DNA Virus Vectors for Vaccine Production in Plants: Spotlight on Geminiviruses

Abstract
:
1. Introduction
2. Concerning Geminiviruses
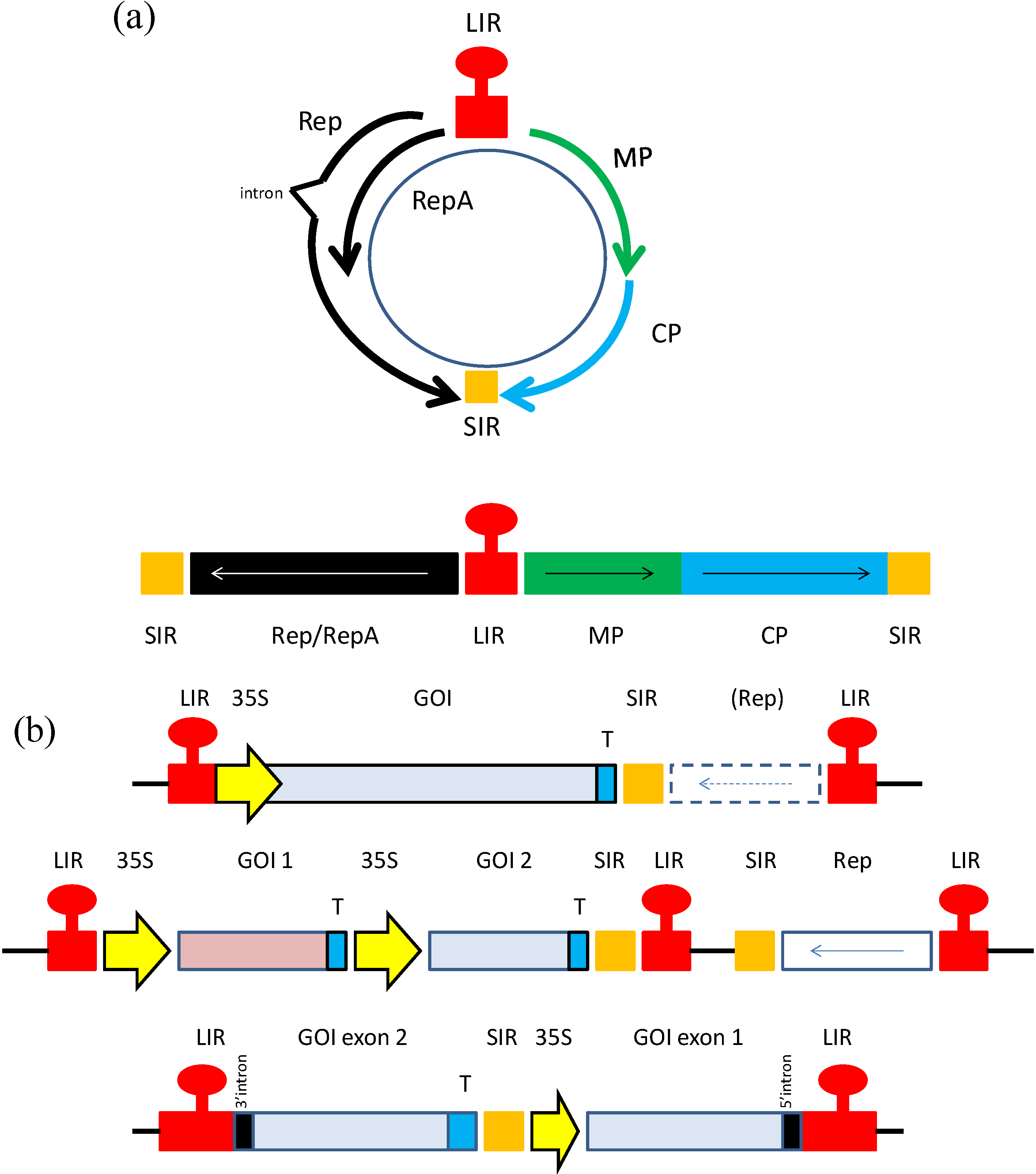
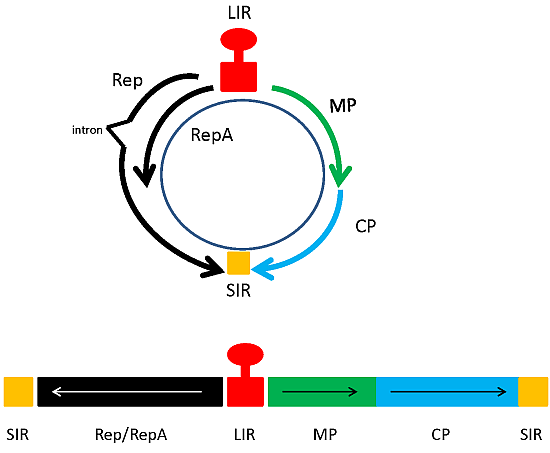
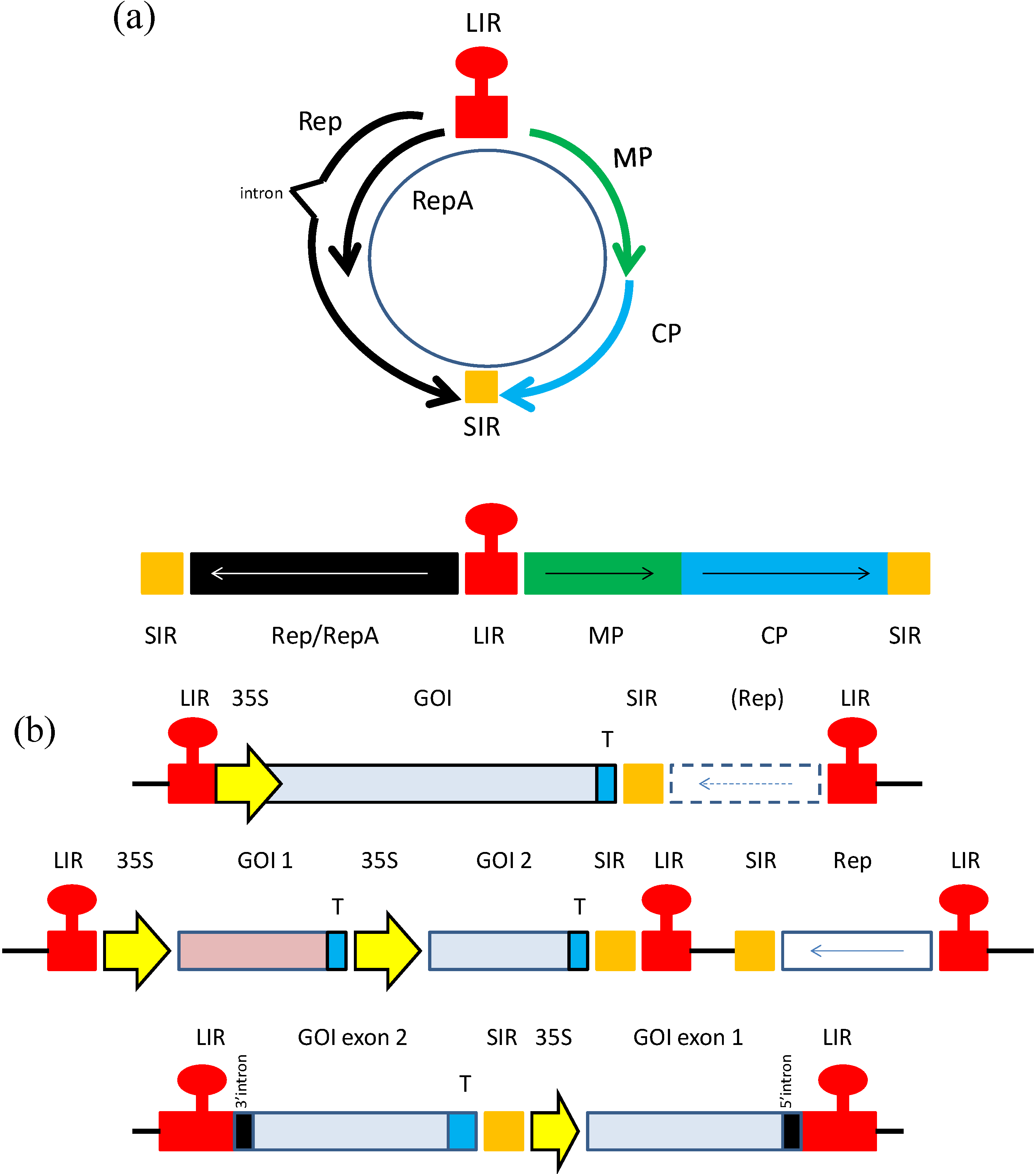
3. Geminiviruses as Expression Vectors
3.1. Early Expression Vector Studies

{kind=link}
{kind=link}
| Geminivirus | Function of Vector | Reference |
|---|---|---|
| Wheat Dwarf Virus Expression Vector [23] Maize streak virus (MSV) | Expression vector | [26,27] |
| Bean yellow dwarf virus (BeYDV) | vaccines, gene silencing, DNA repair | [35,36,37] |
| Mild stain of BeYDV | Vaccine production | [38,39] |
| Beet curly top virus (BCTV) | Vaccine production | [40,41] |
| Tobacco yellow dwarf virus (TYDV) | Vaccine/industrial protein production | [16,42] |
| 5Tobacco curly shoot virus (TbCSV) | Gene silencing | [43] |
| Cabbage leaf-curl virus (CaLCuV) | Gene silencing | [44] |
| African cassava mosaic virus (ACMV) | Gene silencing | [45] |
| Ageratum yellow vein virus (AYVV) | Expression Vector | [46] |
| Abutilon mosaic vírus (AbMV) | Gene silencing | [47] |
| Cotton leaf curl Multan betasatellite (CLCuMB) | Gene silencing | [48] |
3.2. Next Generation Geminivirus Expression Vectors
3.2.1. BeYDV Vaccine Vectors
| Therapeutic Protein | Vector Used | Host Plant | Expression Level | Immunogenicity Tested | Reference |
|---|---|---|---|---|---|
| SEB | BeYDV | N. bethamiana | n/a * | yes | [31] |
| Norwalk Virus VLPs | BeYDV | Tobacco, lettuce | 0.34 mg/g LFW ** | yes | [36] |
| HBVcAg | BeYDV | N. benthamiana | 0.8 mg/g LFW | no | [34] |
| WNV E protein Mab | BeYDV | Tobacco, lettuce | 0.23–0.27 mg/g LFW | yes | [35] |
| Ebola Virus GP1Mab | BeYDV | Tobacco, lettuce | 0.23–0.27 g/g LFW | yes | [35] |
| HPV-1 L1 protein | BeYDV, mild strain | N. benthamiana | n/a | no | [38] |
| HIV-1 type C p24 | BeYDV, mild strain | N. benthamiana | n/a | no | [38] |
| HAV VP1 | BCTV | N. benthamiana | n/a | no | [41] |
| vitronectin | TYDV | N. benthamiana | 2.3% TSP *** | n/a | [42] |
3.2.2. BCTV Vaccine Vector
3.2.3. TYDV Vaccine Vector
4. Other Geminivirus Vectors and Their Uses
5. Conclusions
Acknowledgments
Conflicts of Interest
References
- Hamorsky, K.T.; Kouokam, J.C.; Bennett, L.J.; Baldauf, K.J.; Kajiura, H.; Fujiyama, K.; Matoba, N. Rapid and scalable plant-based production of a cholera toxin B subunit variant to aid in mass vaccination against cholera outbreaks. PLoS Negl. Trop. Dis. 2013, 7, e2046. [Google Scholar] [CrossRef]
- McCormick, A.A.; Reddy, S.; Reinl, S.J.; Cameron, T.I.; Czerwinkski, D.K.; Vojdani, F.; Hanley, K.M.; Garger, S.J.; White, E.L.; Novak, J.; et al. Plant-produced idiotype vaccines for the treatment of non-Hodgkin’s lymphoma: Safety and immunogenicity in a phase I clinical study. Proc. Natl. Acad. Sci. USA 2008, 105, 10131–10136. [Google Scholar] [CrossRef]
- Platis, D.; Drossard, J.; Fischer, R.; Ma, J.K.; Labrou, N.E. New downstream processing strategy for the purification of monoclonal antibodies from transgenic tobacco plants. J. Chromatogr. A 2008, 1211, 80–89. [Google Scholar] [CrossRef]
- Leuzinger, K.; Dent, M.; Hurtado, J.; Stahnke, J.; Lai, H.; Zhou, X.; Chen, Q. Efficient agroinfiltration of plants for high-level transient expression of recombinant proteins. J. Vis. Exp. 2013, 23. [Google Scholar] [CrossRef]
- Bisaro, D.M. Geminivirus DNA replication. In DNA Replication in Eukaryofic Cells; Cold Spring Harbor Laboratory Press: Cold Spring Harbor, NY, USA, 1996. [Google Scholar]
- Lazarowitz, S.G. Geminiviruses: Genome structure and gene function. Crit. Rev. Plant Sci. 1992, 1, 327–349. [Google Scholar] [CrossRef]
- Faurez, F.; Dory, D.; Grasland, B.; Jestin, A. Replication of porcine circoviruses. Virol. J. 2009, 6. [Google Scholar] [CrossRef]
- Delwart, E.; Li, L. Rapidly expanding genetic diversity and host range of the Circoviridae viral family and other Rep encoding small circular ssDNA genomes. Virus Res. 2012, 164, 114–121. [Google Scholar] [CrossRef]
- Krupovic, M.; Ravantti, J.J.; Bamford, D.H. Geminiviruses: A tale of a plasmid becoming a virus. BMC Evol. Biol. 2009, 9. [Google Scholar] [CrossRef]
- Ilyina, T.V.; Koonin, E.V. Conserved sequence motifs in the initiator proteins for rolling circle DNA replication encoded by diverse replicons from eubacteria, eucaryotes and archaebacteria. Nucl. Acids Res. 1992, 20, 3279–3285. [Google Scholar] [CrossRef]
- Saccardo, F.; Cettul, E.; Palmano, S.; Noris, E.; Firrao, G. On the alleged origin of geminiviruses from extrachromosomal DNAs of phytoplasmas. BMC Evol Biol. 2011, 11. [Google Scholar] [CrossRef]
- Katzourakis, A.; Gifford, R.J. Endogenous viral elements in animal genomes. PLoS Genet. 2010, 6, e1001191. [Google Scholar] [CrossRef]
- Belyi, V.A.; Levine, A.J.; Skalka, A.M. Sequences from ancestral single-stranded DNA viruses invertebrate genomes: The parvoviridae and circoviridae are more than 40 to 50 million years old. J. Virol. 2010, 84, 12458–12462. [Google Scholar] [CrossRef]
- Liu, H.; Fu, Y.; Li, B.; Yu, X.; Xie, J.; Cheng, J.; Ghabrial, S.A.; Li, G.; Yi, X.; Jiang, D. Widespread horizontal gene transfer from circular single-stranded DNA viruses to eukaryotic genomes. BMC Evol. Biol. 2011, 11. [Google Scholar] [CrossRef]
- Chen, Q.; He, J.; Phoolcharoen, W.; Mason, H.S. Geminiviral vectors based on bean yellow dwarf virus for production of vaccine antigens and monoclonal antibodies in plants. Hum. Vaccin. 2011, 7, 331–338. [Google Scholar] [CrossRef]
- Dugdale, B.; Mortimer, C.L.; Kato, M.; James, T.A.; Harding, R.M.; Dale, J.L. In plant activation: An inducible, hyperexpression platform for recombinant protein production in plants. Plant Cell 2013, 25, 2429–2443. [Google Scholar] [CrossRef]
- Hanley-Bowdoin, L.; Settlage, S.B.; Orozco, B.M.; Nagar, S.; Robertson, D. Geminiviruses: Models for plant DNA replication, transcription, and cell cycle regulation. Crit. Rev. Plant Sci. 1999, 18, 71–106. [Google Scholar] [CrossRef]
- Jeske, H. Geminiviruses. Curr. Top. Microbiol. Immunol. 2009, 331, 185–226. [Google Scholar]
- Paprotka, T.; Deuschle, K.; Metzler, V.; Jeske, H. Conformation-selective methylation of geminivirus DNA. J. Virol. 2011, 85, 12001–12012. [Google Scholar] [CrossRef]
- Hanley-Bowdoin, L.; Bejarano, E.R.; Robertson, D.; Mansoor, S. Geminiviruses: Masters at redirecting and reprogramming plant processes. Nat. Rev. Microbiol. 2013, 11, 777–788. [Google Scholar] [CrossRef]
- Mikhail, M. Pooggin how can plant DNA viruses evade siRNA-directed DNA methylation and silencing? Int. J. Mol. Sci. 2013, 14, 15233–15259. [Google Scholar] [CrossRef]
- Raja, P.; Sanville, B.C.; Buchmann, R.C.; Bisaro, D.M. Viral genome methylation as an epigenetic defense against geminiviruses. J. Virol. 2008, 82, 8997–9007. [Google Scholar] [CrossRef]
- Trinks, D.; Rajeswaran, R.; Shivaprasad, P.V.; Akbergenov, R.; Oakeley, E.J.; Veluthambi, K.; Hohn, T.; Pooggin, M.M. Suppression of RNA silencing by a geminivirus nuclear protein, AC2, correlates with transactivation of host genes. J. Virol. 2005, 79, 2517–2527. [Google Scholar] [CrossRef]
- Yang, X.; Xie, Y.; Raja, P.; Li, S.; Wolf, J.N.; Shen, Q.; Bisaro, D.M.; Zhou, X. Suppression of methylation-mediated transcriptional gene silencing by βC1-SAHH protein interaction during geminivirus-betasatellite infection. PLoS Pathog. 2011, 7, e1002329. [Google Scholar] [CrossRef]
- Timmermans, M.C.; Das, O.P.; Messing, J. Trans replication and high copy numbers of wheat dwarf virus vectors in maize cells. Nucl. Acids Res. 1992, 20, 4047–4054. [Google Scholar] [CrossRef]
- Matzeit, V.; Schaefer, S.; Kammann, M.; Schalk, H.J.; Schell, J.; Gronenborn, B. Wheat dwarf virus vectors replicate and express foreign genes in cells of monocotyledonous plants. Plant Cell 1991, 3, 247–258. [Google Scholar] [CrossRef]
- Ugaki, M.; Ueda, T.; Timmermans, M.C.; Vieira, J.; Elliston, K.O.; Messing, J. Replication of a geminivirus derived shuttle vector in maize endosperm cells. Nucl. Acids Res. 1991, 19, 371–377. [Google Scholar]
- Davies, J.W.; Stanley, J. Geminivirus genes and vectors. Trends Genet. 1989, 5, 77–81. [Google Scholar] [CrossRef]
- Palmer, K.E.; Rybicki, E.P. Investigation of the potential of maize streak virus to act as an infectious gene vector in maize plants. Arch Virol. 2001, 146, 1089–1104. [Google Scholar] [CrossRef]
- Stanley, J. Geminiviruses: Plant viral vectors. Curr Opin Genet Dev. 1993, 3, 91–96. [Google Scholar] [CrossRef]
- Shen, W.H.; Hohn, B. Vectors based on maize streak virus can replicate to high copy numbers in maize plants. J. Gen Virol. 1995, 76, 965–969. [Google Scholar] [CrossRef]
- Hefferon, K.L.; Dugdale, B.G. Independent expression of Rep and RepA and their roles in regulating bean yellow dwarf virus replication. J. Gen. Virol. 2003, 84, 3465–3472. [Google Scholar] [CrossRef]
- Hefferon, K.L.; Fan, Y. Production of a vaccine protein in plants using a geminivirus expression system. Vaccine 2004, 23, 404–410. [Google Scholar] [CrossRef]
- Huang, Z.; Chen, Q.; Hjelm, B.; Arntzen, C.; Mason, H. A DNA replicon system for rapid high-level production of virus-like particles in plants. Biotechnol. Bioeng. 2009, 103, 706–714. [Google Scholar] [CrossRef]
- Phoolcharoen, W.; Bhoo, S.H.; Lai, H.; Ma, J.; Arntzen, C.J.; Chen, Q.; Mason, H.S. Expression of an immunogenic Ebola immune complex in Nicotiana benthamiana. Plant Biotechnol. J. 2011, 9, 807–816. [Google Scholar] [CrossRef]
- Lai, H.; He, J.; Engle, M.; Diamond, M.S.; Chen, Q. Robust production of virus-like particles and monoclonal antibodies with geminiviral replicon vectors in lettuce. Plant Biotechnol. J. 2012, 10, 95–104. [Google Scholar] [CrossRef]
- Huang, Z.; Phoolcharoen, W.; Lai, H.; Piensook, K.; Cardineau, G.; Zeitlin, L.; Whaley, K.J.; Arntzen, C.J.; Mason, H.S.; Chen, Q. High-level rapid production of full-size monoclonal antibodies in plants by a single-vector DNA replicon system. Biotechnol. Bioeng. 2010, 106, 9–17. [Google Scholar]
- Regnard, G.L.; Halley-Stott, R.P.; Tanzer, F.L.; Hitzeroth, I.I.; Rybicki, E.P. High level protein expression in plants through the use of a novel autonomously replicating geminivirus shuttle vector. Plant Biotechnol. J. 2010, 8, 38–46. [Google Scholar] [CrossRef]
- Rybicki, E.P.; Martin, D.P. Virus-derived ssDNA vectors for the expression of foreign proteins in plants. Curr. Top. Microbiol. Immunol. 2014, 375, 19–45. [Google Scholar]
- Kim, K.I.; Sunter, G.; Bisaro, D.M.; Chung, I.S. Improved expression of recombinant GFP using a replicating vector based on Beet curly top virus in leaf-disks and infiltrated Nicotiana benthamiana leaves. Plant Mol. Biol. 2007, 64, 103–112. [Google Scholar] [CrossRef]
- Chung, H.Y.; Lee, H.H.; Kim, K.I.; Chung, H.Y.; Hwang-Bo, J.; Park, J.H.; Sunter, G.; Kim, J.B.; Shon, D.H.; Kim, W.; et al. Expression of a recombinant chimeric protein of hepatitis A virus VP1-Fc using a replicating vector based on Beet curly top virus in tobacco leaves and its immunogenicity in mice. Plant Cell Rep. 2011, 30, 1513–1521. [Google Scholar] [CrossRef]
- Dugdale, B.; Mortimer, C.L.; Kato, M.; James, T.A.; Harding, R.M.; Dale, J.L. Design and construction of an in-plant activation cassette for transgene expression and recombinant protein production in plants. Nat. Protoc. 2014, 9, 1010–1027. [Google Scholar] [CrossRef]
- Tang, Y.; Wang, F.; Zhao, J.; Xie, K.; Hong, Y.; Liu, Y. Virus-based microRNA expression for gene functional analysis in plants. Plant Physiol. 2010, 153, 632–641. [Google Scholar] [CrossRef]
- Huang, C.; Xie, Y.; Zhou, X. Efficient virus-induced gene silencing in plants using a modified geminivirus DNA1 component. Plant Biotechnol. J. 2009, 7, 254–265. [Google Scholar] [CrossRef]
- Fofana, I.B.; Sangaré, A.; Collier, R.; Taylor, C.; Fauquet, C.M. A geminivirus-induced gene silencing system for gene function validation in cassava. Plant Mol. Biol. 2004, 56, 613–624. [Google Scholar] [CrossRef]
- Tamilselvi, D.; Anand, G.; Swarup, S. A geminivirus AYVV-derived shuttle vector for tobacco BY2 cells. Plant Cell Rep. 2004, 23, 81–90. [Google Scholar]
- Krenz, B.; Wege, C.; Jeske, H. Cell-free construction of disarmed Abutilon mosaic virus-based gene silencing vectors. J. Virol. Methods 2010, 169, 129–137. [Google Scholar] [CrossRef]
- Kharazmi, S.; Behjatnia, S.A.; Hamzehzarghani, H.; Niazi, A. Cotton leaf curl Multan betasatellite as a plant gene delivery vector trans-activated by taxonomically diverse geminiviruses. Arch. Virol. 2012, 157, 1269–1279. [Google Scholar] [CrossRef]
- Zhang, X.; Mason, H. Bean Yellow Dwarf Virus replicons for high-level transgene expression in transgenic plants and cell cultures. Biotechnol. Bioeng. 2006, 93, 271–279. [Google Scholar] [CrossRef]
- Carrillo-Tripp, J.; Shimada-Beltrán, H.; Rivera-Bustamante, R. Use of geminiviral vectors for functional genomics. Curr. Opin. Plant Biol. 2006, 9, 209–215. [Google Scholar] [CrossRef]
- Mach, J. Geminivirus vectors deliver reagents for plant genome engineering. Plant Cell 2014, 26, 2. [Google Scholar] [CrossRef]
- Seaberg, B.L.; Hsieh, Y.C.; Scholthof, K.B.; Scholthof, H.B. Host impact on the stability of a plant virus gene vector as measured by a new fluorescent local lesion passaging assay. J. Virol. Methods 2012, 179, 289–294. [Google Scholar]
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Hefferon, K.L. DNA Virus Vectors for Vaccine Production in Plants: Spotlight on Geminiviruses. Vaccines 2014, 2, 642-653. https://doi.org/10.3390/vaccines2030642
Hefferon KL. DNA Virus Vectors for Vaccine Production in Plants: Spotlight on Geminiviruses. Vaccines. 2014; 2(3):642-653. https://doi.org/10.3390/vaccines2030642
Chicago/Turabian StyleHefferon, Kathleen L. 2014. "DNA Virus Vectors for Vaccine Production in Plants: Spotlight on Geminiviruses" Vaccines 2, no. 3: 642-653. https://doi.org/10.3390/vaccines2030642
APA StyleHefferon, K. L. (2014). DNA Virus Vectors for Vaccine Production in Plants: Spotlight on Geminiviruses. Vaccines, 2(3), 642-653. https://doi.org/10.3390/vaccines2030642