Abstract
Background/Objectives: Lawsonia intracellularis is a bacterium that causes Proliferative Enteropathy, an enteric infection characterized mainly by diarrhea and growth retardation, leading to important economic losses. Acute and chronic infections are easily diagnosed, and their control by vaccination has been proven efficacious. However, subclinical infections, despite being very prevalent, often remain underdiagnosed and uncontrolled in practice. Scarce research is available on the control of subclinical infections by vaccination, and the benefit in these scenarios remains to be elucidated. Two field trials were carried out to (1) determine the association between the growth and fecal shedding of L. intracellularis in unvaccinated and intramuscularly vaccinated pigs in a farm with subclinical infection and (2) assess the impact of intradermal vaccination against L. intracellularis on clinical performance and carcass quality in a herd with subclinical infection. Methods: A pig herd with subclinical infection was selected. Pigs were vaccinated intramuscularly (study 1) or intradermally (study 2) at weaning. Fecal shedding, performance, clinical parameters, and carcass quality were investigated. Results: Growth was negatively associated with the fecal load of L. intracellularis in non-vaccinated pigs, whereas in vaccinated pigs, growth performance was not impacted by fecal load (study 1). Vaccinated pigs presented a significantly lower fecal load, lower prevalence of tail biting (31.7%) compared with controls (54.2%), less back fat, and a greater Lean Meat percentage (study 2). Conclusions: Vaccination against L. intracellularis in a herd with subclinical infection and low fecal bacterial shedding led to a reduction in fecal shedding, a lower prevalence of tail biting, and an improvement in carcass quality.
1. Introduction
Porcine Proliferative Enteropathy is an enteric disease caused by Lawsonia intracellularis, an intracellular Gram-negative bacterium [1]. This pathogen infects immature intestinal epithelial cells mainly from the ileum, leading to the adenomatous proliferation of an immature intestinal mucosa, which is commonly known as ileitis [2]. This thickening of the intestinal mucosa impairs nutrient absorption, followed by diarrhea and subsequent growth retardation [3].
The clinical presentation of the disease may vary from acute to chronic and subclinical forms. The acute form, known as Proliferative Hemorrhagic Enteropathy, is associated with hemorrhagic diarrhea and sudden death. The impact of the chronic form, or Porcine Intestinal Adenomatosis, originates from the impairment of nutrient absorption, resulting in energy leakage through diarrhea and leading to a loss of growth [2]. In subclinical infections, diarrhea is uncommonly seen; however, the gut mucosa is colonized by a moderate number of L. intracellularis bacteria, and intestinal lesions are already extensive [4,5,6], indicating that energy leakage through the loss of amino acids and proteins occurs [7]. Winkelman [8] defined subclinical ileitis as a confirmed laboratory diagnosis through the detection of the bacterium by PCR or antibodies, along with the presence of macro- and/or microscopic lesions, but without any ileitis-associated clinical signs or mortality. The absence of specific clinical signs and ileitis-associated mortality makes this form invisible to the eyes of the animal caretaker or the diagnostician, particularly when precise and reliable growth recordings are unavailable [9].
Numerous surveys have shown high herd prevalence in farms infected with this bacterium in Europe (70–100%) [6], the US (75–78%) [10], Brazil (73–98%) [11,12], and Asia (77–100%) [13,14,15,16,17]. Despite this high prevalence of L. intracellularis-positive farms worldwide, the number of farms diagnosed in practice with either acute or chronic infections remain limited, whereas a significant proportion of subclinically infected farms continues to be underdiagnosed [4,18,19]. In addition, the use of antimicrobials hinders the detection of these subclinical infections. Fortunately, several studies have given visibility to the economic relevance of L. intracellularis subclinical infections and their negative impact on growth [19,20,21]. Interestingly, all these studies have quantified the fecal bacterial load of infected pigs by using a real-time polymerase chain reaction (qPCR) [22,23] and then have demonstrated a positive association between this fecal load and both microscopic lesions and growth retardation caused by L. intracellularis infections. According to Collins [5] and Johanssen [24], fecal shedding needs to be more than 106 GE/g feces and 107 GE/g feces before having an impact on growth. Similarly, Pedersen [25] found that the lower limit for detection in pigs with histological lesions was 4.8 Log10 GE/g feces.
Current uncertainties regarding raw materials supply, pork prices, and global trade redirect the focus of the swine industry to improving feed efficiency and sustainability. Consequently, the optimization of growth and performance using less feed is of paramount importance for the economic viability of every pig herd, particularly those subclinically infected. This strategy is included in the road map of every control program for L. intracellularis infections, and different tools are available to achieve this goal, including antimicrobial treatment, vaccination, and the optimization of management, hygiene, and biosecurity [2]. In the current context of antimicrobial reduction, vaccination is gaining relevance in this control approach. Vaccination, both intramuscular and intradermal, has been proven to be an excellent tool to reduce clinical signs, bacterial shedding in feces, and performance losses in clinically affected farms [26,27,28,29,30,31]. However, it is still unknown whether vaccination could positively impact performance in this large population of farms with subclinical infection and low fecal bacterial shedding. Our study aims to address this knowledge gap, frequently acknowledged in practice by pig producers and practitioners.
The overall aim of this study was to assess the clinical, growth, and carcass performance of intramuscular or intradermal vaccination against L. intracellularis in a herd with subclinical infection and low fecal bacterial shedding. Two studies were carried out:
- Field trial 1 was performed to elucidate whether there was an association between the growth and fecal shedding of L. intracellularis in unvaccinated and intramuscularly vaccinated pigs in a farm with subclinical infection.
- Field trial 2 was performed to assess the impact of intradermal vaccination against L. intracellularis on clinical performance and carcass quality in a herd with subclinical infection.
2. Materials and Methods
2.1. Herd Selection
A herd with a history of subclinical ileitis and a low bacterial load in feces was selected for both studies. In our study, subclinical ileitis was defined (adapted from Winkelman) [8] by the absence of ileitis-associated clinical signs and/or mortality, as well as by the detection of the bacterium by qPCR. A low bacterial fecal load was considered when the quantification of genomic copies by RT-PCR (Bactoreal®, Kit Lawsonia intracellularis, Ingenetix GmbH, Wien, Austria) [32] did not exceed 4 Log10 GE copies/µL. This cut-off was established based on several scientific references [5,24,25] and guided by practical experience in the field (personal communication).
Before the start of the study, cross-sectional fecal sampling was performed, and qPCR testing was performed for the quantification of L. intracellularis. Pigs at 6, 9, 12, 15, 18, and 21 weeks of age were screened. For each age group, a total of 15 fresh fecal samples were collected from the floor of different pens and pooled by 5. Each pool included at least 25 g of feces and was analyzed by PCR for the detection of L. intracellularis, Brachyspira hyodysenteriae (Kylt® BHP Triplex, SAN Group Biotech Germany GmbH, Höltinghausen, Germany), Brachyspira pilosicoli (Kylt® BHP Triplex, SAN Group Biotech Germany GmbH, Höltinghausen, Germany), and Salmonella enterica Tiphimurium (Kylt® Salm, SAN Group Biotech Germany GmbH, Höltinghausen, Germany). Only the samples from 9-, 12-, and 15-week-old pigs tested positive for L. intracellularis by PCR. The bacterial loads for those positive age groups were in the ranges 0–2.9, 1.5–2.7, and 2.8–3.8 Log10 copies/µL, respectively. All samples tested negative by PCR for B. hyodysenteriae and Salmonella Typhimurium, whereas B. pilosicoli was only detected at 18 weeks of age.
The herd was a farrow-to-finish farm housing 400 sows, with a commercial health status, working in a 3-week-batch farrowing system and weaning at 28 days of age. Prior to weaning, piglets were routinely vaccinated against PCV2 and M. hyopneumoniae (Porcilis® PCV M Hyo, MSD Animal Health, Rahway, NJ, USA). No routine antimicrobial medication was administered to the piglets during the nursery period (from weaning to 11 weeks of age), nor during the fattening period (from 12 weeks to slaughter). Only zinc oxide was administered in the first two weeks post-weaning to control post-weaning diarrhea. Prior to the initiation of the study, ethical approval was obtained from University of Leeds (20211404ATKA).
2.2. Field Trial 1
A randomized, controlled, blind, side-by-side trial was performed. In total, 240 pigs were included in the study and individually tagged within 24 h of birth. On the day of vaccination (28 days of age; day 0), all piglets were weighed and ranked within litter from the heaviest to the lightest; then, every alternate pig was allocated to either the Vaccination [V; n = 120; vaccinated with Porcilis® Lawsonia (lyophilized pellet) (MSD Animal Health, Rahway, NJ, USA) mixed with Porcilis® PCV M Hyo (2 mL) (MSD Animal Health, Rahway, NJ, USA)] or Control [C; n = 120; Porcilis® PCV M Hyo (2 mL)] group. Porcilis® Lawsonia consists of both a freeze-dried antigen fraction and a solvent fraction (Emunade®). The freeze-dried antigen could be reconstituted prior to use in the solvent. Alternatively, it could be also reconstituted in Porcilis PCV® M Hyo, which is based on the same adjuvant [26]. Therefore, both vaccines given to Group V could be administered in an associated mixed use. No sham-vaccinated group was included due to ethical reasons. Immediately following vaccination, pigs were weaned into slatted pens (20 pigs per pen) according to the vaccination type, matched as far as possible for litter origin, sex, and live weight. Pigs were fed standard commercial diets following weaning and through to slaughter. These diets provided sufficient nutrient levels to satisfy minimum requirements and normal growth performance. Pigs had ad libitum access to feed and water at all times.
A longitudinal sampling was carried out by collecting feces on days 0, 21, 42, 63, 84, and 105 from the same individual pig at each time point. Fecal samples (n = 30/group) were taken directly from the anus at defecation. The fecal bacterial load of L. intracellularis was assessed by qPCR (Bactoreal®, Kit Lawsonia intracellularis, Ingenetix GmbH, Wien, Austria), and the Area Under the Curve (AUC) was calculated for each group. The qPCR assay used is a real-time PCR for the detection of L. intracellularis DNA, targeting the 16S rRNA gene of L. intracellularis [32]. The analytical sensitivity is 10 target copies/PCR reaction. The analytical specificity allows for the detection of so-far-known L. intracellularis strains, with no cross-reactions observed to other bacterial isolates according to manufacturer specifications.
Growth performance was investigated using precision farming technology. The daily weights and daily feed intake of the individual pigs were determined throughout the trial by using pig performance testing technology. From weaning to approximately 35 kg, pigs were housed in a specific weaner grower accommodation with the Pig Insight Asserva System (Asserva, Lamballe-Armor, France). These pens were 3.5 m × 2.5 m and housed 20 pigs each. At 35 kg, the pigs were moved into finisher accommodation with the Pig Performance Nedap ProSense system (Nedap livestock management, Groenlo, The Netherlands). Each nursery pen of 20 was split into two fattening pens of 10. The pens in the finisher accommodation were 2.6 m × 3.5 m and housed 10 pigs each. All automated or mechanical equipment was inspected at least once per day. Pigs were sent to slaughter at approximately 110 kg live weight. Average Daily Gain (ADG), Average Daily Feed Intake (ADI), and Feed Conversion Ratio (FCR) were calculated using the data generated by the aforementioned technology. The association between bacterial load and ADG, ADI, and FCR for the 7 days prior to the peak of bacterial shedding was determined.
Pig health was assessed and recorded daily. A record was kept of any veterinary interventions. Mortality was documented during the whole trial and post-mortem examinations were carried out for those casualties at a certified diagnostic lab.
2.3. Field Trial 2
A randomized, controlled, blind, side-by-side trial was performed. In total, 240 pigs were included in the study and individually tagged within 24 h of birth. On the day of vaccination (28 days of age; day 0) all piglets were weighed and ranked within litter from the heaviest to the lightest; then, every alternate pig was allocated to either the Vaccination group [V; n = 120; vaccinated with Porcilis® Lawsonia ID (lyophilized pellet) (MSD Animal Health, Rahway, NJ, USA) mixed with Porcilis® PCV ID (0.2 mL) (MSD Animal Health, Rahway, NJ, USA) and administered concurrently with Porcilis® M Hyo ID Once (0.2 mL) using and IDAL® Twin (Henke Sass Wolf GmbH, Tuttlingen, Germany)] or the Control group [C; n = 120; Porcilis® PCV ID (0.2 mL) and Porcilis® M Hyo ID Once (0.2 mL) administered concurrently using and IDAL® Twin]. No sham-vaccinated group was included due to ethical reasons. Immediately following vaccination, pigs were weaned following the same protocol as described for study 1.
A longitudinal sampling was performed by collecting feces on days 0, 21, 42, 63, 84, 105, and 126 from the same individual pig at each time point. Fecal samples (n = 30/group) were taken directly from the anus at defecation. The fecal bacterial load of L. intracellularis was assessed by a qPCR test (Bactoreal®, Kit Lawsonia intracellularis, Ingenetix GmbH, Wien, Austria), and the AUC was calculated for each group.
Ileitis-associated mortality %, treatment incidences, scour incidences, and tail biting % were registered. At slaughter, carcass quality was assessed by recording carcass weight, back fat level (at P2 level; cm), and Lean Meat % (LM%).
2.4. Statistical Analysis
A Kruskal–Wallis test was run to evaluate bacterial shedding and carcass characteristics. Proportions of mortality, antibiotic treatment, and tail bitten incidences in each group were assessed by Chi-square analysis.
Additionally, in study 1, the performance data were analyzed using a mixed linear model procedure (IBM SPSS Statistics 26) with pig as the experimental unit. Treatment was included as a fixed effect. A random effect of pen nested within treatment was also included. Normality was assessed using the Shapiro–Wilk test. Any data that were not normally distributed were subject to a log10 transformation prior to analysis. Transformed data were back-transformed prior to inclusion in the presented results. A linear regression was used to assess the association between the number of L. intracellularis bacteria in feces and ADG, ADI, and FCR for the 7 days prior to the pigs testing positive. A sample size of 110 pigs per group was estimated to detect a 6% difference in ADG between treatment groups with a power of 90%. Then, 10 pigs were added to top each group up to 120 pigs to cover any losses.
3. Results
3.1. Field Trial 1
3.1.1. Fecal Bacterial Load
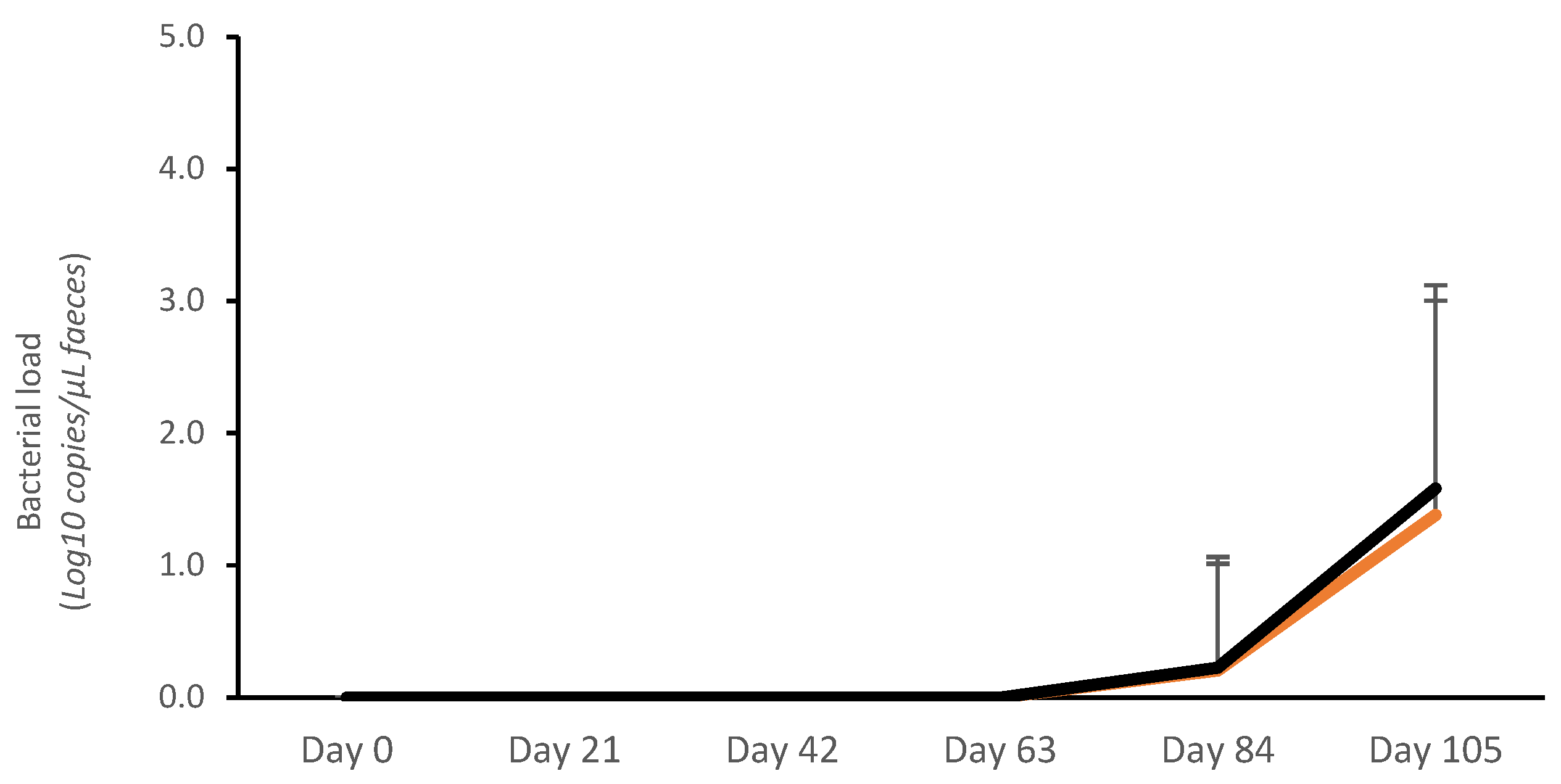
No bacterial shedding was detected from day 0 to day 63. The average bacterial load (Log10 copies/μL) was very low in both vaccinated and control pigs on day 84 (V: 0.19 ± 0.13; C: 0.20 ± 0.14; p > 0.05) and day 105 (V: 1.17 ± 0.27; C: 1.48 ± 0.27; p > 0.05). The average AUC for fecal bacterial load on days 0–105 was numerically lower for vaccinates (14.96 ± 20.64 Log10 copies/μL) than for controls (17.53 ± 19.85 Log10 copies/μL) (p > 0.05) (Figure 1). There was also a numerical reduction in the proportion of pigs that tested positive for L. intracellularis by qPCR in the vaccinated group (46.7%; 14/30) compared with the controls (63.3%; 19/30) (p > 0.05).
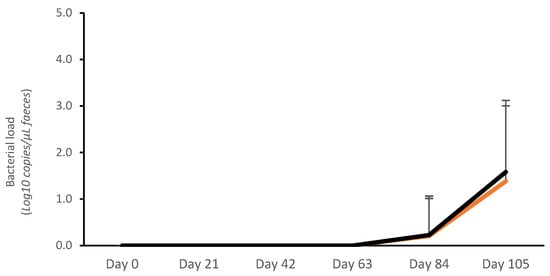
Figure 1.
Average Lawsonia intracellularis shedding in feces in vaccinated pigs (orange) and control pigs (black).
3.1.2. Performance
Live weight was similar for each treatment group on day 0 (V: 8.85 kg; C: 8.86 kg; p > 0.05). Subsequently, there was a tendency for vaccinated pigs to be heavier from day 7 to day 105 (p < 0.1). This average weekly live weight was consistently higher in vaccinated pigs than in the controls, being significant only on day 77 (2.2 ± 0.77 kg). Overall, the treatment groups performed similarly (Table S1).
3.1.3. Association Between Fecal Load and ADG, ADI, and FCR
The peak of bacterial load was confirmed on day 105 (Figure 1), and this was concurrent with the largest proportion of PCR-positive samples. Therefore, the association between bacterial load and ADG, ADI, and FCR was calculated for the 7 days prior to this peak of infection (day 99–day 105). During this period, an association between the number of L. intracellularis bacteria in feces and ADG, ADI, and FCR was described (Table 1).

Table 1.
Association (Pearson correlation coefficient; PCC) between the number of L. intracellularis bacteria in feces and ADG, ADI, and FCR in control non-vaccinated pigs and pigs vaccinated against Lawsonia intracellularis.
A significant negative correlation between the ADG of control pigs and Log10 bacteria per gram of feces was described (R2 = −0.193; p < 0.05); the higher the bacterial load in feces, the lower the rate of gain. This negative correlation was not observed in vaccinated pigs (Figure S1).
There was a negative association between the ADI of control pigs and Log10 bacteria per gram of feces (R2 = −0.111; p < 0.1); the higher the bacterial load in feces, the lower feed intake. This negative correlation was not observed in vaccinated pigs (Figure S2).
A positive correlation between the FCR of control pigs and Log10 bacteria per gram of feces was detected (R2 = 0.136; p < 0.05); the higher the bacterial load in feces, the higher the FCR. No significant correlations were observed between FCR and Log10 bacteria per gram of feces in vaccinated pigs (Figure S3).
3.1.4. Clinical Parameters
Clinical signs compatible with L. intracellularis infection were not recorded. Ileitis-associated mortality was not reported. There was no difference in the number of pigs treated with antibiotics or that died between treatments throughout the trial (Table 2). Tail biting was documented in both groups, and the proportion of pigs affected was not significantly different between the groups.
Table 2.
Clinical parameters recorded in control non-vaccinated pigs and pigs vaccinated against Lawsonia intracellularis. No statistical differences were described between groups.
3.2. Field Trial 2
3.2.1. Fecal Bacterial Load
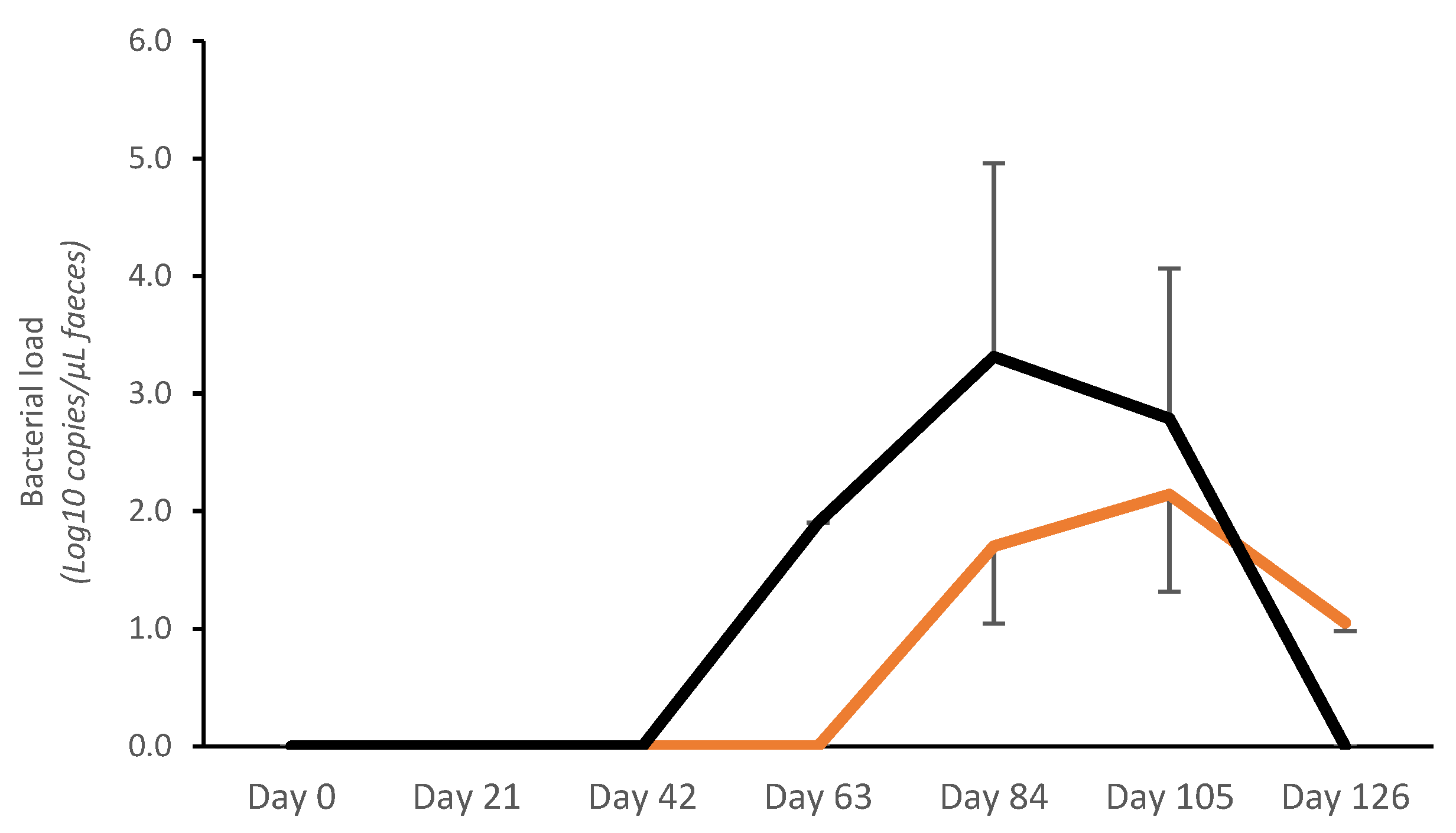
No bacterial shedding was detected before day 63 in controls, whereas in vaccinated pigs, fecal shedding started later, on day 84 (Figure 2). A significantly lower bacterial load (Log10 copies/μL) was detected on day 84 in vaccinated pigs (V: 1.70 ± 0.66; C: 3.31 ± 1.65; p < 0.05). The average AUC (bacterial shedding Log10 copies/μL) from day 0 to day 126 was significantly lower in vaccinates (20.72 ± 25.93) compared with the controls (40.23 ± 39.10) (p < 0.05) (Figure 2).
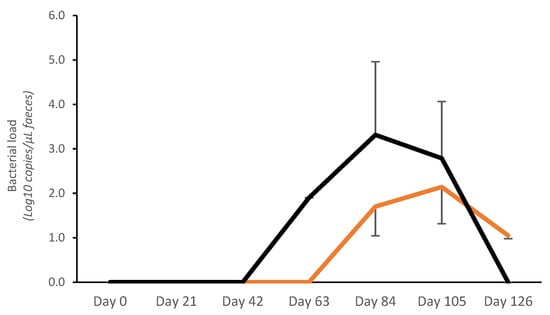
Figure 2.
Average Lawsonia intracellularis shedding in feces in vaccinated pigs (orange) and control pigs (black). Average Area Under the Curve (bacterial shedding Log10 copies/μL) from day 0 to day 126 was significantly lower for vaccinates (20.72 ± 25.93) compared with controls (40.23 ± 39.10) (p < 0.05).
3.2.2. Clinical Parameters
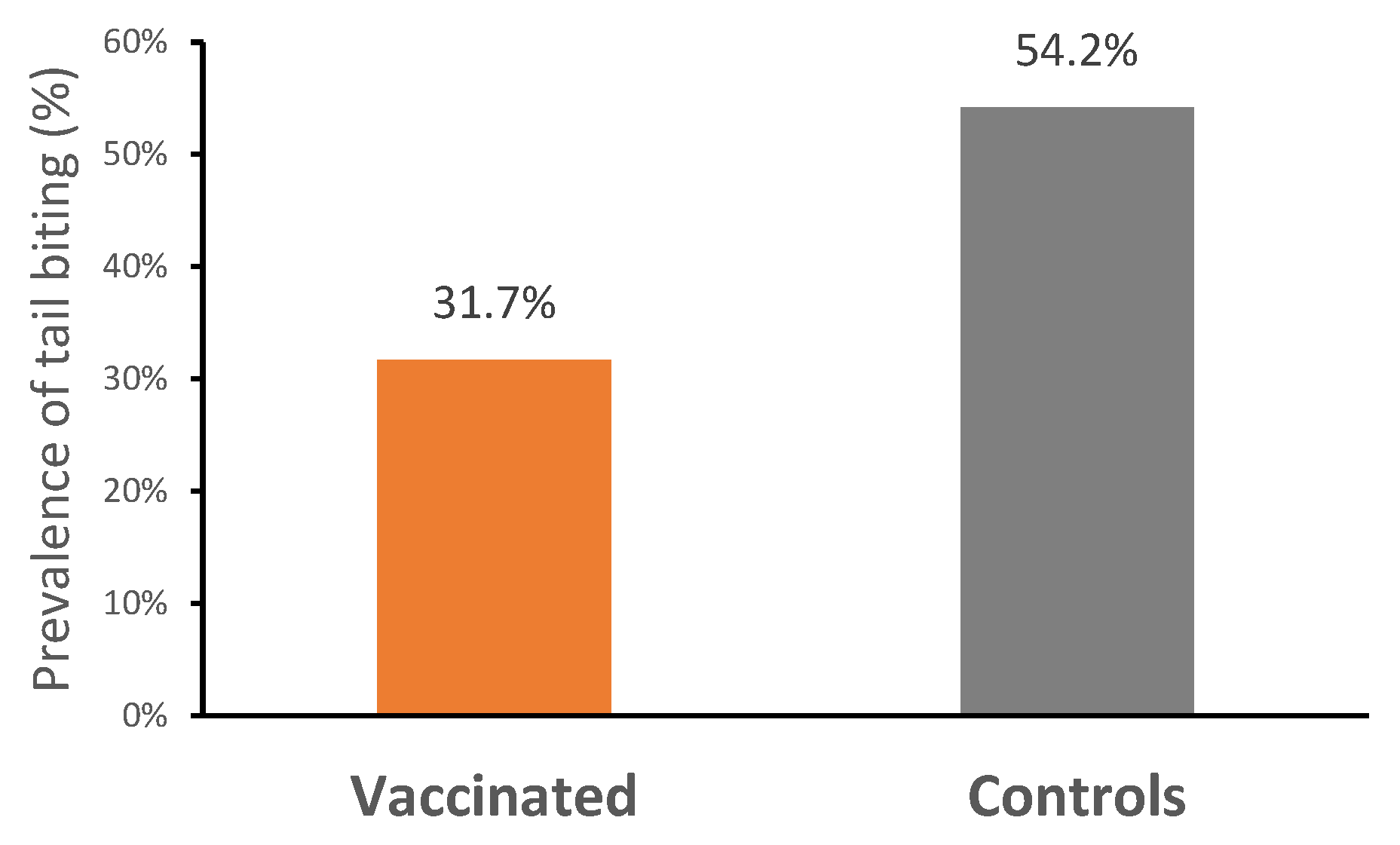
The mortality rate was very low across the entire study (V: 0%; C: 0.8%; p > 0.05), and no ileitis-associated mortality was recorded. During the first 21 days of the study, a total of 14 piglets were removed from the trial (V: 10; C: 4), due to a failure to adapt to the electronic feeders, which led to low feed intake and poor growth. There was no difference in the number of pigs treated with antibiotics between treatments throughout the trial (Table 3). Treatment incidences related to scour were similar for both groups (V: 11; C: 12; p > 0.05). Vaccinated pigs had a significantly lower prevalence of tail biting (31.67%; n = 38) compared with control pigs (54.17%; n = 65) (p < 0.05) (Figure 3).
Table 3.
Clinical parameters and carcass quality in vaccinated and control pigs.
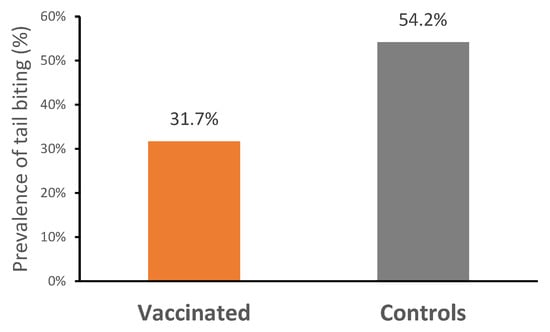
Figure 3.
Prevalence of tail biting in vaccinated and control pigs (p < 0.05).
3.2.3. Carcass Quality
Vaccinated pigs had less back fat (10.5 ± 0.14 mm vs. 10.9 ± 0.14 mm; p < 0.07) and greater LM% (62.7 ± 0.12% vs. 62.1 ± 0.12%; p < 0.05) compared with non-vaccinated pigs (Table 3). No significant differences were seen in the other parameters.
4. Discussion
This study was designed to assess the benefits of vaccination against L. intracellularis in a herd with subclinical infection and low fecal bacterial shedding. Taking both field trials together, piglet vaccination on a farm with subclinical infection led to lower fecal bacterial shedding, a partial reduction in tail biting, and an improvement in carcass quality. The fecal load of L. intracellularis had a negative effect only on the growth of non-vaccinated pigs, whereas growth in vaccinated pigs was not impacted.
While subclinical L. intracellularis infections are very common, they remain often underdiagnosed. Consequently, no control strategies are set in place, and the economic performance of those herds is negatively impacted. There is limited research regarding the control of subclinical infections. This study reveals the impact of subclinical infection and sheds light on the use of vaccination to help control any negative impacts, addressing a knowledge gap commonly encountered in the field by pig farmers and practitioners. It also elucidates the relationship between fecal shedding and growth in subclinical cases. In addition, this is the first scientific report showing a partial reduction in tail biting after L. intracellularis vaccination.
The preliminary screening carried out before the start of the trials confirmed that the selected farm suffered from a subclinical L. intracellularis infection. The absence of both ileitis-like signs and mortality corroborated the suitability of the farm for the study. A low bacterial load in feces was detected before the start of the study. This load was lower than the threshold previously described as the minimum limit for growth retardation [5,21]. However, it was comparable to the cut-off set by Pedersen et al. [4] for the presence of microscopic lesions caused by L. intracellularis. It is acknowledged that a high-shedding farm would be more suitable to test vaccine efficacy. However, the study aimed to investigate whether vaccination was still beneficial under these specific conditions.
Fecal bacterial shedding was reduced in vaccinated pigs compared with the controls in trial 2. It is important to note the lack of significance between groups in trial 1. As the overall load was extremely low in trial 1 compared with the threshold for detection of histological lesions (4.8 Log10 GE/g feces) [25], non-significant results are not surprising. However, both the lower AUC and the smaller proportion of PCR-positive pigs in the vaccinated group, even if only numerically different, supported this trend of lower shedding. It is important to note the differences in dynamics and fecal load between the batches of both trials. This is in line with previous reports [31,33] that revealed that the variability in the dynamics and epidemiology of L. intracellularis, even between consecutive batches, was a common observation in the field. To avoid underdiagnosing subclinical infections, there is a need for serial longitudinal fecal sampling, even including several batches. This emphasizes the unsuitability of single time fecal sampling for the same purpose. Due to the labor-intensive protocol and high cost of facility usage, our research did not include multiple batches as replicates, which could be a limitation. However, the pre-screening of the herd prior to the start and its history of subclinical ileitis had confirmed the suitability of the herd for the purpose of this study.
Despite a tendency for vaccinated pigs to grow consistently better and faster than controls every week of the study, both treatment groups performed similarly at end of the field trial 1. The lack of significant differences could be explained by the late L. intracellularis infection, occurring only four weeks before slaughter, together with the low bacterial shedding in the investigated batch. It is hypothesized that vaccinated pigs did not have enough time between infection and slaughter to express their full potential in growth, as there was not enough time for the unvaccinated, non-protected controls to suffer from the expected retardation in growth. The great variation in infection dynamics between batches likely played a role here. While the pre-screening revealed a low bacterial load (<4 Log10 copies/µL) present between 9 and 15 weeks of age before the start of the study, the batch included in the trial showed a totally different infection pattern, i.e., a late infection around 22 weeks of age characterized by very low fecal shedding (<2 Log10 copies/µL). A future corrective measure when investigating performance in farms with subclinical infections would be the inclusion of several batches as a replicate.
Several attempts have been made to find a relationship between growth rate and the level of antibodies against L. intracellularis. However, they have not been successful. In one of those studies [21], a lack of association was shown between reduced growth rate and the level of antibodies against this bacterium. However, the same authors found also that fecal shedding, as measured by qPCR, was a better indicator of growth retardation. More specifically, a high fecal load (>106 copies/g of feces) was a significant risk factor for low growth. Similarly, an Australian study [5] described that the fecal shedding of L. intracellularis correlated positively with the severity of histopathological lesions of Proliferative Enteropathy and negatively with ADG. In this experimental challenge trial, pigs shedding from 107 to 108 copies/g experienced a large growth retardation (ADG: −131 g/day) compared with non-challenged pigs, whereas when lower shedding was detected (106–107), a smaller impact on ADG was reported (ADG: −15 g/day). These observations confirmed that antibodies reflect the immune response to infection, whereas fecal bacterial load mirrors the infection level in the intestine. Another Danish longitudinal field study also reported a negative correlation between bacterial load in feces and ADG in naturally infected but non-vaccinated pigs [20].
While the aforementioned studies described this association in animals clinically affected with high bacterial shedding, others have also reported a similar association in pigs with subclinical infections [19]. These authors were able to experimentally induce the transition from clinical to subclinical infection and impair performance by inoculating different doses of L. intracellularis, demonstrating a dose-dependent response. Growth retardation (ADG) was the most sensitive indicator of disease, and it revealed the impact of subclinical infections. Lower ADG (−37%; −63 g/day) and feed efficiency (−27%) were recorded at the lowest challenge dose (3.2 × 104 organisms). In the absence of clinical signs of disease, pigs from the lowest challenge group presented sporadic shedding as well as macro- and microscopic lesions. The severity of gross and histological lesions was also dose-dependent.
Abundant research has been carried out under both experimental and field conditions, including both clinically and subclinically infected pigs. However, it is still unknown whether vaccination could have an influence on this association in farms with subclinical infection and low fecal bacterial shedding. One of the aims of our study was to answer this. In our case, an increasing number of L. intracellularis log10 bacteria/g of feces was significantly associated with decreasing ADG, but this observation was only confirmed in non-vaccinated pigs: the higher the fecal bacterial load, the lower the rate of weight gain. Conversely, the growth of vaccinated pigs was not negatively affected by the load of L. intracellularis in feces. This supports the field observations suggesting that vaccination protects against the disease even if the bacterium is still detected by PCR during routine diagnosis in vaccinated pigs. Now it also indicates that growth is not impacted. This is in line with the benefits shown by both intramuscular and intradermal vaccination on the reduction in L. intracellularis shedding in both intestinal mucosa and feces [26,28]. However, it was also shown that the bacterium could still be found, and total elimination was not achieved.
Our findings confirm the impact of these subclinical infections on growth and corroborate the observations by Paradis et al. [19] but slightly conflict with the ones by Pedersen et al. [20]. In their study, no differences in ADG were reported between negative pigs and pigs with low numbers of bacteria in feces [20]. In order to reduce bias, our farm selection followed similar standards to those included in the Danish study. Although our study herd was not an SPF farm, the preliminary screening confirmed the absence of most enteric diseases that may have interfered with L. intracellularis. The correlation between growth and shedding was performed taking into consideration only the period of seven days prior to the collection of the samples, which was the same period used in the association modeled by Pedersen et al. [20]. The assay used to quantify bacterial fecal load differs between studies. To the authors’ knowledge, there is only one study comparing different PCR assays for this bacterium [34]. In the research study, two RT-PCRs were included, and both assays presented similar sensitivity and specificity in detecting L. intracellularis in fecal samples. The first assay [32], which is the same commercial assay used in our study, presented 97% sensitivity and 85% specificity, as well as 100% analytical specificity. In addition, it was reported that the lower detection limit of the commercial kit was 4 × 104 GE/g of feces. However, the lower quantification limit, robustness, and precision of the test were not reported. For the second assay, the diagnostic performance of the test was reported concisely [22], showing similarities to the previous one (sensitivity of 97%; specificity of 34%; analytical specificity of 100%; lower limit of quantification of 101 GE/µL). In view of this similar diagnostic performance, it was decided to use the commercial kit [32], which had high availability across Europe and is used commercially to support diagnosis in practice.
Regarding ADI and FCR, our study reported a similar trend to the association described for ADG. In non-vaccinated pigs, the higher the bacterial load, the lower the feed intake and the lower the feed efficiency reported. This was not the case for vaccinated pigs. It is assumed that non-vaccinated, unprotected pigs could have suffered from the disease to a higher extent compared with vaccinated ones, developing more severe intestinal lesions, having a lower appetite, and facing suboptimal nutrient absorption which eventually led to lower performance. Unfortunately, this assumption could not be confirmed as an ileum lesion scoring was not carried out. In a previous study [4], no correlation was found between fecal load and gross lesions. However, these authors demonstrated that an increase in histopathology and immunohistochemistry scores was positively correlated with L. intracellularis shedding. The lack of microscopic investigation of intestinal tissue is a limitation of our study. Assessing histological lesions would have been beneficial in explaining the growth differences between groups and potentially linking those differences to nutrient absorption at the gut level. However, it is important to note that interpreting histopathology results in practice is challenging, especially when this is performed only at slaughter time rather than through serial investigations, as lesions may heal by the time of slaughter [35].
The use of precision farming technology for the recording and investigation of growth performance was a great asset for the research. It facilitated the detection of patterns and small deviations, while routine weighing at entry and before slaughter would have missed this information. Moreover, it optimized the detection of piglets with lower feed intake and accelerated their exclusion from the study, avoiding important bias. Both precision farming technologies used for live weight recording (the Pig Insight Asserva System and the Pig Performance Nedap ProSense system) presented the advantage of recording it multiple times per pig per day, increasing accuracy and leading to a data set with multiple dependent measurements. Therefore, a Generalized Mixed Linear Model, which can account for the non-independence of the data by adding random effects, was used.
As expected in a farm with subclinical infection, no ileitis-associated clinical signs nor mortality was reported in either group during the study. Similarly, antimicrobial incidences and overall mortality were low and equal between groups. Intramuscular vaccination against L. intracellularis has been reported as an alternative tool to reduce antimicrobial use in cases of both acute and chronic ileitis [36,37]. Further research is needed to elucidate if antimicrobial consumption can be also reduced in farms with subclinical infections, where prophylactic use is often still implemented, especially outside of Europe.
Tail biting was reported in both studies. Surprisingly, only vaccinated pigs from trial 2 presented a significantly lower prevalence compared with the controls. To the authors’ knowledge, this is the first scientific report of partial reduction in tail biting after vaccination against L. intracellularis. It is unknown why no differences were reported between groups in trial 1. The low bacterial load in feces in trial 1, as well as the use of different routes of administration in each study, might have influenced this outcome, but this study design does not allow us to demonstrate causality and make such a conclusion. Similarly, from the data reported in this study, it cannot be concluded that vaccination against L. intracellularis will have a protective effect against tail biting, as there are some limitations. Firstly, the study did not aim to investigate this question or elucidate the mechanism of action. Therefore, it was not designed in such a way to answer this. Secondly, more thorough research would have been needed, including histology of tail lesions, stress markers, microbiome analysis, and/or a complete risk factor analysis. The multifactorial nature of tail biting is continuously being explored, and all mechanisms behind this behavior are not yet fully understood, making it difficult to predict and control [38]. Risk factors for tail biting may be categorized in those related to the pig or to the environment. Pig-associated factors include but are not limited to genetics, gender, health status, growth, age, and behavior. Environmental factors include feeding, space allowance, climate, flooring, access to manipulable materials, housing, etc. All pig- and environment-related factors described above were similar between vaccinated and non-vaccinated pigs. Pigs were randomized at arrival, housed in the same compartment under uniform environmental conditions across the barn, under similar stock density, receiving the same diet, and having similar access to manipulable materials. Therefore, the absence of any bias that might have influenced the tail biting, growth, or fecal shedding in favor of one group or another could be confirmed. Recent research has provided new evidence of a relationship between the gut microbiome and tail biting behavior, pointing at the gut–brain axis as a potential mechanism to partially explain this phenomenon [39,40,41,42,43]. While studies indicate that L. intracellularis can modulate the porcine gut microbiome following either infection [9,44,45] or vaccination [46], no documented association exists between tail biting behavior and this bacterial infection to date. Furthermore, the unexpected findings in our study underscore the need for more extensive research employing an interventional design to investigate behavioral patterns and stress factors associated with gut microbiome dysbiosis and the development of tail lesions.
At slaughter, vaccinated pigs performed better than controls in terms of Lean Meat percentage, presenting less back fat. A similar improvement in carcass quality after L. intracellularis intramuscular vaccination has been previously shown [47]. In this study, the authors reported greater carcass weight followed by an increased percentage of lean meat in the ham, loin, and shoulder after intramuscular vaccination. Carcass weight was also increased in another study carried out in Brazil after intramuscular vaccination [48]. In this case, vaccination also improved the integrity of the intestinal wall, leading to an increased yield of casing obtention (meters of casing/intestine) and sausage fill (kg of sausage/meter of casing). Gut inflammation in pigs can significantly impact energy deposition in the carcass. When pigs experience gut inflammation, it often leads to a reduction in feed intake and nutrient absorption, which can negatively affect growth rates and overall carcass quality. Inflammation can trigger the release of pro-inflammatory cytokines, which may divert energy away from muscle synthesis and towards immune responses, leading to increased fat deposition instead of muscle growth [49]. Conversely, low gut inflammation is associated with improved nutrient absorption and metabolic efficiency. When inflammation is minimized, the body can redirect energy towards anabolic processes, including muscle formation. This is facilitated by enhanced biosynthetic pathways, such as protein synthesis and lipid metabolism, which favor muscle growth over fat deposition [50]. Therefore, managing gut health is crucial to optimizing muscle formation and improving carcass quality in pigs. In our study, it was presumed that the reduction in L. intracellularis organisms at the gut level in vaccinated pigs led to lower gut inflammation, better gut integrity, and consequently, an improved digestion process and more efficient nutrient absorption. Gut microbiota plays a pivotal role in pig metabolism by leading the fermentation of dietary fiber, the production of Short-Chain Fatty Acids (SCFAs), and the absorption of their metabolites at the intestinal level [51]. SCFAs are instrumental in the protein and fat metabolism of skeletal muscle, affecting muscle fiber formation and intramuscular adipogenesis [51]. Interestingly, they are known to have antimicrobial properties by inhibiting the growth of common enteric pathogenic bacteria and hence supporting the control of intestinal infections [52,53]. However, this study also acknowledges its limitations, particularly the lack of investigation into the specific impacts of vaccination on gut inflammation and gut microbiota composition and how this impacts growth performance and carcass quality. Microbiome analysis, histopathology, metabolic data, and investigations into nutrient absorption will be essential to fully understanding these relationships and develop effective strategies for optimizing energy deposition in the carcass of pigs. By addressing these gaps, we can further enhance the benefits of vaccination and improve the overall efficiency and sustainability of swine production systems.
5. Conclusions
In this study, it was shown that vaccination against L. intracellularis in a herd with subclinical infection and low fecal bacterial shedding led to a reduction in fecal shedding, a lower prevalence of tail biting, and an improvement in carcass quality. Notably, it was found that growth performance in non-vaccinated pigs was negatively correlated with the fecal load of L. intracellularis, whereas vaccinated pigs exhibited no such negative impact, indicating a protective effect of vaccination.
These findings suggest that vaccination can serve as a valuable tool for enhancing farm profitability by improving lean meat yield and mitigating behavioral issues such as tail biting, which can lead to economic losses. By integrating vaccination into herd management practices, producers may not only improve animal welfare but also optimize production efficiency.
Further research is essential to fully elucidating the mechanisms by which vaccination influences tail biting behavior and energy deposition in pig carcasses, paving the way for more effective strategies in swine health management.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/vaccines13070728/s1, Table S1. Overall performance of control non-vaccinated pigs and pigs vaccinated against Lawsonia intracellularis. Figure S1. Pearson correlation coefficient between Average Daily Gain (ADG) and bacterial load in feces (qPCR Law; Log10 copies/µL of feces) in control (left) and vaccinated (right) pigs. Figure S2. Pearson correlation coefficient between Average Daily Intake (ADI) and bacterial load in feces (qPCR Law; Log10 copies/µL of feces) in control (left) and vaccinated (right) pigs. Figure S3. Pearson correlation coefficient between Feed Conversion Ratio (FCR) and bacterial load in feces (qPCR Law; Log10 copies/µL of feces) in control (left) and vaccinated (right) pigs.
Author Contributions
Conceptualization, R.D.P.S., H.S., S.v.B., and A.E.T.; methodology, R.D.P.S., H.S., S.v.B., and A.E.T.; formal analysis, A.E.T.; investigation, A.E.T.; resources, R.D.P.S.; writing—original draft preparation, R.D.P.S.; writing—review and editing, H.S., S.v.B., and A.E.T. All authors have read and agreed to the published version of the manuscript.
Funding
This research study was partially funded by MSD Animal Health (Rahway, NJ, USA) and incorporated by Royal Charter with registered number RC000658.
Institutional Review Board Statement
The animal study protocol was approved by the Ethics Committee of University of Leeds (20211404ATKA; 14 March 2021).
Informed Consent Statement
Not applicable.
Data Availability Statement
Please add the corresponding content of this part.
Acknowledgments
We wish to thank the research team for their help in conducting the in vivo part at the Research farm from the University of Leeds, UK, and the lab personal at Center of Diagnostics Solutions in Boxmeer, The Netherlands.
Conflicts of Interest
This study was partially funded by MSD Animal Health. R.D.P.S., H.W. and S.v.B. are employees of MSD Animal Health. The remaining authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Abbreviations
The following abbreviations are used in this manuscript:
| qPCR | real-time quantitative polymerase chain reaction |
| V | Vaccination group |
| C | Control group |
| AUC | Area Under the Curve |
| ADG | Average Daily Gain |
| ADI | Average Daily Feed Intake |
| FCR | Feed Conversion Ratio |
| LM% | Lean Meat percentage |
| SE | Standard Error |
| SPF | Specific Pathogen Free |
| SCFAs | Short-Chain Fatty Acids |
References
- Lawson, G.H.K.; McOrist, S.; Sabri, J.; Mackie, R.A. Intracellular Bacteria of Porcine Proliferative Enteropathy: Cultivation and Maintenance In Vitro. J. Clin. Microbiol. 1993, 31, 1136–1142. [Google Scholar] [CrossRef] [PubMed]
- Vannucci, F.A.; Gebhart, C.J.; McOrist, S. Proliferative Enteropathy. In Book Diseases of Swine, 11th ed.; Zimmerman, J.J., Karriker, L.A., Ramirez, A., Schwartz, K.J., Stevenson, G.W., Zhang, J., Eds.; John Wiley & Sons, Inc.: New York, NY, USA, 2019; pp. 898–911. [Google Scholar]
- McOrist, S.; Gebhart, C.J.; Boid, R.; Barns, S.M. Characterization of Lawsonia intracellularis gen. nov., sp.nov, the obligately intracellular bacterium of porcine proliferative enteropathy. Int. J. Syst. Bacteriol. 1995, 45, 820–825. [Google Scholar] [CrossRef]
- Pedersen, K.S.; Ståhl, M.; Guedes, R.M.C.; Angen, Ø.; Nielsen, J.P.; Jensen, T.J. Association between faecal load of Lawsonia intracellularis and pathological findings of proliferative enteropathy in pigs with diarrhoea. BMC Vet. Res. 2012, 8, 198. [Google Scholar] [CrossRef]
- Collins, A.M.; Barchia, I.M. The critical threshold of Lawsonia intracellularis in pig faeces that causes reduced average daily weight gains in experimentally challenged pigs. Vet. Microbiol. 2014, 168, 455–458. [Google Scholar] [CrossRef]
- Arnold, M.; Crienen, A.; Swam, H.; von Berg, S.; Jolie, R.; Nathues, H. Prevalence of Lawsonia intracellularis in pig herds in different European countries. Porc. Health Manag. 2019, 5, 31. [Google Scholar] [CrossRef] [PubMed]
- Vannucci, F.B.; Borges, E.L.; de Oliveira, J.S.V.; Guedes, R.M.C. Intestinal absorption and histomorphometry of Syrian hamsters (Mesocricetus auratus) experimentally infected with Lawsonia intracellularis. Vet. Microbiol. 2010, 145, 286–291. [Google Scholar] [CrossRef]
- Winkelman, N. Subclinical ileitis: Diagnostic monitoring, R2, and economics. In Proceedings of the 49th Annual Meeting of the American Association of Swine Veterinarians AASV, San Diego, CA, USA, 3–6 March 2018; pp. 488–493. [Google Scholar]
- Karuppannan, A.K.; Opriessnig, T. Lawsonia intracellularis: Revisiting the Disease Ecology and Control of This Fastidious Pathogen in Pigs. Front. Vet. Sci. 2018, 5, 181. [Google Scholar] [CrossRef] [PubMed]
- Marsteller, T.A.; Armbruster, G.; Bane, D.P.; Gebhart, C.P.; Muller, R.; Weatherford, J.; Thacker, B. Monitoring the prevalence of Lawsonia intracellularis IgG antibodies using serial sampling in growing and breeding swine herds. J. Swine Health Prod. 2003, 11, 127–130. [Google Scholar] [CrossRef] [PubMed]
- Baldasso, D.Z.; Guizzo, J.A.; Dazzi, C.C.; Paraboni Frandoloso, G.C.; Feronato, C.; von Berg, S.; Carvalho Guedes, R.M.; Wilson, H.L.; Kreutz, L.C.; Frandoloso, R. Development and validation of a flow cytometry antibody test for Lawsonia intracellularis. Front. Immunol. 2023, 14, 1145072. [Google Scholar] [CrossRef]
- Barbosa, J.C.R.; Nicolino, R.R.; Gabardo, M.P.; Gonçalves, J.P.M.; Suarez-Duarte, M.E.; Laub, R.P.; Daniel, A.G.S.; Mariano, G.H.S.; Costa, C.M.; Correia, P.A.; et al. Subsistence swine farming: Seroprevalence and risk factors associated with Lawsonia intracellularis infection in the state of Minas Gerais Brazil in 2016. Trop. Anim. Health Prod. 2023, 55, 314. [Google Scholar] [CrossRef]
- McOrist, S. Prevalence and impact of proliferative enteropathy = ileitis, in East Asia. In Proceedings of the 2nd Asian Pig Veterinary Society. Congress. APVS, Shangri-La, Pasig City, Philippines, 19–21 September 2005. [Google Scholar]
- Yeh, J.Y.; Lee, J.H.; Yeh, H.R.; Kim, A.; Lee, J.Y.; Hwang, J.M.; Kang, B.K.; Kim, J.M.; Choi, I.S.; Lee, J.B. Antimicrobial susceptibility testing of two Lawsonia intracellularis isolates associated with proliferative hemorrhagic enteropathy and porcine intestinal adenomatosis in South Korea. Antimicrob. Agents Chemother. 2011, 55, 4451–4453. [Google Scholar] [CrossRef] [PubMed]
- Wu, Z.; Ling, Y.; Tian, D.; Pan, Q.; Heegaard, P.M.H.; He, C. Seroprevalence of Lawsonia intracellularis antibodies in intensive pig farms in China. BMC Vet. Res. 2014, 10, 100. [Google Scholar] [CrossRef]
- Nguyen, T.C.; Maghfira, C.R.; Young, N.; Samatiwat, K.; Kaewpaluk, C.; Laothanakit, A.; Mungprasittichai, P.; Thanawongnuwech, R.; Panyasing, Y. Seroprevalence of Lawsonia intracellularis Antibodies among Large-scale Swine Farming Systems in Thailand. Thai. J. Vet. Med. 2024, 53, 481–486. [Google Scholar] [CrossRef]
- Wang, L.; Wu, W.; Zhao, L.; Zhu, Z.; Yao, X.; Fan, J.; Chen, H.; Song, W.; Huang, X.; Hua, L.; et al. Fecal PCR survey and genome analysis of Lawsonia intracellularis in China. Front. Vet. Sci. 2024, 11, 1324768. [Google Scholar] [CrossRef] [PubMed]
- Steven McOrist. Defining the full costs of endemic porcine proliferative enteropathy. Vet. J. 2005, 170, 8–9. [Google Scholar] [CrossRef] [PubMed]
- Paradis, M.A.; Gebhart, C.J.; Toole, D.; Vessie, G.; Winkelman, N.L.; Bauer, A.A.; Wilson, J.B.; McClure, C.A. Subclinical ileitis: Diagnostic and performance parameters in a multi-dose mucosal homogenate challenge model. J. Swine Health Prod. 2012, 20, 137–141. [Google Scholar] [CrossRef]
- Pedersen, K.S.; Skrubel, R.; Stege, H.; Angen, Ø.; Ståhl, M.; Hjulsager, C.; Larsen, L.E.; Nielsen, J.P. Association between average daily gain, faecal dry matter content and concentration of Lawsonia intracellularis in faeces. Acta Vet. Scand. 2012, 54, 58. [Google Scholar] [CrossRef] [PubMed]
- Johansen, M.; Nielsen, M.; Dahl, J.; Svensmark, B.; Bækbo, P.; Kristensen, C.S.; Hjulsager, C.K.; Jensen, T.K.; Ståhl, M.; Larsen, L.E.; et al. Investigation of the association of growth rate in grower-finishing pigs with the quantification of Lawsonia intracellularis and porcine circovirus type 2. Prev. Vet. Med. 2013, 108, 63–72. [Google Scholar] [CrossRef]
- Nathues, H.; Holthaus, K.; grosse Beilage, E. Quantification of Lawsonia intracellularis in porcine faeces by real-time PCR. J. Appl. Microbiol. 2009, 107, 16. [Google Scholar] [CrossRef]
- Ståhl, M.; Kokotovic, B.; Hjulsager, C.K.; Breum, S.Ø.; Angen, Ø. The use of quantitative PCR for identification and quantification of Brachyspira pilosicoli, Lawsonia intracellularis and Escherichia coli fimbrial types F4 and F18 in pig feces. Vet. Microbiol. 2011, 151, 307–314. [Google Scholar] [CrossRef]
- Johansen, M.; Nielsen, M.; Dahl, J.; Svensmark, B.; Bækbo, P.; Ståhl, M.; Hjulsager, C.K.; Angen, Ø. Effects of Lawsonia intracellularis on Average Daily Gain in finisher pigs. In Proceedings of the 22nd International Pig Veterinary Society Congress IPVS, Jeju, Republic of South Korea, 10–13 June 2012. [Google Scholar]
- Pedersen, K.S.; Stege, H.; Jensen, T.K.; Guedes, R.; Stahl, M.; Nielsen, J.P.; Hjulsager, C.; Larsen, L.E.; Angen, Ø. Diagnostic performance of fecal real-time polymerase chain reaction for detection of Lawsonia intracellularis-associated proliferative enteropathy in nursery pigs. J. Vet. Diagn. Investig. 2013, 25, 336–340. [Google Scholar] [CrossRef]
- Jacobs, A.A.C.; Harks, F.; Hazenberg, L.; Hoeijmakers, M.J.H.; Nell, T.; Pel, S.; Segers, R.P.A.M. Efficacy of a novel inactivated Lawsonia intracellularis vaccine in pigs against experimental infection and under field conditions. Vaccine 2019, 37, 2149–2157. [Google Scholar] [CrossRef] [PubMed]
- Jacobs, A.A.C.; Harks, F.; Pauwels, R.; Cao, Q.; Holtslag, H.; Pel, S.; Segers, R.P.A.M. Efficacy of a novel intradermal Lawsonia intracellularis vaccine in pigs against experimental infection and under field conditions. Porcine Health Manag. 2020, 6, 25. [Google Scholar] [CrossRef] [PubMed]
- Horsington, J.; Witvliet, M.; Jacobs, A.A.C.; Segers, R.P.A.M. Efficacy of Simultaneous Intradermal Vaccination of Swine against Porcine Circovirus 2, Porcine Reproductive and Respiratory Syndrome Virus, Mycoplasma hyopneumoniae and Lawsonia intracellularis. Animals 2021, 11, 2225. [Google Scholar] [CrossRef] [PubMed]
- Musse, S.L.; Nielsen, G.B.; Stege, H.; Weber, N.R.; Houe, H. Productivity parameters, antimicrobial consumption, and prevalence of enteric pathogens before and after intramuscular vaccination against Lawsonia intracellularis in naturally infected Danish weaner and finisher pig herds. Prev. Vet. Med. 2023, 217, 105973. [Google Scholar] [CrossRef]
- Musse, S.L.; Nielsen, G.B.; Stege, H.; Weber, N.R.; Houe, H. Effect of intramuscular vaccination against Lawsonia intracellularis on production parameters, diarrhea occurrence, antimicrobial treatment, bacterial shedding, and lean meat percentage in two Danish naturally infected finisher pig herds. Prev. Vet. Med. 2023, 212, 105837. [Google Scholar] [CrossRef]
- Del Pozo Sacristan, R.; Lunt, J.; Berkshire, D.; Swam, H. Reduction on Lawsonia intracellularis faecal shedding in pigs chronically infected after intramuscular vaccination against Lawsonia intracellularis. In Proceedings of the 13th European Symposium of Porcine Health Management ESPHM, Budapest, Hungary, 11–13 May 2022. [Google Scholar]
- Lindecrona, R.H.; Jensen, T.K.; Andersen, P.H.; Moller, K. Application of 5’ nuclease assay for detection of Lawsonia intracellularis in fecal samples from pigs. J. Clin. Microbiol. 2002, 40, 984–987. [Google Scholar] [CrossRef]
- Musse, S.L.; Nielsen, G.B.; Stege, H.; Weber, N.R.; Houe, H. Difference in Lawsonia intracellularis between batches and days post entry must be considered when performing diagnostics of ileitis in finisher herds. In Proceedings of the 13th European Symposium of Porcine Health Management ESPHM, Budapest, Hungary, 11–13 May 2022. [Google Scholar]
- Pedersen, K.S.; Holyoake, P.; Stege, H.; Nielsen, J.P. Diagnostic performance of different fecal Lawsonia intracellularis-specific polymerase chain reaction assays as diagnostic tests for proliferative enteropathy in pigs: A review. J. Vet. Diagn. Investig. 2010, 22, 487–494. [Google Scholar] [CrossRef]
- Brandt, D.; Kaim, U.; Baumgartner, W.; Wendt, M. Evaluation of Lawsonia intracellularis infection in a group of pigs in a subclinically affected herd from weaning to slaughter. Vet. Microbiol. 2010, 146, 361–365. [Google Scholar] [CrossRef]
- Del Pozo Sacristan, R.; Lunt, J.; Berkshire, D.; Swam, H. Intramuscular vaccination against Lawsonia intracellularis as a tool to reduce antimicrobial consumption—A case report. In Proceedings of the 26th International Pig Veterinary Society Congress IPVS, Rio De Janeiro, Brazil, 21–24 June 2022. [Google Scholar]
- Marcos Cienfuegos, M.; Jimenez, M.; Menjon, R.; Llorente, C.; Perez, M.L.; Bollo, J.; Romero, A.; Santos, L.; Serrano, D.; Tejedor, T. Control of ileitis in growers and finisher pigs after Lawsonia intracellularis parenteral vaccination. In Proceedings of the 14th European Symposium of Porcine Health Management ESPHM, Thessaloniki, Greece, 31 May–2 June 2023. [Google Scholar]
- Henry, M.; Jansen, H.; Amezcua, M.D.R.; O’Sullivan, T.L.; Niel, L.; Shoveller, A.K.; Friendship, R.M. Tail-Biting in Pigs: A Scoping Review. Animals 2021, 11, 2002. [Google Scholar] [CrossRef]
- Rabhi, N.; Thibodeau, A.; Côté, J.C.; Devillers, N.; Laplante, B.; Fravalo, P.; Larivière-Gauthier, G.; Thériault, W.P.; Faucitano, L.; Beauchamp, G.; et al. Association Between Tail-Biting and Intestinal Microbiota Composition in Pigs. Front. Vet. Sci. 2020, 7, 563762. [Google Scholar] [CrossRef] [PubMed]
- Verbeek, E.; Keeling, L.; Landberg, R.; Lindberg, J.E.; Dicksved, J. The gut microbiota and microbial metabolites are associated with tail biting in pigs. Sci. Rep. 2021, 11, 20547. [Google Scholar] [CrossRef] [PubMed]
- Konig, E.; Heponiemi, P.; Kivinen, S.; Rakkolainen, J.; Beasley, S.; Hukkinen, V.; Norring, M.; Sali, V.; Salminen, S.; Heinonen, M.; et al. Relation of Faecal lactobacilli to manipulative behaviour in pigs. In Proceedings of the 55th Congress of the International Society for Applied Ethology (ISAE), Ohird, North Macedonia, 4–8 September 2022. [Google Scholar]
- Valros, A. Managing tail biting in pigs: Preventing risk factors or docking tails? In Book Advances in Pig Welfare, 2nd ed.; Camerlink, I., Baxter, E.M., Eds.; Woodhead Publishing: Sawston, UK, 2024; pp. 261–287. [Google Scholar] [CrossRef]
- Kobek-Kjeldager, C.; Schönherz, A.A.; Canibe, N.; Pedersen, L.J. Diet and microbiota-gut-brain axis in relation to tail biting in pigs: A review. Appl. Anim. Behav. Sci. 2022, 246, 105514. [Google Scholar] [CrossRef]
- Molbak, L.; Johnsen, K.; Boye, M.; Jensen, T.K.; Johansen, M.; Moller, K.; Leser, T.D. The microbiota of pigs influenced by diet texture and severity of Lawsonia intracellularis infection. Vet. Microbiol. 2008, 128, 96–107. [Google Scholar] [CrossRef] [PubMed]
- Borewicz, K.A.; Kim, H.B.; Singer, R.S.; Gebhart, C.J.; Sreevatsan, S.; Johnson, T.; Isaacson, R.E. Changes in the porcine intestinal microbiome in response to infection with Salmonella enterica and Lawsonia intracellularis. PLoS ONE 2015, 10, e0139106. [Google Scholar] [CrossRef]
- Hankel, J.; Sander, S.; Muthukumarasamy, U.; Strowig, T.; Kamphues, J.; Jung, K.; Visscher, C. Microbiota of vaccinated and non-vaccinated clinically inconspicuous and conspicuous piglets under natural Lawsonia intracellularis infection. Front. Vet. Sci. 2022, 9, 1004506. [Google Scholar] [CrossRef]
- Marcos Cienfuegos, M.; Jimenez, M.; Menjon, R.; Llorente, C.; Perez, M.L.; Bollo, J.; Romero, A.; Santos, L.; Serrano, D.; Tejedor, T. Carcass performance at slaughterhouse after parenteral ileitis vaccination. In Proceedings of the 14th European Symposium of Porcine Health Management ESPHM, Thessaloniki, Greece, 31 May–2 June 2023. [Google Scholar]
- Kamoei, M.; Massaferro, L.; Luiz, H.; Figueiredo MPereira, S.; Marques, B. Immunization against ileitis in swine: A farm to slaughterhouse performance study. In Proceedings of the 13th European Symposium of Porcine Health Management ESPHM, Budapest, Hungary, 11–13 May 2022. [Google Scholar]
- Pluske, J.R.; Hampson, D.J.; Williams, I.H. Factors influencing the structure and function of the small intestine in the weaned pig: A review. Livest. Prod. Sci. 1996, 46, 215–236. [Google Scholar] [CrossRef]
- Pluske, J.R.; Turpin, D.L.; Kim, J.C. Gastrointestinal tract (gut) health in the young pig. Anim. Nutr. 2018, 4, 187–196. [Google Scholar]
- Han, Q.; Huang, X.; Yan, F.; Yin, J.; Xiao, Y. The Role of Gut Microbiota in the Skeletal Muscle Development and Fat Deposition in Pigs. Antibiotics 2022, 11, 793. [Google Scholar] [CrossRef]
- Boyen, F.; Haesebrouck, F.; Vanparys, A.; Volf, J.; Mahu, M.; Van Immerseel, F.; Rychlik, I.; Dewulf, J.; Ducatelle, R.; Pasmans, F. Coated fatty acids alter virulence properties of Salmonella Typhimurium and decrease intestinal colonization of pigs. Vet. Microbiol. 2008, 132, 319–327. [Google Scholar] [CrossRef]
- van Riet, M.M.J.; Vartiainen, S.; Jurgens, G.; Seppala, A.; Rikkola, K.; Vermaut, S.; Peeters, I. Inhibitory effect of SCFA and MCFA on contaminants of liquid pig feed and intestinal bacteria. In Proceedings of the European Federation of Animal Science Congress, Ghent, Belgium, 26–30 August 2019. [Google Scholar]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).