
Direct Delivery of Antigens to Dendritic Cells via Antibodies Specific for Endocytic Receptors as a Promising Strategy for Future Therapies

 , ,
, ,
Abstract
:1. Introduction
2. Pattern Recognition Receptors
3. Monocyte-Derived DCs in Immunotherapeutic Approaches
4. Delivery of Antigens to DC Subsets by Usage of Recombinant Antibodies
5. Antigen Targeting Receptors
5.1. DC Subsets
5.2. Antigen Targeting to DCs and DC Subsets by Utilizing C Type Lectin Receptors
5.3. Other Receptors Used for Antigen Delivery to APCs
6. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Sharpe, A.H.; Freeman, G.J. The B7-CD28 superfamily. Nat. Rev. Immunol. 2002, 2, 116–126. [Google Scholar] [CrossRef] [PubMed]
- Steinman, R.M. Dendritic cells: Understanding immunogenicity. Eur. J. Immunol. 2007. [Google Scholar] [CrossRef] [PubMed]
- Dudziak, D.; Kamphorst, A.O.; Heidkamp, G.F.; Buchholz, V.R.; Trumpfheller, C.; Yamazaki, S.; Cheong, C.; Liu, K.; Lee, H.W.; Park, C.G.; et al. Differential antigen processing by dendritic cell subsets in vivo. Science 2007, 315, 107–111. [Google Scholar] [CrossRef] [PubMed]
- Steinman, R.M.; Hawiger, D.; Liu, K.; Bonifaz, L.; Bonnyay, D.; Mahnke, K.; Iyoda, T.; Ravetch, J.; Dhodapkar, M.; Inaba, K.; et al. Dendritic cell function in vivo during the steady state: A role in peripheral tolerance. Ann. NY Acad. Sci. 2003, 987, 15–25. [Google Scholar] [CrossRef] [PubMed]
- Steinman, R.M.; Hawiger, D.; Nussenzweig, M.C. Tolerogenic dendritic cells. Ann. Rev. Immunol. 2003, 21, 685–711. [Google Scholar] [CrossRef] [PubMed]
- Steinman, R.M.; Nussenzweig, M.C. Avoiding horror autotoxicus: The importance of dendritic cells in peripheral T cell tolerance. Proc. Nat. Acad. Sci. USA 2002, 99, 351–358. [Google Scholar] [CrossRef] [PubMed]
- Yamazaki, S.; Dudziak, D.; Heidkamp, G.F.; Fiorese, C.; Bonito, A.J.; Inaba, K.; Nussenzweig, M.C.; Steinman, R.M. CD8+ CD205+ splenic dendritic cells are specialized to induce Foxp3+ regulatory T cells. J. Immunol. 2008, 181, 6923–6933. [Google Scholar] [CrossRef] [PubMed]
- Hawiger, D.; Inaba, K.; Dorsett, Y.; Guo, M.; Mahnke, K.; Rivera, M.; Ravetch, J.V.; Steinman, R.M.; Nussenzweig, M.C. Dendritic cells induce peripheral T cell unresponsiveness under steady state conditions in vivo. J. Exp. Med. 2001, 194, 769–779. [Google Scholar] [CrossRef] [PubMed]
- Hammad, H.; Lambrecht, B.N. Dendritic cells and airway epithelial cells at the interface between innate and adaptive immune responses. Allergy 2011, 66, 579–587. [Google Scholar] [CrossRef] [PubMed]
- Joffre, O.; Nolte, M.A.; Sporri, R.; Reis e Sousa, C. Inflammatory signals in dendritic cell activation and the induction of adaptive immunity. Immunol. Rev. 2009, 227, 234–247. [Google Scholar] [CrossRef] [PubMed]
- Merad, M.; Sathe, P.; Helft, J.; Miller, J.; Mortha, A. The dendritic cell lineage: Ontogeny and function of dendritic cells and their subsets in the steady state and the inflamed setting. Ann. Rev. Immunol. 2013, 31, 563–604. [Google Scholar] [CrossRef] [PubMed]
- Banchereau, J.; Steinman, R.M. Dendritic cells and the control of immunity. Nature 1998, 392, 245–252. [Google Scholar] [CrossRef] [PubMed]
- Shortman, K.; Liu, Y.J. Mouse and human dendritic cell subtypes. Nat. Rev. Immunol. 2002, 2, 151–161. [Google Scholar] [CrossRef] [PubMed]
- Akira, S.; Uematsu, S.; Takeuchi, O. Pathogen recognition and innate immunity. Cell 2006, 124, 783–801. [Google Scholar] [CrossRef] [PubMed]
- Bowie, A.; O’Neill, L.A. The interleukin-1 receptor/toll-like receptor superfamily: Signal generators for pro-inflammatory interleukins and microbial products. J. Leukoc. Biol. 2000, 67, 508–514. [Google Scholar] [PubMed]
- Brencicova, E.; Diebold, S.S. Nucleic acids and endosomal pattern recognition: How to tell friend from foe? Front. Cell. Infect. Microbiol. 2013. [Google Scholar] [CrossRef] [PubMed]
- Cambi, A.; Figdor, C.G. Dual function of C-type lectin-like receptors in the immune system. Curr. Opin. Cell Biol. 2003, 15, 539–546. [Google Scholar] [CrossRef] [PubMed]
- Figdor, C.G.; van Kooyk, Y.; Adema, G.J. C-type lectin receptors on dendritic cells and langerhans cells. Nat. Rev. Immunol. 2002, 2, 77–84. [Google Scholar] [CrossRef] [PubMed]
- Ottenhoff, T.H.; Bevan, M.J. Host-pathogen interactions. Curr. Opin. Immunol. 2004, 16, 439–442. [Google Scholar] [CrossRef] [PubMed]
- Segura, E.; Villadangos, J.A. Antigen presentation by dendritic cells in vivo. Curr. Opin. Immunol. 2009, 21, 105–110. [Google Scholar] [CrossRef] [PubMed]
- Swiecki, M.; Colonna, M. The multifaceted biology of plasmacytoid dendritic cells. Nat. Rev. Immunol. 2015, 15, 471–485. [Google Scholar] [CrossRef] [PubMed]
- Takeuchi, O.; Akira, S. Pattern recognition receptors and inflammation. Cell 2010, 140, 805–820. [Google Scholar] [CrossRef] [PubMed]
- Pufnock, J.S.; Cigal, M.; Rolczynski, L.S.; Andersen-Nissen, E.; Wolfl, M.; McElrath, M.J.; Greenberg, P.D. Priming CD8+ T cells with dendritic cells matured using TLR4 and TLR7/8 ligands together enhances generation of CD8+ T cells retaining CD28. Blood 2011, 117, 6542–6551. [Google Scholar] [CrossRef] [PubMed]
- Shen, H.; Tesar, B.M.; Walker, W.E.; Goldstein, D.R. Dual signaling of MyD88 and trif is critical for maximal TLR4-induced dendritic cell maturation. J. Immunol. 2008, 181, 1849–1858. [Google Scholar] [CrossRef] [PubMed]
- Delamarre, L.; Holcombe, H.; Mellman, I. Presentation of exogenous antigens on major histocompatibility complex (MHC) class I and MHC class II molecules is differentially regulated during dendritic cell maturation. J. Exp. Med. 2003, 198, 111–122. [Google Scholar] [CrossRef] [PubMed]
- Kaufmann, S.H. The contribution of immunology to the rational design of novel antibacterial vaccines. Nat. Rev. Microbiol. 2007, 5, 491–504. [Google Scholar] [CrossRef] [PubMed]
- Bowie, A.G.; Unterholzner, L. Viral evasion and subversion of pattern-recognition receptor signalling. Nat. Rev. Immunol. 2008, 8, 911–922. [Google Scholar] [CrossRef] [PubMed]
- Varki, A.; Etzler, M.E.; Cummings, R.D.; Esko, J.D. Discovery and classification of glycan-binding proteins. In Essentials of Glycobiology, 2nd ed.; Varki, A., Cummings, R.D., Esko, J.D., Freeze, H.H., Stanley, P., Bertozzi, C.R., Hart, G.W., Etzler, M.E., Eds.; Cold Spring Harbor: New York, NY, USA, 2009. [Google Scholar]
- Geijtenbeek, T.B.; Gringhuis, S.I. Signalling through C-type lectin receptors: Shaping immune responses. Nat. Rev. Immunol. 2009, 9, 465–479. [Google Scholar] [CrossRef] [PubMed]
- Van Kooyk, Y.; Rabinovich, G.A. Protein-glycan interactions in the control of innate and adaptive immune responses. Nat. Immunol. 2008, 9, 593–601. [Google Scholar] [CrossRef] [PubMed]
- Giraldi-Guimaraes, A.; de Freitas, H.T.; Coelho Bde, P.; Macedo-Ramos, H.; Mendez-Otero, R.; Cavalcante, L.A.; Baetas-da-Cruz, W. Bone marrow mononuclear cells and mannose receptor expression in focal cortical ischemia. Brain Res. 2012, 1452, 173–184. [Google Scholar] [CrossRef] [PubMed]
- Hoque, R.; Malik, A.F.; Gorelick, F.; Mehal, W.Z. Sterile inflammatory response in acute pancreatitis. Pancreas 2012, 41, 353–357. [Google Scholar] [CrossRef] [PubMed]
- Mathew, A.; Lindsley, T.A.; Sheridan, A.; Bhoiwala, D.L.; Hushmendy, S.F.; Yager, E.J.; Ruggiero, E.A.; Crawford, D.R. Degraded mitochondrial DNA is a newly identified subtype of the damage associated molecular pattern (DAMP) family and possible trigger of neurodegeneration. J. Alzheimers Dis. 2012, 30, 617–627. [Google Scholar] [PubMed]
- Sancho, D.; Joffre, O.P.; Keller, A.M.; Rogers, N.C.; Martinez, D.; Hernanz-Falcon, P.; Rosewell, I.; Reis e Sousa, C. Identification of a dendritic cell receptor that couples sensing of necrosis to immunity. Nature 2009, 458, 899–903. [Google Scholar] [CrossRef] [PubMed]
- Anton, L.C.; Yewdell, J.W. Translating drips: MHC class I immunosurveillance of pathogens and tumors. J. Leukoc. Biol. 2014, 95, 551–562. [Google Scholar] [CrossRef] [PubMed]
- Lutz, M.B.; Schuler, G. Immature, semi-mature and fully mature dendritic cells: Which signals induce tolerance or immunity? Trends Immunol. 2002, 23, 445–449. [Google Scholar] [CrossRef]
- Schwartz, R.H. Acquisition of immunologic self-tolerance. Cell 1989, 57, 1073–1081. [Google Scholar] [CrossRef]
- Copier, J.; Bodman-Smith, M.; Dalgleish, A. Current status and future applications of cellular therapies for cancer. Immunotherapy 2011, 3, 507–516. [Google Scholar] [CrossRef] [PubMed]
- Keler, T.; He, L.; Ramakrishna, V.; Champion, B. Antibody-targeted vaccines. Oncogene 2007, 26, 3758–3767. [Google Scholar] [CrossRef] [PubMed]
- Osada, T.; Nagaoka, K.; Takahara, M.; Yang, X.Y.; Liu, C.X.; Guo, H.; Roy Choudhury, K.; Hobeika, A.; Hartman, Z.; Morse, M.A.; et al. Precision cancer immunotherapy: Optimizing dendritic cell-based strategies to induce tumor antigen-specific t-cell responses against individual patient tumors. J. Immunother. 2015, 38, 155–164. [Google Scholar] [CrossRef] [PubMed]
- Palucka, K.; Banchereau, J. Cancer immunotherapy via dendritic cells. Nat. Rev. Cancer 2012, 12, 265–277. [Google Scholar] [CrossRef] [PubMed]
- Schuler, P.J.; Harasymczuk, M.; Visus, C.; Deleo, A.; Trivedi, S.; Lei, Y.; Argiris, A.; Gooding, W.; Butterfield, L.H.; Whiteside, T.L.; et al. Phase I dendritic cell p53 peptide vaccine for head and neck cancer. Clin. Cancer Res. 2014, 20, 2433–2444. [Google Scholar] [CrossRef] [PubMed]
- Tacken, P.J.; Figdor, C.G. Targeted antigen delivery and activation of dendritic cells in vivo: Steps towards cost effective vaccines. Semin. Immunol. 2011, 23, 12–20. [Google Scholar] [CrossRef] [PubMed]
- Steinman, R.M.; Banchereau, J. Taking dendritic cells into medicine. Nature 2007, 449, 419–426. [Google Scholar] [CrossRef] [PubMed]
- Jonuleit, H.; Kuhn, U.; Muller, G.; Steinbrink, K.; Paragnik, L.; Schmitt, E.; Knop, J.; Enk, A.H. Pro-inflammatory cytokines and prostaglandins induce maturation of potent immunostimulatory dendritic cells under fetal calf serum-free conditions. Eur. J. Immunol. 1997, 27, 3135–3142. [Google Scholar] [CrossRef] [PubMed]
- Romani, N.; Koide, S.; Crowley, M.; Witmer-Pack, M.; Livingstone, A.M.; Fathman, C.G.; Inaba, K.; Steinman, R.M. Presentation of exogenous protein antigens by dendritic cells to T cell clones. Intact protein is presented best by immature, epidermal langerhans cells. J. Exp. Med. 1989, 169, 1169–1178. [Google Scholar] [CrossRef] [PubMed]
- Sallusto, F.; Cella, M.; Danieli, C.; Lanzavecchia, A. Dendritic cells use macropinocytosis and the mannose receptor to concentrate macromolecules in the major histocompatibility complex class II compartment: Downregulation by cytokines and bacterial products. J. Exp. Med. 1995, 182, 389–400. [Google Scholar] [CrossRef] [PubMed]
- Sallusto, F.; Lanzavecchia, A. Efficient presentation of soluble antigen by cultured human dendritic cells is maintained by granulocyte/macrophage colony-stimulating factor plus interleukin 4 and downregulated by tumor necrosis factor alpha. J. Exp. Med. 1994, 179, 1109–1118. [Google Scholar] [CrossRef] [PubMed]
- Schuler-Thurner, B.; Schultz, E.S.; Berger, T.G.; Weinlich, G.; Ebner, S.; Woerl, P.; Bender, A.; Feuerstein, B.; Fritsch, P.O.; Romani, N.; et al. Rapid induction of tumor-specific type 1 T helper cells in metastatic melanoma patients by vaccination with mature, cryopreserved, peptide-loaded monocyte-derived dendritic cells. J. Exp. Med. 2002, 195, 1279–1288. [Google Scholar] [CrossRef] [PubMed]
- Guilliams, M.; Bruhns, P.; Saeys, Y.; Hammad, H.; Lambrecht, B.N. The function of fcgamma receptors in dendritic cells and macrophages. Nat. Rev. Immunol. 2014, 14, 94–108. [Google Scholar] [CrossRef] [PubMed]
- de Vries, I.J.; Lesterhuis, W.J.; Scharenborg, N.M.; Engelen, L.P.; Ruiter, D.J.; Gerritsen, M.J.; Croockewit, S.; Britten, C.M.; Torensma, R.; Adema, G.J.; et al. Maturation of dendritic cells is a prerequisite for inducing immune responses in advanced melanoma patients. Clin. Cancer Res. 2003, 9, 5091–5100. [Google Scholar] [PubMed]
- Dhodapkar, M.V.; Krasovsky, J.; Steinman, R.M.; Bhardwaj, N. Mature dendritic cells boost functionally superior CD8(+) t-cell in humans without foreign helper epitopes. J. Clin. Investig. 2000, 105, 9–14. [Google Scholar] [CrossRef] [PubMed]
- Mahnke, K.; Schmitt, E.; Bonifaz, L.; Enk, A.H.; Jonuleit, H. Immature, but not inactive: The tolerogenic function of immature dendritic cells. Immunol. Cell Biol. 2002, 80, 477–483. [Google Scholar] [CrossRef] [PubMed]
- Schuler, G. Dendritic cells in cancer immunotherapy. Eur. J. Immunol. 2010, 40, 2123–2130. [Google Scholar] [CrossRef] [PubMed]
- Fong, L.; Brockstedt, D.; Benike, C.; Wu, L.; Engleman, E.G. Dendritic cells injected via different routes induce immunity in cancer patients. J. Immunol. 2001, 166, 4254–4259. [Google Scholar] [CrossRef] [PubMed]
- Jonuleit, H.; Giesecke-Tuettenberg, A.; Tuting, T.; Thurner-Schuler, B.; Stuge, T.B.; Paragnik, L.; Kandemir, A.; Lee, P.P.; Schuler, G.; Knop, J.; et al. A comparison of two types of dendritic cell as adjuvants for the induction of melanoma-specific t-cell responses in humans following intranodal injection. Int. J. Cancer. 2001, 93, 243–251. [Google Scholar] [CrossRef] [PubMed]
- Oshita, C.; Takikawa, M.; Kume, A.; Miyata, H.; Ashizawa, T.; Iizuka, A.; Kiyohara, Y.; Yoshikawa, S.; Tanosaki, R.; Yamazaki, N.; et al. Dendritic cell-based vaccination in metastatic melanoma patients: Phase ii clinical trial. Oncol. Rep. 2012, 28, 1131–1138. [Google Scholar] [PubMed]
- Tada, F.; Abe, M.; Hirooka, M.; Ikeda, Y.; Hiasa, Y.; Lee, Y.; Jung, N.C.; Lee, W.B.; Lee, H.S.; Bae, Y.S.; et al. Phase I/II study of immunotherapy using tumor antigen-pulsed dendritic cells in patients with hepatocellular carcinoma. Int. J. Oncol. 2012, 41, 1601–1609. [Google Scholar] [PubMed]
- Ardon, H.; Van Gool, S.W.; Verschuere, T.; Maes, W.; Fieuws, S.; Sciot, R.; Wilms, G.; Demaerel, P.; Goffin, J.; Van Calenbergh, F.; et al. Integration of autologous dendritic cell-based immunotherapy in the standard of care treatment for patients with newly diagnosed glioblastoma: Results of the HGG-2006 phase I/II trial. Cancer Immunol. Immunother. 2012, 61, 2033–2044. [Google Scholar] [CrossRef] [PubMed]
- Celluzzi, C.M.; Falo, L.D., Jr. Physical interaction between dendritic cells and tumor cells results in an immunogen that induces protective and therapeutic tumor rejection. J. Immunol. 1998, 160, 3081–3085. [Google Scholar] [PubMed]
- Chang, A.E.; Redman, B.G.; Whitfield, J.R.; Nickoloff, B.J.; Braun, T.M.; Lee, P.P.; Geiger, J.D.; Mule, J.J. A phase I trial of tumor lysate-pulsed dendritic cells in the treatment of advanced cancer. Clin. Cancer Res. 2002, 8, 1021–1032. [Google Scholar] [PubMed]
- El Ansary, M.; Mogawer, S.; Elhamid, S.A.; Alwakil, S.; Aboelkasem, F.; Sabaawy, H.E.; Abdelhalim, O. Immunotherapy by autologous dendritic cell vaccine in patients with advanced HCC. J. Cancer Res. Clin. Oncol. 2013, 139, 39–48. [Google Scholar] [CrossRef] [PubMed]
- Gao, D.; Li, C.; Xie, X.; Zhao, P.; Wei, X.; Sun, W.; Liu, H.C.; Alexandrou, A.T.; Jones, J.; Zhao, R.; et al. Autologous tumor lysate-pulsed dendritic cell immunotherapy with cytokine-induced killer cells improves survival in gastric and colorectal cancer patients. PLoS ONE 2014, 9, e93886. [Google Scholar] [CrossRef] [PubMed]
- Hunyadi, J.; Andras, C.; Szabo, I.; Szanto, J.; Szluha, K.; Sipka, S.; Kovacs, P.; Kiss, A.; Szegedi, G.; Altorjay, I.; et al. Autologous dendritic cell based adoptive immunotherapy of patients with colorectal cancer-a phase I-II study. Pathol. Oncol. Res. 2014, 20, 357–365. [Google Scholar] [CrossRef] [PubMed]
- Matsushita, H.; Enomoto, Y.; Kume, H.; Nakagawa, T.; Fukuhara, H.; Suzuki, M.; Fujimura, T.; Homma, Y.; Kakimi, K. A pilot study of autologous tumor lysate-loaded dendritic cell vaccination combined with sunitinib for metastatic renal cell carcinoma. J. Immunother. Cancer 2014. [Google Scholar] [CrossRef] [PubMed]
- Nouri-Shirazi, M.; Banchereau, J.; Bell, D.; Burkeholder, S.; Kraus, E.T.; Davoust, J.; Palucka, K.A. Dendritic cells capture killed tumor cells and present their antigens to elicit tumor-specific immune responses. J. Immunol. 2000, 165, 3797–3803. [Google Scholar] [CrossRef] [PubMed]
- Aarntzen, E.H.; Schreibelt, G.; Bol, K.; Lesterhuis, W.J.; Croockewit, A.J.; de Wilt, J.H.; van Rossum, M.M.; Blokx, W.A.; Jacobs, J.F.; Duiveman-de Boer, T.; et al. Vaccination with mRNA-electroporated dendritic cells induces robust tumor antigen-specific CD4+ and CD8+ T cells responses in stage III and IV melanoma patients. Clin. Cancer Res. 2012, 18, 5460–5470. [Google Scholar] [CrossRef] [PubMed]
- Dannull, J.; Haley, N.R.; Archer, G.; Nair, S.; Boczkowski, D.; Harper, M.; De Rosa, N.; Pickett, N.; Mosca, P.J.; Burchette, J.; et al. Melanoma immunotherapy using mature DCS expressing the constitutive proteasome. J. Clin. Investig. 2013, 123, 3135–3145. [Google Scholar] [CrossRef] [PubMed]
- Heiser, A.; Maurice, M.A.; Yancey, D.R.; Wu, N.Z.; Dahm, P.; Pruitt, S.K.; Boczkowski, D.; Nair, S.K.; Ballo, M.S.; Gilboa, E.; et al. Induction of polyclonal prostate cancer-specific CTL using dendritic cells transfected with amplified tumor RNA. J. Immunol. 2001, 166, 2953–2960. [Google Scholar] [CrossRef] [PubMed]
- Nair, S.K.; Morse, M.; Boczkowski, D.; Cumming, R.I.; Vasovic, L.; Gilboa, E.; Lyerly, H.K. Induction of tumor-specific cytotoxic T lymphocytes in cancer patients by autologous tumor RNA-transfected dendritic cells. Ann. Surg. 2002, 235, 540–549. [Google Scholar] [CrossRef] [PubMed]
- Schaft, N.; Wellner, V.; Wohn, C.; Schuler, G.; Dorrie, J. CD8(+) T-cell priming and boosting: More antigen-presenting DC, or more antigen per DC? Cancer Immunol. Immunother. 2013, 62, 1769–1780. [Google Scholar] [CrossRef] [PubMed]
- Schuurhuis, D.H.; Verdijk, P.; Schreibelt, G.; Aarntzen, E.H.; Scharenborg, N.; de Boer, A.; van de Rakt, M.W.; Kerkhoff, M.; Gerritsen, M.J.; Eijckeler, F.; et al. In situ expression of tumor antigens by messenger RNA-electroporated dendritic cells in lymph nodes of melanoma patients. Cancer Res. 2009, 69, 2927–2934. [Google Scholar] [CrossRef] [PubMed]
- Van Craenenbroeck, A.H.; Smits, E.L.; Anguille, S.; Van de Velde, A.; Stein, B.; Braeckman, T.; Van Camp, K.; Nijs, G.; Ieven, M.; Goossens, H.; et al. Induction of cytomegalovirus-specific T cell responses in healthy volunteers and allogeneic stem cell recipients using vaccination with messenger RNA-transfected dendritic cells. Transplantation 2015, 99, 120–127. [Google Scholar] [CrossRef] [PubMed]
- Bagley, K.C.; Lewis, G.K.; Fouts, T.R. Adjuvant activity of the catalytic a1 domain of cholera toxin for retroviral antigens delivered by genegun. Clin. Vaccine Immunol. 2011, 18, 922–930. [Google Scholar] [CrossRef] [PubMed]
- Condon, C.; Watkins, S.C.; Celluzzi, C.M.; Thompson, K.; Falo, L.D., Jr. DNA-based immunization by in vivo transfection of dendritic cells. Nat. Med. 1996, 2, 1122–1128. [Google Scholar] [CrossRef] [PubMed]
- Harrison, R.A.; Richards, A.; Laing, G.D.; Theakston, R.D. Simultaneous genegun immunisation with plasmids encoding antigen and GM-CSF: Significant enhancement of murine antivenom IgG1 titres. Vaccine 2002, 20, 1702–1706. [Google Scholar] [CrossRef]
- Whiteside, T.L.; Gambotto, A.; Albers, A.; Stanson, J.; Cohen, E.P. Human tumor-derived genomic DNA transduced into a recipient cell induces tumor-specific immune responses ex vivo. Proc. Nat. Acad. Sci. USA 2002, 99, 9415–9420. [Google Scholar] [CrossRef] [PubMed]
- Vik-Mo, E.O.; Nyakas, M.; Mikkelsen, B.V.; Moe, M.C.; Due-Tonnesen, P.; Suso, E.M.; Saeboe-Larssen, S.; Sandberg, C.; Brinchmann, J.E.; Helseth, E.; et al. Therapeutic vaccination against autologous cancer stem cells with mRNA-transfected dendritic cells in patients with glioblastoma. Cancer Immunol. Immunother. 2013, 62, 1499–1509. [Google Scholar] [CrossRef] [PubMed]
- Hammad, H.; Lambrecht, B.N. Dendritic cells and epithelial cells: Linking innate and adaptive immunity in asthma. Nat. Rev. Immunol. 2008, 8, 193–204. [Google Scholar] [CrossRef] [PubMed]
- Kissick, H.T.; Sanda, M.G. The role of active vaccination in cancer immunotherapy: Lessons from clinical trials. Curr. Opin. Immunol. 2015, 35, 15–22. [Google Scholar] [CrossRef] [PubMed]
- Nestle, F.O.; Banchereau, J.; Hart, D. Dendritic cells: On the move from bench to bedside. Nat. Med. 2001, 7, 761–765. [Google Scholar] [CrossRef] [PubMed]
- Mayanagi, S.; Kitago, M.; Sakurai, T.; Matsuda, T.; Fujita, T.; Higuchi, H.; Taguchi, J.; Takeuchi, H.; Itano, O.; Aiura, K.; et al. Phase I pilot study of wilms tumor gene 1 peptide-pulsed dendritic cell vaccination combined with gemcitabine in pancreatic cancer. Cancer Sci. 2015, 106, 397–406. [Google Scholar] [CrossRef] [PubMed]
- Mitchell, D.A.; Batich, K.A.; Gunn, M.D.; Huang, M.N.; Sanchez-Perez, L.; Nair, S.K.; Congdon, K.L.; Reap, E.A.; Archer, G.E.; Desjardins, A.; et al. Tetanus toxoid and CCL3 improve dendritic cell vaccines in mice and glioblastoma patients. Nature 2015, 519, 366–369. [Google Scholar] [CrossRef] [PubMed]
- Chodon, T.; Comin-Anduix, B.; Chmielowski, B.; Koya, R.C.; Wu, Z.; Auerbach, M.; Ng, C.; Avramis, E.; Seja, E.; Villanueva, A.; et al. Adoptive transfer of MART-1 T-cell receptor transgenic lymphocytes and dendritic cell vaccination in patients with metastatic melanoma. Clin. Cancer Res. 2014, 20, 2457–2465. [Google Scholar] [CrossRef] [PubMed]
- Dhodapkar, M.V.; Sznol, M.; Zhao, B.; Wang, D.; Carvajal, R.D.; Keohan, M.L.; Chuang, E.; Sanborn, R.E.; Lutzky, J.; Powderly, J.; et al. Induction of antigen-specific immunity with a vaccine targeting NY-ESO-1 to the dendritic cell receptor DEC-205. Sci. Transl. Med. 2014. [Google Scholar] [CrossRef] [PubMed]
- Nestle, F.O.; Alijagic, S.; Gilliet, M.; Sun, Y.; Grabbe, S.; Dummer, R.; Burg, G.; Schadendorf, D. Vaccination of melanoma patients with peptide—Or tumor lysate—Pulsed dendritic cells. Nat. Med. 1998, 4, 328–332. [Google Scholar] [CrossRef] [PubMed]
- Poschke, I.; Lovgren, T.; Adamson, L.; Nystrom, M.; Andersson, E.; Hansson, J.; Tell, R.; Masucci, G.V.; Kiessling, R. A phase I clinical trial combining dendritic cell vaccination with adoptive T cell transfer in patients with stage IV melanoma. Cancer Immunol. Immunother. 2014, 63, 1061–1071. [Google Scholar] [CrossRef] [PubMed]
- Thurner, B.; Haendle, I.; Roder, C.; Dieckmann, D.; Keikavoussi, P.; Jonuleit, H.; Bender, A.; Maczek, C.; Schreiner, D.; von den Driesch, P.; et al. Vaccination with MAGE-3A1 peptide-pulsed mature, monocyte-derived dendritic cells expands specific cytotoxic T cells and induces regression of some metastases in advanced stage IV melanoma. J. Exp. Med. 1999, 190, 1669–1678. [Google Scholar] [CrossRef] [PubMed]
- Aarntzen, E.H.; De Vries, I.J.; Lesterhuis, W.J.; Schuurhuis, D.; Jacobs, J.F.; Bol, K.; Schreibelt, G.; Mus, R.; De Wilt, J.H.; Haanen, J.B.; et al. Targeting CD4(+) T-helper cells improves the induction of antitumor responses in dendritic cell-based vaccination. Cancer Res. 2013, 73, 19–29. [Google Scholar] [CrossRef] [PubMed]
- Engell-Noerregaard, L.; Hansen, T.H.; Andersen, M.H.; Thor Straten, P.; Svane, I.M. Review of clinical studies on dendritic cell-based vaccination of patients with malignant melanoma: Assessment of correlation between clinical response and vaccine parameters. Cancer Immunol. Immunother. 2009, 58, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Hobo, W.; Strobbe, L.; Maas, F.; Fredrix, H.; Greupink-Draaisma, A.; Esendam, B.; de Witte, T.; Preijers, F.; Levenga, H.; van Rees, B.; et al. Immunogenicity of dendritic cells pulsed with MAGE3, survivin and B-cell maturation antigen mRNA for vaccination of multiple myeloma patients. Cancer Immunol. Immunother. 2013, 62, 1381–1392. [Google Scholar] [CrossRef] [PubMed]
- Murthy, V.; Moiyadi, A.; Sawant, R.; Sarin, R. Clinical considerations in developing dendritic cell vaccine based immunotherapy protocols in cancer. Curr. Mol. Med. 2009, 9, 725–731. [Google Scholar] [CrossRef] [PubMed]
- Butterfield, L.H. Cancer vaccines. BMJ 2015. [Google Scholar] [CrossRef] [PubMed]
- Mahoney, K.M.; Rennert, P.D.; Freeman, G.J. Combination cancer immunotherapy and new immunomodulatory targets. Nat. Rev. Drug Discov. 2015, 14, 561–584. [Google Scholar] [CrossRef] [PubMed]
- Pizzurro, G.A.; Barrio, M.M. Dendritic cell-based vaccine efficacy: Aiming for hot spots. Front. Immunol. 2015. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Melero, I.; Gaudernack, G.; Gerritsen, W.; Huber, C.; Parmiani, G.; Scholl, S.; Thatcher, N.; Wagstaff, J.; Zielinski, C.; Faulkner, I.; et al. Therapeutic vaccines for cancer: An overview of clinical trials. Nat. Rev. Clin. Oncol. 2014, 11, 509–524. [Google Scholar] [CrossRef] [PubMed]
- Hodi, F.S.; O’Day, S.J.; McDermott, D.F.; Weber, R.W.; Sosman, J.A.; Haanen, J.B.; Gonzalez, R.; Robert, C.; Schadendorf, D.; Hassel, J.C.; et al. Improved survival with ipilimumab in patients with metastatic melanoma. N. Engl. J. Med. 2010, 363, 711–723. [Google Scholar] [CrossRef] [PubMed]
- Lawson, D.H.; Lee, S.; Zhao, F.; Tarhini, A.A.; Margolin, K.A.; Ernstoff, M.S.; Atkins, M.B.; Cohen, G.I.; Whiteside, T.L.; Butterfield, L.H.; et al. Randomized, placebo-controlled, phase III trial of yeast-derived granulocyte-macrophage colony-stimulating factor (GM-CSF) versus peptide vaccination versus GM-CSF plus peptide vaccination versus placebo in patients with no evidence of disease after complete surgical resection of locally advanced and/or stage IV melanoma: A trial of the eastern cooperative oncology group-american college of radiology imaging network cancer research group (e4697). J. Clin. Oncol. 2015, 33, 4066–4076. [Google Scholar] [PubMed]
- Noguchi, M.; Moriya, F.; Suekane, S.; Matsuoka, K.; Arai, G.; Matsueda, S.; Sasada, T.; Yamada, A.; Itoh, K. Phase II study of personalized peptide vaccination for castration-resistant prostate cancer patients who failed in docetaxel-based chemotherapy. Prostate 2012, 72, 834–845. [Google Scholar] [CrossRef] [PubMed]
- Pol, J.; Bloy, N.; Buque, A.; Eggermont, A.; Cremer, I.; Sautes-Fridman, C.; Galon, J.; Tartour, E.; Zitvogel, L.; Kroemer, G.; et al. Trial watch: Peptide-based anticancer vaccines. Oncoimmunology 2015. [Google Scholar] [CrossRef] [PubMed]
- Schuster, J.; Lai, R.K.; Recht, L.D.; Reardon, D.A.; Paleologos, N.A.; Groves, M.D.; Mrugala, M.M.; Jensen, R.; Baehring, J.M.; Sloan, A.; et al. A phase ii, multicenter trial of rindopepimut (CDX-110) in newly diagnosed glioblastoma: The act III study. Neuro-Oncology 2015, 17, 854–861. [Google Scholar] [CrossRef] [PubMed]
- Schwartzentruber, D.J.; Lawson, D.H.; Richards, J.M.; Conry, R.M.; Miller, D.M.; Treisman, J.; Gailani, F.; Riley, L.; Conlon, K.; Pockaj, B.; et al. Gp100 peptide vaccine and interleukin-2 in patients with advanced melanoma. N. Engl. J. Med. 2011, 364, 2119–2127. [Google Scholar] [CrossRef] [PubMed]
- McNeel, D.G.; Dunphy, E.J.; Davies, J.G.; Frye, T.P.; Johnson, L.E.; Staab, M.J.; Horvath, D.L.; Straus, J.; Alberti, D.; Marnocha, R.; et al. Safety and immunological efficacy of a DNA vaccine encoding prostatic acid phosphatase in patients with stage d0 prostate cancer. J. Clin. Oncol. 2009, 27, 4047–4054. [Google Scholar] [CrossRef] [PubMed]
- Pavlenko, M.; Roos, A.K.; Lundqvist, A.; Palmborg, A.; Miller, A.M.; Ozenci, V.; Bergman, B.; Egevad, L.; Hellstrom, M.; Kiessling, R.; et al. A phase I trial of DNA vaccination with a plasmid expressing prostate-specific antigen in patients with hormone-refractory prostate cancer. Br. J. Cancer 2004, 91, 688–694. [Google Scholar] [CrossRef] [PubMed]
- Fehres, C.M.; Kalay, H.; Bruijns, S.C.; Musaafir, S.A.; Ambrosini, M.; van Bloois, L.; van Vliet, S.J.; Storm, G.; Garcia-Vallejo, J.J.; van Kooyk, Y. Cross-presentation through langerin and DC-SIGN targeting requires different formulations of glycan-modified antigens. J. Control. Release 2015, 203, 67–76. [Google Scholar] [CrossRef] [PubMed]
- Mansourian, M.; Badiee, A.; Jalali, S.A.; Shariat, S.; Yazdani, M.; Amin, M.; Jaafari, M.R. Effective induction of anti-tumor immunity using p5 HER-2/neu derived peptide encapsulated in fusogenic DOTAP cationic liposomes co-administrated with CpG-ODN. Immunol. Lett. 2014, 162, 87–93. [Google Scholar] [CrossRef] [PubMed]
- Varypataki, E.M.; van der Maaden, K.; Bouwstra, J.; Ossendorp, F.; Jiskoot, W. Cationic liposomes loaded with a synthetic long peptide and poly(i:C): A defined adjuvanted vaccine for induction of antigen-specific T cell cytotoxicity. AAPS J. 2015, 17, 216–226. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.L.; Park, K.; Soo, R.A.; Sun, Y.; Tyroller, K.; Wages, D.; Ely, G.; Yang, J.C.; Mok, T. Inspire: A phase III study of the BLP25 liposome vaccine (l-BLP25) in asian patients with unresectable stage III non-small cell lung cancer. BMC Cancer 2011. [Google Scholar] [CrossRef] [PubMed]
- Berd, D. M-vax: An autologous, hapten-modified vaccine for human cancer. Expert Rev. Vaccines 2004, 3, 521–527. [Google Scholar] [CrossRef] [PubMed]
- Hersey, P.; Coates, A.S.; McCarthy, W.H.; Thompson, J.F.; Sillar, R.W.; McLeod, R.; Gill, P.G.; Coventry, B.J.; McMullen, A.; Dillon, H.; et al. Adjuvant immunotherapy of patients with high-risk melanoma using vaccinia viral lysates of melanoma: Results of a randomized trial. J. Clin. Oncol. 2002, 20, 4181–4190. [Google Scholar] [CrossRef] [PubMed]
- Sondak, V.K.; Liu, P.Y.; Tuthill, R.J.; Kempf, R.A.; Unger, J.M.; Sosman, J.A.; Thompson, J.A.; Weiss, G.R.; Redman, B.G.; Jakowatz, J.G.; et al. Adjuvant immunotherapy of resected, intermediate-thickness, node-negative melanoma with an allogeneic tumor vaccine: Overall results of a randomized trial of the southwest oncology group. J. Clin. Oncol. 2002, 20, 2058–2066. [Google Scholar] [CrossRef] [PubMed]
- Bijker, M.S.; Melief, C.J.; Offringa, R.; van der Burg, S.H. Design and development of synthetic peptide vaccines: Past, present and future. Expert Rev. Vaccines 2007, 6, 591–603. [Google Scholar] [CrossRef] [PubMed]
- Vremec, D.; O’Keeffe, M.; Wilson, A.; Ferrero, I.; Koch, U.; Radtke, F.; Scott, B.; Hertzog, P.; Villadangos, J.; Shortman, K. Factors determining the spontaneous activation of splenic dendritic cells in culture. Innate Immun. 2011, 17, 338–352. [Google Scholar] [CrossRef] [PubMed]
- Kamphorst, A.O.; Guermonprez, P.; Dudziak, D.; Nussenzweig, M.C. Route of antigen uptake differentially impacts presentation by dendritic cells and activated monocytes. J. Immunol. 2010, 185, 3426–3435. [Google Scholar] [CrossRef] [PubMed]
- Joshi, M.D.; Unger, W.J.; Storm, G.; van Kooyk, Y.; Mastrobattista, E. Targeting tumor antigens to dendritic cells using particulate carriers. J. Control. Release 2012, 161, 25–37. [Google Scholar] [CrossRef] [PubMed]
- Paulis, L.E.; Mandal, S.; Kreutz, M.; Figdor, C.G. Dendritic cell-based nanovaccines for cancer immunotherapy. Curr. Opin. Immunol. 2013, 25, 389–395. [Google Scholar] [CrossRef] [PubMed]
- Reddy, S.T.; Swartz, M.A.; Hubbell, J.A. Targeting dendritic cells with biomaterials: Developing the next generation of vaccines. Trends Immunol. 2006, 27, 573–579. [Google Scholar] [CrossRef] [PubMed]
- Ducancel, F.; Muller, B.H. Molecular engineering of antibodies for therapeutic and diagnostic purposes. MAbs 2012, 4, 445–457. [Google Scholar] [CrossRef] [PubMed]
- Feige, M.J.; Buchner, J. Principles and engineering of antibody folding and assembly. Biochim. Biophys. Acta 2014, 1844, 2024–2031. [Google Scholar] [CrossRef] [PubMed]
- Lazar, G.A.; Dang, W.; Karki, S.; Vafa, O.; Peng, J.S.; Hyun, L.; Chan, C.; Chung, H.S.; Eivazi, A.; Yoder, S.C.; et al. Engineered antibody fc variants with enhanced effector function. Proc. Nat. Acad. Sci. USA 2006, 103, 4005–4010. [Google Scholar] [CrossRef] [PubMed]
- Li, F.; Ravetch, J.V. Inhibitory fcgamma receptor engagement drives adjuvant and anti-tumor activities of agonistic CD40 antibodies. Science 2011, 333, 1030–1034. [Google Scholar] [CrossRef] [PubMed]
- Lux, A.; Nimmerjahn, F. Impact of differential glycosylation on IgG activity. Adv. Exp. Med. Biol. 2011, 780, 113–124. [Google Scholar] [PubMed]
- Nimmerjahn, F.; Ravetch, J.V. Translating basic mechanisms of IgG effector activity into next generation cancer therapies. Cancer Immun. 2012, 12, 13. [Google Scholar] [PubMed]
- Roopenian, D.C.; Akilesh, S. Fcrn: The neonatal fc receptor comes of age. Nat. Rev. Immunol. 2007, 7, 715–725. [Google Scholar] [CrossRef] [PubMed]
- Ward, E.S.; Devanaboyina, S.C.; Ober, R.J. Targeting fcrn for the modulation of antibody dynamics. Mol. Immunol. 2015, 67, 131–141. [Google Scholar] [CrossRef] [PubMed]
- Dong, H.; Stanek, O.; Salvador, F.R.; Langer, U.; Morillon, E.; Ung, C.; Sebo, P.; Leclerc, C.; Majlessi, L. Induction of protective immunity against mycobacterium tuberculosis by delivery of esx antigens into airway dendritic cells. Mucosal Immunol. 2013, 6, 522–534. [Google Scholar] [CrossRef] [PubMed]
- Helft, J.; Ginhoux, F.; Bogunovic, M.; Merad, M. Origin and functional heterogeneity of non-lymphoid tissue dendritic cells in mice. Immunol. Rev. 2010, 234, 55–75. [Google Scholar] [CrossRef] [PubMed]
- Hadeiba, H.; Sato, T.; Habtezion, A.; Oderup, C.; Pan, J.; Butcher, E.C. CCR9 expression defines tolerogenic plasmacytoid dendritic cells able to suppress acute graft-versus-host disease. Nat. Immunol. 2008, 9, 1253–1260. [Google Scholar] [CrossRef] [PubMed]
- Wendland, M.; Czeloth, N.; Mach, N.; Malissen, B.; Kremmer, E.; Pabst, O.; Forster, R. CCR9 is a homing receptor for plasmacytoid dendritic cells to the small intestine. Proc. Nat. Acad. Sci. USA 2007, 104, 6347–6352. [Google Scholar] [CrossRef] [PubMed]
- Ardavin, C. Origin, precursors and differentiation of mouse dendritic cells. Nat. Rev. Immunol. 2003, 3, 582–590. [Google Scholar] [CrossRef] [PubMed]
- Cella, M.; Jarrossay, D.; Facchetti, F.; Alebardi, O.; Nakajima, H.; Lanzavecchia, A.; Colonna, M. Plasmacytoid monocytes migrate to inflamed lymph nodes and produce large amounts of type I interferon. Nat. Med. 1999, 5, 919–923. [Google Scholar] [PubMed]
- Gilliet, M.; Cao, W.; Liu, Y.J. Plasmacytoid dendritic cells: Sensing nucleic acids in viral infection and autoimmune diseases. Nat. Rev. Immunol. 2008, 8, 594–606. [Google Scholar] [CrossRef] [PubMed]
- Gregorio, J.; Meller, S.; Conrad, C.; Di Nardo, A.; Homey, B.; Lauerma, A.; Arai, N.; Gallo, R.L.; Digiovanni, J.; Gilliet, M. Plasmacytoid dendritic cells sense skin injury and promote wound healing through type I interferons. J. Exp. Med. 2010, 207, 2921–2930. [Google Scholar] [CrossRef] [PubMed]
- Rajagopal, D.; Paturel, C.; Morel, Y.; Uematsu, S.; Akira, S.; Diebold, S.S. Plasmacytoid dendritic cell-derived type I interferon is crucial for the adjuvant activity of toll-like receptor 7 agonists. Blood 2010, 115, 1949–1957. [Google Scholar] [CrossRef] [PubMed]
- Segura, E.; Wong, J.; Villadangos, J.A. Cutting edge: B220+CCR9− dendritic cells are not plasmacytoid dendritic cells but are precursors of conventional dendritic cells. J. Immunol. 2009, 183, 1514–1517. [Google Scholar] [CrossRef] [PubMed]
- Siegal, F.P.; Kadowaki, N.; Shodell, M.; Fitzgerald-Bocarsly, P.A.; Shah, K.; Ho, S.; Antonenko, S.; Liu, Y.J. The nature of the principal type 1 interferon-producing cells in human blood. Science 1999, 284, 1835–1837. [Google Scholar] [CrossRef] [PubMed]
- Chappell, C.P.; Giltiay, N.V.; Draves, K.E.; Chen, C.; Hayden-Ledbetter, M.S.; Shlomchik, M.J.; Kaplan, D.H.; Clark, E.A. Targeting antigens through blood dendritic cell antigen 2 on plasmacytoid dendritic cells promotes immunologic tolerance. J. Immunol. 2014, 192, 5789–5801. [Google Scholar] [CrossRef] [PubMed]
- Di Pucchio, T.; Chatterjee, B.; Smed-Sorensen, A.; Clayton, S.; Palazzo, A.; Montes, M.; Xue, Y.; Mellman, I.; Banchereau, J.; Connolly, J.E. Direct proteasome-independent cross-presentation of viral antigen by plasmacytoid dendritic cells on major histocompatibility complex class I. Nat. Immunol. 2008, 9, 551–557. [Google Scholar] [CrossRef] [PubMed]
- Flores, M.; Desai, D.D.; Downie, M.; Liang, B.; Reilly, M.P.; McKenzie, S.E.; Clynes, R. Dominant expression of the inhibitory fcgammariib prevents antigen presentation by murine plasmacytoid dendritic cells. J. Immunol. 2009, 183, 7129–7139. [Google Scholar] [CrossRef] [PubMed]
- Hoeffel, G.; Ripoche, A.C.; Matheoud, D.; Nascimbeni, M.; Escriou, N.; Lebon, P.; Heshmati, F.; Guillet, J.G.; Gannage, M.; Caillat-Zucman, S.; et al. Antigen crosspresentation by human plasmacytoid dendritic cells. Immunity 2007, 27, 481–492. [Google Scholar] [CrossRef] [PubMed]
- Loschko, J.; Heink, S.; Hackl, D.; Dudziak, D.; Reindl, W.; Korn, T.; Krug, A.B. Antigen targeting to plasmacytoid dendritic cells via Siglec-H inhibits Th cell-dependent autoimmunity. J. Immunol. 2011, 187, 6346–6356. [Google Scholar] [CrossRef] [PubMed]
- Loschko, J.; Schlitzer, A.; Dudziak, D.; Drexler, I.; Sandholzer, N.; Bourquin, C.; Reindl, W.; Krug, A.B. Antigen delivery to plasmacytoid dendritic cells via BST2 induces protective T cell-mediated immunity. J. Immunol. 2011, 186, 6718–6725. [Google Scholar] [CrossRef] [PubMed]
- Meyer-Wentrup, F.; Benitez-Ribas, D.; Tacken, P.J.; Punt, C.J.; Figdor, C.G.; de Vries, I.J.; Adema, G.J. Targeting DCIR on human plasmacytoid dendritic cells results in antigen presentation and inhibits IFN-alpha production. Blood 2008, 111, 4245–4253. [Google Scholar] [CrossRef] [PubMed]
- Mouries, J.; Moron, G.; Schlecht, G.; Escriou, N.; Dadaglio, G.; Leclerc, C. Plasmacytoid dendritic cells efficiently cross-prime naive T cells in vivo after TLR activation. Blood 2008, 112, 3713–3722. [Google Scholar] [CrossRef] [PubMed]
- Reizis, B.; Bunin, A.; Ghosh, H.S.; Lewis, K.L.; Sisirak, V. Plasmacytoid dendritic cells: Recent progress and open questions. Ann. Rev. Immunol. 2011, 29, 163–183. [Google Scholar] [CrossRef] [PubMed]
- Villadangos, J.A.; Schnorrer, P. Intrinsic and cooperative antigen-presenting functions of dendritic-cell subsets in vivo. Nat. Rev. Immunol. 2007, 7, 543–555. [Google Scholar] [CrossRef] [PubMed]
- Gurka, S.; Hartung, E.; Becker, M.; Kroczek, R.A. Mouse conventional dendritic cells can be universally classified based on the mutually exclusive expression of XCR1 and sirpalpha. Front. Immunol. 2015. [Google Scholar] [CrossRef] [PubMed]
- Miller, J.C.; Brown, B.D.; Shay, T.; Gautier, E.L.; Jojic, V.; Cohain, A.; Pandey, G.; Leboeuf, M.; Elpek, K.G.; Helft, J.; et al. Deciphering the transcriptional network of the dendritic cell lineage. Nat. Immunol. 2012, 13, 888–899. [Google Scholar] [CrossRef] [PubMed]
- Malissen, B.; Tamoutounour, S.; Henri, S. The origins and functions of dendritic cells and macrophages in the skin. Nat. Rev. Immunol. 2014, 14, 417–428. [Google Scholar] [CrossRef] [PubMed]
- Langlet, C.; Tamoutounour, S.; Henri, S.; Luche, H.; Ardouin, L.; Gregoire, C.; Malissen, B.; Guilliams, M. CD64 expression distinguishes monocyte-derived and conventional dendritic cells and reveals their distinct role during intramuscular immunization. J. Immunol. 2012, 188, 1751–1760. [Google Scholar] [CrossRef] [PubMed]
- Durand, M.; Segura, E. The known unknowns of the human dendritic cell network. Front. Immunol. 2015. [Google Scholar] [CrossRef] [PubMed]
- Guilliams, M.; Ginhoux, F.; Jakubzick, C.; Naik, S.H.; Onai, N.; Schraml, B.U.; Segura, E.; Tussiwand, R.; Yona, S. Dendritic cells, monocytes and macrophages: A unified nomenclature based on ontogeny. Nat. Rev. Immunol. 2014, 14, 571–578. [Google Scholar] [CrossRef] [PubMed]
- Heidkamp, G.F.; Lehmann, C.H.K.; Heger, L.; Baransk, A.; Hoffmann, A.; Lühr, J.; Dudziak, D. Functional specialization of dendritic cell subsets. In Encyclopedia of Cell Biology, 1st ed.; Academic Press: Waltham, UK, 2016; pp. 588–604. [Google Scholar]
- Palucka, K.; Banchereau, J. Dendritic-cell-based therapeutic cancer vaccines. Immunity 2013, 39, 38–48. [Google Scholar] [CrossRef] [PubMed]
- Reynolds, G.; Haniffa, M. Human and mouse mononuclear phagocyte networks: A tale of two species? Front. Immunol. 2015. [Google Scholar] [CrossRef] [PubMed]
- Vu Manh, T.P.; Bertho, N.; Hosmalin, A.; Schwartz-Cornil, I.; Dalod, M. Investigating evolutionary conservation of dendritic cell subset identity and functions. Front. Immunol. 2015. [Google Scholar] [CrossRef] [PubMed]
- Bonifaz, L.C.; Bonnyay, D.P.; Charalambous, A.; Darguste, D.I.; Fujii, S.; Soares, H.; Brimnes, M.K.; Moltedo, B.; Moran, T.M.; Steinman, R.M. In vivo targeting of antigens to maturing dendritic cells via the DEC-205 receptor improves T cell vaccination. J. Exp. Med. 2004, 199, 815–824. [Google Scholar] [CrossRef] [PubMed]
- Lindquist, R.L.; Shakhar, G.; Dudziak, D.; Wardemann, H.; Eisenreich, T.; Dustin, M.L.; Nussenzweig, M.C. Visualizing dendritic cell networks in vivo. Nat. Immunol. 2004, 5, 1243–1250. [Google Scholar] [CrossRef] [PubMed]
- Shakhar, G.; Lindquist, R.L.; Skokos, D.; Dudziak, D.; Huang, J.H.; Nussenzweig, M.C.; Dustin, M.L. Stable T cell-dendritic cell interactions precede the development of both tolerance and immunity in vivo. Nat. Immunol. 2005, 6, 707–714. [Google Scholar] [CrossRef] [PubMed]
- Soares, H.; Waechter, H.; Glaichenhaus, N.; Mougneau, E.; Yagita, H.; Mizenina, O.; Dudziak, D.; Nussenzweig, M.C.; Steinman, R.M. A subset of dendritic cells induces CD4+ T cells to produce IFN-gamma by an IL-12-independent but CD70-dependent mechanism in vivo. J. Exp. Med. 2007, 204, 1095–1106. [Google Scholar] [CrossRef] [PubMed]
- Jiang, W.; Swiggard, W.J.; Heufler, C.; Peng, M.; Mirza, A.; Steinman, R.M.; Nussenzweig, M.C. The receptor DEC-205 expressed by dendritic cells and thymic epithelial cells is involved in antigen processing. Nature 1995, 375, 151–155. [Google Scholar] [CrossRef] [PubMed]
- Inaba, K.; Swiggard, W.J.; Inaba, M.; Meltzer, J.; Mirza, A.; Sasagawa, T.; Nussenzweig, M.C.; Steinman, R.M. Tissue distribution of the DEC-205 protein that is detected by the monoclonal antibody NLDC-145. I. Expression on dendritic cells and other subsets of mouse leukocytes. Cell. Immunol. 1995, 163, 148–156. [Google Scholar] [CrossRef] [PubMed]
- Kraal, G.; Breel, M.; Janse, M.; Bruin, G. Langerhans’ cells, veiled cells, and interdigitating cells in the mouse recognized by a monoclonal antibody. J. Exp. Med. 1986, 163, 981–997. [Google Scholar] [CrossRef] [PubMed]
- Witmer-Pack, M.D.; Swiggard, W.J.; Mirza, A.; Inaba, K.; Steinman, R.M. Tissue distribution of the DEC-205 protein that is detected by the monoclonal antibody NLDC-145. II. Expression in situ in lymphoid and nonlymphoid tissues. Cell. Immunol. 1995, 163, 157–162. [Google Scholar] [CrossRef] [PubMed]
- Mahnke, K.; Qian, Y.; Knop, J.; Enk, A.H. Induction of CD4+/CD25+ regulatory T cells by targeting of antigens to immature dendritic cells. Blood 2003, 101, 4862–4869. [Google Scholar] [CrossRef] [PubMed]
- Iyoda, T.; Shimoyama, S.; Liu, K.; Omatsu, Y.; Akiyama, Y.; Maeda, Y.; Takahara, K.; Steinman, R.M.; Inaba, K. The CD8+ dendritic cell subset selectively endocytoses dying cells in culture and in vivo. J. Exp. Med. 2002, 195, 1289–1302. [Google Scholar] [CrossRef] [PubMed]
- Lahoud, M.H.; Ahmet, F.; Zhang, J.G.; Meuter, S.; Policheni, A.N.; Kitsoulis, S.; Lee, C.N.; O’Keeffe, M.; Sullivan, L.C.; Brooks, A.G.; et al. DEC-205 is a cell surface receptor for CPG oligonucleotides. Proc. Nat. Acad. Sci. USA 2012, 109, 16270–16275. [Google Scholar] [CrossRef] [PubMed]
- Shrimpton, R.E.; Butler, M.; Morel, A.S.; Eren, E.; Hue, S.S.; Ritter, M.A. CD205 (DEC-205): A recognition receptor for apoptotic and necrotic self. Mol. Immunol. 2009, 46, 1229–1239. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.S.; Park, C.G.; Zhang, P.; Bartra, S.S.; Plano, G.V.; Klena, J.D.; Skurnik, M.; Hinnebusch, B.J.; Chen, T. Plasminogen activator Pla of Yersinia pestis utilizes murine DEC-205 (CD205) as a receptor to promote dissemination. J. Biol. Chem. 2008, 283, 31511–31521. [Google Scholar] [CrossRef] [PubMed]
- Heidkamp, G.F.; Neubert, K.; Haertel, E.; Nimmerjahn, F.; Nussenzweig, M.C.; Dudziak, D. Efficient generation of a monoclonal antibody against the human C-type lectin receptor DCIR by targeting murine dendritic cells. Immunol. Lett. 2010, 132, 69–78. [Google Scholar] [CrossRef] [PubMed]
- Henriques, H.R.; Rampazo, E.V.; Goncalves, A.J.; Vicentin, E.C.; Amorim, J.H.; Panatieri, R.H.; Amorim, K.N.; Yamamoto, M.M.; Ferreira, L.C.; Alves, A.M.; et al. Targeting the non-structural protein 1 from dengue virus to a dendritic cell population confers protective immunity to lethal virus challenge. PLoS Negl. Trop. Dis. 2013. [Google Scholar] [CrossRef] [PubMed]
- Idoyaga, J.; Fiorese, C.; Zbytnuik, L.; Lubkin, A.; Miller, J.; Malissen, B.; Mucida, D.; Merad, M.; Steinman, R.M. Specialized role of migratory dendritic cells in peripheral tolerance induction. J. Clin. Investig. 2013, 123, 844–854. [Google Scholar] [CrossRef] [PubMed]
- Bates, E.E.; Fournier, N.; Garcia, E.; Valladeau, J.; Durand, I.; Pin, J.J.; Zurawski, S.M.; Patel, S.; Abrams, J.S.; Lebecque, S.; et al. APCs express DCIR, a novel C-type lectin surface receptor containing an immunoreceptor tyrosine-based inhibitory motif. J. Immunol. 1999, 163, 1973–1983. [Google Scholar] [PubMed]
- Klechevsky, E.; Flamar, A.L.; Cao, Y.; Blanck, J.P.; Liu, M.; O’Bar, A.; Agouna-Deciat, O.; Klucar, P.; Thompson-Snipes, L.; Zurawski, S.; et al. Cross-priming CD8+ T cells by targeting antigens to human dendritic cells through DCIR. Blood 2010, 116, 1685–1697. [Google Scholar] [CrossRef] [PubMed]
- Bloem, K.; Vuist, I.M.; van den Berk, M.; Klaver, E.J.; van Die, I.; Knippels, L.M.; Garssen, J.; Garcia-Vallejo, J.J.; van Vliet, S.J.; van Kooyk, Y. DCIR interacts with ligands from both endogenous and pathogenic origin. Immunol. Lett. 2014, 158, 33–41. [Google Scholar] [CrossRef] [PubMed]
- Jin, W.; Li, C.; Du, T.; Hu, K.; Huang, X.; Hu, Q. DC-SIGN plays a stronger role than DCIR in mediating HIV-1 capture and transfer. Virology 2014. [Google Scholar] [CrossRef] [PubMed]
- Lambert, A.A.; Gilbert, C.; Richard, M.; Beaulieu, A.D.; Tremblay, M.J. The C-type lectin surface receptor DCIR acts as a new attachment factor for HIV-1 in dendritic cells and contributes to trans- and cis-infection pathways. Blood 2008, 112, 1299–1307. [Google Scholar] [CrossRef] [PubMed]
- Idoyaga, J.; Cheong, C.; Suda, K.; Suda, N.; Kim, J.Y.; Lee, H.; Park, C.G.; Steinman, R.M. Cutting edge: Langerin/CD207 receptor on dendritic cells mediates efficient antigen presentation on MHC I and II products in vivo. J. Immunol. 2008, 180, 3647–3650. [Google Scholar] [CrossRef] [PubMed]
- Neubert, K.; Lehmann, C.H.; Heger, L.; Baranska, A.; Staedtler, A.M.; Buchholz, V.R.; Yamazaki, S.; Heidkamp, G.F.; Eissing, N.; Zebroski, H.; et al. Antigen delivery to CD11c+CD8− dendritic cells induces protective immune responses against experimental melanoma in mice in vivo. J. Immunol. 2014, 192, 5830–5838. [Google Scholar] [CrossRef] [PubMed]
- Boscardin, S.B.; Hafalla, J.C.; Masilamani, R.F.; Kamphorst, A.O.; Zebroski, H.A.; Rai, U.; Morrot, A.; Zavala, F.; Steinman, R.M.; Nussenzweig, R.S.; et al. Antigen targeting to dendritic cells elicits long-lived T cell help for antibody responses. J. Exp. Med. 2006, 203, 599–606. [Google Scholar] [CrossRef] [PubMed]
- Macho-Fernandez, E.; Cruz, L.J.; Ghinnagow, R.; Fontaine, J.; Bialecki, E.; Frisch, B.; Trottein, F.; Faveeuw, C. Targeted delivery of α-galactosylceramide to CD8α+ dendritic cells optimizes type I NKT cell-based antitumor responses. J. Immunol. 2014, 193, 961–969. [Google Scholar] [CrossRef] [PubMed]
- Do, Y.; Koh, H.; Park, C.G.; Dudziak, D.; Seo, P.; Mehandru, S.; Choi, J.H.; Cheong, C.; Park, S.; Perlin, D.S.; et al. Targeting of LCRV virulence protein from yersinia pestis to dendritic cells protects mice against pneumonic plague. Eur. J. Immunol. 2010, 40, 2791–2796. [Google Scholar] [CrossRef] [PubMed]
- Trumpfheller, C.; Finke, J.S.; Lopez, C.B.; Moran, T.M.; Moltedo, B.; Soares, H.; Huang, Y.; Schlesinger, S.J.; Park, C.G.; Nussenzweig, M.C.; et al. Intensified and protective CD4+ T cell immunity in mice with anti-dendritic cell HIV gag fusion antibody vaccine. J. Exp. Med. 2006, 203, 607–617. [Google Scholar] [CrossRef] [PubMed]
- Idoyaga, J.; Lubkin, A.; Fiorese, C.; Lahoud, M.H.; Caminschi, I.; Huang, Y.; Rodriguez, A.; Clausen, B.E.; Park, C.G.; Trumpfheller, C.; et al. Comparable T helper 1 (Th1) and CD8 T-cell immunity by targeting HIV gag p24 to CD8 dendritic cells within antibodies to langerin, DEC205, and Clec9A. Proc. Nat. Acad. Sci. USA 2011, 108, 2384–2389. [Google Scholar] [CrossRef] [PubMed]
- Longhi, M.P.; Trumpfheller, C.; Idoyaga, J.; Caskey, M.; Matos, I.; Kluger, C.; Salazar, A.M.; Colonna, M.; Steinman, R.M. Dendritic cells require a systemic type I interferon response to mature and induce CD4+ Th1 immunity with poly IC as adjuvant. J. Exp. Med. 2009, 206, 1589–1602. [Google Scholar] [CrossRef] [PubMed]
- Ruane, D.; Chorny, A.; Lee, H.; Faith, J.; Pandey, G.; Shan, M.; Simchoni, N.; Rahman, A.; Garg, A.; Weinstein, E.G.; et al. Microbiota regulate the ability of lung dendritic cells to induce IgA class-switch recombination and generate protective gastrointestinal immune responses. J. Exp. Med. 2016, 213, 53–73. [Google Scholar] [CrossRef] [PubMed]
- Spiering, R.; Margry, B.; Keijzer, C.; Petzold, C.; Hoek, A.; Wagenaar-Hilbers, J.; van der Zee, R.; van Eden, W.; Kretschmer, K.; Broere, F. DEC205+ dendritic cell-targeted tolerogenic vaccination promotes immune tolerance in experimental autoimmune arthritis. J. Immunol. 2015, 194, 4804–4813. [Google Scholar] [CrossRef] [PubMed]
- Matos, I.; Mizenina, O.; Lubkin, A.; Steinman, R.M.; Idoyaga, J. Targeting antigens to dendritic cells in vivo induces protective immunity. PLoS ONE 2013, 8, e67453. [Google Scholar]
- Wang, B.; Zaidi, N.; He, L.Z.; Zhang, L.; Kuroiwa, J.M.; Keler, T.; Steinman, R.M. Targeting of the non-mutated tumor antigen HER2/neu to mature dendritic cells induces an integrated immune response that protects against breast cancer in mice. Breast Cancer Res. 2012. [Google Scholar] [CrossRef] [PubMed]
- Rampazo, E.V.; Amorim, K.N.; Yamamoto, M.M.; Panatieri, R.H.; Rodrigues, M.M.; Boscardin, S.B. Antigen targeting to dendritic cells allows the identification of a CD4 T-cell epitope within an immunodominant trypanosoma cruzi antigen. PLoS ONE 2015, 10, e0117778. [Google Scholar] [CrossRef] [PubMed]
- Mukhopadhaya, A.; Hanafusa, T.; Jarchum, I.; Chen, Y.G.; Iwai, Y.; Serreze, D.V.; Steinman, R.M.; Tarbell, K.V.; DiLorenzo, T.P. Selective delivery of beta cell antigen to dendritic cells in vivo leads to deletion and tolerance of autoreactive CD8+ T cells in nod mice. Proc. Nat. Acad. Sci. USA 2008, 105, 6374–6379. [Google Scholar] [CrossRef] [PubMed]
- Petzold, C.; Riewaldt, J.; Koenig, T.; Schallenberg, S.; Kretschmer, K. Dendritic cell-targeted pancreatic beta-cell antigen leads to conversion of self-reactive CD4(+) T cells into regulatory T cells and promotes immunotolerance in NOD mice. Rev Diabet. Stud. 2010, 7, 47–61. [Google Scholar] [CrossRef] [PubMed]
- Charalambous, A.; Oks, M.; Nchinda, G.; Yamazaki, S.; Steinman, R.M. Dendritic cell targeting of survivin protein in a xenogeneic form elicits strong CD4+ T cell immunity to mouse survivin. J. Immunol. 2006, 177, 8410–8421. [Google Scholar] [CrossRef] [PubMed]
- Mukherjee, G.; Geliebter, A.; Babad, J.; Santamaria, P.; Serreze, D.V.; Freeman, G.J.; Tarbell, K.V.; Sharpe, A.; DiLorenzo, T.P. DEC-205-mediated antigen targeting to steady-state dendritic cells induces deletion of diabetogenic CD8(+) T cells independently of PD-1 and PD-L1. Int. Immunol. 2013, 25, 651–660. [Google Scholar] [CrossRef] [PubMed]
- Stern, J.N.; Keskin, D.B.; Kato, Z.; Waldner, H.; Schallenberg, S.; Anderson, A.; von Boehmer, H.; Kretschmer, K.; Strominger, J.L. Promoting tolerance to proteolipid protein-induced experimental autoimmune encephalomyelitis through targeting dendritic cells. Proc. Nat. Acad. Sci. USA 2010, 107, 17280–17285. [Google Scholar] [CrossRef] [PubMed]
- Platt, C.D.; Ma, J.K.; Chalouni, C.; Ebersold, M.; Bou-Reslan, H.; Carano, R.A.; Mellman, I.; Delamarre, L. Mature dendritic cells use endocytic receptors to capture and present antigens. Proc. Nat. Acad. Sci. USA 2010, 107, 4287–4292. [Google Scholar] [CrossRef] [PubMed]
- Ring, S.; Maas, M.; Nettelbeck, D.M.; Enk, A.H.; Mahnke, K. Targeting of autoantigens to DEC205(+) dendritic cells in vivo suppresses experimental allergic encephalomyelitis in mice. J. Immunol. 2013, 191, 2938–2947. [Google Scholar] [CrossRef] [PubMed]
- Ettinger, M.; Gratz, I.K.; Gruber, C.; Hauser-Kronberger, C.; Johnson, T.S.; Mahnke, K.; Thalhamer, J.; Hintner, H.; Peckl-Schmid, D.; Bauer, J.W. Targeting of the hNC16A collagen domain to dendritic cells induces tolerance to human type XVII collagen. Exp. Dermatol. 2012, 21, 395–398. [Google Scholar] [PubMed]
- van Broekhoven, C.L.; Parish, C.R.; Demangel, C.; Britton, W.J.; Altin, J.G. Targeting dendritic cells with antigen-containing liposomes: A highly effective procedure for induction of antitumor immunity and for tumor immunotherapy. Cancer Res. 2004, 64, 4357–4365. [Google Scholar] [CrossRef] [PubMed]
- Bonifaz, L.; Bonnyay, D.; Mahnke, K.; Rivera, M.; Nussenzweig, M.C.; Steinman, R.M. Efficient targeting of protein antigen to the dendritic cell receptor DEC-205 in the steady state leads to antigen presentation on major histocompatibility complex class I products and peripheral CD8+ T cell tolerance. J. Exp. Med. 2002, 196, 1627–1638. [Google Scholar] [CrossRef] [PubMed]
- Flacher, V.; Tripp, C.H.; Haid, B.; Kissenpfennig, A.; Malissen, B.; Stoitzner, P.; Idoyaga, J.; Romani, N. Skin langerin+ dendritic cells transport intradermally injected anti-DEC-205 antibodies but are not essential for subsequent cytotoxic CD8+ T cell responses. J. Immunol. 2012, 188, 2146–2155. [Google Scholar] [CrossRef] [PubMed]
- Flacher, V.; Tripp, C.H.; Stoitzner, P.; Haid, B.; Ebner, S.; Del Frari, B.; Koch, F.; Park, C.G.; Steinman, R.M.; Idoyaga, J.; et al. Epidermal Langerhans cells rapidly capture and present antigens from C-type lectin-targeting antibodies deposited in the dermis. J. Investig. Dermatol. 2010, 130, 755–762. [Google Scholar] [CrossRef] [PubMed]
- Flynn, B.J.; Kastenmuller, K.; Wille-Reece, U.; Tomaras, G.D.; Alam, M.; Lindsay, R.W.; Salazar, A.M.; Perdiguero, B.; Gomez, C.E.; Wagner, R.; et al. Immunization with HIV gag targeted to dendritic cells followed by recombinant new york vaccinia virus induces robust T-cell immunity in nonhuman primates. Proc. Nat. Acad. Sci. USA 2011, 108, 7131–7136. [Google Scholar] [CrossRef] [PubMed]
- Gurer, C.; Strowig, T.; Brilot, F.; Pack, M.; Trumpfheller, C.; Arrey, F.; Park, C.G.; Steinman, R.M.; Munz, C. Targeting the nuclear antigen 1 of Epstein-Barr virus to the human endocytic receptor DEC-205 stimulates protective T-cell responses. Blood 2008, 112, 1231–1239. [Google Scholar] [CrossRef] [PubMed]
- Bozzacco, L.; Trumpfheller, C.; Siegal, F.P.; Mehandru, S.; Markowitz, M.; Carrington, M.; Nussenzweig, M.C.; Piperno, A.G.; Steinman, R.M. DEC-205 receptor on dendritic cells mediates presentation of HIV gag protein to CD8+ T cells in a spectrum of human MHC I haplotypes. Proc. Nat. Acad. Sci. USA 2007, 104, 1289–1294. [Google Scholar] [CrossRef] [PubMed]
- Tsuji, T.; Matsuzaki, J.; Kelly, M.P.; Ramakrishna, V.; Vitale, L.; He, L.Z.; Keler, T.; Odunsi, K.; Old, L.J.; Ritter, G.; et al. Antibody-targeted NY-ESO-1 to mannose receptor or DEC-205 in vitro elicits dual human CD8+ and CD4+ T cell responses with broad antigen specificity. J. Immunol. 2011, 186, 1218–1227. [Google Scholar] [CrossRef] [PubMed]
- Birkholz, K.; Schwenkert, M.; Kellner, C.; Gross, S.; Fey, G.; Schuler-Thurner, B.; Schuler, G.; Schaft, N.; Dorrie, J. Targeting of DEC-205 on human dendritic cells results in efficient mhc class II-restricted antigen presentation. Blood 2010, 116, 2277–2285. [Google Scholar] [CrossRef] [PubMed]
- White, A.L.; Tutt, A.L.; James, S.; Wilkinson, K.A.; Castro, F.V.; Dixon, S.V.; Hitchcock, J.; Khan, M.; Al-Shamkhani, A.; Cunningham, A.F.; et al. Ligation of CD11c during vaccination promotes germinal centre induction and robust humoral responses without adjuvant. Immunology 2010, 131, 141–151. [Google Scholar] [CrossRef] [PubMed]
- Sancho, D.; Mourao-Sa, D.; Joffre, O.P.; Schulz, O.; Rogers, N.C.; Pennington, D.J.; Carlyle, J.R.; Reis e Sousa, C. Tumor therapy in mice via antigen targeting to a novel, DC-restricted C-type lectin. J. Clin. Investig. 2008, 118, 2098–2110. [Google Scholar] [CrossRef] [PubMed]
- Joffre, O.P.; Sancho, D.; Zelenay, S.; Keller, A.M.; Reis e Sousa, C. Efficient and versatile manipulation of the peripheral CD4+ T-cell compartment by antigen targeting to DNGR-1/CLEC9A. Eur. J. Immunol. 2010, 40, 1255–1265. [Google Scholar] [CrossRef] [PubMed]
- Caminschi, I.; Proietto, A.I.; Ahmet, F.; Kitsoulis, S.; Shin Teh, J.; Lo, J.C.; Rizzitelli, A.; Wu, L.; Vremec, D.; van Dommelen, S.L.; et al. The dendritic cell subtype-restricted C-type lectin Clec9A is a target for vaccine enhancement. Blood 2008, 112, 3264–3273. [Google Scholar] [CrossRef] [PubMed]
- Lahoud, M.H.; Ahmet, F.; Kitsoulis, S.; Wan, S.S.; Vremec, D.; Lee, C.N.; Phipson, B.; Shi, W.; Smyth, G.K.; Lew, A.M.; et al. Targeting antigen to mouse dendritic cells via Clec9A induces potent CD4 T cell responses biased toward a follicular helper phenotype. J. Immunol. 2011, 187, 842–850. [Google Scholar] [CrossRef] [PubMed]
- Schreibelt, G.; Klinkenberg, L.J.; Cruz, L.J.; Tacken, P.J.; Tel, J.; Kreutz, M.; Adema, G.J.; Brown, G.D.; Figdor, C.G.; de Vries, I.J. The C-type lectin receptor Clec9A mediates antigen uptake and (cross−) presentation by human blood BDCA3+ myeloid dendritic cells. Blood 2012, 119, 2284–2292. [Google Scholar] [CrossRef] [PubMed]
- Lahoud, M.H.; Proietto, A.I.; Ahmet, F.; Kitsoulis, S.; Eidsmo, L.; Wu, L.; Sathe, P.; Pietersz, S.; Chang, H.W.; Walker, I.D.; et al. The C-type lectin clec12a present on mouse and human dendritic cells can serve as a target for antigen delivery and enhancement of antibody responses. J. Immunol. 2009, 182, 7587–7594. [Google Scholar] [CrossRef] [PubMed]
- Carter, R.W.; Thompson, C.; Reid, D.M.; Wong, S.Y.; Tough, D.F. Preferential induction of CD4+ T cell responses through in vivo targeting of antigen to dendritic cell-associated C-type lectin-1. J. Immunol. 2006, 177, 2276–2284. [Google Scholar] [CrossRef] [PubMed]
- Duluc, D.; Joo, H.; Ni, L.; Yin, W.; Upchurch, K.; Li, D.; Xue, Y.; Klucar, P.; Zurawski, S.; Zurawski, G.; et al. Induction and activation of human Th17 by targeting antigens to dendritic cells via Dectin-1. J. Immunol. 2014, 192, 5776–5788. [Google Scholar] [CrossRef] [PubMed]
- Ni, L.; Gayet, I.; Zurawski, S.; Duluc, D.; Flamar, A.L.; Li, X.H.; O’Bar, A.; Clayton, S.; Palucka, A.K.; Zurawski, G.; et al. Concomitant activation and antigen uptake via human Dectin-1 results in potent antigen-specific CD8+ T cell responses. J. Immunol. 2010, 185, 3504–3513. [Google Scholar] [CrossRef] [PubMed]
- Flacher, V.; Tripp, C.H.; Mairhofer, D.G.; Steinman, R.M.; Stoitzner, P.; Idoyaga, J.; Romani, N. Murine langerin+ dermal dendritic cells prime CD8+ T cells while langerhans cells induce cross-tolerance. EMBO Mol. Med. 2014, 6, 1191–1204. [Google Scholar] [CrossRef] [PubMed]
- Tacken, P.J.; de Vries, I.J.; Gijzen, K.; Joosten, B.; Wu, D.; Rother, R.P.; Faas, S.J.; Punt, C.J.; Torensma, R.; Adema, G.J.; et al. Effective induction of naive and recall T-cell responses by targeting antigen to human dendritic cells via a humanized anti-DC-SIGN antibody. Blood 2005, 106, 1278–1285. [Google Scholar] [CrossRef] [PubMed]
- Hesse, C.; Ginter, W.; Forg, T.; Mayer, C.T.; Baru, A.M.; Arnold-Schrauf, C.; Unger, W.W.; Kalay, H.; van Kooyk, Y.; Berod, L.; et al. In vivo targeting of human DC-SIGN drastically enhances CD8(+) T-cell-mediated protective immunity. Eur. J. Immunol. 2013, 43, 2543–2553. [Google Scholar] [CrossRef] [PubMed]
- Tacken, P.J.; Ginter, W.; Berod, L.; Cruz, L.J.; Joosten, B.; Sparwasser, T.; Figdor, C.G.; Cambi, A. Targeting DC-SIGN via its neck region leads to prolonged antigen residence in early endosomes, delayed lysosomal degradation, and cross-presentation. Blood 2011, 118, 4111–4119. [Google Scholar] [CrossRef] [PubMed]
- Pereira, C.F.; Torensma, R.; Hebeda, K.; Kretz-Rommel, A.; Faas, S.J.; Figdor, C.G.; Adema, G.J. In vivo targeting of DC-SIGN-positive antigen-presenting cells in a nonhuman primate model. J. Immunother. 2007, 30, 705–714. [Google Scholar] [CrossRef] [PubMed]
- Kretz-Rommel, A.; Qin, F.; Dakappagari, N.; Torensma, R.; Faas, S.; Wu, D.; Bowdish, K.S. In vivo targeting of antigens to human dendritic cells through DC-SIGN elicits stimulatory immune responses and inhibits tumor growth in grafted mouse models. J. Immunother. 2007, 30, 715–726. [Google Scholar] [CrossRef] [PubMed]
- He, L.Z.; Ramakrishna, V.; Connolly, J.E.; Wang, X.T.; Smith, P.A.; Jones, C.L.; Valkova-Valchanova, M.; Arunakumari, A.; Treml, J.F.; Goldstein, J.; et al. A novel human cancer vaccine elicits cellular responses to the tumor-associated antigen, human chorionic gonadotropin beta. Clin. Cancer Res. 2004, 10, 1920–1927. [Google Scholar] [CrossRef] [PubMed]
- Ramakrishna, V.; Treml, J.F.; Vitale, L.; Connolly, J.E.; O’Neill, T.; Smith, P.A.; Jones, C.L.; He, L.Z.; Goldstein, J.; Wallace, P.K.; et al. Mannose receptor targeting of tumor antigen pmel17 to human dendritic cells directs anti-melanoma T cell responses via multiple HLA molecules. J. Immunol. 2004, 172, 2845–2852. [Google Scholar] [CrossRef] [PubMed]
- Ramakrishna, V.; Vasilakos, J.P.; Tario, J.D., Jr.; Berger, M.A.; Wallace, P.K.; Keler, T. Toll-like receptor activation enhances cell-mediated immunity induced by an antibody vaccine targeting human dendritic cells. J. Transl. Med. 2007. [Google Scholar] [CrossRef] [PubMed]
- Castro, F.V.; Tutt, A.L.; White, A.L.; Teeling, J.L.; James, S.; French, R.R.; Glennie, M.J. CD11c provides an effective immunotarget for the generation of both CD4 and CD8 T cell responses. Eur. J. Immunol. 2008, 38, 2263–2273. [Google Scholar] [CrossRef] [PubMed]
- Wei, H.; Wang, S.; Zhang, D.; Hou, S.; Qian, W.; Li, B.; Guo, H.; Kou, G.; He, J.; Wang, H.; et al. Targeted delivery of tumor antigens to activated dendritic cells via CD11c molecules induces potent antitumor immunity in mice. Clin. Cancer Res. 2009, 15, 4612–4621. [Google Scholar] [CrossRef] [PubMed]
- Dickgreber, N.; Stoitzner, P.; Bai, Y.; Price, K.M.; Farrand, K.J.; Manning, K.; Angel, C.E.; Dunbar, P.R.; Ronchese, F.; Fraser, J.D.; et al. Targeting antigen to MHC class II molecules promotes efficient cross-presentation and enhances immunotherapy. J. Immunol. 2009, 182, 1260–1269. [Google Scholar] [CrossRef] [PubMed]
- Delneste, Y.; Magistrelli, G.; Gauchat, J.; Haeuw, J.; Aubry, J.; Nakamura, K.; Kawakami-Honda, N.; Goetsch, L.; Sawamura, T.; Bonnefoy, J.; et al. Involvement of LOX-1 in dendritic cell-mediated antigen cross-presentation. Immunity 2002, 17, 353–362. [Google Scholar] [CrossRef]
- Joo, H.; Li, D.; Dullaers, M.; Kim, T.W.; Duluc, D.; Upchurch, K.; Xue, Y.; Zurawski, S.; Le Grand, R.; Liu, Y.J.; et al. C-type lectin-like receptor LOX-1 promotes dendritic cell-mediated class-switched B cell responses. Immunity 2014, 41, 592–604. [Google Scholar] [CrossRef] [PubMed]
- Li, D.; Romain, G.; Flamar, A.L.; Duluc, D.; Dullaers, M.; Li, X.H.; Zurawski, S.; Bosquet, N.; Palucka, A.K.; Le Grand, R.; et al. Targeting self- and foreign antigens to dendritic cells via DC-asgpr generates IL-10-producing suppressive CD4+ T cells. J. Exp. Med. 2012, 209, 109–121. [Google Scholar] [CrossRef] [PubMed]
- Tagliani, E.; Guermonprez, P.; Sepulveda, J.; Lopez-Bravo, M.; Ardavin, C.; Amigorena, S.; Benvenuti, F.; Burrone, O.R. Selection of an antibody library identifies a pathway to induce immunity by targeting CD36 on steady-state CD8α+ dendritic cells. J. Immunol. 2008, 180, 3201–3209. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Raper, A.; Sugita, N.; Hingorani, R.; Salio, M.; Palmowski, M.J.; Cerundolo, V.; Crocker, P.R. Characterization of Siglec-H as a novel endocytic receptor expressed on murine plasmacytoid dendritic cell precursors. Blood 2006, 107, 3600–3608. [Google Scholar] [CrossRef] [PubMed]
- Sapoznikov, A.; Fischer, J.A.; Zaft, T.; Krauthgamer, R.; Dzionek, A.; Jung, S. Organ-dependent in vivo priming of naive CD4+, but not CD8+, T cells by plasmacytoid dendritic cells. J. Exp. Med. 2007, 204, 1923–1933. [Google Scholar] [CrossRef] [PubMed]
- Price, J.D.; Tarbell, K.V. The role of dendritic cell subsets and innate immunity in the pathogenesis of type 1 diabetes and other autoimmune diseases. Front. Immunol. 2015. [Google Scholar] [CrossRef] [PubMed]
- Ruane, D.; Do, Y.; Brane, L.; Garg, A.; Bozzacco, L.; Kraus, T.; Caskey, M.; Salazar, A.; Trumpheller, C.; Mehandru, S. A dendritic cell targeted vaccine induces long-term HIV-specific immunity within the gastrointestinal tract. Mucosal Immunol. 2016. [Google Scholar] [CrossRef] [PubMed]
- Engering, A.; Geijtenbeek, T.B.; van Vliet, S.J.; Wijers, M.; van Liempt, E.; Demaurex, N.; Lanzavecchia, A.; Fransen, J.; Figdor, C.G.; Piguet, V.; et al. The dendritic cell-specific adhesion receptor DC-SIGN internalizes antigen for presentation to T cells. J. Immunol. 2002, 168, 2118–2126. [Google Scholar] [CrossRef] [PubMed]
- Lai, W.K.; Sun, P.J.; Zhang, J.; Jennings, A.; Lalor, P.F.; Hubscher, S.; McKeating, J.A.; Adams, D.H. Expression of DC-SIGN and DC-SIGNR on human sinusoidal endothelium: A role for capturing hepatitis C virus particles. Am. J. Pathol. 2006, 169, 200–208. [Google Scholar] [CrossRef] [PubMed]
- Segura, E.; Valladeau-Guilemond, J.; Donnadieu, M.H.; Sastre-Garau, X.; Soumelis, V.; Amigorena, S. Characterization of resident and migratory dendritic cells in human lymph nodes. J. Exp. Med. 2012, 209, 653–660. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Vallejo, J.J.; van Kooyk, Y. The physiological role of DC-SIGN: A tale of mice and men. Trends Immunol. 2013, 34, 482–486. [Google Scholar] [CrossRef] [PubMed]
- Park, C.G.; Takahara, K.; Umemoto, E.; Yashima, Y.; Matsubara, K.; Matsuda, Y.; Clausen, B.E.; Inaba, K.; Steinman, R.M. Five mouse homologues of the human dendritic cell C-type lectin, DC-SIGN. Int. Immunol. 2001, 13, 1283–1290. [Google Scholar] [CrossRef] [PubMed]
- Powlesland, A.S.; Ward, E.M.; Sadhu, S.K.; Guo, Y.; Taylor, M.E.; Drickamer, K. Widely divergent biochemical properties of the complete set of mouse DC-SIGN-related proteins. J. Biol. Chem. 2006, 281, 20440–20449. [Google Scholar] [CrossRef] [PubMed]
- Schaefer, M.; Reiling, N.; Fessler, C.; Stephani, J.; Taniuchi, I.; Hatam, F.; Yildirim, A.O.; Fehrenbach, H.; Walter, K.; Ruland, J.; et al. Decreased pathology and prolonged survival of human DC-SIGN transgenic mice during mycobacterial infection. J. Immunol. 2008, 180, 6836–6845. [Google Scholar] [CrossRef] [PubMed]
- Caparros, E.; Munoz, P.; Sierra-Filardi, E.; Serrano-Gomez, D.; Puig-Kroger, A.; Rodriguez-Fernandez, J.L.; Mellado, M.; Sancho, J.; Zubiaur, M.; Corbi, A.L. DC-SIGN ligation on dendritic cells results in ERK and PI3K activation and modulates cytokine production. Blood 2006, 107, 3950–3958. [Google Scholar] [CrossRef] [PubMed]
- Bachem, A.; Guttler, S.; Hartung, E.; Ebstein, F.; Schaefer, M.; Tannert, A.; Salama, A.; Movassaghi, K.; Opitz, C.; Mages, H.W.; et al. Superior antigen cross-presentation and XCR1 expression define human CD11c+CD141+ cells as homologues of mouse CD8+ dendritic cells. J. Exp. Med. 2010, 207, 1273–1281. [Google Scholar] [CrossRef] [PubMed]
- Crozat, K.; Guiton, R.; Guilliams, M.; Henri, S.; Baranek, T.; Schwartz-Cornil, I.; Malissen, B.; Dalod, M. Comparative genomics as a tool to reveal functional equivalences between human and mouse dendritic cell subsets. Immunol. Rev. 2010, 234, 177–198. [Google Scholar] [CrossRef] [PubMed]
- Jongbloed, S.L.; Kassianos, A.J.; McDonald, K.J.; Clark, G.J.; Ju, X.; Angel, C.E.; Chen, C.J.; Dunbar, P.R.; Wadley, R.B.; Jeet, V.; et al. Human CD141+ (BDCA-3)+ dendritic cells (DCs) represent a unique myeloid DC subset that cross-presents necrotic cell antigens. J. Exp. Med. 2010, 207, 1247–1260. [Google Scholar] [CrossRef] [PubMed]
- Lundberg, K.; Rydnert, F.; Greiff, L.; Lindstedt, M. Human blood dendritic cell subsets exhibit discriminative pattern recognition receptor profiles. Immunology 2014, 142, 279–288. [Google Scholar] [CrossRef] [PubMed]
- Poulin, L.F.; Salio, M.; Griessinger, E.; Anjos-Afonso, F.; Craciun, L.; Chen, J.L.; Keller, A.M.; Joffre, O.; Zelenay, S.; Nye, E.; et al. Characterization of human DNGR-1+ BDCA3+ leukocytes as putative equivalents of mouse CD8α+ dendritic cells. J. Exp. Med. 2010, 207, 1261–1271. [Google Scholar] [CrossRef] [PubMed]
- Watchmaker, P.B.; Lahl, K.; Lee, M.; Baumjohann, D.; Morton, J.; Kim, S.J.; Zeng, R.; Dent, A.; Ansel, K.M.; Diamond, B.; et al. Comparative transcriptional and functional profiling defines conserved programs of intestinal DC differentiation in humans and mice. Nat. Immunol. 2014, 15, 98–108. [Google Scholar] [CrossRef] [PubMed]
- Chatterjee, B.; Smed-Sorensen, A.; Cohn, L.; Chalouni, C.; Vandlen, R.; Lee, B.C.; Widger, J.; Keler, T.; Delamarre, L.; Mellman, I. Internalization and endosomal degradation of receptor-bound antigens regulate the efficiency of cross presentation by human dendritic cells. Blood 2012, 120, 2011–2020. [Google Scholar] [CrossRef] [PubMed]
- Keler, T.; Ramakrishna, V.; Fanger, M.W. Mannose receptor-targeted vaccines. Expert Opin. Biol. Ther. 2004, 4, 1953–1962. [Google Scholar] [CrossRef] [PubMed]
- Drummond, R.A.; Brown, G.D. Signalling C-type lectins in antimicrobial immunity. PLoS Pathog. 2013, 9. [Google Scholar] [CrossRef] [PubMed]
- Killick, K.E.; Ni Cheallaigh, C.; O’Farrelly, C.; Hokamp, K.; MacHugh, D.E.; Harris, J. Receptor-mediated recognition of mycobacterial pathogens. Cell. Microbiol. 2013, 15, 1484–1495. [Google Scholar] [CrossRef] [PubMed]
- Willment, J.A.; Marshall, A.S.; Reid, D.M.; Williams, D.L.; Wong, S.Y.; Gordon, S.; Brown, G.D. The human beta-glucan receptor is widely expressed and functionally equivalent to murine Dectin-1 on primary cells. Eur. J. Immunol. 2005, 35, 1539–1547. [Google Scholar] [CrossRef] [PubMed]
- Ruland, J. CARD9 signaling in the innate immune response. Ann. N. Y. Acad. Sci. 2008, 1143, 35–44. [Google Scholar] [CrossRef] [PubMed]
- Robinson, M.J.; Sancho, D.; Slack, E.C.; LeibundGut-Landmann, S.; Reis e Sousa, C. Myeloid C-type lectins in innate immunity. Nat. Immunol. 2006, 7, 1258–1265. [Google Scholar] [CrossRef] [PubMed]
- Reid, D.M.; Montoya, M.; Taylor, P.R.; Borrow, P.; Gordon, S.; Brown, G.D.; Wong, S.Y. Expression of the beta-glucan receptor, Dectin-1, on murine leukocytes in situ correlates with its function in pathogen recognition and reveals potential roles in leukocyte interactions. J. Leukoc. Biol. 2004, 76, 86–94. [Google Scholar] [CrossRef] [PubMed]
- Romani, N.; Clausen, B.E.; Stoitzner, P. Langerhans cells and more: Langerin-expressing dendritic cell subsets in the skin. Immunol. Rev. 2010, 234, 120–141. [Google Scholar] [CrossRef] [PubMed]
- Valladeau, J.; Ravel, O.; Dezutter-Dambuyant, C.; Moore, K.; Kleijmeer, M.; Liu, Y.; Duvert-Frances, V.; Vincent, C.; Schmitt, D.; Davoust, J.; et al. Langerin, a novel C-type lectin specific to langerhans cells, is an endocytic receptor that induces the formation of birbeck granules. Immunity 2000, 12, 71–81. [Google Scholar] [CrossRef]
- Valladeau, J.; Duvert-Frances, V.; Pin, J.J.; Kleijmeer, M.J.; Ait-Yahia, S.; Ravel, O.; Vincent, C.; Vega, F., Jr.; Helms, A.; Gorman, D.; et al. Immature human dendritic cells express asialoglycoprotein receptor isoforms for efficient receptor-mediated endocytosis. J. Immunol. 2001, 167, 5767–5774. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Griffiths, M.N.; Burton, D.R.; Ghazal, P. Rapid antibody responses by low-dose, single-step, dendritic cell-targeted immunization. Proc. Nat. Acad. Sci. USA 2000, 97, 847–852. [Google Scholar] [CrossRef] [PubMed]
- Krug, A.; French, A.R.; Barchet, W.; Fischer, J.A.; Dzionek, A.; Pingel, J.T.; Orihuela, M.M.; Akira, S.; Yokoyama, W.M.; Colonna, M. TLR9-dependent recognition of MCMV by IPC and DC generates coordinated cytokine responses that activate antiviral NK cell function. Immunity 2004, 21, 107–119. [Google Scholar] [CrossRef] [PubMed]
- Blasius, A.L.; Giurisato, E.; Cella, M.; Schreiber, R.D.; Shaw, A.S.; Colonna, M. Bone marrow stromal cell antigen 2 is a specific marker of type I IFN-producing cells in the naive mouse, but a promiscuous cell surface antigen following IFN stimulation. J. Immunol. 2006, 177, 3260–3265. [Google Scholar] [CrossRef] [PubMed]
- Pugholm, L.H.; Petersen, L.R.; Sondergaard, E.K.; Varming, K.; Agger, R. Enhanced humoral responses induced by targeting of antigen to murine dendritic cells. Scand. J. Immunol. 2015, 82, 515–522. [Google Scholar] [CrossRef] [PubMed]
- Maisonnasse, P.; Bouguyon, E.; Piton, G.; Ezquerra, A.; Urien, C.; Deloizy, C.; Bourge, M.; Leplat, J.J.; Simon, G.; Chevalier, C.; et al. The respiratory DC/macrophage network at steady-state and upon influenza infection in the swine biomedical model. Mucosal Immunol. 2015. [Google Scholar] [CrossRef] [PubMed]
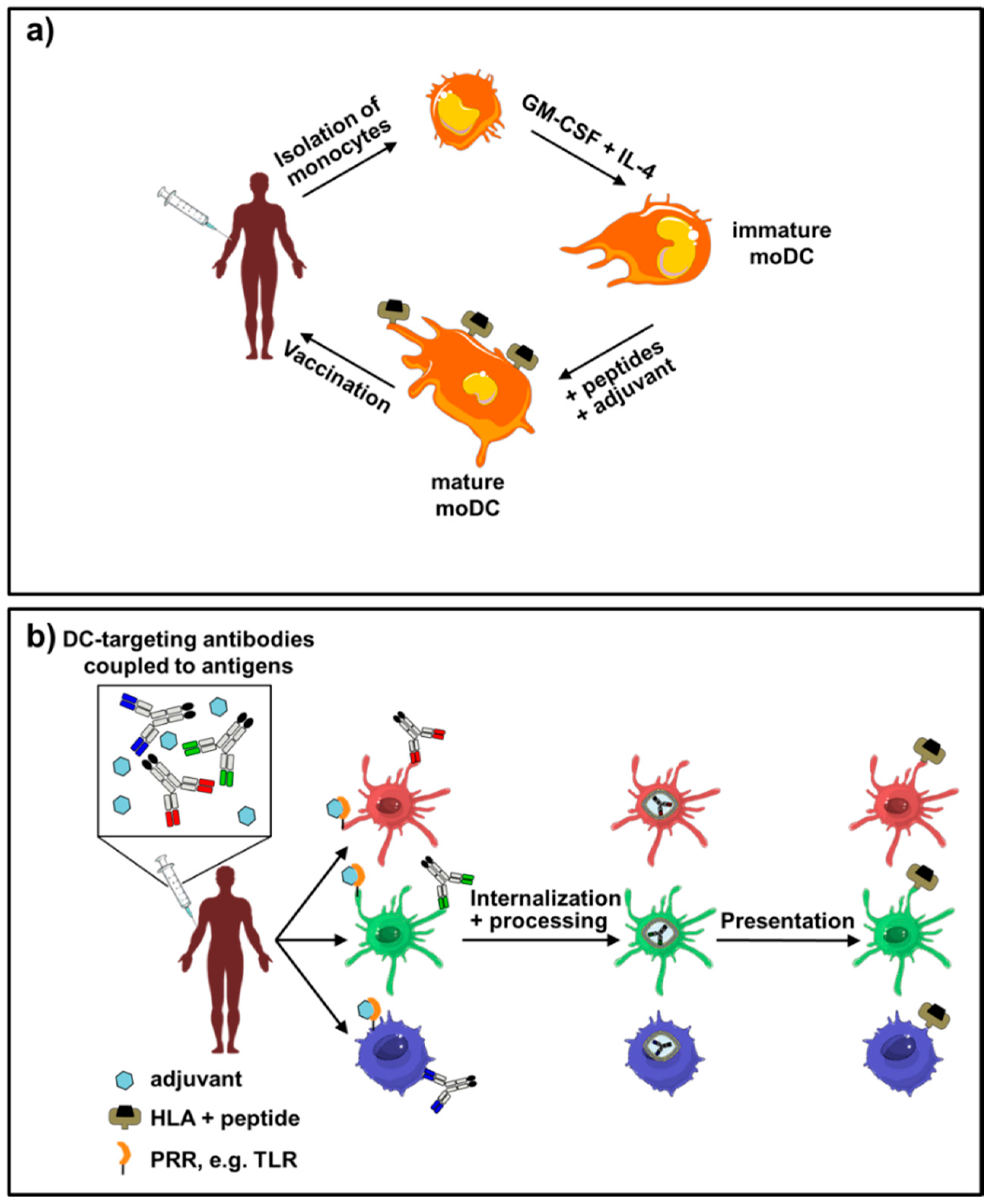

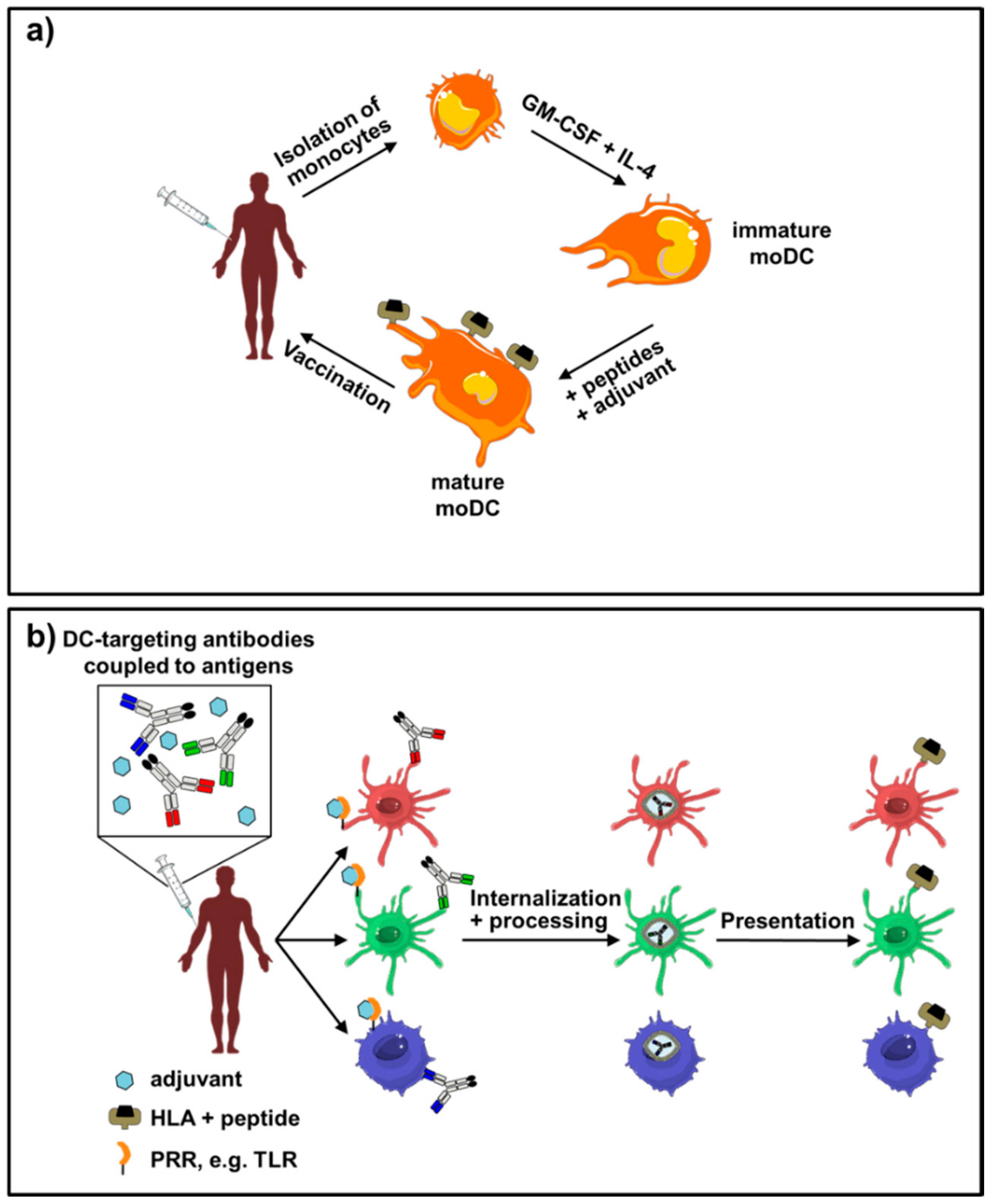{kind=link}
| Targeted Population | Antibody Type/Coupling | Used Antigen | In Vivo Model | Outcome | Ref. | |
|---|---|---|---|---|---|---|
| DEC205 | Murine CD8+ DEC205+ DCs | Fusion-protein, no FcR-binding | ovalbumin | Transfer of OT-I and OT-II transgenic T cells | Strong cross-presentation by CD8+ DCs targeted with a DEC205 antibody due to the expression of MHC-I machinery | [3] |
| Transfer of DO11.10 transgenic T cells | TGFβ-dependent induction of FoxP3+ Tregs | [7] | ||||
| Immunization of naïve C57Bl/6 mice | Induction of strong CD8+ and weaker CD4+ T cell responses | [178] | ||||
| Immunization of naïve C57Bl/6 mice | Ex vivo induced OT-I and OT-II T cell proliferation | [114] | ||||
| Transfer of OT-II transgenic T cells into BDCA-2 transgenic C57Bl/6 mice | CD4+ T cell proliferation, differentiation, and humoral responses | [137] | ||||
| Ova-expressing B16F10 melanoma cells, protective and therapeutic model | Induction of therapeutic and protective anti-tumor immune responses, ova-specific CD4+ and CD8+ T cell responses in naive C57Bl/6 mice, strong ova-specific mixed IgG1/IgG2a antibody response | [179] | ||||
| ovalbumin/ova-NP, circumsporozoite protein (CSP) | Immunization of naïve C57Bl/6, B10.BR, and BALB/c mice; transfer of CSP transgenic T cells | Induction of T helper cell responses, induction of high titers of hapten-specific IgG, stronger antibody response in comparison to immunization with irradiated sporozoites | [180] | |||
| ovalbumin/α-GalCer | Ova-expressing B16F10 and EG7, transfer of transgenic OT-I cells | Delivery of ova and α-GalCer to CD8+ DCs induces iNKT cell, CD8+ T cell, and protective and therapeutic anti-tumor responses | [181] | |||
| LcrV | Lethal aerosol challenge with Y. pestis | Induction of LcrV-specific antibody response, poorer survival in comparison to targeting with DCIR2 | [182] | |||
| NS-1 (Dengue virus) | Lethal intracranial challenge with DENV2 NGC | Improved survival by targeting NS-1 via DEC205 to CD8+ DCs, stronger TH1 response and IFNγ+ T cells | [171] | |||
| HIV gag p24 and p41 | Intranasal challenge with vaccinia-gag | Strong and broad T cell and antibody response to HIV gag, reduced severity of vaccinia-gag infection | [183] | |||
| HIV gag p24 | Immunization of naïve C57Bl/6 mice | Induction of IFNγ+ CD4+ and CD8+ T cell responses | [184] | |||
| Immunization of naïve mice in combination with several adjuvants | Poly(I:C) strongest adjuvant, signaling via IFNAR necessary for activation of DCs and induction of CD8+ T cell response | [185] | ||||
| Immunization of naïve C57Bl/6 mice | Long-term HIV-specific immunity within the gastrointestinal tract | [186] | ||||
| human cartilage proteoglycan (PG) | PG-induced arthritis with transfer of transgenic PG-specific T cells | Targeting PG to DCs induced reduced arthritis score, lower titer of PG-specific IgG1 and IgG2a, and lower proliferation of CD4+ T cells | [187] | |||
| stress-inducible 1 protein of L. major (LmSTI1a) | Intradermal or subcutaneous challenge with L. major | Induction of IFNγ+ CD4+ T cell responses, improved survival after challenge with L. major in BALB/c mice | [188] | |||
| Her2/neu | Protective breast cancer model, injection of NT2.5 tumor cells into FVB/N mice | Induction of IFNγ+ CD4+ and CD8+ T cell responses, protection in a breast cancer tumor model | [189] | |||
| Trypanosoma cruzi amastigote surface protein 2 (ASP-2) | Immunization of naïve C57Bl/6 mice | Induction of IFNγ+ CD4+ T cells, identification of an immunogenic epitope of ASP-2 | [190] | |||
| MimA2 | Transfer of diabetogenic transgenic AI4 T cells in NOD mice | Induction of tolerance to MimA2 due to deletion of the transferred T cells | [191] | |||
| BDC2.5 mimitope peptide 1040-63/pro-Insulin | Transfer of transgenic BDC2.5 T cells in NOD mice | Induction of Treg cells by delivery of BDC2.5 mimitope via DEC205, delayed onset of diabetes by delivery of pro-insulin to DCs via DEC205 | [192] | |||
| LACK (L. major) | Transfer of LACK-specific transgenic T cells | Targeting DEC205 induces IFNγ+ TH1 cells independent of IL-12, but dependent on CD70 | [160] | |||
| survivin | Immunization of naïve mice, depletion of Tregs | Mainly specific T cells against human survivin; depletion of Tregs enhances T cell response against survivin | [193] | |||
| IGRP | Immunization of naïve NOD mice | Reduced type-I diabetes | [194] | |||
| PLP(139-151) | Immunization of SJL/J mice after transfer of 5B6 transgenic T cells | Reduced experimental autoimmune encephalomyelitis (EAE) | [195] | |||
| HEL | Transfer of 3A9 transgenic T cells | T cell tolerance in response to antibody alone, memory response after co-injection of αCD40 | [8] | |||
| Fusion protein, FcR-binding? | ovalbumin | Transfer of OT-I and OT-II transgenic T cells | Even mature DCs take up antigens via targeting antibodies; induction of OT-I and OT-II T cell activation and proliferation | [196] | ||
| Fusion-protein, single chain Fv | myelin oligodendrocyte glycoprotein (MOG) | MOG-induced experimental allergic encephalomyelitis (EAE) | Induction of protective and therapeutic responses against EAE by targeting MOG to CD8+ DCs | [197] | ||
| Fusion-protein, single chain Fv | hNC16A collagen domain | Immunization of naïve C57Bl/6 mice | Reduced graft rejection | [198] | ||
| Single chain Fv | ova/Plasma membrane vesicles of B16F10-ova cells | Vaccination with liposomes or plasma membrane vesicles; i.v. injection of B16F10-ova cells (Lung metastasis model) | Antigen can be delivered in liposomes or plasma membrane vesicles via coupled scFv against DEC205 to DCs; reduction of tumor growth due to tumor-specific T cells | [199] | ||
| Murine DEC205+ LN DCs, CD8+ DEC205+ spleen DCs | Chemically coupled, FcR-binding? | ovalbumin | Transfer of OT-I and OT-II transgenic T cells; challenge with MO4-ova and vaccinia-ova | Induction of IFNγ+ memory T cell responses; vaccination induces protection against MO4-ova and vaccinia-ova; in vivo targeting protects in contrast to ex vivo loaded DCs | [157] | |
| Murine CD11c+ LN DCs | Chemically coupled, FcR-binding? | ovalbumin | Transfer of OT-I transgenic T cells | T cell tolerance in response to antibody alone, memory response after co-injection of αCD40 | [200] | |
| Murine CD11c+ DCs | Chemically coupled, FcR-binding? | ovalbumin/TNCB | Delayed-type hypersensitivity and contact hypersensitivity model | Induction of Tregs, tolerance in DTH and CHS models | [165] | |
| Murine dermal and LN DCs | Fusion protein, no FcR-binding | ovalbumin | Transfer of target cells in vaccinated mice | αDEC205 targets LCs and dermal DCs; induces cytotoxic CD8+ T cell responses independent of Langerin+ DCs (Langerin-DTR mice) | [201] | |
| Murine LCs and dermal DCs | Fusion protein, no FcR-binding | ovalbumin | Naïve C57Bl/6 mice | LCs are targeted by both Langerin and DEC205 antibodies; only DEC205 induces CD4+ and CD8+ T cell responses | [202] | |
| DEC205+ DCs | Fusion protein, no FcR-binding | HIV gag p24 | Immunization of non-human primates | Induction of broad CD4+ and CD8+ T cell responses against p24 after targeting of DEC205 in combination with poly ICLC | [203] | |
| Human DCs | Fusion protein, monoclonal humanized antibody | NY-ESO-1 | Phase I trial with NY-ESO-1 positive patients | 13/45 patients with stabilized disease, 2/45 with tumor regression; no dose-limiting or grade 3 toxicities reported | [85] | |
| Human moDCs, DEC205+ cells in humanized mice | Fusion protein, FcR-binding | EBNA-1 | Vaccination of humanized NOG mice | Activation of EBNA-1 specific autologous T cells; protection against EBV-infected B cells; induction of EBNA-1-specific T cells in humanized mice | [204] | |
| Human moDCs | Fusion protein, FcR-binding | HIV gag p24 | - | Stronger IFNγ+ CD8+ T cell responses by targeting with DEC205 in comparison to DC-SIGN or MMR | [205] | |
| NY-ESO-1 | - | Induction of CD4+ and CD8+ T cell responses in PBMCs of NY-ESO-1 seropositive breast cancer patients | [206] | |||
| Fusion protein, single chain Fv | MAGE-A3 epitope | - | Induction of proliferation of TCR-transfected CD4+ T cells | [207] | ||
| DCIR2 | Murine CD8− DCIR2+ DCs | Fusion-protein, no FcR-binding | ovalbumin | Transfer of OT-I and OT-II transgenic T cells | Mainly CD4+ T cell response, stronger expression of MHC-II machinery in CD8− DCs | [3] |
| Ova-expressing B16F10 melanoma cells, protective and therapeutic model | Induction of therapeutic and protective anti-tumor immune responses, ova-specific CD4+ and CD8+ T cell responses in naive C57Bl/6 mice, strong ova-specific mixed IgG1/IgG2a antibody response | [179] | ||||
| Immunization of naïve C57Bl/6 mice | Ex vivo induced OT-I and OT-II T cell proliferation | [114] | ||||
| Immunization of naïve C57Bl/6 mice | Induction of mainly CD4+ and weaker CD8+ T cell responses | [178] | ||||
| Murine dermal and LN DCs | Fusion protein, no FcR-binding | LcrV | Lethal aerosol challenge with Y. pestis | Induction of LcrV-specific antibody response, improved survival after targeting of DCIR2 | [182] | |
| Murine LCs and dermal DCs | Fusion protein, no FcR-binding | LACK (L. major) | Transfer of LACK-specific transgenic T cells | Targeting DCIR2 induces IFNγ+ TH1 cells dependent on IL-12, but independent of CD70 | [160] | |
| DEC205+ DCs | Chemically coupled, Fab-fragment | ovalbumin | Immunization of naïve C57Bl/6 and BALB/c mice | Induction of antibody responses | [208] | |
| CD34+-derived LCs, epidermal LCs, CD11c+ blood DCs, blood pDCs | Fusion protein, no FcR-binding | FluMP/MART-1/HIV gag p24 | - | Induction of IFNγ+ CD8+ T cell responses | [174] | |
| Human blood pDCs | Chemically coupled, FcR-binding? | KLH | - | Proliferation of T cells (not further defined) | [143] | |
| Clec9a | Murine CD8+ DCs | Fusion protein, no FcR-binding | HIV gag p24 | Immunization of naïve C57Bl/6 mice | Induction of IFNγ+ CD4+ and CD8+ T cell responses | [184] |
| Chemically coupled, FcR-binding? | SIINFEKL/epitopes of gp100, TRP-1, TRP-2 | Challenge with B16-ova cells | Protective and therapeutic responses in B16-ova melanoma model | [209] | ||
| ova-peptide (323-339) | Transfer of OT-II cells in C57Bl/6 mice | Differential polarization of naive CD4+ T cells dependent on the adjuvant | [210] | |||
| ovalbumin | Immunization of naïve mice | Induction of antibody response without adjuvant and independent of MyD88-signaling; induction of OT-I and OT-II transgenic T cell proliferation | [211] | |||
| Murine CD8+ DCs | Fusion protein, single chain Fv | ovalbumin | Transfer of OT-I and OT-II cells | Comparable OT-I T cell activation between DEC205 and Clec9a, superior CD4+ T cell responses after targeting of Clec9a | [212] | |
| Human blood BDCA-3+ DCs | Biotin-labeled KLH, gp110-filled nanoparticles, FcR-binding? | KLH/gp100 | - | Induction of KLH+ CD4+ T cell responses, cross-presentation to CD8+ T cells | [213] | |
| Clec12a | Murine CD8+ DCs and pDCs | Chemically coupled, FcR-binding? | ovalbumin | Transfer of OT-I and OT-II cells in C57Bl/6 and CD11c-DTR mice | Induction of OT-I and OT-II responses by CD11c+ cells | [214] |
| Dectin-1 | Murine DN DCs | Chemically coupled, FcR-binding? | ovalbumin | Immunization of naïve C57Bl/6 mice, transfer of OT-I and OT-II transgenic T cells | Stronger CD4+ T cell response and weaker CD8+ T cell response in comparison to DEC205-targeting; induction of strong antibody response, especially after i.v. injection | [215] |
| Human IFNα moDCs | Fusion protein, FcR-binding? | hemagglutinin | - | Re-stimulation of memory TH17 cells via antigen-targeting to Dectin-1 | [216] | |
| Human IL-4 or IFNγ DCs | Fusion to hIgG4 | Flu M1, MART-1 (26–35) | - | Activation of moDCs by Dectin-1 antibody, expansion of Flu M1 spec & MART-1 CD8+ T cells, differentiation of naïve CD8+ T cells into Flu M1-specific | [217] | |
| Langerin | Murine CD8+ DEC205+ DCs | Fusion-protein, no FcR-binding | HIV gag p24 | Immunization of naïve C57Bl/6 mice | Induction of IFNγ+ CD4+ and CD8+ T cell responses | [184] |
| Murine CD11c+ Langerin+ DCs | Fusion-protein, no FcR-binding | ovalbumin | Immunization of naïve C57Bl/6 mice | Induction of CD4+ and CD8+ T cell responses | [178] | |
| Murine LCs and dermal DCs | Fusion-protein, no FcR-binding | ovalbumin | B16-ova model | Targeting of LCs with Langerin and Imiquimod led to cross-tolerance and impaired secondary memory response using DEC205 as targeting antibody | [218] | |
| Naïve C57Bl/6 mice | LCs are targeted by both Langerin and DEC205 antibodies; only DEC205 induces CD4+ and CD8+ T cell responses | [202] | ||||
| Murine LCs and dermal DCs | Fusion-protein, no FcR-binding | MOGp | Naïve C57Bl/6 mice, partly transfer of MOG-specific T cells | Langerin+ cells can induce tolerance in vivo | [172] | |
| DC-SIGN | Human moDCs | Chemically coupled, FcR-binding? | KLH | - | Proliferation of T cells | [219] |
| Murine CD11c+ cells expressing hDC-SIGN | Chemically coupled, FcR-binding? | ovalbumin | Infection of humanized mice with Listeria monocytogenes | Vaccination with DC-SIGN-ova protects humanized mice from infection with Listeria monocytogenes | [220] | |
| Transfer of OT-I T cells into hDC-SIGN expressing C57Bl/6 mice | Prolonged antigen residence in early endosomes, delayed lysosomal degradation, and cross-presentation | [221] | ||||
| DC-SIGN+ APCs | - | - | Injection of cynomolgus macaque | APCs in LNs of macaques were targeted | [222] | |
| Human DC-SIGN+ cells | Chemically coupled (KLH), FcR-binding?; fused scFv (tetanus toxoid peptides) | KLH/tetanus toxoid peptides | Immunization of humanized mice | Induction of cell proliferation after targeting of KLH to DC-SIGN+ cells; protection after transfer of Raji-cells | [223] | |
| MR | Human moDCs | Fusion-protein, FcR-binding? | chorionic gonadotropin β | - | Induction of autologous T cell responses against CGβ; cytotoxic T cells show lysis against CGβ+ tumor cell lines (HLA-partially matched) | [224] |
| pmel17 | - | Induction of autologous T cell responses against pmel17; cytotoxic T cells show lysis against pmel17+ melanoma cell lines (HLA-partially matched) | [225] | |||
| chorionic gonadotropin β | - | TLR ligands boosted cytotoxic T cell response induced by antibody targeting | [226] | |||
| NY-ESO-1 | - | Antigen targeting to MR induces activation of CD4+ and CD8+ T cells | [206] | |||
| CD11c | Murine CD11c+ cells | Chemically coupled, Fab-fragment | ovalbumin | Immunization of naïve mice; transferof OT-I and OT-II transgenic T cells; EL4 as target cells | CD11c superior in the generation of CD8+ and CD4+ T cells; targeting leads to in vivo lysis of EL4 cells; induces endogenous ova-specific T cells | [227] |
| Immunization of naïve C57Bl/6 and BALB/c mice | Induction of antibody responses | [208] | ||||
| Fusion protein, single chain Fv | Her2/neu | Challenge with D2F2/E2 breast cancer cells; spontaneous breast cancer model | Induces protective and therapeutic immune responses against Her2/neu expressing tumor cells; delays tumor growth and onset in a spontaneous breast cancer model | [228] | ||
| MHC-II | MHC-II+ APCs | Chemically coupled, Fab-fragment | ovalbumin | Immunization of naïve C57Bl/6 and BALB/c mice | Induction of antibody responses | [208] |
| MHC-II+ APCs | Chemically coupled, Fab-fragment | ovalbumin | Immunization of naïve C57Bl/6 and BALB/c mice | Induction of antibody responses | [208] | |
| Chemically coupled, superantigen M1 | ovalbumin | In vivo killing assays; challenge with B16F10-ova melanoma cells | Cross-presentation by all splenic DC subsets; induces T cell responses with in vivo killing activity; therapeutic response in B16F10-ova melanoma model | [229] | ||
| LOX-1 | Not further defined APCs in vivo | Chemically coupled, FcR-binding? | ovalbumin | Challenge of C57Bl/6 mice with E.G7 tumor cells expressing ova | Targeting ova to LOX-1 induces protective immune response against E.G7 cells | [230] |
| Macaque blood CD11c+ and CD14+ cells | Fusion protein, FcR-binding? | hemagglutinin | Immunization of rhesus macaques; challenge of macaques with Influenza | Induction of HA-specific antibodies in macaques, higher antibody titer in comparison of Dectin-1 targeting | [231] | |
| Human IFNα moDCs | Fusion protein, FcR-binding? | hemagglutinin | - | LOX-1 targeting induces activation of TH1 cells | [232] | |
| DC-ASGPR | Human IFNα moDCs | Fusion protein, FcR-binding? | hemagglutinin | - | DC-ASGPR targeting induces the secretion of IL-10 by DCs and the polarization/re-stimulation of suppressive IL-10+ T cells | [232] |
| CD36 | Murine CD8+ DCs | Fusion protein, scFV | ovalbumin | Challenge of C57Bl/6 mice with E.G7 tumor cells expressing ova | Targeting ova to CD36 induces protective immune response against EG7 tumor cells expressing ova, memory OT-I T cell response without adjuvant | [233] |
| Siglec-H | Murine plasmacytoid DCs | Fusion protein, no FcR-binding | ovalbumin, pHEL, pMOG | EAE | Less severe EAE after targeting of MOG to Siglec-H on pDCs, less T cell polarization, lower antibody titers even after injection of an adjuvant | [141] |
| ovalbumin | Transfer of OT-II transgenic T cells into BDCA-2 transgenic C57Bl/6 mice | CD4+ T cell proliferation, differentiation, and humoral responses | [137] | |||
| Murine pDCs | Chemically coupled, FcR-binding? | ovalbumin | Immunization of naïve mice, boost with Vaccinia virus expressing ova | Targeting ova via Siglec-H to pDCs induces ova-specific CD8+ T cells only when CpG is co-injected | [234] | |
| BST-2 | Murine pDCs | Fusion protein, no FcR-binding | ovalbumin, pHEL | Vaccinia virus expressing ova, B16F10-ova melanoma cells | Targeting ova to pDCs via BST-2 protects against VV-infection and B16F10-ova cells; induces activation of OT-I and OT-II cells as well as antibody titer | [142] |
| Chemically coupled, F(ab)2-fragment | ovalbumin | Transfer of OT-I and OT-II cells | Targeting of ova to pDCs induces OT-II cell proliferation in lymph nodes but not in spleen | [235] | ||
| BDCA-2 | Murine pDCs | Fusion protein, no FcR-binding | ovalbumin | Transfer of OT-II transgenic T cells into BDCA-2 transgenic C57Bl/6 mice | CD4+ T cell proliferation, differentiation, and humoral responses | [137] |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons by Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lehmann, C.H.K.; Heger, L.; Heidkamp, G.F.; Baranska, A.; Lühr, J.J.; Hoffmann, A.; Dudziak, D. Direct Delivery of Antigens to Dendritic Cells via Antibodies Specific for Endocytic Receptors as a Promising Strategy for Future Therapies. Vaccines 2016, 4, 8. https://doi.org/10.3390/vaccines4020008
Lehmann CHK, Heger L, Heidkamp GF, Baranska A, Lühr JJ, Hoffmann A, Dudziak D. Direct Delivery of Antigens to Dendritic Cells via Antibodies Specific for Endocytic Receptors as a Promising Strategy for Future Therapies. Vaccines. 2016; 4(2):8. https://doi.org/10.3390/vaccines4020008
Chicago/Turabian StyleLehmann, Christian H. K., Lukas Heger, Gordon F. Heidkamp, Anna Baranska, Jennifer J. Lühr, Alana Hoffmann, and Diana Dudziak. 2016. "Direct Delivery of Antigens to Dendritic Cells via Antibodies Specific for Endocytic Receptors as a Promising Strategy for Future Therapies" Vaccines 4, no. 2: 8. https://doi.org/10.3390/vaccines4020008
APA StyleLehmann, C. H. K., Heger, L., Heidkamp, G. F., Baranska, A., Lühr, J. J., Hoffmann, A., & Dudziak, D. (2016). Direct Delivery of Antigens to Dendritic Cells via Antibodies Specific for Endocytic Receptors as a Promising Strategy for Future Therapies. Vaccines, 4(2), 8. https://doi.org/10.3390/vaccines4020008