensrud, K.M. CRISP Polypeptides as Contraceptives and Inhibitors of Sperm Capacitation. Patent US7405202B2, 29 July 2008. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
| Title of the Patent | Patent Number | Details on the Invention | Inventors | Reference |
|---|---|---|---|---|
| Sperm antigen corresponding to a sperm autoantigenic epitope and methods of using the same | US5175148 | Usage of rabbit sperm membrane autoantigen for detecting auto immune infertility and as an immune contraceptive agent. | Michael G. O’Rand, Esther E. Widgren | O’Rand and Widgren, 1992 [232] |
| DNA immunocontraceptive vaccines and uses thereof | WO2005093043A1 | Recombinant bacteria possessing sperm specific protein-lactate dehydrogenase C (LDH C). On administration, antibodies will be produced against LDH C leading to less sperm in males and immunocontraceptive in females. | Barrie G. Kitto, Daniel C. Hirschhorn | Kitto and Hirschhorn, 2005 [233] |
| Sperm antigen corresponding to a sperm zona binding protein autoantigenic epitope | US5480799 A | Human seminal plasma 17 protein and its antigenic fragments were used as an immunocontraceptive. | Michael G. O’Rand, Esther E. Widgren, Richard T. Richardson, Isabel A. Lea | O’Rand et al., 1996 [234] |
| Fertility control | US4879285 | Platelet activating factor (PAF) is an enhancer of embryo viability. PAF antibody in conjugation with estrogen was proposed to have a contraceptive effect. | Christopher O’Neill | O’Neill, 1989 [235] |
| Use of mullerian inhibiting substance (MIS) as a contraceptive agent | US4753794 | MIS interferes in the maturation of oocytes and thus prevents formation of the embryo. | Patricia Donahoe | Donahoe, 1988 [236] |
| Antigenic modification of polypeptides | US4713366A | Both endogenous and exogenous proteins are modified to become more antigenic so as to produce antibodies against host components, including sperm. | Vernon C. Stevens | Stevens, 1987 [237] |
| Human sperm specific lysozyme-like proteins | US7125550B2 | Testis specific proteins C 19 and C 23 were modified to be used as contraceptive agents. | John C. Herr, Friederike Jayes, Arabinda Mandal, Jagathpala Shetty, Michael J. Wolkowicz | Herr et al., 2006 [238] |
| Compositions and methods for reducing or preventing fertilization in fish and birds | US6790457B1 | Immunocontraceptive vaccine was developed using a homolog of the Zona pellucid protein along with a carrier. | Robert Brown, Bill Pohajdak, Janet Horrocks, Leslie McLaren | Brown et al., 2004 [239] |
| Topical application of antibodies for contraception and for prophylaxis against sexually transmitted diseases | US6355235B1 | Antibody molecules capable of immobilizing sperm can be used in men as a topical application, and in women, these antibodies were made to be released using intrauterine/intravaginal devices. | Richard A. Cone, Kevin J. Whaley | Coneand Whaley, 2002 [240] |
| Method to prevent fertilization in mammals by administering a single dose of zona-pellucida-derived antigens, liposome, and Freund’s adjuvant | US5736141A | A liposome mediated delivery of zona pellucida antigens through vaccination was reported to produce antibodies detected for 22 months. | Robert Brown, Michael Mezei, Bill Pohajdak, Warwick Charles Kimmins | Brown et al., 1998 [241] |
| Chimeric contraceptive vaccines | US6045799A | Carrier protein related with a protein of reproductive function was used. | Jerry J. Reeves, Kevin P. Bertrand, Yuzhi Zhang | Reeves et al., 2000 [242] |
| Antigens for immunocontraception | US7056515B2 | Immunocontraceptive vaccines were produced using zona pellucida protein fragments from carnivorous animals. | Robert George Brown, Marc Mansour, Bill Pohajdak | Brown et al., 2006 [243] |
| Rabies-virus-based recombinant immunocontraceptive compositions and methods of use | US8524247B2 | Nucleic acid information of an immunocontraceptive protein in a recombinant rabies virus was proposed to inhibit fertility. | Xianfu Wu, Charles Rupprecht | Wuand Rupprecht, 2013 [244] |
| Methods and compositions for stable transgenic plant pharmaceuticals and their use as contraceptives | CA2441699A1 | Pharmaceutical proteins capable of acting as an immunocontraceptive were proposed to be produced in transgenic plants. | Dwayne Kirk, Hugh Mason, Amanda Walmsley, Charles Arntzen | Kirk et al., 2004 [245] |
| Protein for the immunocastration of mammals | US8940693B2 | A fusion protein for immunocastration was used in this method. | Leonardo Enrique Saenziturriaga | Saenziturriaga, 2015 [246] |
| Methods of inhibiting gonad Maturation | US 2015/0104474 A1 | Nucleic acid molecules possessing compounds capable of eliciting immune response against gonad maturation in juvenile animals was developed | Igor Babiak, Reid Hole | Babiak and Hole, 2018 [247] |
| Immunogenic LHRH (synthetic luteinizing hormone releasing hormone) compositions and methods relating thereto | US8741303B2 | Fertility potential was modified using LHRH with five amino acids attached to a C terminal. | Michael Kerin McNamara | McNamara, 2014 [248] |
| Intratesticular injection of immunogens | US 2014/0271716 A1 | Chemical sterilization agent along with immunogen against disease pathogens when introduced in testis tissue can bring effective contraception. | Min Wang | Wang, 2014 [249] |
| Antibody-mediated immunocontraception | US20140223591A1 | Methods and compositions for contraception (including vector-based approaches) and manipulation or other reproduction-associated traits. | Bruce A. Hay, Juan Li | Hay and Li, 2014 [250] |
| Method for the prediction of immune-dependent male infertility development in patients after varicocelectomy | UA102086U | Laboratory-based methods and standards for determining the probability of male infertility out of immune reaction in patients having undergone a varicocelectomy. | Myroslavivna H.A., Volodymyrivna, C.V., Andriiovych, N.Y., Yosypivna, K.I., Maciej, K. | Myroslavivna et al., 2015 [251] |
| Medicine for treating immune male infertility | CN105079573A | A polyherbal preparation against infertility by improving immune functions and eliminating hypersensitivity and autoimmune responses. | Qingdao Huaren Tech Incubator | Qingdao Huaren Tech Incubator, 2015 [252] |
| Pharmaceutical composition for curing immune infertility | CN105560668A | Involves traditional Chinese medicine using a polyherbal combination for curing immune infertility. | Chen Hong | Hong, 2016 [253] |
| Detection methods and kits of anti-ACTL7a antibody and anti-GAPDH-2 (glyceraldehyde-3-phosphate dehydrogenase) antibody, and uses of kits | CN106153938A | This involves the diagnosis of immunological infertility through the detection of antifertility antibodies against ACTL7a (Actin-like protein 7A) and GAPDH-2 using kits provided with a purified ACTL7a antigen-coated plate and/or a purified GAPDH-2-antigen-coated plate. | Fu Jun, Song Wei, Wang Linfang, Yan Shiying, Wang Yong, Zhang Xiaodong, Yao Rongyan, Luo Yanyun. | Jun et al., 2016 [254] |
| Albizzia julibrissin and Rhizoma cyperi pill | CN107050378(A) | Involves the preparation of a watered pill prepared out of 19 Chinese traditional medicines with a high efficiency in treating positive antisperm antibodies. | Cui Yue | Yue, 2017 [255] |
| Placental chondroitin sulphate an immunogenic composition and application | WO2019075893A1 | A contraceptive drug formulation for humans and other animals using placental chondroitin sulphate A to prevent implantation of the embryo onto the endometrium. Also supplied with an immunopotentiator thatcan terminatepregnancy. | Zhang Juzuo, Fan Xiujun, Zhang Jian, Chen Zhilong, Wang Baozhen, Huang Chen, Chen Jie, Li Mengxia | Juzuo, 2019 [256] |
| Multi-functional beverage containing peptides | CN109363037 (A) | This is a novel formulation of peptides from various sources with multiple health benefits to male subjects including immune boosting and being effective against infertility. | Wang Jinming | Jinming, 2019 [257] |
| Macro web bombinator antisperm polypeptide and its preparation process and use in medicine | CN1142182C | A polypeptide prepared out of novel process possessing sperm-brake activity and the peptide can be therapeutically applicable for contraception and against pancreatitis. | Yun Zhang, Zhangyun, Lai Wei, Zheng Yongtang, Li Wenhui | Zhang et al., 2004 [258] |
| Biological female contraceptives | ES2671345T3 | Commensal Lactobacillus bacterium of the female genital tract has been genetically modified to express antibodies with an anti-sperm property so as to be used as an effective biological anti-fertilizing agent. | Rachel Teitelbaum | Teitelbaum, 2018 [259] |
| Biologic female contraceptives | JP2019017389A | Commensal microbial organisms genetically engineered to carry fragments of anti-sperm activity and applied for preventing fertilization. | Teitelbaum Rachel | Rachel, 2019 [260] |
| Biologic female contraceptives | KR101927747B1 | Symbiotic organism genetically modified to provide effective contraception in females. | Rachel Tetelbaum | Tetelbaum, 2013 [261] |
| Male contraceptive compositions and methods of use | JP6022442B2 | It describes the novel methods and compositions for developing compounds based on inhibitors of the bromodomain testis-specific protein (BRDt) for effective contraception in male subjects. | Bradner, James Elliott, Martin Matzuk, Jun Qi | Bradner et al., 2016 [262] |
| CRISP (Cysteine-rich secretory protein) polypeptides as contraceptives and inhibitors of sperm capacitation | US7405202B2 | It describes the usage of a CRISP polypeptide for contraception through inhibiting fertilization events in sperm such as capacitation and acrosomal reactions. | David W. Hamilton, Kenneth P. Roberts, Kathy M. Ensrud | Hamilton et al., 2008 [263] |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
A. S., V.; Dhama, K.; Chakraborty, S.; Abdul Samad, H.; K. Latheef, S.; Sharun, K.; Khurana, S.K.; K., A.; Tiwari, R.; Bhatt, P.; et al. Role of Antisperm Antibodies in Infertility, Pregnancy, and Potential for Contraceptive and Antifertility Vaccine Designs: Research Progress and Pioneering Vision. Vaccines 2019, 7, 116. https://doi.org/10.3390/vaccines7030116
A. S. V, Dhama K, Chakraborty S, Abdul Samad H, K. Latheef S, Sharun K, Khurana SK, K. A, Tiwari R, Bhatt P, et al. Role of Antisperm Antibodies in Infertility, Pregnancy, and Potential for Contraceptive and Antifertility Vaccine Designs: Research Progress and Pioneering Vision. Vaccines. 2019; 7(3):116. https://doi.org/10.3390/vaccines7030116
Chicago/Turabian StyleA. S., Vickram, Kuldeep Dhama, Sandip Chakraborty, Hari Abdul Samad, Shyma K. Latheef, Khan Sharun, Sandip Kumar Khurana, Archana K., Ruchi Tiwari, Prakash Bhatt, and et al. 2019. "Role of Antisperm Antibodies in Infertility, Pregnancy, and Potential for Contraceptive and Antifertility Vaccine Designs: Research Progress and Pioneering Vision" Vaccines 7, no. 3: 116. https://doi.org/10.3390/vaccines7030116
APA StyleA. S., V., Dhama, K., Chakraborty, S., Abdul Samad, H., K. Latheef, S., Sharun, K., Khurana, S. K., K., A., Tiwari, R., Bhatt, P., K., V., & Chaicumpa, W. (2019). Role of Antisperm Antibodies in Infertility, Pregnancy, and Potential for Contraceptive and Antifertility Vaccine Designs: Research Progress and Pioneering Vision. Vaccines, 7(3), 116. https://doi.org/10.3390/vaccines7030116