
Comparative Study of Two Immunisation Protocols in Goats Using Thiol-Sepharose Chromatography-Enriched Extracts from Adult Haemonchus contortus Worms
 , , ,
, , ,  ,
,  and
and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Parasites and Immunogens
2.2. Immunization Trials
- Group 1 (n = 6): goats immunized with a total of 625 μg TBSP in Freund complete and incomplete adjuvants and challenged with H. contortus; immunization in this group of animals was performed by weekly intramuscular injections of 50, 75, 100, 100, and 300 μg of TBSP extract over five consecutive weeks. Freund complete adjuvant was used for the first immunization (50 μg), and Freund incomplete adjuvant for the following immunizations. This protocol has previously been used in several experiments in goats demonstrating an immunoprotective effect against H. contortus. [19,20].
- Group 2 (n = 6): goats immunized with a total of 200 μg TBSP in Freund complete and incomplete adjuvants and challenged with H. contortus; immunization in this group of animals was performed by weekly intramuscular injections of 100 μg TBSP extract over two consecutive weeks. Freund complete adjuvant was used for the first immunization and Freund incomplete adjuvant for the second.
- Group 3 (n = 6): Freund adjuvant control. Animals in control group 3 underwent the same inoculation protocol as animals in group 1, but elution buffer was used instead of PBS-TSBP.
- Group 4 (n = 3): This group contained uninfected and unimmunised animals and served as a control for serological analysis.
2.3. Haematological and Parasitological Analysis
2.4. ELISA
2.5. Histology
2.6. Statistical Analysis
3. Results
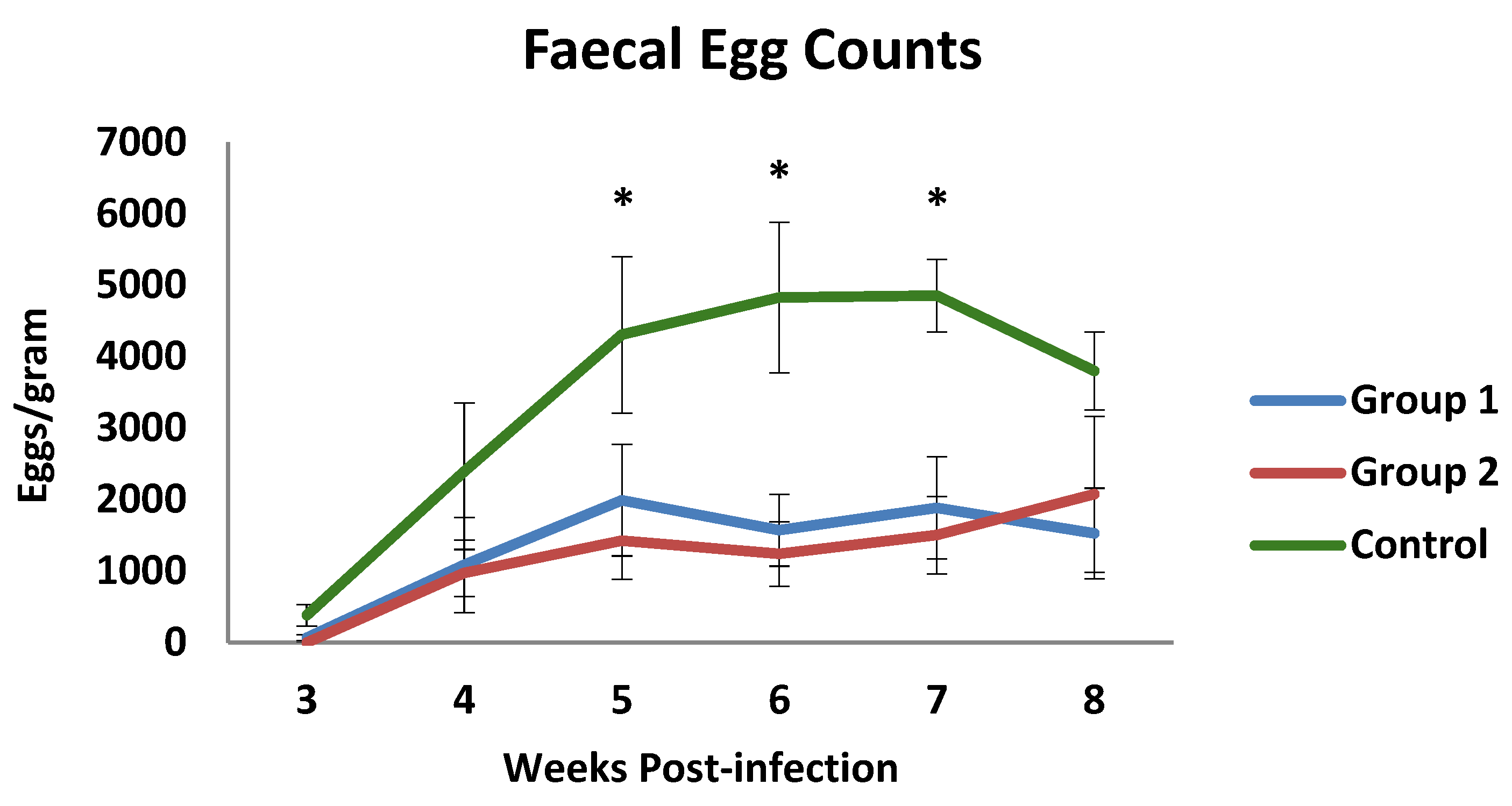
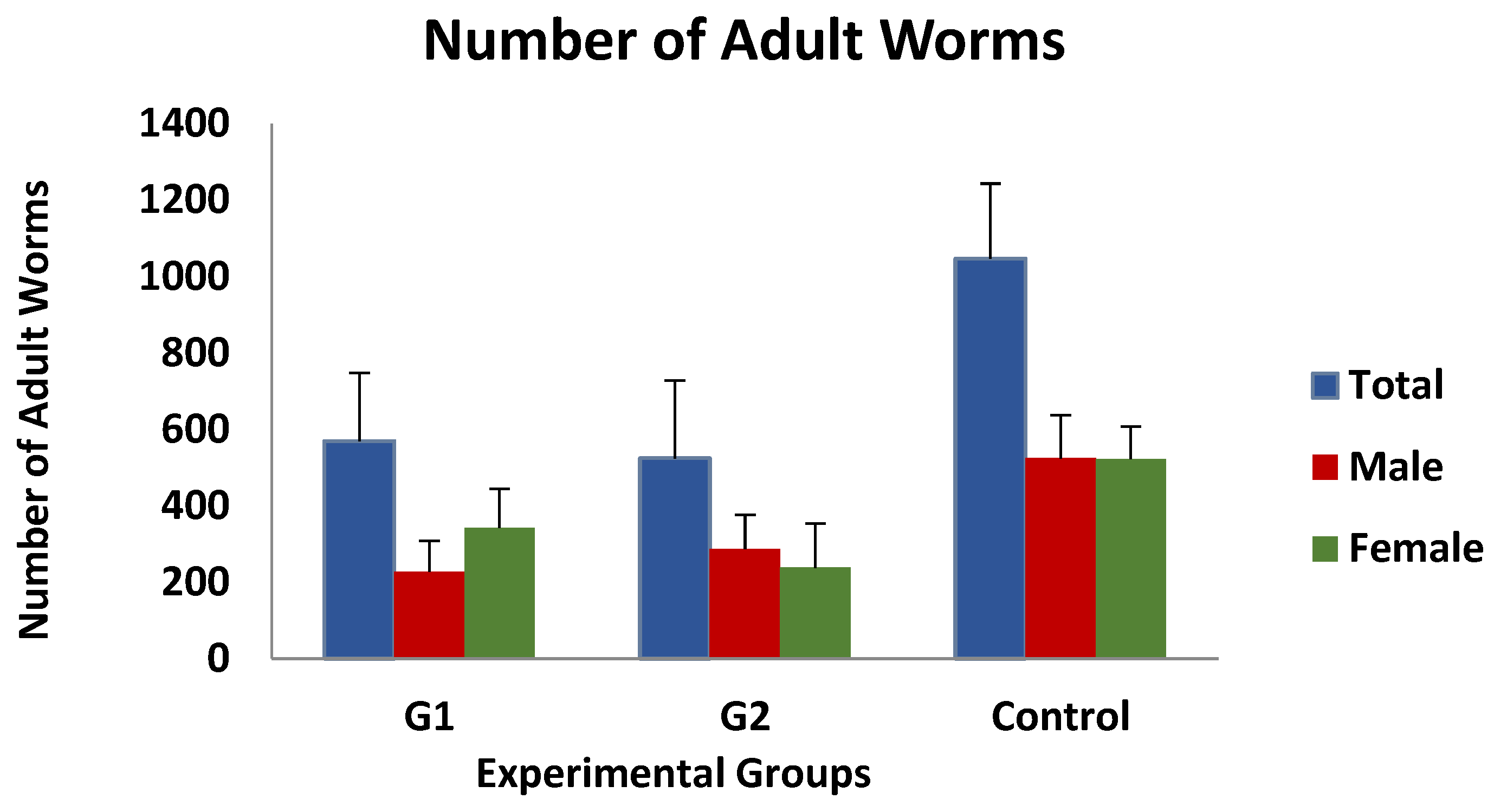
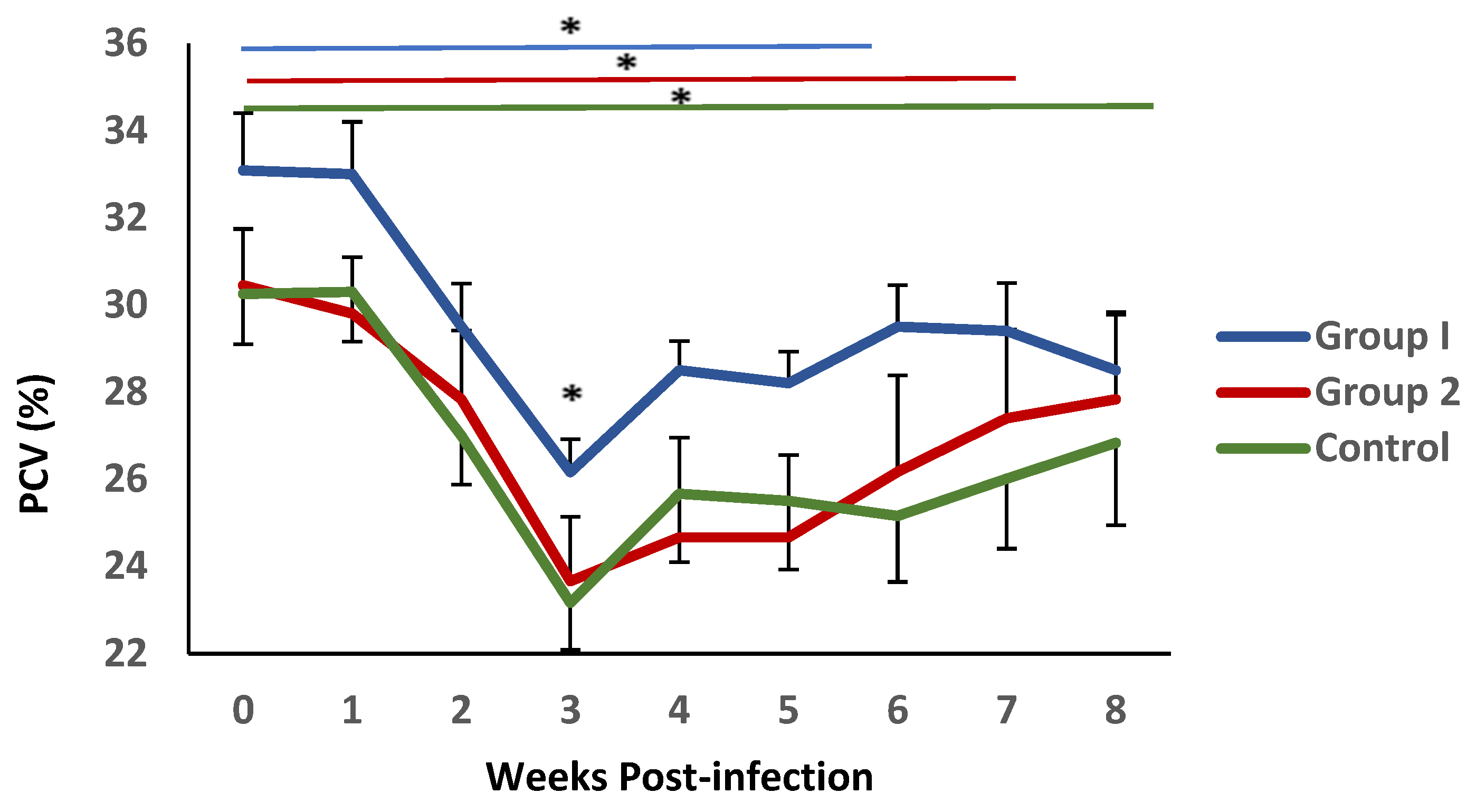
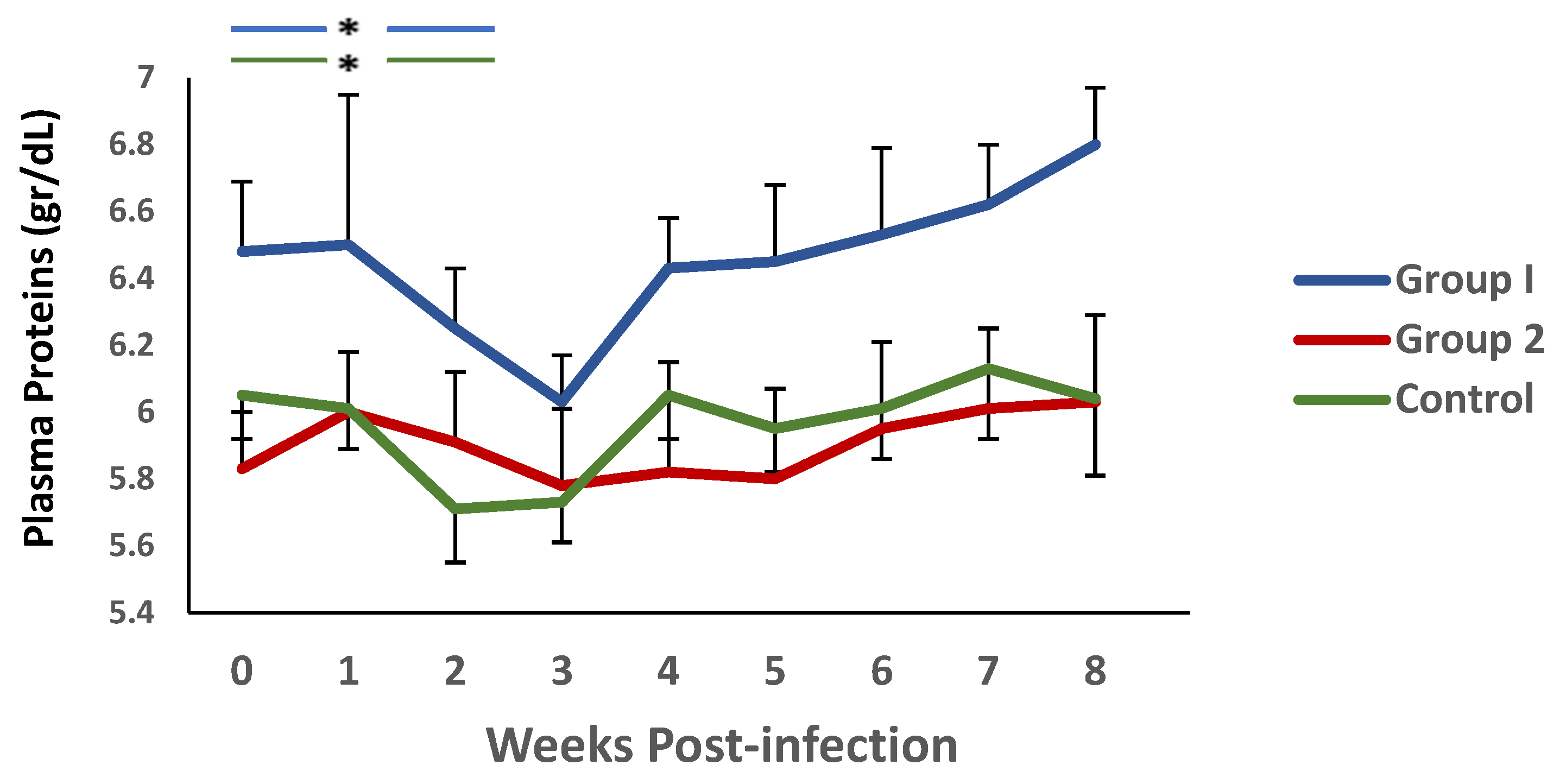
3.1. Parasitological and Haematological Results
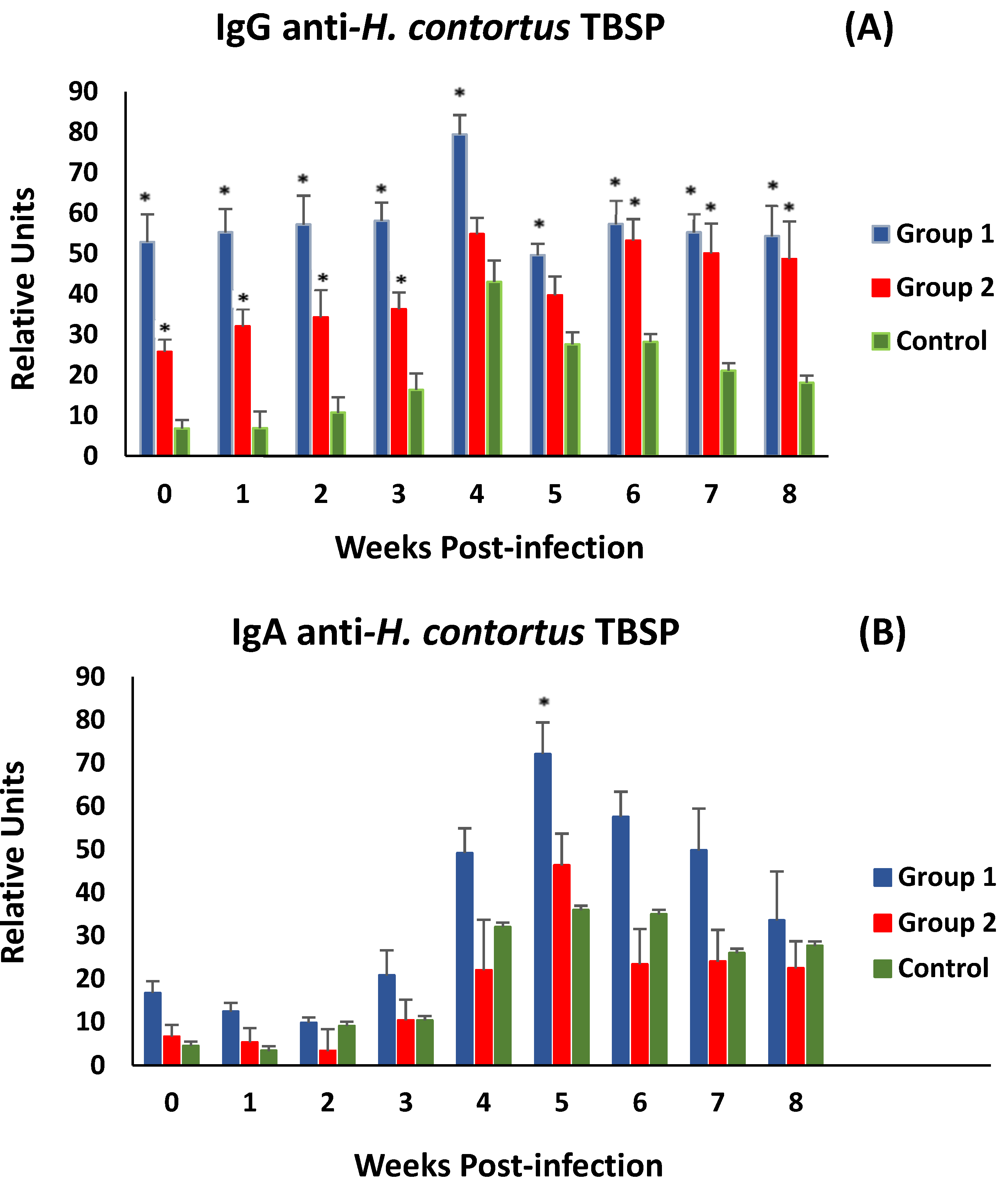
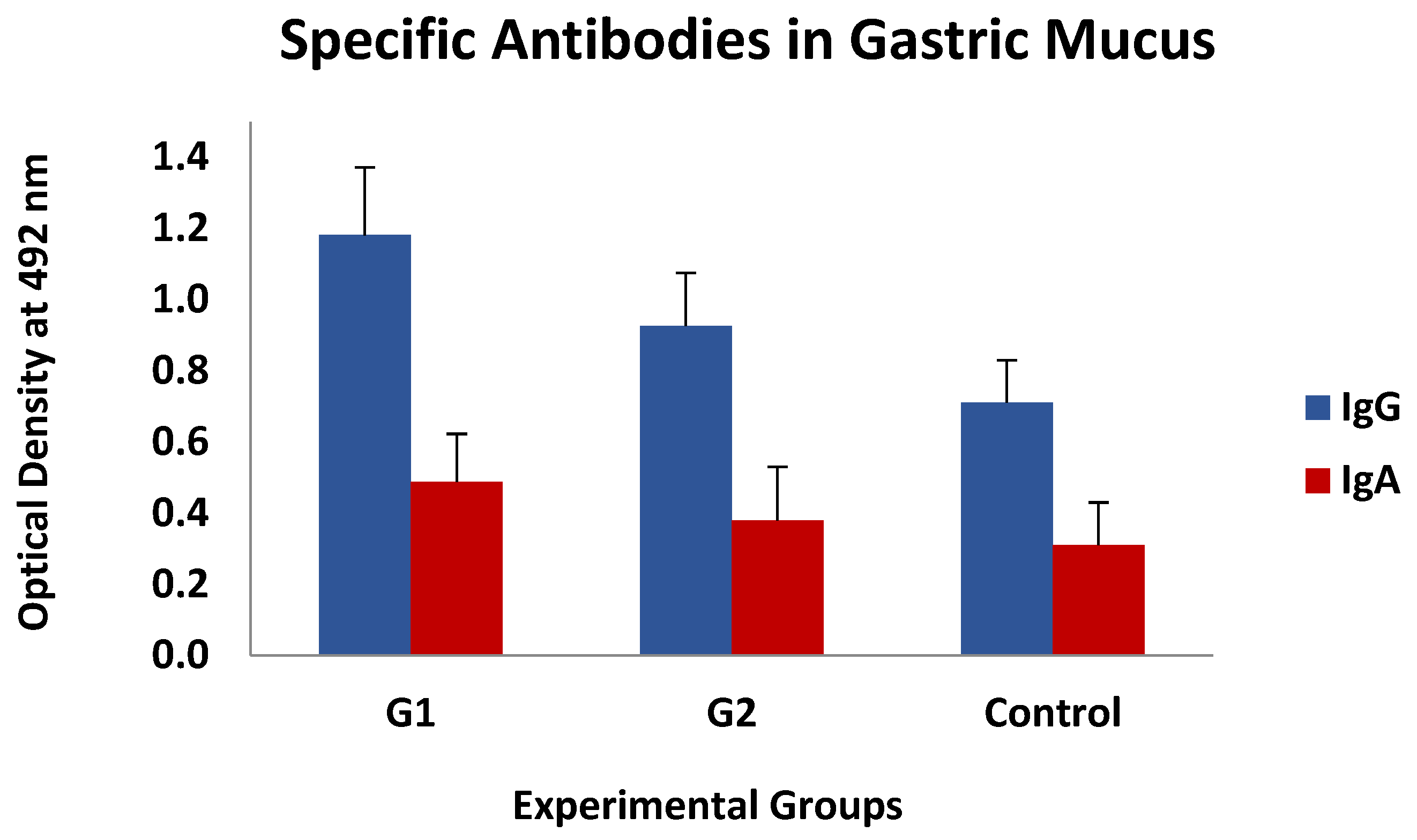
3.2. ELISA Tests
3.3. Histology
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Besier, R.B.; Kahn, L.P.; Sargison, N.D.; van Wyk, J.A. Diagnosis; Treatment and Management of Haemonchus contortus in Small Ruminants. Adv. Parasitol. 2016, 93, 181–238. [Google Scholar] [PubMed]
- Taylor, M.A.; Coop, R.L.; Wall, R.L. Parasites of Sheep and Goats (Parasites of the Digestive System). In Veterinary Parasitology, 4th ed.; Taylor, M.A., Coop, R.L., Wall, R.L., Eds.; John Wiley & Sons: Hoboken, NJ, USA, 2016; pp. 436–474. [Google Scholar]
- Britton, C.; Emery, D.L.; McNeilly, T.N.; Nisbet, A.J.; Stear, M.J. The potential for vaccines against scour worms of small ruminants. Int. J. Parasitol. 2020, 50, 533–553. [Google Scholar] [CrossRef]
- Zaheer, T.; Abbas, R.Z.; Imran, M.; Abbas, A.; Butt, A.; Aslam, S.; Ahmad, J. Vaccines against chicken coccidiosis with particular reference to previous decade: Progress; challenges; and opportunities. Parasitol. Res. 2022, 121, 2749–2763. [Google Scholar] [CrossRef]
- Claerebout, E.; Geldhof, P. Helminth vaccines in ruminants: From development to application. Vet. Clin. N. Am. Food Anim. Pract. 2020, 36, 159–171. [Google Scholar] [CrossRef]
- Meier, L.; Torgenson, P.R.; Hertzberg, H. Vaccination of goats against Haemonchus contortus with the gut membrane proteins H11/H-gal-GP. Vet. Parasitol. 2016, 229, 15–21. [Google Scholar] [CrossRef]
- Schallig, H.D.; Van Leeuwen, M.A.; Cornelissen, A.W. Protective immunity induced by vaccination with two Haemonchus contortus excretory secretory proteins in sheep. Parasite Immunol. 1997, 19, 447–453. [Google Scholar] [CrossRef]
- Matthews, J.B.; Geldhof, P.; Tzelos, T.; Claerebout, E. Progress in the development of subunit vaccines for gastrointestinal nematodes of ruminants. Parasite Immunol. 2016, 38, 744–753. [Google Scholar] [CrossRef] [PubMed]
- Bu, Y.; Jia, C.; Tian, X.; Aimulajiang, K.; Memon, M.A.; Yan, R.; Song, X.; Xu, L.; Li, X. Immunization of goats with recombinant protein 14-3-3 Isoform 2 (rHcftt-2) induced moderate protection against Haemonchus contortus challenge. Pathogens 2020, 9, 46. [Google Scholar] [CrossRef] [PubMed]
- Hasan, M.; Zhang, Z.; Fu, Y.; Wang, T.; Yan, R.; Li, X.; Ehsan, M.; Haseeb, M.; Lakho, S.A.; Haider, A.; et al. Immunization with PLGA nanoparticle-encapsulated Haemonchus contortus excretory-secretory protein rHcES-15 enhances protection in mice. Vaccines 2023, 11, 1794. [Google Scholar] [CrossRef]
- Piedrafita, D.; Preston, S.; Kemp, J.; de Veer, M.; Sherrard, J.; Kraska, T.; Elhay, M.; Meeusen, E. The effect of different adjuvants on immune parameters and protection following vaccination of sheep with a larval-specific antigen of the gastrointestinal nematode Haemonchus contortus. PLoS ONE 2013, 8, e78357. [Google Scholar] [CrossRef]
- González-Sánchez, M.E.; Cuquerella, M.; Alunda, J.M. Vaccination of lambs against Haemonchus contortus with the recombinant rHc23 antigen. Vet. Parasitol. 2018, 252, 8–13. [Google Scholar]
- Sun, W.; Song, X.; Yan, R.; Xu, L.; Li, X. Vaccination of goats with a glutathione peroxidase DNA vaccine induced partial protection against Haemonchus contortus infection. Vet. Parasitol. 2011, 182, 239–247. [Google Scholar] [CrossRef]
- Han, K.; Xu, L.; Yan, R.; Song, X.; Li, X. Vaccination of goats with glyceraldehyde-3-phosphate dehydrogenase DNA vaccine induced partial protection against Haemonchus contortus. Vet. Immunol. Immunopathol. 2012, 149, 177–195. [Google Scholar] [CrossRef]
- Kebeta, M.M.; Hine, B.C.; Walkden-Brown, S.W.; Kahn, L.P.; Doyle, E.K. Protective efficacy of Barbervax® in Merino weaner sheep trickle infected with five doses of Haemonchus contortus infective larvae. Vet. Parasitol. 2021, 292, 109386. [Google Scholar] [CrossRef]
- Smith, D. Barbervax®, a Vaccine for Haemonchus contortus Infection of Sheep: Attempts to Extend the Registration Claim to Include Goats; Final Report for Project B.AHE.0215; Meat & Livestock Australia: North Sydney, NSW, Australia, 2016. [Google Scholar]
- Adduci, I.; Sajovitz, F.; Hinney, B.; Lichtmannsperger, K.; Joachim, A.; Wittek, T.; Yan, S. Haemonchosis in sheep and goats, control strategies and development of vaccines against Haemonchus contortus. Animals 2022, 12, 2339. [Google Scholar] [CrossRef] [PubMed]
- Molina, J.M.; Martín, S.; Hernández, Y.I.; González, J.F.; Ferrer, O.; Ruiz, A. Immunoprotective effect of cysteine proteinase fractions from two Haemonchus contortus strains adapted to sheep and goats. Vet. Parasitol. 2012, 188, 53–59. [Google Scholar] [CrossRef] [PubMed]
- Molina, J.M.; Hernández, Y.I.; Martín, S.; Ferrer, O.; Rodríguez, F.; Ruiz, A. Immune response in goats vaccinated with thiol-binding proteins from Haemonchus contortus. Parasite Immunol. 2018, 40, e12569. [Google Scholar] [CrossRef] [PubMed]
- Molina, J.M.; Hernández, Y.I.; Ferrer, O.; Conde, M.M.; Rodríguez, F.; Ruiz, A. Immunization with thiol-binding proteins from Haemonchus contortus adult worms partially protects goats against infection during prepatency. Exp. Parasitol. 2023, 248, 108512. [Google Scholar] [CrossRef]
- Risa, D.; Mamo, G.; Waktole, H.; Haile, G.; Terefe, G. Goats are more susceptible to Haemonchus contortus infection than sheep under similar experimental settings. Sci. Rep. 2024, 14, 25379. [Google Scholar] [CrossRef]
- Ministery of Agriculture Fisheries and Food (MAFF). Manual of Veterinary Laboratory Diagnostic Techniques, Reference Book No. 418; Her Majesty’s Stationery Office: London, UK, 1989.
- Martín, S.; Molina, J.M.; Hernández, Y.I.; Ferrer, O.; Muñoz, M.C.; López, A.; Ortega, L.; Ruiz, A. Influence of immunoprotection 579 on genetic variability of cysteine proteinases from Haemonchus contortus adult worms. Int. J. Parasitol. 2015, 45, 831–840. [Google Scholar] [CrossRef]
- Camas-Pereyra, R.; Bautista-García, G.A.; Pérez-Anzúrez, G.; Duran-Cortes, Z.C.; Reyes-Guerrero, D.E.; Maza-Lopez, J.; Olmedo-Juárez, A.; López-Arellano, M.E. Excreted/secreted 15-kDa proteins and serine protease peptides from Haemonchus contortus act as immune-response enhancers in lambs. Pathogens 2024, 13, 604. [Google Scholar] [CrossRef] [PubMed]
- Kaplan, R.M.; Burke, J.M. Managing anthelmintic resistance in gastrointestinal nematodes of small ruminants. J. Anim. Sci. 2009, 87 (Suppl. S14), E47–E56. [Google Scholar]
- García Casas, C.; Moreno, A.; Caporte, J.; de la Haba, M.R. Characterization of the Canary racial goat groups with erythrocyte genetic markers. Small Rumin. Res. 1992, 7, 361–368. [Google Scholar] [CrossRef]
- Bassetto, C.C.; Almeida, F.A.; Newlands, G.F.J.; Smith, W.D.; Amarante, A.F.T. Repeated vaccination against Haemonchus contortus provides continuous protection to young grazing sheep. Vet. Parasitol. 2020, 287, 109273. [Google Scholar] [CrossRef]
- Albuquerque, A.C.A.; Bassetto, C.C.; Almeida, F.A.; Hildersley, K.A.; McNeilly, T.N.; Britton, C.; Amarante, A.F. Differences in immune responses to Haemonchus contortus infection in the susceptible Ile de France and the resistant Santa Ines sheep under different anthelmintic treatment regimens. Vet. Res. 2019, 50, 104. [Google Scholar] [CrossRef]
- Zhao, G.; Yan, R.; Muleke, C.I.; Sun, Y.; Xu, L.; Li, X. Vaccination of goats with DNA vaccines encoding H11 and IL-2 induces partial protection against Haemonchus contortus infection. Vet. J. 2021, 191, 94–100. [Google Scholar] [CrossRef]
- Pérez, J.; García, P.M.; Hernández, S.; Mozos, E.; Cámara, S.; Martínez-Moreno, A. Experimental haemonchosis in goats: Effects of single and multiple infections in the host response. Vet. Parasitol. 2003, 111, 333–342. [Google Scholar] [CrossRef]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Conde-Felipe, M.M.; Molina, J.A.; Ruiz, A.; Ferrer, O.; Del Rio, M.C.; Carmelo, E.; Hernández-Fernaud, J.R.; Rodríguez, F.; Molina, J.M. Comparative Study of Two Immunisation Protocols in Goats Using Thiol-Sepharose Chromatography-Enriched Extracts from Adult Haemonchus contortus Worms. Vaccines 2025, 13, 708. https://doi.org/10.3390/vaccines13070708
Conde-Felipe MM, Molina JA, Ruiz A, Ferrer O, Del Rio MC, Carmelo E, Hernández-Fernaud JR, Rodríguez F, Molina JM. Comparative Study of Two Immunisation Protocols in Goats Using Thiol-Sepharose Chromatography-Enriched Extracts from Adult Haemonchus contortus Worms. Vaccines. 2025; 13(7):708. https://doi.org/10.3390/vaccines13070708
Chicago/Turabian StyleConde-Felipe, Magnolia M., José Adrián Molina, Antonio Ruiz, Otilia Ferrer, Mª Cristina Del Rio, Emma Carmelo, Juan R. Hernández-Fernaud, Francisco Rodríguez, and José Manuel Molina. 2025. "Comparative Study of Two Immunisation Protocols in Goats Using Thiol-Sepharose Chromatography-Enriched Extracts from Adult Haemonchus contortus Worms" Vaccines 13, no. 7: 708. https://doi.org/10.3390/vaccines13070708
APA StyleConde-Felipe, M. M., Molina, J. A., Ruiz, A., Ferrer, O., Del Rio, M. C., Carmelo, E., Hernández-Fernaud, J. R., Rodríguez, F., & Molina, J. M. (2025). Comparative Study of Two Immunisation Protocols in Goats Using Thiol-Sepharose Chromatography-Enriched Extracts from Adult Haemonchus contortus Worms. Vaccines, 13(7), 708. https://doi.org/10.3390/vaccines13070708