
SARS-CoV-2 Variant-Specific Antibodies in Vaccinated Inflammatory Bowel Disease Patients

 ,
,  , , , and
, , , and
Abstract
1. Introduction
2. Materials and Methods
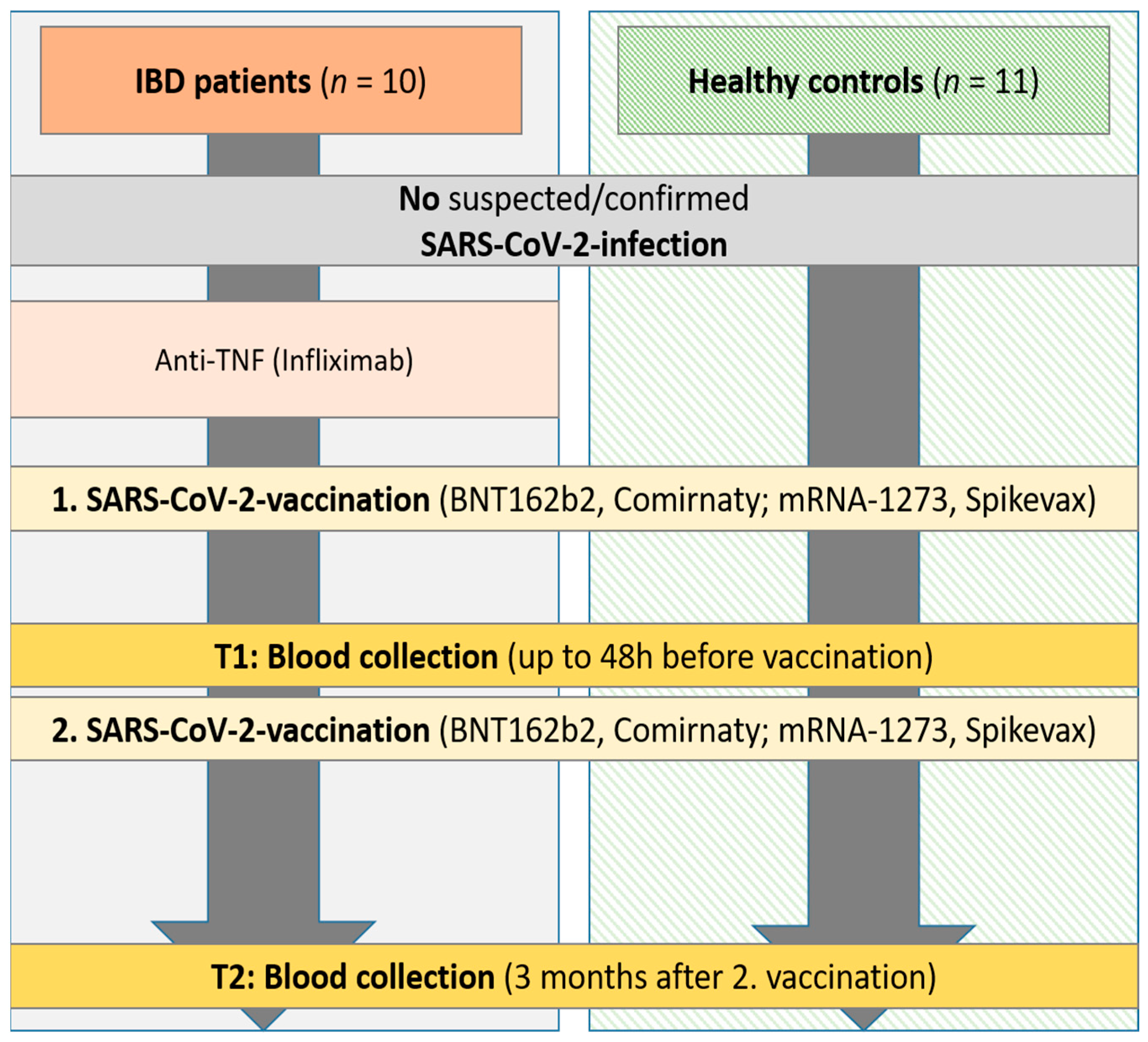
2.1. Study Subjects and Samples
2.2. Characterization of Sera with Commercial Assays
2.3. Characterization of the Neutralizing Capacity of Sera with In-House Assays
2.3.1. VSV-Pseudotyped Virus Neutralization Assay
2.3.2. Surrogate Virus Neutralization Assay
2.4. Statistical Analysis
3. Results
3.1. Cohort Characteristics
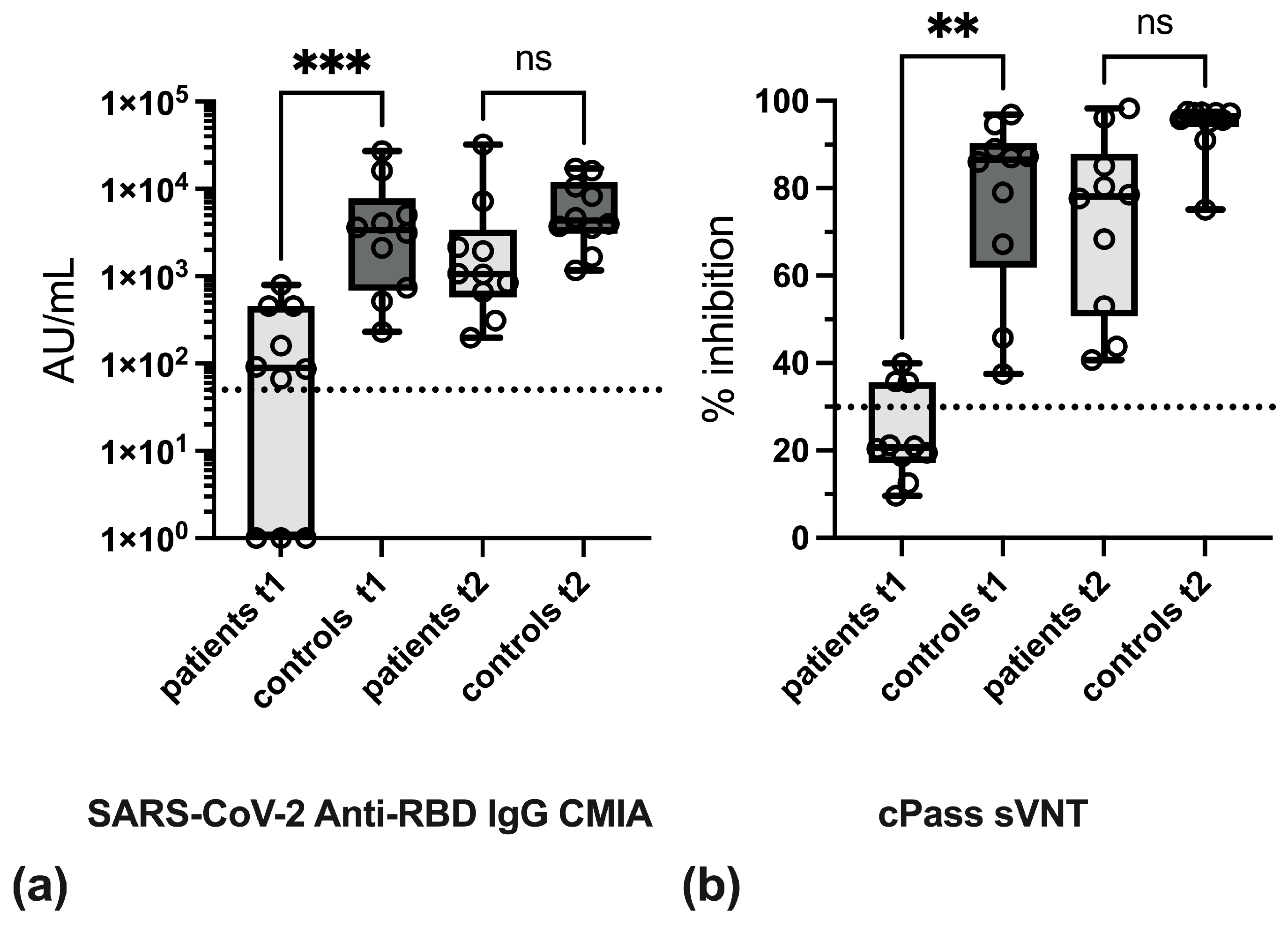
3.2. Effect of Anti-TNF Therapy on Antibody Levels
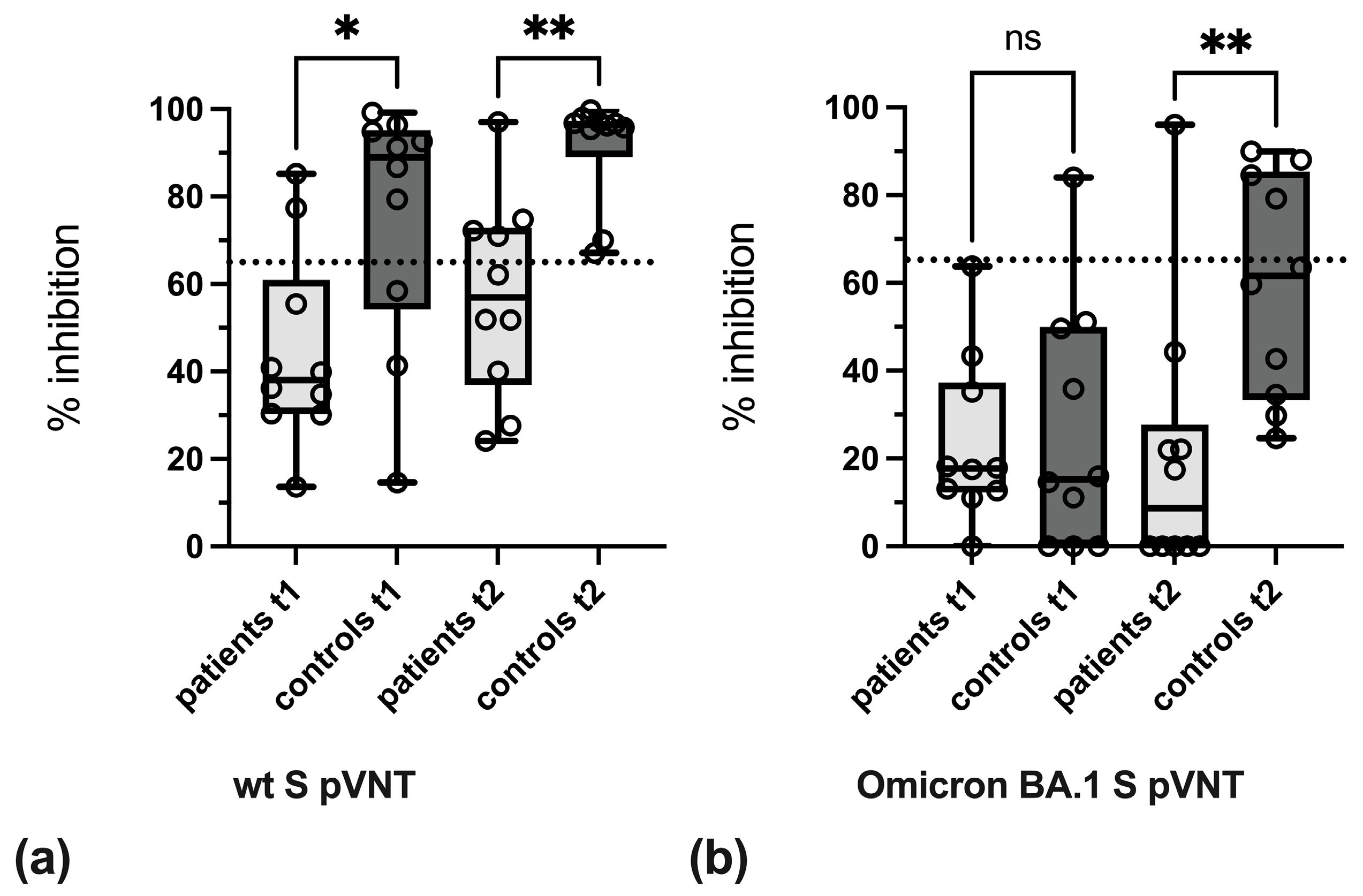
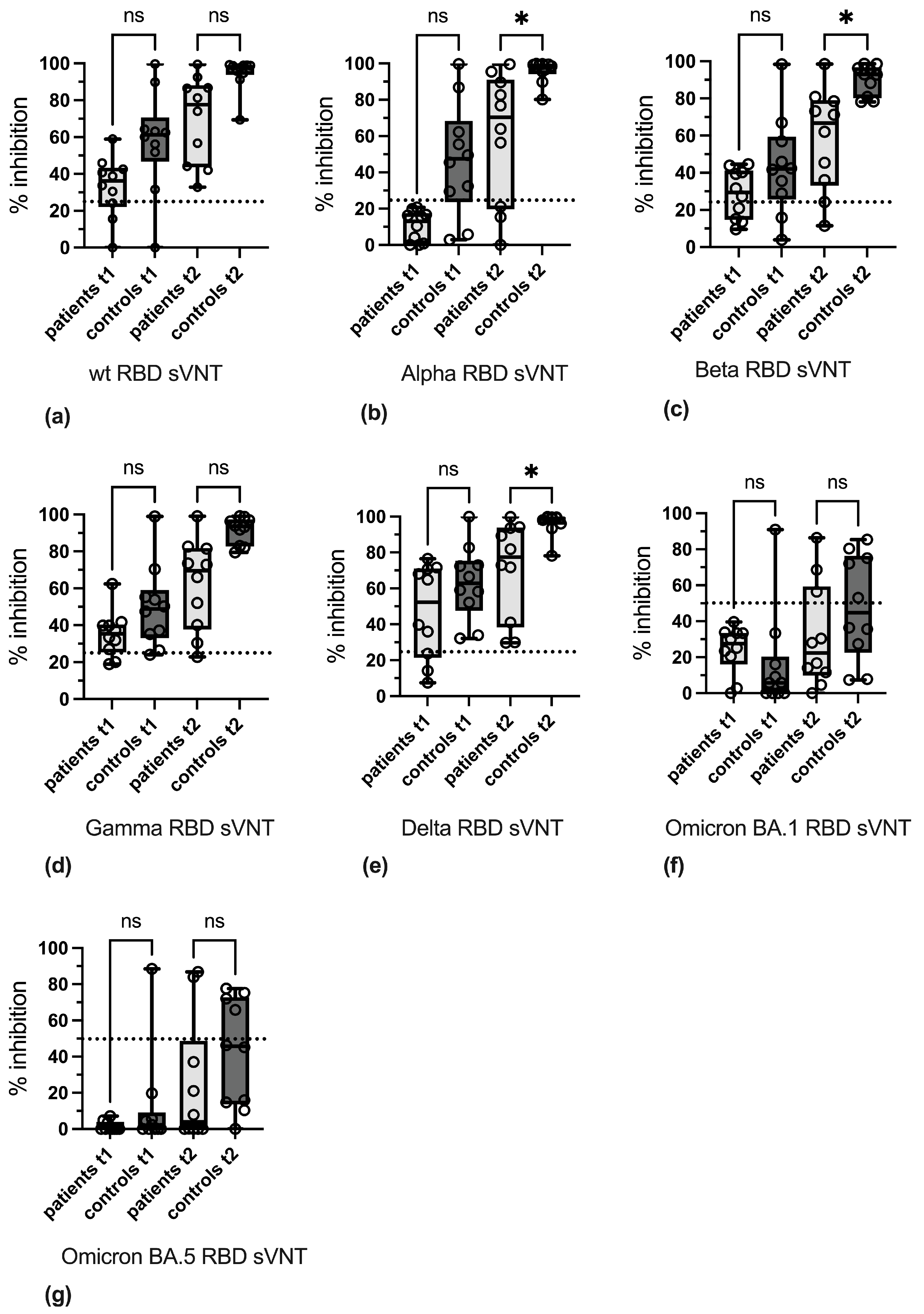
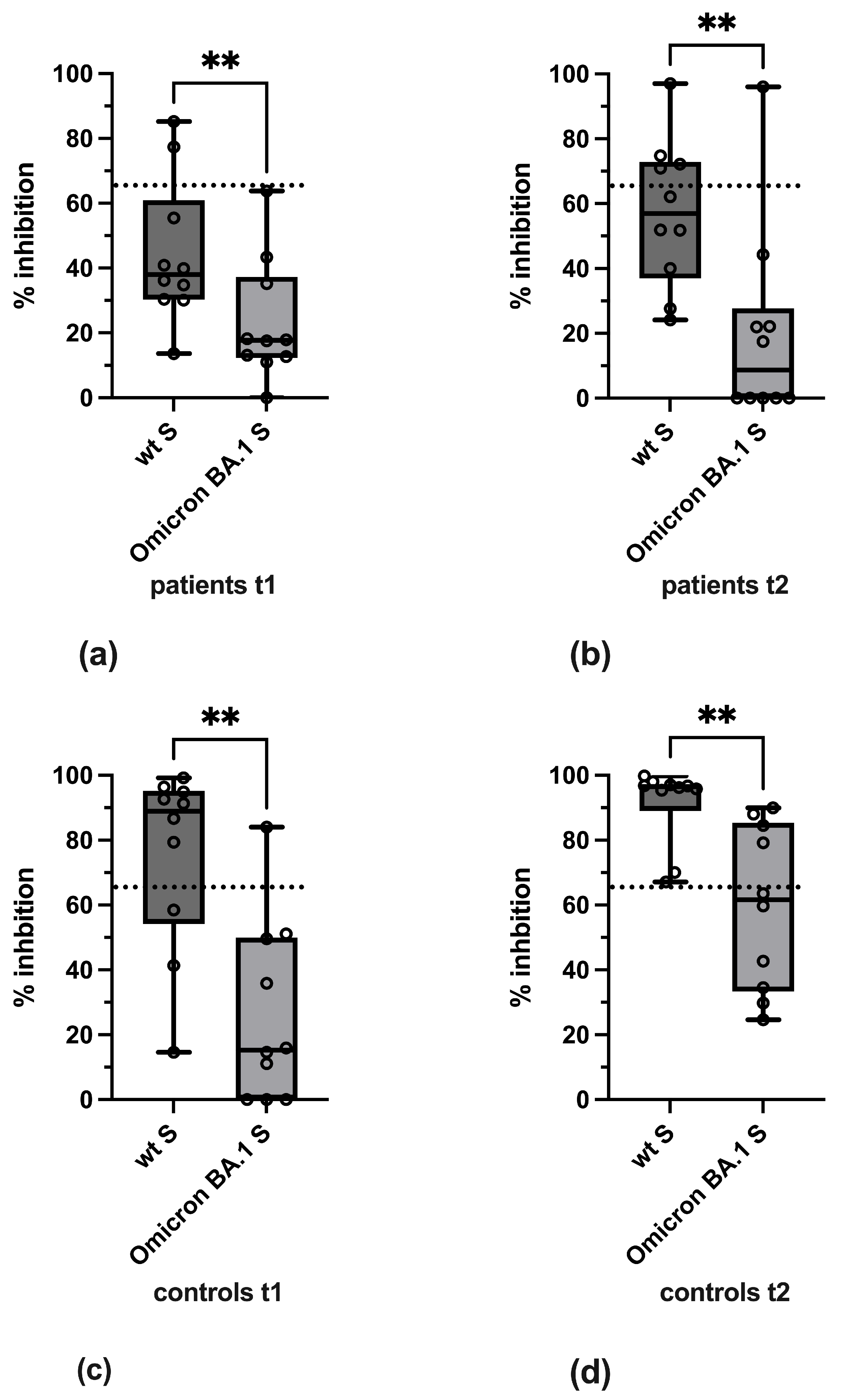
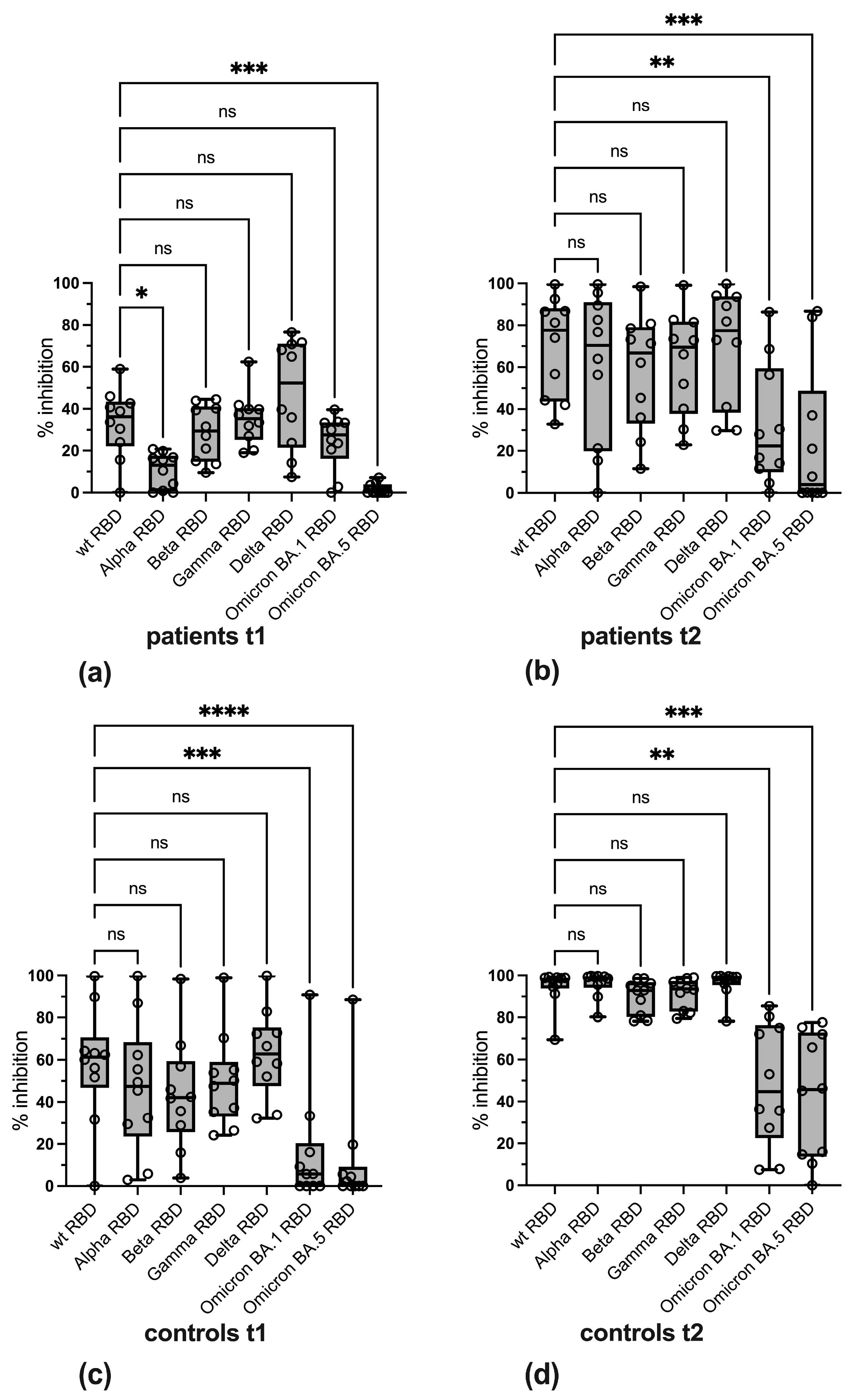
3.3. Variant-Specific Differences in Reactivity
3.4. Qualitative Evaluation of SARS-CoV-2-Specific Antibody Reactivity
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| Abbreviation | Definition |
| AU | arbitrary units |
| BAU | binding antibody units |
| CMIA | chemiluminescence microparticle assay |
| hACE2 | human angiotensin-converting enzyme 2 |
| NLuc | nanoluciferase |
| pVNT | VSV-pseudotyped virus neutralization assay |
| RBD | receptor-binding domain |
| ROC | receiver operating characteristic |
| SARS-CoV-2 | severe acute respiratory syndrome coronavirus 2 |
| sVNT | surrogate virus neutralization test |
| TNF | tumor necrosis factor |
References
- Wu, F.; Zhao, S.; Yu, B.; Chen, Y.M.; Wang, W.; Song, Z.G.; Hu, Y.; Tao, Z.W.; Tian, J.H.; Pei, Y.Y.; et al. A new coronavirus associated with human respiratory disease in China. Nature 2020, 579, 265–269. [Google Scholar] [CrossRef] [PubMed]
- Coronaviridae Study Group of the International Committee on Taxonomy of Viruses. The species Severe acute respiratory syndrome-related coronavirus: Classifying 2019-nCoV and naming it SARS-CoV-2. Nat. Microbiol. 2020, 5, 536–544. [Google Scholar] [CrossRef]
- Vollenberg, R.; Matern, P.; Nowacki, T.; Fuhrmann, V.; Padberg, J.S.; Ochs, K.; Schutte-Nutgen, K.; Strauss, M.; Schmidt, H.; Tepasse, P.R. Prone Position in Mechanically Ventilated COVID-19 Patients: A Multicenter Study. J. Clin. Med. 2021, 10, 1046. [Google Scholar] [CrossRef] [PubMed]
- Torres Acosta, M.A.; Singer, B.D. Pathogenesis of COVID-19-induced ARDS: Implications for an ageing population. Eur. Respir. J. 2020, 56, 2002049. [Google Scholar] [CrossRef]
- Zhou, F.; Yu, T.; Du, R.; Fan, G.; Liu, Y.; Liu, Z.; Xiang, J.; Wang, Y.; Song, B.; Gu, X.; et al. Clinical course and risk factors for mortality of adult inpatients with COVID-19 in Wuhan, China: A retrospective cohort study. Lancet 2020, 395, 1054–1062. [Google Scholar] [CrossRef]
- Huang, C.; Wang, Y.; Li, X.; Ren, L.; Zhao, J.; Hu, Y.; Zhang, L.; Fan, G.; Xu, J.; Gu, X.; et al. Clinical features of patients infected with 2019 novel coronavirus in Wuhan, China. Lancet 2020, 395, 497–506. [Google Scholar] [CrossRef] [PubMed]
- Chen, N.; Zhou, M.; Dong, X.; Qu, J.; Gong, F.; Han, Y.; Qiu, Y.; Wang, J.; Liu, Y.; Wei, Y.; et al. Epidemiological and clinical characteristics of 99 cases of 2019 novel coronavirus pneumonia in Wuhan, China: A descriptive study. Lancet 2020, 395, 507–513. [Google Scholar] [CrossRef]
- Wang, D.; Hu, B.; Hu, C.; Zhu, F.; Liu, X.; Zhang, J.; Wang, B.; Xiang, H.; Cheng, Z.; Xiong, Y.; et al. Clinical Characteristics of 138 Hospitalized Patients with 2019 Novel Coronavirus-Infected Pneumonia in Wuhan, China. JAMA 2020, 323, 1061–1069. [Google Scholar] [CrossRef]
- Kessel, C.; Vollenberg, R.; Masjosthusmann, K.; Hinze, C.; Wittkowski, H.; Debaugnies, F.; Nagant, C.; Corazza, F.; Vely, F.; Kaplanski, G.; et al. Discrimination of COVID-19 From Inflammation-Induced Cytokine Storm Syndromes Using Disease-Related Blood Biomarkers. Arthritis Rheumatol. 2021, 73, 1791–1799. [Google Scholar] [CrossRef]
- Jose, R.J.; Manuel, A. COVID-19 cytokine storm: The interplay between inflammation and coagulation. Lancet Respir. Med. 2020, 8, e46–e47. [Google Scholar] [CrossRef]
- Yang, L.; Xie, X.; Tu, Z.; Fu, J.; Xu, D.; Zhou, Y. The signal pathways and treatment of cytokine storm in COVID-19. Signal Transduct. Target. Ther. 2021, 6, 255. [Google Scholar] [CrossRef]
- Nasserie, T.; Hittle, M.; Goodman, S.N. Assessment of the Frequency and Variety of Persistent Symptoms Among Patients with COVID-19: A Systematic Review. JAMA Netw. Open 2021, 4, e2111417. [Google Scholar] [CrossRef]
- Deer, R.R.; Rock, M.A.; Vasilevsky, N.; Carmody, L.; Rando, H.; Anzalone, A.J.; Basson, M.D.; Bennett, T.D.; Bergquist, T.; Boudreau, E.A.; et al. Characterizing Long COVID: Deep Phenotype of a Complex Condition. EBioMedicine 2021, 74, 103722. [Google Scholar] [CrossRef]
- Varghese, J.; Sandmann, S.; Ochs, K.; Schrempf, I.M.; Frommel, C.; Dugas, M.; Schmidt, H.H.; Vollenberg, R.; Tepasse, P.R. Persistent symptoms and lab abnormalities in patients who recovered from COVID-19. Sci. Rep. 2021, 11, 12775. [Google Scholar] [CrossRef] [PubMed]
- Davis, H.E.; McCorkell, L.; Vogel, J.M.; Topol, E.J. Long COVID: Major findings, mechanisms and recommendations. Nat. Rev. Microbiol. 2023, 21, 133–146. [Google Scholar] [CrossRef] [PubMed]
- Ungaro, R.; Mehandru, S.; Allen, P.B.; Peyrin-Biroulet, L.; Colombel, J.F. Ulcerative colitis. Lancet 2017, 389, 1756–1770. [Google Scholar] [CrossRef]
- Torres, J.; Mehandru, S.; Colombel, J.F.; Peyrin-Biroulet, L. Crohn’s disease. Lancet 2017, 389, 1741–1755. [Google Scholar] [CrossRef] [PubMed]
- Ali, T.; Kaitha, S.; Mahmood, S.; Ftesi, A.; Stone, J.; Bronze, M.S. Clinical use of anti-TNF therapy and increased risk of infections. Drug Health Patient Saf. 2013, 5, 79–99. [Google Scholar] [CrossRef]
- Vollenberg, R.; Lorentzen, E.U.; Kuhn, J.; Nowacki, T.M.; Meier, J.A.; Trebicka, J.; Tepasse, P.R. Humoral Immunity in Immunosuppressed IBD Patients after the Third SARS-CoV-2 Vaccination: A Comparison with Healthy Control Subjects. Vaccines 2023, 11, 1411. [Google Scholar] [CrossRef]
- Vollenberg, R.; Tepasse, P.R.; Kuhn, J.E.; Hennies, M.; Strauss, M.; Rennebaum, F.; Schomacher, T.; Boeckel, G.; Lorentzen, E.; Bokemeyer, A.; et al. Humoral Immune Response in IBD Patients Three and Six Months after Vaccination with the SARS-CoV-2 mRNA Vaccines mRNA-1273 and BNT162b2. Biomedicines 2022, 10, 171. [Google Scholar] [CrossRef]
- Vollenberg, R.; Tepasse, P.R.; Lorentzen, E.; Nowacki, T.M. Impaired Humoral Immunity with Concomitant Preserved T Cell Reactivity in IBD Patients on Treatment with Infliximab 6 Month after Vaccination with the SARS-CoV-2 mRNA Vaccine BNT162b2: A Pilot Study. J. Pers. Med. 2022, 12, 694. [Google Scholar] [CrossRef] [PubMed]
- Kennedy, N.A.; Goodhand, J.R.; Bewshea, C.; Nice, R.; Chee, D.; Lin, S.; Chanchlani, N.; Butterworth, J.; Cooney, R.; Croft, N.M.; et al. Anti-SARS-CoV-2 antibody responses are attenuated in patients with IBD treated with infliximab. Gut 2021, 70, 865–875. [Google Scholar] [CrossRef] [PubMed]
- Kennedy, N.A.; Lin, S.; Goodhand, J.R.; Chanchlani, N.; Hamilton, B.; Bewshea, C.; Nice, R.; Chee, D.; Cummings, J.F.; Fraser, A.; et al. Infliximab is associated with attenuated immunogenicity to BNT162b2 and ChAdOx1 nCoV-19 SARS-CoV-2 vaccines in patients with IBD. Gut 2021, 70, 1884–1893. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Le, K.; Zhou, X.; Alexander, J.L.; Lin, S.; Bewshea, C.; Chanchlani, N.; Nice, R.; McDonald, T.J.; Lamb, C.A.; et al. Neutralising antibody potency against SARS-CoV-2 wild-type and omicron BA.1 and BA.4/5 variants in patients with inflammatory bowel disease treated with infliximab and vedolizumab after three doses of COVID-19 vaccine (CLARITY IBD): An analysis of a prospective multicentre cohort study. Lancet Gastroenterol. Hepatol. 2023, 8, 145–156. [Google Scholar] [CrossRef]
- Lin, S.; Kennedy, N.A.; Saifuddin, A.; Sandoval, D.M.; Reynolds, C.J.; Seoane, R.C.; Kottoor, S.H.; Pieper, F.P.; Lin, K.M.; Butler, D.K.; et al. Antibody decay, T cell immunity and breakthrough infections following two SARS-CoV-2 vaccine doses in inflammatory bowel disease patients treated with infliximab and vedolizumab. Nat. Commun. 2022, 13, 1379. [Google Scholar] [CrossRef]
- Caldera, F.; Rolak, S.; Farraye, F.A.; Necela, B.M.; Cogen, D.; Zona, E.E.; Schell, T.L.; Ramirez, O.R.; Almasry, M.; Chun, K.; et al. Higher and Sustained Cell-Mediated Immune Responses After 3 Doses of mRNA COVID-19 Vaccine in Patients with Inflammatory Bowel Disease on Anti-Tumor Necrosis Factor Therapy. Clin. Transl. Gastroenterol. 2024, 15, e00688. [Google Scholar] [CrossRef]
- Martinez-Dominguez, S.J.; Garcia-Mateo, S.; Sainz-Arnal, P.; Martinez-Garcia, J.; Gallego-Llera, B.; Lozano-Limones, M.J.; Hidalgo, S.; Gargallo-Puyuelo, C.J.; Latre-Santos, M.; Nocito-Colon, M.M.L.; et al. Unravelling the cellular response to the SARS-COV-2 vaccine in inflammatory bowel disease patients on biologic drugs. Sci. Rep. 2023, 13, 23061. [Google Scholar] [CrossRef]
- Schell, T.L.; Caldera, F. A Practical Update on COVID-19 and Inflammatory Bowel Disease: COVID-19 Disease Risk and Vaccine Safety and Efficacy. Gastroenterol. Hepatol. 2024, 20, 88–97. [Google Scholar]
- Harvey, W.T.; Carabelli, A.M.; Jackson, B.; Gupta, R.K.; Thomson, E.C.; Harrison, E.M.; Ludden, C.; Reeve, R.; Rambaut, A.; Consortium, C.-G.U.; et al. SARS-CoV-2 variants, spike mutations and immune escape. Nat. Rev. Microbiol. 2021, 19, 409–424. [Google Scholar] [CrossRef]
- Viana, R.; Moyo, S.; Amoako, D.G.; Tegally, H.; Scheepers, C.; Althaus, C.L.; Anyaneji, U.J.; Bester, P.A.; Boni, M.F.; Chand, M.; et al. Rapid epidemic expansion of the SARS-CoV-2 Omicron variant in southern Africa. Nature 2022, 603, 679–686. [Google Scholar] [CrossRef]
- Pulliam, J.R.C.; van Schalkwyk, C.; Govender, N.; von Gottberg, A.; Cohen, C.; Groome, M.J.; Dushoff, J.; Mlisana, K.; Moultrie, H. Increased risk of SARS-CoV-2 reinfection associated with emergence of Omicron in South Africa. Science 2022, 376, eabn4947. [Google Scholar] [CrossRef] [PubMed]
- Christensen, P.A.; Olsen, R.J.; Long, S.W.; Snehal, R.; Davis, J.J.; Ojeda Saavedra, M.; Reppond, K.; Shyer, M.N.; Cambric, J.; Gadd, R.; et al. Signals of Significantly Increased Vaccine Breakthrough, Decreased Hospitalization Rates, and Less Severe Disease in Patients with Coronavirus Disease 2019 Caused by the Omicron Variant of Severe Acute Respiratory Syndrome Coronavirus 2 in Houston, Texas. Am. J. Pathol. 2022, 192, 642–652. [Google Scholar] [CrossRef] [PubMed]
- Martin-Arranz, M.D.; Garcia-Ramirez, L.; Hernandez-Perez, M.; Montero Vega, D.; Martin-Arranz, E.; Sanchez-Azofra, M.; Poza Cordon, J.; Rueda Garcia, J.L.; Noci Belda, J.; Verges Martinez-Meco, T.; et al. Seroprevalence of ANTI-SARS-CoV-2 antibodies in patients with inflammatory bowel disease. Sci. Rep. 2023, 13, 7044. [Google Scholar] [CrossRef]
- Schoefbaenker, M.; Neddermeyer, R.; Guenther, T.; Mueller, M.M.; Romberg, M.L.; Classen, N.; Hennies, M.T.; Hrincius, E.R.; Ludwig, S.; Kuehn, J.E.; et al. Surrogate Virus Neutralisation Test Based on Nanoluciferase-Tagged Antigens to Quantify Inhibitory Antibodies against SARS-CoV-2 and Characterise Omicron-Specific Reactivity in a Vaccination Cohort. Vaccines 2023, 11, 1832. [Google Scholar] [CrossRef]
- Springer, D.N.; Perkmann, T.; Jani, C.M.; Mucher, P.; Pruger, K.; Marculescu, R.; Reuberger, E.; Camp, J.V.; Graninger, M.; Borsodi, C.; et al. Reduced Sensitivity of Commercial Spike-Specific Antibody Assays after Primary Infection with the SARS-CoV-2 Omicron Variant. Microbiol. Spectr. 2022, 10, e0212922. [Google Scholar] [CrossRef]
- Tan, C.W.; Chia, W.N.; Qin, X.; Liu, P.; Chen, M.I.; Tiu, C.; Hu, Z.; Chen, V.C.; Young, B.E.; Sia, W.R.; et al. A SARS-CoV-2 surrogate virus neutralization test based on antibody-mediated blockage of ACE2-spike protein-protein interaction. Nat. Biotechnol. 2020, 38, 1073–1078. [Google Scholar] [CrossRef]
- von Rhein, C.; Scholz, T.; Henss, L.; Kronstein-Wiedemann, R.; Schwarz, T.; Rodionov, R.N.; Corman, V.M.; Tonn, T.; Schnierle, B.S. Comparison of potency assays to assess SARS-CoV-2 neutralizing antibody capacity in COVID-19 convalescent plasma. J. Virol. Methods 2021, 288, 114031. [Google Scholar] [CrossRef]
- Graninger, M.; Jani, C.M.; Reuberger, E.; Pruger, K.; Gaspar, P.; Springer, D.N.; Borsodi, C.; Weidner, L.; Rabady, S.; Puchhammer-Stockl, E.; et al. Comprehensive Comparison of Seven SARS-CoV-2-Specific Surrogate Virus Neutralization and Anti-Spike IgG Antibody Assays Using a Live-Virus Neutralization Assay as a Reference. Microbiol. Spectr. 2023, 11, e0231422. [Google Scholar] [CrossRef] [PubMed]
- Mohamed, F.F.; Anhlan, D.; Schofbanker, M.; Schreiber, A.; Classen, N.; Hensel, A.; Hempel, G.; Scholz, W.; Kuhn, J.; Hrincius, E.R.; et al. Hypericum perforatum and Its Ingredients Hypericin and Pseudohypericin Demonstrate an Antiviral Activity against SARS-CoV-2. Pharmaceuticals 2022, 15, 530. [Google Scholar] [CrossRef]
- Schoefbaenker, M.; Gunther, T.; Lorentzen, E.U.; Romberg, M.L.; Hennies, M.T.; Neddermeyer, R.; Muller, M.M.; Mellmann, A.; Bojarzyn, C.R.; Lenz, G.; et al. Characterisation of the antibody-mediated selective pressure driving intra-host evolution of SARS-CoV-2 in prolonged infection. PLoS Pathog. 2024, 20, e1012624. [Google Scholar] [CrossRef]
- Glockner, S.; Hornung, F.; Baier, M.; Weis, S.; Pletz, M.W.; Deinhardt-Emmer, S.; Loffler, B.; The CoNAN Study Group. Robust Neutralizing Antibody Levels Detected after Either SARS-CoV-2 Vaccination or One Year after Infection. Viruses 2021, 13, 2003. [Google Scholar] [CrossRef] [PubMed]
- Rockstroh, A.; Wolf, J.; Fertey, J.; Kalbitz, S.; Schroth, S.; Lubbert, C.; Ulbert, S.; Borte, S. Correlation of humoral immune responses to different SARS-CoV-2 antigens with virus neutralizing antibodies and symptomatic severity in a German COVID-19 cohort. Emerg. Microbes Infect. 2021, 10, 774–781. [Google Scholar] [CrossRef] [PubMed]
- Dolscheid-Pommerich, R.; Bartok, E.; Renn, M.; Kummerer, B.M.; Schulte, B.; Schmithausen, R.M.; Stoffel-Wagner, B.; Streeck, H.; Saschenbrecker, S.; Steinhagen, K.; et al. Correlation between a quantitative anti-SARS-CoV-2 IgG ELISA and neutralization activity. J. Med. Virol. 2022, 94, 388–392. [Google Scholar] [CrossRef]
- Grunau, B.; Prusinkiewicz, M.; Asamoah-Boaheng, M.; Golding, L.; Lavoie, P.M.; Petric, M.; Levett, P.N.; Haig, S.; Barakauskas, V.; Karim, M.E.; et al. Correlation of SARS-CoV-2 Viral Neutralizing Antibody Titers with Anti-Spike Antibodies and ACE-2 Inhibition among Vaccinated Individuals. Microbiol. Spectr. 2022, 10, e0131522. [Google Scholar] [CrossRef]
- Earle, K.A.; Ambrosino, D.M.; Fiore-Gartland, A.; Goldblatt, D.; Gilbert, P.B.; Siber, G.R.; Dull, P.; Plotkin, S.A. Evidence for antibody as a protective correlate for COVID-19 vaccines. Vaccine 2021, 39, 4423–4428. [Google Scholar] [CrossRef] [PubMed]
- Krammer, F. A correlate of protection for SARS-CoV-2 vaccines is urgently needed. Nat. Med. 2021, 27, 1147–1148. [Google Scholar] [CrossRef]
- Martin Perez, C.; Aguilar, R.; Jimenez, A.; Salmeron, G.; Canyelles, M.; Rubio, R.; Vidal, M.; Cuamba, I.; Barrios, D.; Diaz, N.; et al. Correlates of protection and determinants of SARS-CoV-2 breakthrough infections 1 year after third dose vaccination. BMC Med. 2024, 22, 103. [Google Scholar] [CrossRef]
- Bubonja-Sonje, M.; Baticic, L.; Abram, M.; Cekinovic Grbesa, D. Diagnostic accuracy of three SARS-CoV2 antibody detection assays, neutralizing effect and longevity of serum antibodies. J. Virol. Methods 2021, 293, 114173. [Google Scholar] [CrossRef]
- Slotkin, R.; Kyriakides, T.C.; Kundu, A.; Stack, G.; Sutton, R.E.; Gupta, S. Correlation of a commercial platform’s results with post-vaccination SARS-CoV-2 neutralizing antibody response and clinical host factors. PLoS ONE 2023, 18, e0289713. [Google Scholar] [CrossRef]
- Infantino, M.; Pieri, M.; Nuccetelli, M.; Grossi, V.; Lari, B.; Tomassetti, F.; Calugi, G.; Pancani, S.; Benucci, M.; Casprini, P.; et al. The WHO International Standard for COVID-19 serological tests: Towards harmonization of anti-spike assays. Int. Immunopharmacol. 2021, 100, 108095. [Google Scholar] [CrossRef]
- Zhu, F.; Althaus, T.; Tan, C.W.; Costantini, A.; Chia, W.N.; Van Vinh Chau, N.; Van Tan, L.; Mattiuzzo, G.; Rose, N.J.; Voiglio, E.; et al. WHO international standard for SARS-CoV-2 antibodies to determine markers of protection. Lancet Microbe 2022, 3, e81–e82. [Google Scholar] [CrossRef] [PubMed]
- Muecksch, F.; Wise, H.; Templeton, K.; Batchelor, B.; Squires, M.; McCance, K.; Jarvis, L.; Malloy, K.; Furrie, E.; Richardson, C.; et al. Longitudinal variation in SARS-CoV-2 antibody levels and emergence of viral variants: A serological analysis. Lancet Microbe 2022, 3, e493–e502. [Google Scholar] [CrossRef] [PubMed]
- Giavarina, D.; Carta, M. Improvements and limits of anti SARS-CoV-2 antibodies assays by WHO (NIBSC 20/136) standardization. Diagnosis 2021, 9, 274–279. [Google Scholar] [CrossRef]
- Saker, K.; Escuret, V.; Pitiot, V.; Massardier-Pilonchery, A.; Paul, S.; Mokdad, B.; Langlois-Jacques, C.; Rabilloud, M.; Goncalves, D.; Fabien, N.; et al. Evaluation of Commercial Anti-SARS-CoV-2 Antibody Assays and Comparison of Standardized Titers in Vaccinated Health Care Workers. J. Clin. Microbiol. 2022, 60, e0174621. [Google Scholar] [CrossRef]
- Perkmann, T.; Perkmann-Nagele, N.; Koller, T.; Mucher, P.; Radakovics, A.; Marculescu, R.; Wolzt, M.; Wagner, O.F.; Binder, C.J.; Haslacher, H. Anti-Spike Protein Assays to Determine SARS-CoV-2 Antibody Levels: A Head-to-Head Comparison of Five Quantitative Assays. Microbiol. Spectr. 2021, 9, e0024721. [Google Scholar] [CrossRef] [PubMed]
- Karger, A.B.; Brien, J.D.; Christen, J.M.; Dhakal, S.; Kemp, T.J.; Klein, S.L.; Pinto, L.A.; Premkumar, L.; Roback, J.D.; Binder, R.A.; et al. The Serological Sciences Network (SeroNet) for COVID-19: Depth and Breadth of Serology Assays and Plans for Assay Harmonization. mSphere 2022, 7, e0019322. [Google Scholar] [CrossRef]
- Kroidl, I.; Winter, S.; Rubio-Acero, R.; Bakuli, A.; Geldmacher, C.; Eser, T.M.; Deak, F.; Horn, S.; Zielke, A.; Ahmed, M.I.M.; et al. Studying temporal titre evolution of commercial SARS-CoV-2 assays reveals significant shortcomings of using BAU standardization for comparison. Virol. J. 2023, 20, 200. [Google Scholar] [CrossRef]
- Walmsley, S.; Nabipoor, M.; Lovblom, L.E.; Ravindran, R.; Colwill, K.; McGeer, A.; Dayam, R.M.; Manase, D.; Gingras, A.C.; On behalf of the STOPCoV Team. Predictors of Breakthrough SARS-CoV-2 Infection after Vaccination. Vaccines 2023, 12, 36. [Google Scholar] [CrossRef]
- Zedan, H.T.; Yassine, H.M.; Al-Sadeq, D.W.; Liu, N.; Qotba, H.; Nicolai, E.; Pieri, M.; Bernardini, S.; Abu-Raddad, L.J.; Nasrallah, G.K. Evaluation of commercially available fully automated and ELISA-based assays for detecting anti-SARS-CoV-2 neutralizing antibodies. Sci. Rep. 2022, 12, 19020. [Google Scholar] [CrossRef]
- Gutlin, Y.; Albertos Torres, D.; Gensch, A.; Schlotterbeck, A.K.; Stoger, L.; Heller, S.; Infanti, L.; Barut, G.T.; Thiel, V.; Leuzinger, K.; et al. Anti-SARS-CoV-2 total immunoglobulin and neutralising antibody responses in healthy blood donors throughout the COVID-19 pandemic: A longitudinal observational study. Swiss Med. Wkly. 2024, 154, 3408. [Google Scholar] [CrossRef]
- Swadzba, J.; Anyszek, T.; Panek, A.; Chojeta, A.; Wyrzykowska, K.; Martin, E. Head-to-Head Comparison of 5 Anti-SARS-CoV-2 Assays Performance in One Hundred COVID-19 Vaccinees, over an 8-Month Course. Diagnostics 2022, 12, 1426. [Google Scholar] [CrossRef] [PubMed]
- Cao, Y.; Wang, J.; Jian, F.; Xiao, T.; Song, W.; Yisimayi, A.; Huang, W.; Li, Q.; Wang, P.; An, R.; et al. Omicron escapes the majority of existing SARS-CoV-2 neutralizing antibodies. Nature 2022, 602, 657–663. [Google Scholar] [CrossRef] [PubMed]
- Planas, D.; Saunders, N.; Maes, P.; Guivel-Benhassine, F.; Planchais, C.; Buchrieser, J.; Bolland, W.H.; Porrot, F.; Staropoli, I.; Lemoine, F.; et al. Considerable escape of SARS-CoV-2 Omicron to antibody neutralization. Nature 2022, 602, 671–675. [Google Scholar] [CrossRef]
- Shroff, R.T.; Chalasani, P.; Wei, R.; Pennington, D.; Quirk, G.; Schoenle, M.V.; Peyton, K.L.; Uhrlaub, J.L.; Ripperger, T.J.; Jergovic, M.; et al. Immune responses to two and three doses of the BNT162b2 mRNA vaccine in adults with solid tumors. Nat. Med. 2021, 27, 2002–2011. [Google Scholar] [CrossRef]
- Springer, D.N.; Holtl, E.; Pruger, K.; Puchhammer-Stockl, E.; Aberle, J.H.; Stiasny, K.; Weseslindtner, L. Measuring Variant-Specific Neutralizing Antibody Profiles after Bivalent SARS-CoV-2 Vaccinations Using a Multivariant Surrogate Virus Neutralization Microarray. Vaccines 2024, 12, 94. [Google Scholar] [CrossRef] [PubMed]
- Springer, D.N.; Traugott, M.; Reuberger, E.; Kothbauer, K.B.; Borsodi, C.; Nageli, M.; Oelschlagel, T.; Kelani, H.; Lammel, O.; Deutsch, J.; et al. A Multivariant Surrogate Neutralization Assay Identifies Variant-Specific Neutralizing Antibody Profiles in Primary SARS-CoV-2 Omicron Infection. Diagnostics 2023, 13, 2278. [Google Scholar] [CrossRef]
- Santos da Silva, E.; Servais, J.Y.; Kohnen, M.; Arendt, V.; Staub, T.; The CON-VINCE Consortium; The CoVaLux Consortium; Kruger, R.; Fagherazzi, G.; Wilmes, P.; et al. Validation of a SARS-CoV-2 Surrogate Neutralization Test Detecting Neutralizing Antibodies against the Major Variants of Concern. Int. J. Mol. Sci. 2023, 24, 14965. [Google Scholar] [CrossRef]
- Ulrich, L.; Halwe, N.J.; Taddeo, A.; Ebert, N.; Schon, J.; Devisme, C.; Trueb, B.S.; Hoffmann, B.; Wider, M.; Fan, X.; et al. Enhanced fitness of SARS-CoV-2 variant of concern Alpha but not Beta. Nature 2022, 602, 307–313. [Google Scholar] [CrossRef]
- Han, P.; Li, L.; Liu, S.; Wang, Q.; Zhang, D.; Xu, Z.; Han, P.; Li, X.; Peng, Q.; Su, C.; et al. Receptor binding and complex structures of human ACE2 to spike RBD from omicron and delta SARS-CoV-2. Cell 2022, 185, 630–640.e10. [Google Scholar] [CrossRef]
- Liu, Y.; Liu, J.; Plante, K.S.; Plante, J.A.; Xie, X.; Zhang, X.; Ku, Z.; An, Z.; Scharton, D.; Schindewolf, C.; et al. The N501Y spike substitution enhances SARS-CoV-2 infection and transmission. Nature 2022, 602, 294–299. [Google Scholar] [CrossRef]
- Geisen, U.M.; Rose, R.; Neumann, F.; Ciripoi, M.; Vullriede, L.; Reid, H.M.; Berner, D.K.; Bertoglio, F.; Hoff, P.; Hust, M.; et al. The long term vaccine-induced anti-SARS-CoV-2 immune response is impaired in quantity and quality under TNFalpha blockade. J. Med. Virol. 2022, 94, 5780–5789. [Google Scholar] [CrossRef] [PubMed]
- Kirsebom, F.C.M.; Andrews, N.; Stowe, J.; Ramsay, M.; Lopez Bernal, J. Duration of protection of ancestral-strain monovalent vaccines and effectiveness of bivalent BA.1 boosters against COVID-19 hospitalisation in England: A test-negative case-control study. Lancet Infect. Dis. 2023, 23, 1235–1243. [Google Scholar] [CrossRef] [PubMed]
- Depery, L.; Bally, I.; Amen, A.; Nemoz, B.; Buisson, M.; Grossi, L.; Truffot, A.; Germi, R.; Guilligay, D.; Veloso, M.; et al. Anti-SARS-CoV-2 serology based on ancestral RBD antigens does not correlate with the presence of neutralizing antibodies against Omicron variants. Microbiol. Spectr. 2025, 13, e0156824. [Google Scholar] [CrossRef]
- Garcia, M.J.; Rodriguez-Duque, J.C.; Pascual, M.; Rivas, C.; Castro, B.; Raso, S.; Lopez-Hoyos, M.; Arias-Loste, M.T.; Rivero, M. Prevalence of antinuclear antibodies in inflammatory bowel disease and seroconversion after biological therapy. Ther. Adv. Gastroenterol. 2022, 15, 17562848221077837. [Google Scholar] [CrossRef]
- Muratori, P.; Lenzi, M.; Muratori, L.; Granito, A. Antinuclear antibodies in COVID 19. Clin. Transl. Sci. 2021, 14, 1627–1628. [Google Scholar] [CrossRef] [PubMed]
- Pascolini, S.; Granito, A.; Muratori, L.; Lenzi, M.; Muratori, P. Coronavirus disease associated immune thrombocytopenia: Causation or correlation? J. Microbiol. Immunol. Infect. 2021, 54, 531–533. [Google Scholar] [CrossRef]
- Hromic-Jahjefendic, A.; Lundstrom, K.; Adilovic, M.; Aljabali, A.A.A.; Tambuwala, M.M.; Serrano-Aroca, A.; Uversky, V.N. Autoimmune response after SARS-CoV-2 infection and SARS-CoV-2 vaccines. Autoimmun. Rev. 2024, 23, 103508. [Google Scholar] [CrossRef]
- Curman, P.; Kridin, K.; Zirpel, H.; Hernandez, G.; Akyuz, M.; Thaci, D.; Schmidt, E.; Ludwig, R.J. COVID-19 infection is associated with an elevated risk for autoimmune blistering diseases while COVID-19 vaccination decreases the risk: A large-scale population-based cohort study of 112 million individuals. J. Am. Acad. Dermatol. 2025, 92, 452–463. [Google Scholar] [CrossRef]
- Allocca, M.; Chaparro, M.; Gonzalez, H.A.; Bosca-Watts, M.M.; Palmela, C.; D’Amico, F.; Zacharopoulou, E.; Kopylov, U.; Ellul, P.; Bamias, G.; et al. Patients with Inflammatory Bowel Disease Are Not at Increased Risk of COVID-19: A Large Multinational Cohort Study. J. Clin. Med. 2020, 9, 3533. [Google Scholar] [CrossRef]
- Meyer, A.; Semenzato, L.; Zureik, M.; Weill, A.; Carbonnel, F.; Dray-Spira, R. Risk of severe COVID-19 in patients treated with IBD medications: A French nationwide study. Aliment. Pharmacol. Ther. 2021, 54, 160–166. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Patients | Infliximab (n = 10) | Healthy Controls (n = 11) | p-Value | |
|---|---|---|---|---|
| Patient characteristics | Age, years median (IQR) | 50 (31–65) | 35 (23–66) | 0.285 |
| Sex, male (%) | 4 (40) | 4 (36) | 0.863 | |
| SARS-CoV-2 vaccine | Comirnaty BNT162b2 (%) | 10 (100) | 4 (23) | |
| Spikevax mRNA-1273 (%) | 0 (0) | 7 (77) | ||
| IBD | Crohn’s disease (%) | 8 (80) | 0 (0) | |
| Ulcerative colitis (%) | 2 (20) | 0 (0) | ||
| Medication | Prednisolone p.o. (%) | 0 (0) | 0 (0) | |
| Budesonide p.o. (%) | 1 (10) | 0 (0) | ||
| Budesonide supp. (%) | 0 (0) | 0 (0) | ||
| Mesalazine p.o. (%) | 6 (60) | 0 (0) | ||
| Mesalazine supp. (%) | 0 (0) | 0 (0) | ||
| Pre-existing conditions | Cardiovascular disease (%) | 3 (30) | 0 (0) | 0.251 |
| Respiratory disease (%) | 0 (0) | 0 (0) | 1.000 | |
| Kidney insufficiency (%) | 0 (0) | 0 (0) | 1.000 | |
| Metastatic neoplasm (%) | 0 (0) | 0 (0) | 1.000 | |
| Diabetes (%) | 0 (0) | 0 (0) | 1.000 | |
| Hematologic malignancy (%) | 0 (0) | 0 (0) | 1.000 |
| Assay | Area 1 | p Value 1 | Adapted Cut-Off Value 1 | Sensitivity (%) at Adapted Cut-Off Value 1 | Specificity (%) at Adapted Cut-Off Value 1 |
|---|---|---|---|---|---|
| BA.1 S pVNT | 0.8670 | <0.0001 | ≥20.09% | 82.61 | 88.24 |
| Anti-RBD CMIA | 0.8619 | 0.0001 | ≥1126 AU/mL | 82.61 | 88.24 |
| cPass sVNT | 0.8542 | 0.0002 | ≥49.40% | 91.3 | 70.59 |
| wt RBD sVNT | 0.8338 | 0.0004 | ≥59.55% | 78.26 | 76.47 |
| Alpha RBD sVNT | 0.8414 | 0.0003 | ≥25.35% | 86.96 | 70.59 |
| Beta RBD sVNT | 0.8159 | 0.0007 | ≥72.27% | 60.87 | 94.12 |
| Gamma RBD sVNT | 0.8338 | 0.0004 | ≥54.60% | 73.91 | 82.35 |
| Delta RBD sVNT | 0.8235 | 0.0005 | ≥72.07% | 78.26 | 82.35 |
| BA.1 RBD sVNT | 0.6176 | 0.2082 | ≥34.73% | 43.48 | 88.24 |
| BA.5 RBD sVNT | 0.7187 | 0.0193 | ≥9.095% | 60.87 | 94.12 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lorentzen, E.U.; Vollenberg, R.; Neddermeyer, R.; Schoefbaenker, M.; Hrincius, E.R.; Ludwig, S.; Tepasse, P.-R.; Kuehn, J.E. SARS-CoV-2 Variant-Specific Antibodies in Vaccinated Inflammatory Bowel Disease Patients. Vaccines 2025, 13, 595. https://doi.org/10.3390/vaccines13060595
Lorentzen EU, Vollenberg R, Neddermeyer R, Schoefbaenker M, Hrincius ER, Ludwig S, Tepasse P-R, Kuehn JE. SARS-CoV-2 Variant-Specific Antibodies in Vaccinated Inflammatory Bowel Disease Patients. Vaccines. 2025; 13(6):595. https://doi.org/10.3390/vaccines13060595
Chicago/Turabian StyleLorentzen, Eva Ulla, Richard Vollenberg, Rieke Neddermeyer, Michael Schoefbaenker, Eike R. Hrincius, Stephan Ludwig, Phil-Robin Tepasse, and Joachim Ewald Kuehn. 2025. "SARS-CoV-2 Variant-Specific Antibodies in Vaccinated Inflammatory Bowel Disease Patients" Vaccines 13, no. 6: 595. https://doi.org/10.3390/vaccines13060595
APA StyleLorentzen, E. U., Vollenberg, R., Neddermeyer, R., Schoefbaenker, M., Hrincius, E. R., Ludwig, S., Tepasse, P.-R., & Kuehn, J. E. (2025). SARS-CoV-2 Variant-Specific Antibodies in Vaccinated Inflammatory Bowel Disease Patients. Vaccines, 13(6), 595. https://doi.org/10.3390/vaccines13060595