Efforts to Improve the Seasonal Influenza Vaccine


{kind=link}
{kind=link}
Abstract
:1. Introduction
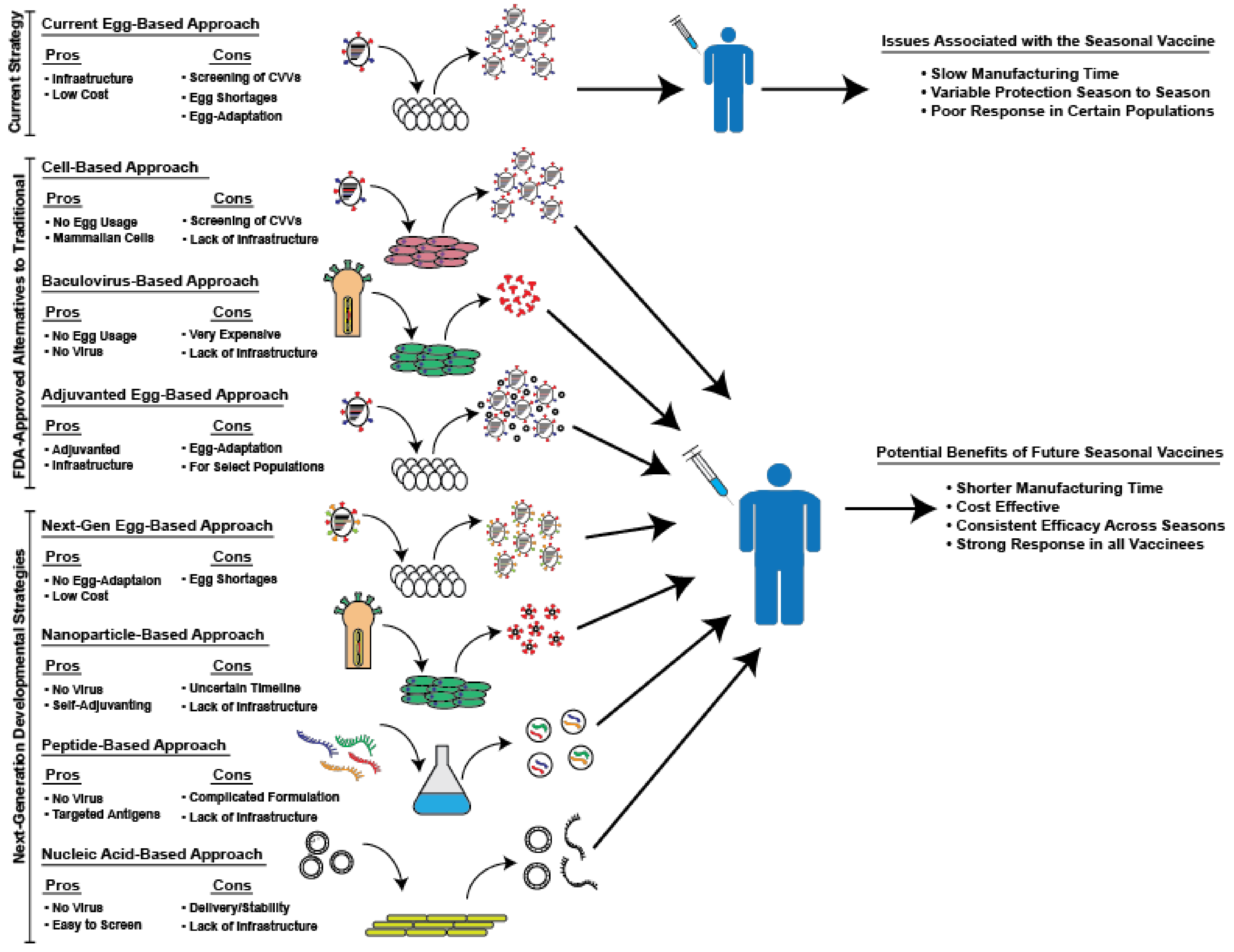
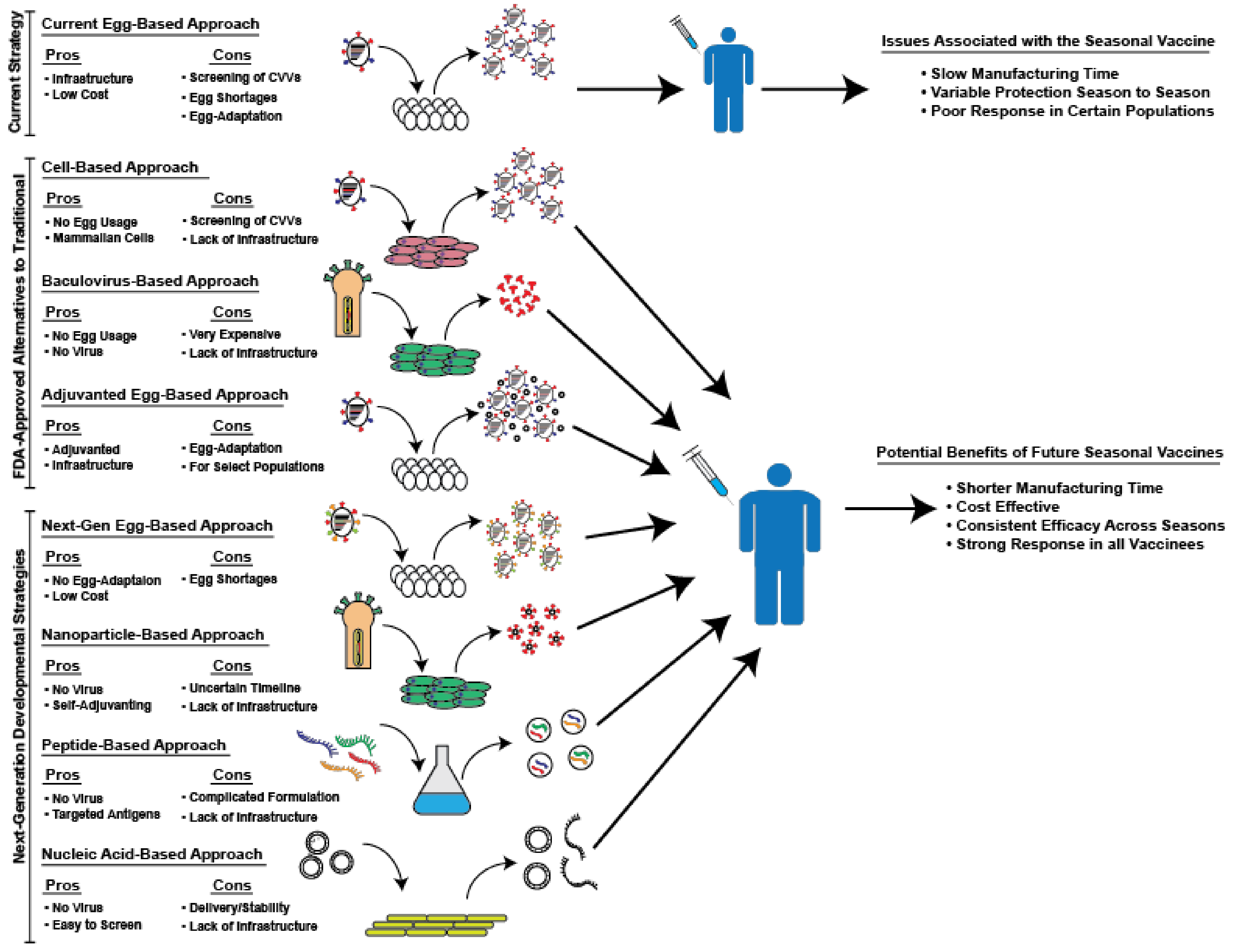
2. Currently Available, FDA-Approved Alternatives to Traditional Egg-Based Vaccines
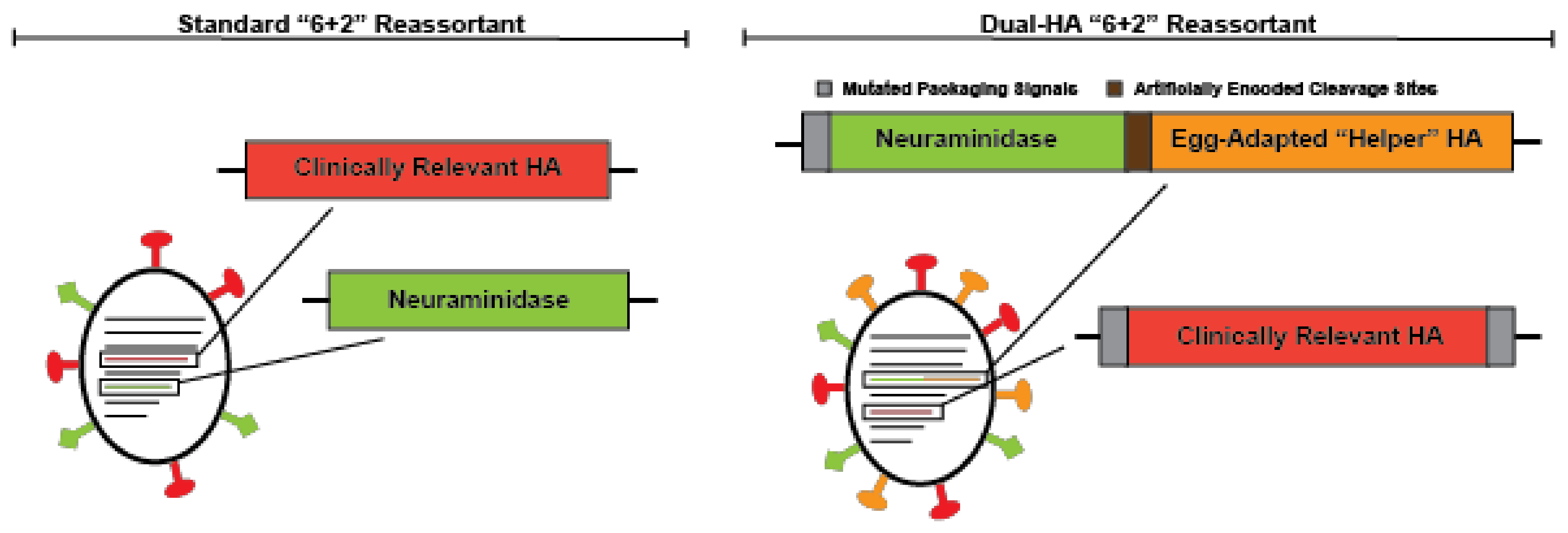
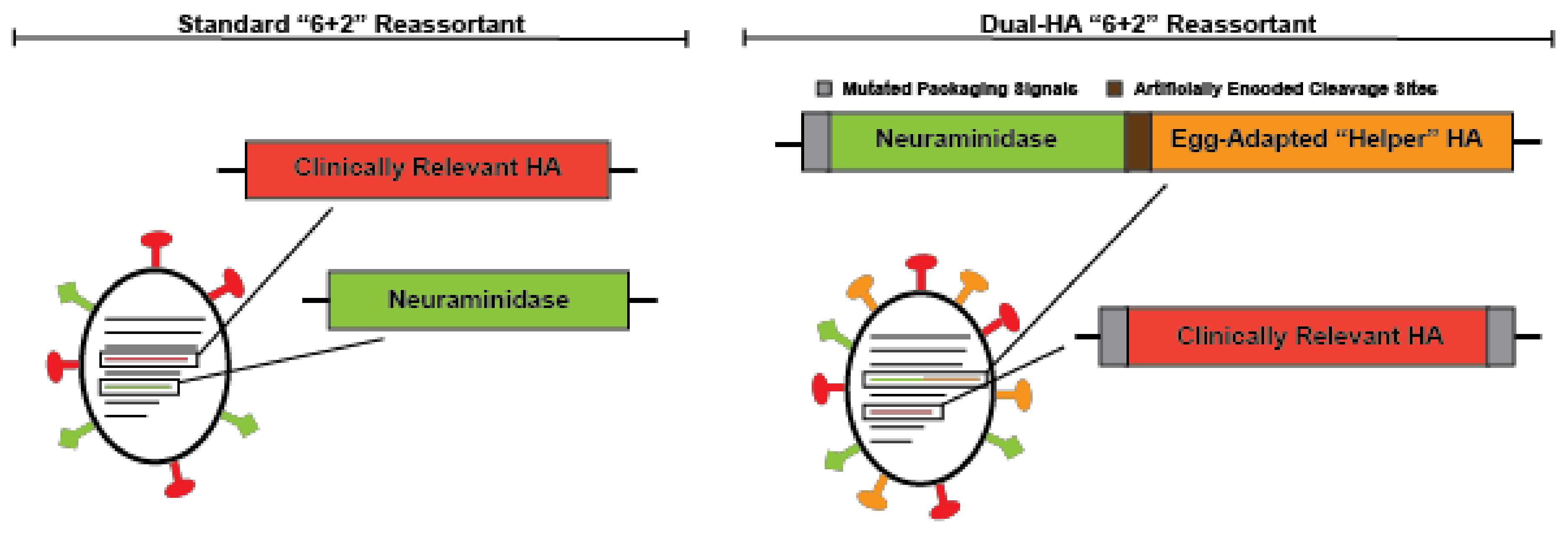
3. Next-Generation Seasonal Influenza Vaccines Currently in Development
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Shaw, M.L.; Palese, P. Orthomyxoviruses. In Fields Virology; Knipe, D.M., Howley, P.M., Eds.; Lippincott Williams and Wilkins: Philadelphia, PA, USA, 2013; pp. 1151–1185. [Google Scholar]
- World Health Organization (WHO). Influenza (Seasonal). Available online: http://www.who.int/mediacentre/factsheets/fs211/en/ (accessed on 12 February 2018).
- CDC. Key Facts about Seasonal Flu Vaccine. Available online: https://www.cdc.gov/flu/protect/keyfacts.htm (accessed on 12 February 2018).
- CDC. Quadrivalent Influenza Vaccine. Available online: https://www.cdc.gov/flu/protect/vaccine/quadrivalent.htm (accessed on 12 February 2018).
- CDC. Selecting Viruses for the Seasonal Influenza Vaccine. Available online: https://www.cdc.gov/flu/about/season/vaccine-selection.htm (accessed on 12 February 2018).
- De Jong, J.C.; Rimmelzwaan, G.F.; Fouchier, R.A.; Osterhaus, A.D. Influenza virus: A master of metamorphosis. J. Infect. 2000, 40, 218–228. [Google Scholar] [CrossRef] [PubMed]
- Steinhauer, D.A.; Holland, J.J. Rapid evolution of rna viruses. Annu. Rev. Microbiol. 1987, 41, 409–433. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.; Webster, R.G.; Webby, R.J. Influenza virus: Dealing with a drifting and shifting pathogen. Viral Immunol. 2018. [Google Scholar] [CrossRef] [PubMed]
- Shapshak, P.; Chiappelli, F.; Somboonwit, C.; Sinnott, J. The influenza pandemic of 2009: Lessons and implications. Mol. Diagn. Ther. 2011, 15, 63–81. [Google Scholar] [CrossRef] [PubMed]
- CDC. How Influenza (Flu) Vaccines Are Made. Available online: https://www.cdc.gov/flu/protect/vaccine/how-fluvaccine-made.htm (accessed on 12 February 2018).
- Pasteur, S. Sanofi pasteur: Fluzone quadrivalent (influenza vaccine) package insert. In Sanofi Pasteur; Sanofi Pasteur: Swiftwater, PA, USA, 2017; pp. 1–37. [Google Scholar]
- GlaxoSmithKline. Gsk: Fluarix quadrivalent influenza vaccine package insert. In GlaxoSmithKline; GlaxoSmithKline: Research Triangl Park, NC, USA, 2017; pp. 1–21. [Google Scholar]
- Seqirus. Seqirus: Afluria quadrivalent influenza vaccine package insert. In Seqirus; Seqirus: Summit, NJ, USA, 2017; pp. 1–23. [Google Scholar]
- Wong, S.S.; Webby, R.J. Traditional and new influenza vaccines. Clin. Microbiol. Rev. 2013, 26, 476–492. [Google Scholar] [CrossRef] [PubMed]
- Weir, J.P. Food and Drug Administration Supplement Approval. 2016. Available online: https://www.fda.gov/downloads/BiologicsBloodVaccines/Vaccines/ApprovedProducts/UCM522280.pdf (accessed on 12 February 2018).
- Couch, R.B. Seasonal inactivated influenza virus vaccines. Vaccine 2008, 26 (Suppl. 4), D5–D9. [Google Scholar] [CrossRef] [PubMed]
- Tate, M.D.; Job, E.R.; Deng, Y.M.; Gunalan, V.; Maurer-Stroh, S.; Reading, P.C. Playing hide and seek: How glycosylation of the influenza virus hemagglutinin can modulate the immune response to infection. Viruses 2014, 6, 1294–1316. [Google Scholar] [CrossRef] [PubMed]
- Krause, J.C.; Crowe, J.E., Jr. Committing the oldest sins in the newest kind of ways-antibodies targeting the influenza virus type a hemagglutinin globular head. Microbiol. Spectr. 2014, 2. [Google Scholar] [CrossRef] [PubMed]
- CDC. Seasonal Influenza Vaccine Supply & Distribution. Available online: https://www.cdc.gov/flu/about/qa/index.htm (accessed on 12 February 2018).
- McLean, K.A.; Goldin, S.; Nannei, C.; Sparrow, E.; Torelli, G. The 2015 global production capacity of seasonal and pandemic influenza vaccine. Vaccine 2016, 34, 5410–5413. [Google Scholar] [CrossRef] [PubMed]
- NCIRD. CDC Vaccine Price List; NCIRD: Atlanta, GA, USA, 2018.
- Ping, J.; Lopes, T.J.; Nidom, C.A.; Ghedin, E.; Macken, C.A.; Fitch, A.; Imai, M.; Maher, E.A.; Neumann, G.; Kawaoka, Y. Development of high-yield influenza a virus vaccine viruses. Nat. Commun. 2015, 6, 8148. [Google Scholar] [CrossRef] [PubMed]
- Ping, J.; Lopes, T.J.; Neumann, G.; Kawaoka, Y. Development of high-yield influenza B virus vaccine viruses. Proc. Natl. Acad. Sci. USA 2016, 113, E8296–E8305. [Google Scholar] [CrossRef] [PubMed]
- Stohr, K.; Bucher, D.; Colgate, T.; Wood, J. Influenza virus surveillance, vaccine strain selection and manufacture. Methods Mol. Biol. 2012, 865, 147–162. [Google Scholar] [PubMed]
- Skowronski, D.M.; Janjua, N.Z.; De Serres, G.; Sabaiduc, S.; Eshaghi, A.; Dickinson, J.A.; Fonseca, K.; Winter, A.L.; Gubbay, J.B.; Krajden, M.; et al. Low 2012–2013 influenza vaccine effectiveness associated with mutation in the egg-adapted h3n2 vaccine strain not antigenic drift in circulating viruses. PLoS ONE 2014, 9, e92153. [Google Scholar] [CrossRef] [PubMed]
- Mochalova, L.; Gambaryan, A.; Romanova, J.; Tuzikov, A.; Chinarev, A.; Katinger, D.; Katinger, H.; Egorov, A.; Bovin, N. Receptor-binding properties of modern human influenza viruses primarily isolated in vero and mdck cells and chicken embryonated eggs. Virology 2003, 313, 473–480. [Google Scholar] [CrossRef]
- World Health Organization (WHO). Addendum to the Recommended Composition of Influenza Virus Vaccines for Use in the 2003–2004 Influenza Season; WHO: Geneva, Switzerland, 2003; p. 77. [Google Scholar]
- CDC. 2003–2004 U.S. Influenza Season Summary. Available online: https://www.cdc.gov/flu/weekly/weeklyarchives2003-2004/03-04summary.htm (accessed on 12 February 2018).
- Zost, S.J.; Parkhouse, K.; Gumina, M.E.; Kim, K.; Diaz Perez, S.; Wilson, P.C.; Treanor, J.J.; Sant, A.J.; Cobey, S.; Hensley, S.E. Contemporary H3N2 influenza viruses have a glycosylation site that alters binding of antibodies elicited by egg-adapted vaccine strains. Proc. Natl. Acad. Sci. USA 2017, 114, 12578–12583. [Google Scholar] [CrossRef] [PubMed]
- Widjaja, L.; Ilyushina, N.; Webster, R.G.; Webby, R.J. Molecular changes associated with adaptation of human influenza a virus in embryonated chicken eggs. Virology 2006, 350, 137–145. [Google Scholar] [CrossRef] [PubMed]
- Nicholls, J.M.; Chan, R.W.; Russell, R.J.; Air, G.M.; Peiris, J.S. Evolving complexities of influenza virus and its receptors. Trends Microbiol. 2008, 16, 149–157. [Google Scholar] [CrossRef] [PubMed]
- Imai, M.; Kawaoka, Y. The role of receptor binding specificity in interspecies transmission of influenza viruses. Curr. Opin. Virol. 2012, 2, 160–167. [Google Scholar] [CrossRef] [PubMed]
- Cobey, S.; Hensley, S.E. Immune history and influenza virus susceptibility. Curr. Opin. Virol. 2017, 22, 105–111. [Google Scholar] [CrossRef] [PubMed]
- Paules, C.I.; Sullivan, S.G.; Subbarao, K.; Fauci, A.S. Chasing seasonal influenza—The need for a universal influenza vaccine. N. Engl. J. Med. 2018, 378, 7–9. [Google Scholar] [CrossRef] [PubMed]
- Flannery, B.; Chung, J.R.; Belongia, E.A.; McLean, H.Q.; Gaglani, M.; Murthy, K.; Zimmerman, R.K.; Nowalk, M.P.; Jackson, M.L.; Jackson, L.A.; et al. Interim estimates of 2017–2018 seasonal influenza vaccine effectiveness—United States, February 2018. Morb. Mortal. Wkly. Rep. 2018, 67, 180–185. [Google Scholar] [CrossRef] [PubMed]
- CDC. Cell-Based Flu Vaccines. Available online: https://www.cdc.gov/flu/protect/vaccine/cell-based.htm (accessed on 12 February 2018).
- Hegde, N.R. Cell culture-based influenza vaccines: A necessary and indispensable investment for the future. Hum. Vaccines Immunother. 2015, 11, 1223–1234. [Google Scholar] [CrossRef] [PubMed]
- Milian, E.; Kamen, A.A. Current and emerging cell culture manufacturing technologies for influenza vaccines. BioMed Res. Int. 2015, 2015, 504831. [Google Scholar] [CrossRef] [PubMed]
- Hutter, J.; Rodig, J.V.; Hoper, D.; Seeberger, P.H.; Reichl, U.; Rapp, E.; Lepenies, B. Toward animal cell culture-based influenza vaccine design: Viral hemagglutinin N-glycosylation markedly impacts immunogenicity. J. Immunol. 2013, 190, 220–230. [Google Scholar] [CrossRef] [PubMed]
- An, Y.; Rininger, J.A.; Jarvis, D.L.; Jing, X.; Ye, Z.; Aumiller, J.J.; Eichelberger, M.; Cipollo, J.F. Comparative glycomics analysis of influenza hemagglutinin (H5N1) produced in vaccine relevant cell platforms. J. Proteome Res. 2013, 12, 3707–3720. [Google Scholar] [CrossRef] [PubMed]
- Branswell, H. Flu Vaccine Grown without Eggs Provided Measurably Better Protection This Season, FDA Says. Available online: https://www.pbs.org/newshour/health/flu-vaccine-grown-without-eggs-provided-measurably-better-protection-this-season-fda-says (accessed on 10 March 2018).
- Lin, Y.; Wharton, S.A.; Whittaker, L.; Dai, M.; Ermetal, B.; Lo, J.; Pontoriero, A.; Baumeister, E.; Daniels, R.S.; McCauley, J.W. The characteristics and antigenic properties of recently emerged subclade 3c.3a and 3c.2a human influenza a(H3N2) viruses passaged in mdck cells. Influenza Other Respir. Viruses 2017, 11, 263–274. [Google Scholar] [CrossRef] [PubMed]
- CDC. Seasonal Influenza Vaccine Supply for the U.S. 2017–2018 Influenza Season. Available online: https://www.cdc.gov/flu/about/qa/vaxsupply.htm (accessed on 19 March 2018).
- CDC. Flublok Seasonal Influenza (Flu) Vaccine. Available online: https://www.cdc.gov/flu/protect/vaccine/qa_flublok-vaccine.htm (accessed on 12 February 2018).
- Cox, M.M.; Hashimoto, Y. A fast track influenza virus vaccine produced in insect cells. J. Invertebr. Pathol. 2011, 107, S31–S41. [Google Scholar] [CrossRef] [PubMed]
- Buckland, B.; Boulanger, R.; Fino, M.; Srivastava, I.; Holtz, K.; Khramtsov, N.; McPherson, C.; Meghrous, J.; Kubera, P.; Cox, M.M. Technology transfer and scale-up of the flublok recombinant hemagglutinin (ha) influenza vaccine manufacturing process. Vaccine 2014, 32, 5496–5502. [Google Scholar] [CrossRef] [PubMed]
- Cox, M.M.; Izikson, R.; Post, P.; Dunkle, L. Safety, efficacy and immunogenicity of flublok in the prevention of seasonal influenza in adults. Ther. Adv. Vaccines 2015, 3, 97–108. [Google Scholar] [CrossRef] [PubMed]
- NCIRD. Archived CDC Vaccine Price List as of 2 February 2017. Available online: https://www.cdc.gov/vaccines/programs/vfc/awardees/vaccine-management/price-list/2017/2017-02-02.html (accessed on 22 February 2018).
- CDC. Fluad™ Flu Vaccine with Adjuvant. Available online: https://www.cdc.gov/flu/protect/vaccine/adjuvant.htm (accessed on 20 March 2018).
- O’Hagan, D.T.; Ott, G.S.; Nest, G.V.; Rappuoli, R.; Giudice, G.D. The history of mf59((r)) adjuvant: A phoenix that arose from the ashes. Expert Rev. Vaccines 2013, 12, 13–30. [Google Scholar] [CrossRef] [PubMed]
- O’Hagan, D.T.; Ott, G.S.; De Gregorio, E.; Seubert, A. The mechanism of action of mf59—An innately attractive adjuvant formulation. Vaccine 2012, 30, 4341–4348. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization (WHO). Squalene-Based Adjuvants in Vaccines. Available online: http://www.who.int/vaccine_safety/committee/topics/adjuvants/squalene/questions_and_answers/en/ (accessed on 20 March 2018).
- Cruz-Valdez, A.; Valdez-Zapata, G.; Patel, S.S.; Castelli, F.V.; Garcia, M.G.; Jansen, W.T.; Arora, A.K.; Heijnen, E. Mf59-adjuvanted influenza vaccine (fluad(r)) elicits higher immune responses than a non-adjuvanted influenza vaccine (fluzone(r)): A randomized, multicenter, phase iii pediatric trial in Mexico. Hum. Vaccines Immunother. 2018, 14, 386–395. [Google Scholar] [CrossRef] [PubMed]
- O’Hagan, D.T.; Rappuoli, R.; De Gregorio, E.; Tsai, T.; Del Giudice, G. Mf59 adjuvant: The best insurance against influenza strain diversity. Expert Rev. Vaccines 2011, 10, 447–462. [Google Scholar] [CrossRef] [PubMed]
- Harding, A.T.; Heaton, B.E.; Dumm, R.E.; Heaton, N.S. Rationally designed influenza virus vaccines that are antigenically stable during growth in eggs. mBio 2017, 8, e00669-17. [Google Scholar] [CrossRef] [PubMed]
- Lu, B.; Zhou, H.; Ye, D.; Kemble, G.; Jin, H. Improvement of influenza a/Fujian/411/02 (H3N2) virus growth in embryonated chicken eggs by balancing the hemagglutinin and neuraminidase activities, using reverse genetics. J. Virol. 2005, 79, 6763–6771. [Google Scholar] [CrossRef] [PubMed]
- Nicolson, C.; Major, D.; Wood, J.M.; Robertson, J.S. Generation of influenza vaccine viruses on vero cells by reverse genetics: An H5N1 candidate vaccine strain produced under a quality system. Vaccine 2005, 23, 2943–2952. [Google Scholar] [CrossRef] [PubMed]
- Smith, G.; Liu, Y.; Flyer, D.; Massare, M.J.; Zhou, B.; Patel, N.; Ellingsworth, L.; Lewis, M.; Cummings, J.F.; Glenn, G. Novel hemagglutinin nanoparticle influenza vaccine with matrix-m adjuvant induces hemagglutination inhibition, neutralizing and protective responses in ferrets against homologous and drifted a(H3N2) subtypes. Vaccine 2017, 35, 5366–5372. [Google Scholar] [CrossRef] [PubMed]
- Kanekiyo, M.; Wei, C.J.; Yassine, H.M.; McTamney, P.M.; Boyington, J.C.; Whittle, J.R.; Rao, S.S.; Kong, W.P.; Wang, L.; Nabel, G.J. Self-assembling influenza nanoparticle vaccines elicit broadly neutralizing H1N1 antibodies. Nature 2013, 499, 102–106. [Google Scholar] [CrossRef] [PubMed]
- Quan, F.S.; Lee, Y.T.; Kim, K.H.; Kim, M.C.; Kang, S.M. Progress in developing virus-like particle influenza vaccines. Expert Rev. Vaccines 2016, 15, 1281–1293. [Google Scholar] [CrossRef] [PubMed]
- Aguilar, J.C.; Rodriguez, E.G. Vaccine adjuvants revisited. Vaccine 2007, 25, 3752–3762. [Google Scholar] [CrossRef] [PubMed]
- Singh, M.; O’Hagan, D. Advances in vaccine adjuvants. Nat. Biotechnol. 1999, 17, 1075–1081. [Google Scholar] [CrossRef] [PubMed]
- Soema, P.C.; Kompier, R.; Amorij, J.P.; Kersten, G.F. Current and next generation influenza vaccines: Formulation and production strategies. Eur. J. Pharm. Biopharm. 2015, 94, 251–263. [Google Scholar] [CrossRef] [PubMed]
- Soema, P.C.; Rosendahl Huber, S.K.; Willems, G.J.; Jiskoot, W.; Kersten, G.F.; Amorij, J.P. Influenza T-cell epitope-loaded virosomes adjuvanted with cpg as a potential influenza vaccine. Pharm. Res. 2015, 32, 1505–1515. [Google Scholar] [CrossRef] [PubMed]
- Naruse, H.; Ogasawara, K.; Kaneda, R.; Hatakeyama, S.; Itoh, T.; Kida, H.; Miyazaki, T.; Good, R.A.; Onoe, K. A potential peptide vaccine against two different strains of influenza virus isolated at intervals of about 10 years. Proc. Natl. Acad. Sci. USA 1994, 91, 9588–9592. [Google Scholar] [CrossRef] [PubMed]
- Ninomiya, A.; Ogasawara, K.; Kajino, K.; Takada, A.; Kida, H. Intranasal administration of a synthetic peptide vaccine encapsulated in liposome together with an anti-cd40 antibody induces protective immunity against influenza a virus in mice. Vaccine 2002, 20, 3123–3129. [Google Scholar] [CrossRef]
- Sercombe, L.; Veerati, T.; Moheimani, F.; Wu, S.Y.; Sood, A.K.; Hua, S. Advances and challenges of liposome assisted drug delivery. Front. Pharmacol. 2015, 6, 286. [Google Scholar] [CrossRef] [PubMed]
- Moser, C.; Muller, M.; Kaeser, M.D.; Weydemann, U.; Amacker, M. Influenza virosomes as vaccine adjuvant and carrier system. Expert Rev. Vaccines 2013, 12, 779–791. [Google Scholar] [CrossRef] [PubMed]
- Shen, K.Y.; Liu, H.Y.; Li, H.J.; Wu, C.C.; Liou, G.G.; Chang, Y.C.; Leng, C.H.; Liu, S.J. A novel liposomal recombinant lipoimmunogen enhances anti-tumor immunity. J. Control. Release 2016, 233, 57–63. [Google Scholar] [CrossRef] [PubMed]
- Ma, J.H.; Yang, F.R.; Yu, H.; Zhou, Y.J.; Li, G.X.; Huang, M.; Wen, F.; Tong, G. An m2e-based synthetic peptide vaccine for influenza A virus confers heterosubtypic protection from lethal virus challenge. Virol. J. 2013, 10, 227. [Google Scholar] [CrossRef] [PubMed]
- Stachyra, A.; Gora-Sochacka, A.; Sirko, A. DNA vaccines against influenza. Acta Biochim. Pol. 2014, 61, 515–522. [Google Scholar] [PubMed]
- Kramps, T.; Probst, J. Messenger rna-based vaccines: Progress, challenges, applications. Wiley Interdiscip. Rev. RNA 2013, 4, 737–749. [Google Scholar] [CrossRef] [PubMed]
- Vogel, F.R.; Sarver, N. Nucleic acid vaccines. Clin. Microbiol. Rev. 1995, 8, 406–410. [Google Scholar] [PubMed]
- Shen, X.; Soderholm, J.; Lin, F.; Kobinger, G.; Bello, A.; Gregg, D.A.; Broderick, K.E.; Sardesai, N.Y. Influenza A vaccines using linear expression cassettes delivered via electroporation afford full protection against challenge in a mouse model. Vaccine 2012, 30, 6946–6954. [Google Scholar] [CrossRef] [PubMed]
- Ledgerwood, J.E.; Wei, C.J.; Hu, Z.; Gordon, I.J.; Enama, M.E.; Hendel, C.S.; McTamney, P.M.; Pearce, M.B.; Yassine, H.M.; Boyington, J.C.; et al. DNA priming and influenza vaccine immunogenicity: Two phase 1 open label randomised clinical trials. Lancet Infect. Dis. 2011, 11, 916–924. [Google Scholar] [CrossRef]
- Ledgerwood, J.E.; Hu, Z.; Gordon, I.J.; Yamshchikov, G.; Enama, M.E.; Plummer, S.; Bailer, R.; Pearce, M.B.; Tumpey, T.M.; Koup, R.A.; et al. Influenza virus H5 DNA vaccination is immunogenic by intramuscular and intradermal routes in humans. Clin. Vaccine Immunol. 2012, 19, 1792–1797. [Google Scholar] [CrossRef] [PubMed]
- Smith, L.R.; Wloch, M.K.; Ye, M.; Reyes, L.R.; Boutsaboualoy, S.; Dunne, C.E.; Chaplin, J.A.; Rusalov, D.; Rolland, A.P.; Fisher, C.L.; et al. Phase 1 clinical trials of the safety and immunogenicity of adjuvanted plasmid DNA vaccines encoding influenza a virus H5 hemagglutinin. Vaccine 2010, 28, 2565–2572. [Google Scholar] [CrossRef] [PubMed]
- Jones, S.; Evans, K.; McElwaine-Johnn, H.; Sharpe, M.; Oxford, J.; Lambkin-Williams, R.; Mant, T.; Nolan, A.; Zambon, M.; Ellis, J.; et al. DNA vaccination protects against an influenza challenge in a double-blind randomised placebo-controlled phase 1b clinical trial. Vaccine 2009, 27, 2506–2512. [Google Scholar] [CrossRef] [PubMed]
- Petsch, B.; Schnee, M.; Vogel, A.B.; Lange, E.; Hoffmann, B.; Voss, D.; Schlake, T.; Thess, A.; Kallen, K.J.; Stitz, L.; et al. Protective efficacy of in vitro synthesized, specific mrna vaccines against influenza a virus infection. Nat. Biotechnol. 2012, 30, 1210–1216. [Google Scholar] [CrossRef] [PubMed]
- Kaczmarek, J.C.; Kowalski, P.S.; Anderson, D.G. Advances in the delivery of rna therapeutics: From concept to clinical reality. Genome Med. 2017, 9, 60. [Google Scholar] [CrossRef] [PubMed]
- Diken, M.; Kreiter, S.; Selmi, A.; Britten, C.M.; Huber, C.; Tureci, O.; Sahin, U. Selective uptake of naked vaccine rna by dendritic cells is driven by macropinocytosis and abrogated upon dc maturation. Gene Ther. 2011, 18, 702–708. [Google Scholar] [CrossRef] [PubMed]
- Lorenz, C.; Fotin-Mleczek, M.; Roth, G.; Becker, C.; Dam, T.C.; Verdurmen, W.P.; Brock, R.; Probst, J.; Schlake, T. Protein expression from exogenous mrna: Uptake by receptor-mediated endocytosis and trafficking via the lysosomal pathway. RNA Biol. 2011, 8, 627–636. [Google Scholar] [CrossRef] [PubMed]
- Redding, L.; Weiner, D.B. DNA vaccines in veterinary use. Expert Rev. Vaccines 2009, 8, 1251–1276. [Google Scholar] [CrossRef] [PubMed]
- Ranjan, P.; Bowzard, J.B.; Schwerzmann, J.W.; Jeisy-Scott, V.; Fujita, T.; Sambhara, S. Cytoplasmic nucleic acid sensors in antiviral immunity. Trends Mol. Med. 2009, 15, 359–368. [Google Scholar] [CrossRef] [PubMed]
- Kariko, K.; Muramatsu, H.; Welsh, F.A.; Ludwig, J.; Kato, H.; Akira, S.; Weissman, D. Incorporation of pseudouridine into mrna yields superior nonimmunogenic vector with increased translational capacity and biological stability. Mol. Ther. 2008, 16, 1833–1840. [Google Scholar] [CrossRef] [PubMed]
- Pica, N.; Palese, P. Toward a universal influenza virus vaccine: Prospects and challenges. Annu. Rev. Med. 2013, 64, 189–202. [Google Scholar] [CrossRef] [PubMed]
- Sautto, G.A.; Kirchenbaum, G.A.; Ross, T.M. Towards a universal influenza vaccine: Different approaches for one goal. Virol. J. 2018, 15, 17. [Google Scholar] [CrossRef] [PubMed]
- Nachbagauer, R.; Krammer, F. Universal influenza virus vaccines and therapeutic antibodies. Clin. Microbiol. Infect. 2017, 23, 222–228. [Google Scholar] [CrossRef] [PubMed]
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Harding, A.T.; Heaton, N.S. Efforts to Improve the Seasonal Influenza Vaccine. Vaccines 2018, 6, 19. https://doi.org/10.3390/vaccines6020019
Harding AT, Heaton NS. Efforts to Improve the Seasonal Influenza Vaccine. Vaccines. 2018; 6(2):19. https://doi.org/10.3390/vaccines6020019
Chicago/Turabian StyleHarding, Alfred T., and Nicholas S. Heaton. 2018. "Efforts to Improve the Seasonal Influenza Vaccine" Vaccines 6, no. 2: 19. https://doi.org/10.3390/vaccines6020019
APA StyleHarding, A. T., & Heaton, N. S. (2018). Efforts to Improve the Seasonal Influenza Vaccine. Vaccines, 6(2), 19. https://doi.org/10.3390/vaccines6020019