
Immunogenicity and Safety Profile of Two Adjuvanted-PD-L1-Based Vaccine Candidates in Mice, Rats, Rabbits, and Cynomolgus Monkeys

 , , ,
, , ,  and
and
Abstract

1. Introduction
2. Materials and Methods
2.1. Ethical Approval
2.2. Animals
2.3. Vaccine Antigen and Adjuvants
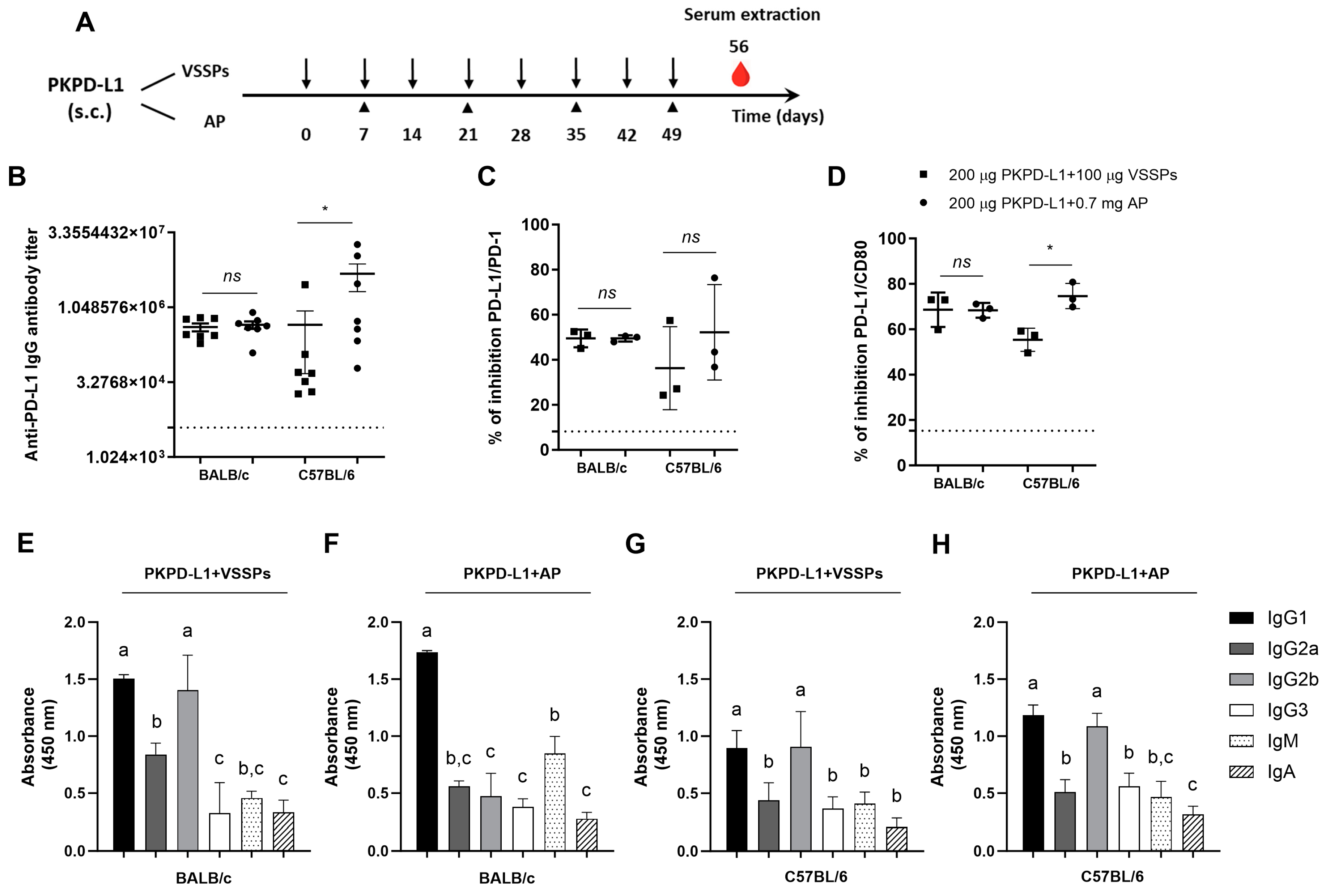
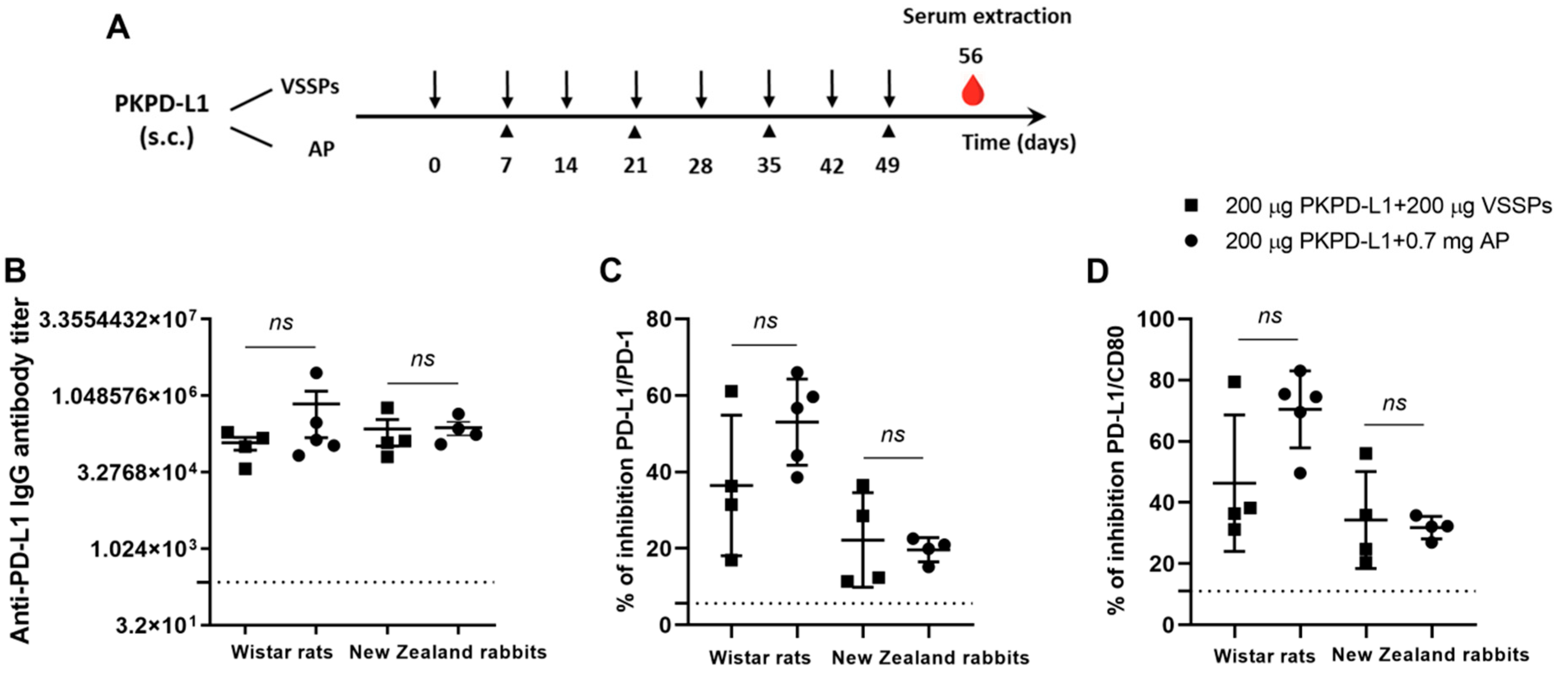
2.4. Immunization Schedules
2.5. ELISA Reagents
2.6. ELISA Procedures
2.6.1. ELISA for Detecting PD-L1 Specific IgG Human Antibodies
2.6.2. Competitive ELISA for Measuring Serum-Mediated Inhibition of PD-1/PD-L1 Interaction
2.7. Specific Anti-PDL1 Antibody Class and Subclass
2.8. Follow-Up of Clinical, Behavioral and Hematological and Serum Biochemical Parameters
2.8.1. Mice
2.8.2. Rats
2.8.3. Rabbits
2.8.4. Non-Human Primates
2.9. Rats and Rabbits Anatomopathological Studies and Organ Weights
2.10. Statistical Analysis
3. Results
3.1. Active Immunization with PKPD-L1Vac Induces Specific Anti-hPD-L1 IgG Antibodies in Mice
3.2. Active Immunization with PKPD-L1Vac Induces Specific Anti-hPD-L1 IgG Antibodies in Rats and Rabbits
3.3. Active Immunization with Recombinant PKPD-L1 Antigen Induces Specific Anti-hPD-L1 IgG Antibodies in Non-Human Primates
3.4. Clinical, Hematological and Biochemical Parameters
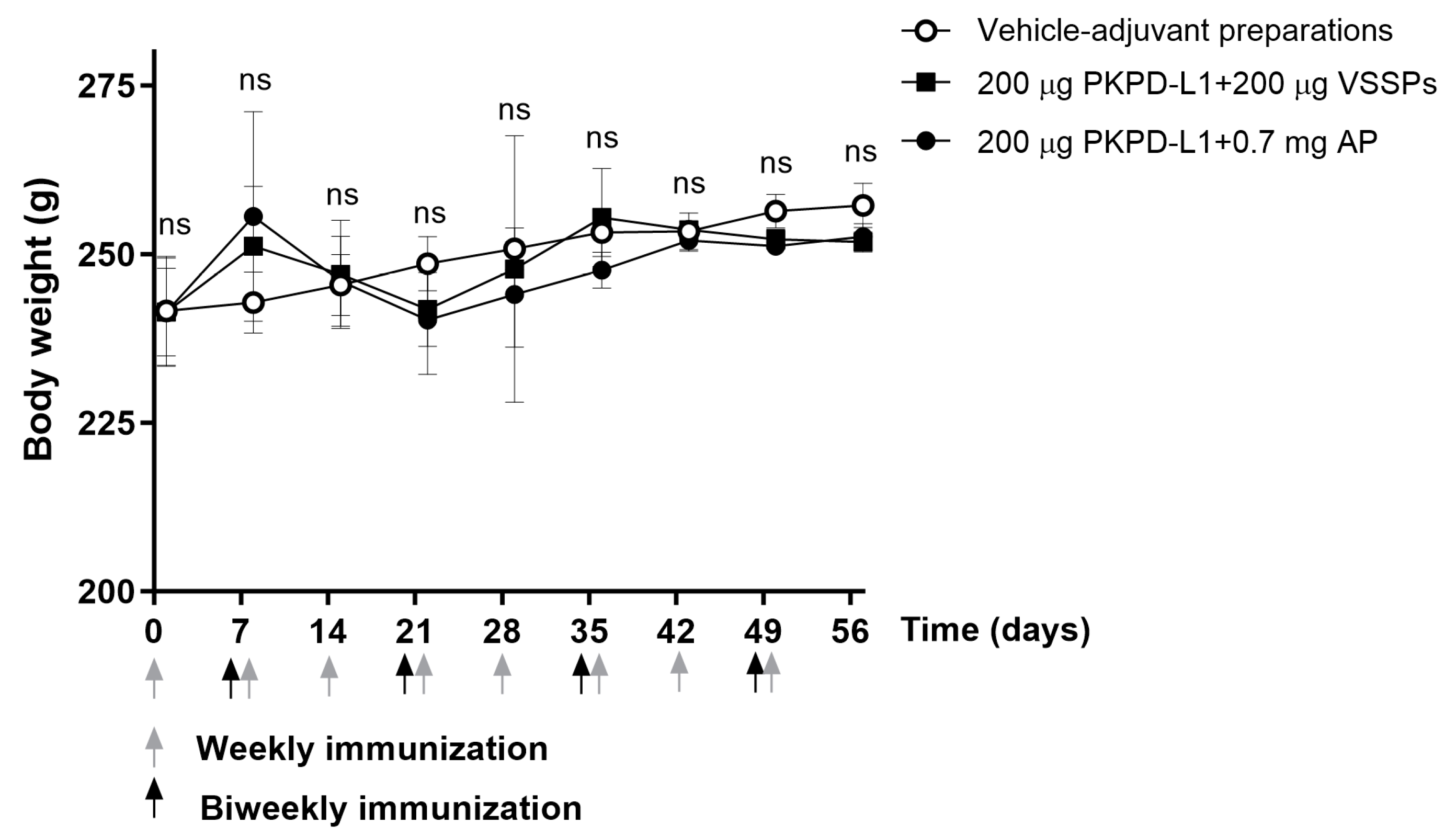
3.4.1. Mice
3.4.2. Rats
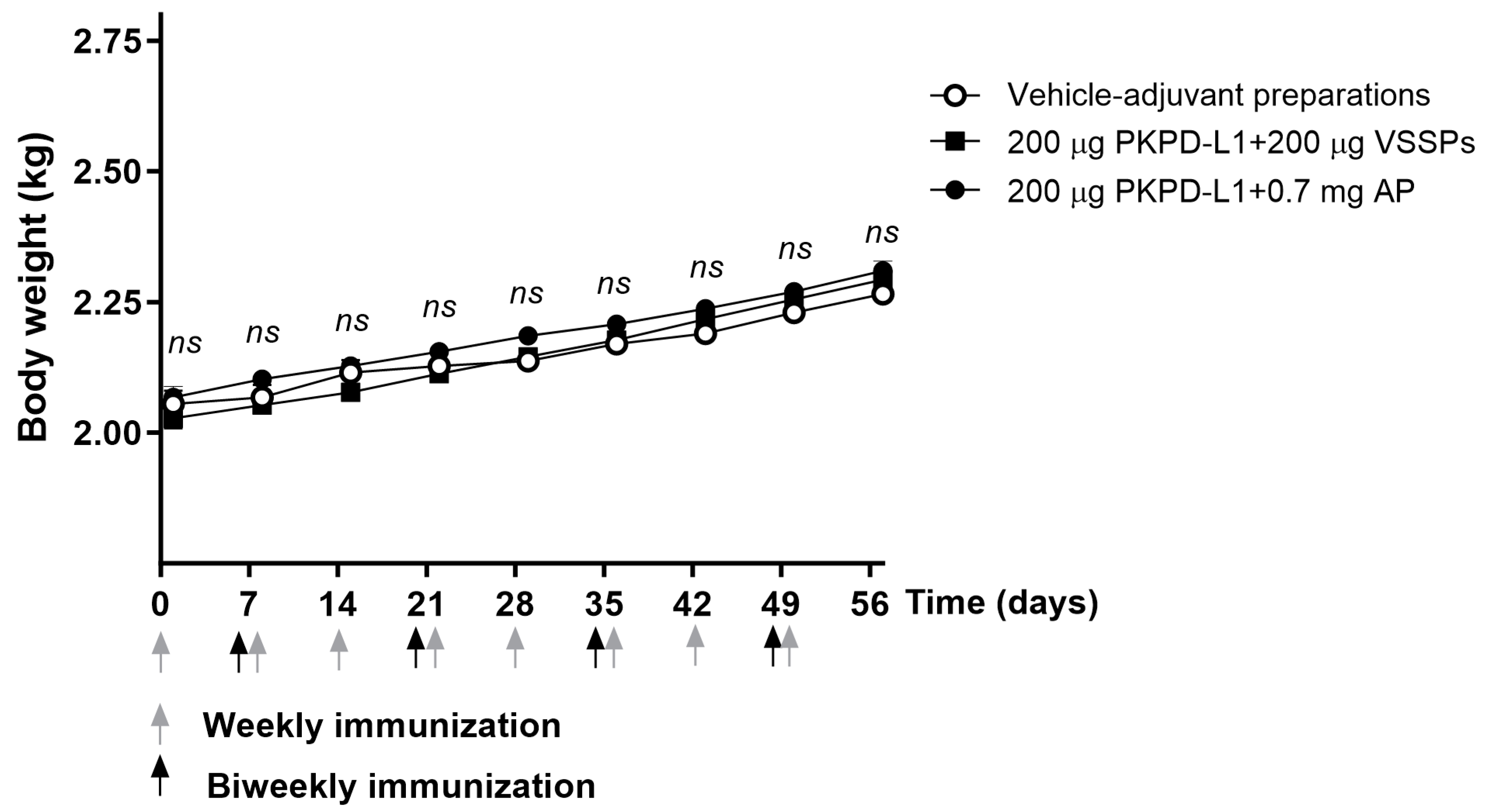
3.4.3. Rabbits
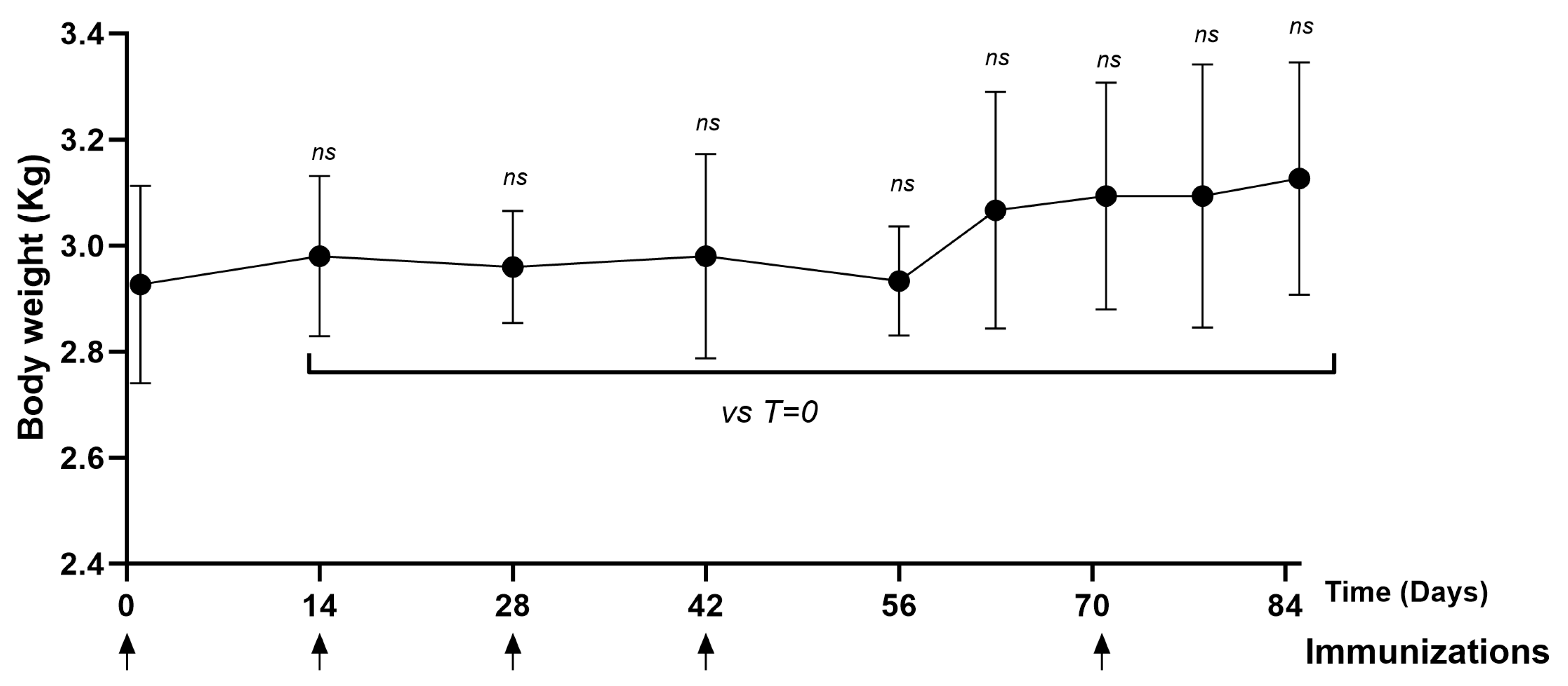
3.4.4. Non-Human Primates (NHPs)
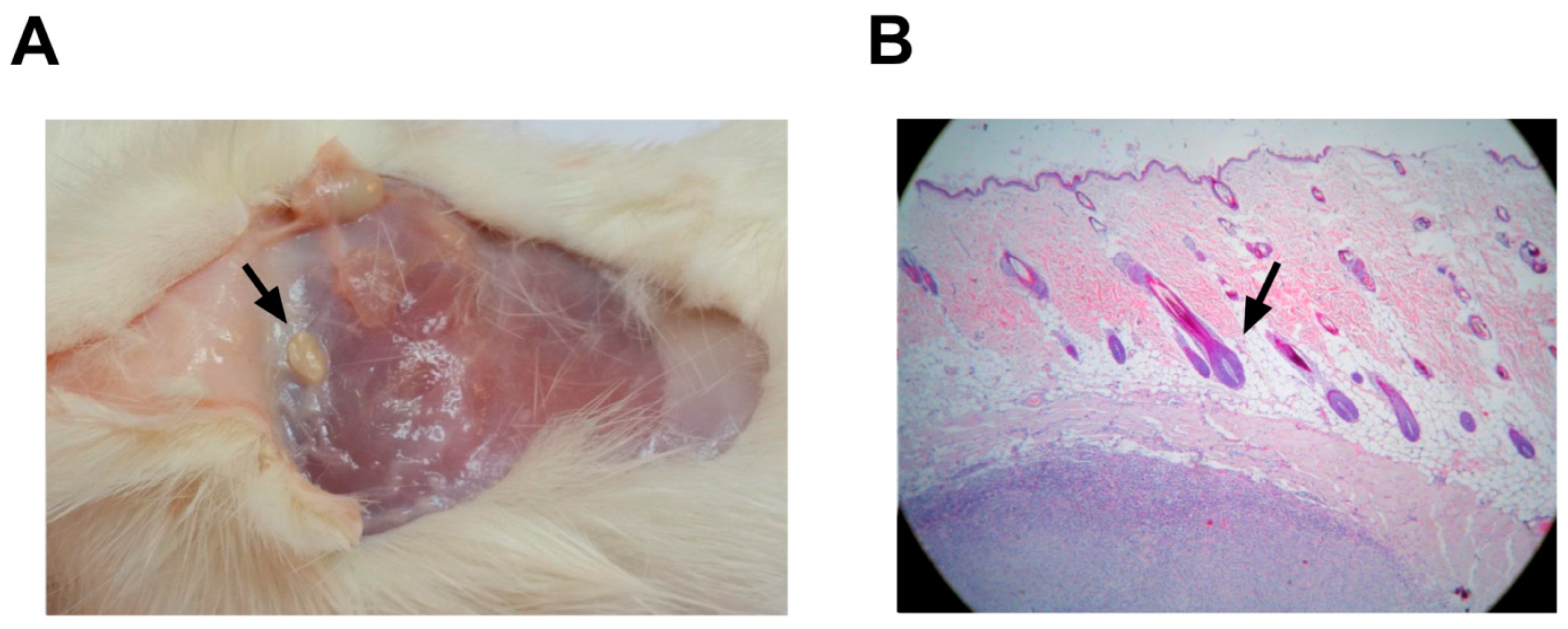
3.5. Anatomopathological Studies in Rats and Rabbits
3.5.1. Rats
3.5.2. Rabbits
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Munari, E.; Mariotti, F.R.; Quatrini, L.; Bertoglio, P.; Tumino, N.; Vacca, P.; Eccher, A.; Ciompi, F.; Brunelli, M.; Martignoni, G.; et al. PD-1/PD-L1 in Cancer: Pathophysiological, Diagnostic and Therapeutic Aspects. Int. J. Mol. Sci. 2021, 22, 5123. [Google Scholar] [CrossRef] [PubMed]
- Ai, L.; Chen, J.; Yan, H.; He, Q.; Luo, P.; Xu, Z.; Yang, X. Research Status and Outlook of PD-1/PD-L1 Inhibitors for Cancer Therapy. Drug Des. Dev. Ther. 2020, 14, 3625–3649. [Google Scholar] [CrossRef] [PubMed]
- Twomey, J.D.; Zhang, B. Cancer Immunotherapy Update: FDA-Approved Checkpoint Inhibitors and Companion Diagnostics. AAPS J. 2021, 23, 39. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Xu, R. Immunotherapy-related adverse events (irAEs): Extraction from FDA drug labels and comparative analysis. JAMIA Open. 2019, 2, 173–178. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Zhang, Z. The history and advances in cancer immunotherapy: Understanding the characteristics of tumor-infiltrating immune cells and their therapeutic implications. Cell. Mol. Immunol. 2020, 17, 807–821. [Google Scholar] [CrossRef]
- Jørgensen, N.G.; Kaae, J.; Grauslund, J.H.; Met, Ö. Vaccination against PD-L1 with IO103 a Novel Immune Modulatory Vaccine in Basal Cell Carcinoma: A Phase IIa Study. Cancers 2021, 13, 911. [Google Scholar] [CrossRef]
- Guo, L.; Overholser, J.; Darby, H.; Ede, N.J.; Kaumaya, P.T.P. A newly discovered PD-L1 B-cell epitope peptide vaccine (PDL1-Vaxx) exhibits potent immune responses and effective anti-tumor immunity in multiple syngeneic mice models and (synergizes) in combination with a dual HER-2 B-cell vaccine (B-Vaxx). Oncoimmunology 2022, 11, 2127691. [Google Scholar] [CrossRef]
- Tian, H.; Kang, Y.; Song, X.; Xu, Y.; Chen, H.; Gong, X.; Zhang, W.; Xu, Y.; Xia, X.; Gao, X.; et al. PDL1-targeted vaccine exhibits potent antitumor activity by simultaneously blocking PD1/PDL1 pathway and activating PDL1-specific immune responses. Cancer Lett. 2020, 476, 170–182. [Google Scholar] [CrossRef]
- Lin, Z.; Zhang, Y.; Cai, H.; Zhou, F.; Gao, H.; Deng, L.; Li, R. A PD-L1-Based Cancer Vaccine Elicits Antitumor Immunity in a Mouse Melanoma Model. Mol. Ther. Oncolytics 2019, 14, 222–232. [Google Scholar] [CrossRef]
- Morera-Díaz, Y.; Canaán-Haden, C.; Sánchez-Ramírez, J.; Bequet-Romero, M.; Gonzalez-Moya, I.; Martínez, R.; Falcón, V.; Palenzuela, D.; Ayala-Ávila, M.; Gavilondo, J.V. Active immunization with a structurally aggregated PD-L1 antigen breaks T and B immune tolerance in non-human primates and exhibits in vivo anti-tumoral effects in immunocompetent mouse tumor models. Cancer Lett. 2023, 561, 216156. [Google Scholar] [CrossRef]
- HogenEsch, H.; O’Hagan, D.T.; Fox, C.B. Optimizing the utilization of aluminum adjuvants in vaccines: You might just get what you want. npj Vaccines 2018, 3, 51. [Google Scholar] [CrossRef] [PubMed]
- Aguilar, F.F.; Barranco, J.J.; Fuentes, E.B.; Aguilera, L.C.; Sáez, Y.L.; Santana, M.D.; Vázquez, E.P.; Baker, R.B.; Acosta, O.R.; Pérez, H.G.; et al. Very small size proteoliposomes (VSSP) and Montanide combination enhance the humoral immuno response in a GnRH based vaccine directed to prostate cancer. Vaccine 2012, 30, 6595–6599. [Google Scholar] [CrossRef] [PubMed]
- Khan, A.N.H.; Emmons, T.R.; Magner, W.J.; Alqassim, E.; Singel, K.L.; Ricciuti, J.; Eng, K.H.; Odunsi, K.; Tomasi, T.B.; Lee, K.; et al. VSSP abrogates murine ovarian tumor-associated myeloid cell-driven immune suppression and induces M1 polarization in tumor-associated macrophages from ovarian cancer patients. Cancer Immunol. Immunother. 2022, 71, 2355–2369. [Google Scholar] [CrossRef] [PubMed]
- Gumusay, O.; Callan, J.; Rugo, H.S. Immunotherapy toxicity: Identification and management. Breast Cancer Res. Treat. 2022, 192, 1–17. [Google Scholar] [CrossRef]
- Hargrave, A.; Mustafa, A.S.; Hanif, A.; Tunio, J.H.; Hanif, S.N.M. Recent Advances in Cancer Immunotherapy with a Focus on FDA-Approved Vaccines and Neoantigen-Based Vaccines. Vaccines 2023, 11, 1633. [Google Scholar] [CrossRef]
- Verma, S.K.; Mahajan, P.; Singh, N.K.; Gupta, A.; Aggarwal, R.; Rappuoli, R.; Johri, A.K. New-age vaccine adjuvants, their development, and future perspective. Front. Immunol. 2023, 14, 1043109. [Google Scholar] [CrossRef]
- Morera, Y.; Bequet-Romero, M.; Ayala, M.; Velazco, J.C.; Pérez, P.P.; Alba, J.S.; Ancizar, J.; Rodríguez, M.; Cosme, K.; Gavilondo, J.V. Immunogenicity and some safety features of a VEGF-based cancer therapeutic vaccine in rats, rabbits and non-human primates. Vaccine 2010, 28, 3453–3461. [Google Scholar] [CrossRef]
- Ramírez, B.S.; Alpízar, Y.A.; Fernández, D.R.; Hidalgo, G.G.; Capote, A.R.; Rodríguez, R.P.; Fernández, L.E. Anti-EGFR activation, anti-proliferative and pro-apoptotic effects of polyclonal antibodies induced by EGFR-based cancer vaccine. Vaccine 2008, 26, 4918–4926. [Google Scholar] [CrossRef]
- Barro, A.M.; Rivero, A.I.; Goñi, A.L.; Navarro, B.O.; Angarica, M.M.; Ramírez, B.S.; Bedoya, D.M.; Triana, C.G.; Rodríguez, A.M.; Parada, Á.C. Non-clinical immuno-toxicological evaluation of HER1 cancer vaccine in non-human primates: A 12-month study. Vaccine 2012, 31, 89–95. [Google Scholar] [CrossRef]
- Zeng, M.; Nourishirazi, E.; Guinet, E.; Nouri-Shirazi, M. The genetic background influences the cellular and humoral immune responses to vaccines. Clin. Exp. Immunol. 2016, 186, 190–204. [Google Scholar] [CrossRef]
- Fornefett, J.; Krause, J.; Klose, K.; Fingas, F.; Hassert, R.; Benga, L.; Grunwald, T.; Müller, U.; Schrödl, W.; Baums, C.G. Comparative analysis of humoral immune responses and pathologies of BALB/c and C57BL/6 wildtype mice experimentally infected with a highly virulent Rodentibacter pneumotropicus (Pasteurella pneumotropica) strain. BMC Microbiol. 2018, 18, 45. [Google Scholar] [CrossRef] [PubMed]
- Song, H.K.; Hwang, D.Y. Use of C57BL/6N mice on the variety of immunological researches. Lab. Anim. Res. 2017, 33, 119–123. [Google Scholar] [CrossRef] [PubMed]
- Jin, S.; Sun, Y.; Liang, X.; Gu, X.; Ning, J.; Xu, Y.; Chen, S.; Pan, L. Emerging new therapeutic antibody derivatives for cancer treatment. Signal Transduct. Target. Ther. 2022, 7, 39. [Google Scholar] [CrossRef] [PubMed]
- Visciano, M.L.; Tagliamonte, M.; Tornesello, M.L.; Buonaguro, F.M.; Buonaguro, L. Effects of adjuvants on IgG subclasses elicited by virus-like particles. J. Transl. Med. 2012, 10, 4. [Google Scholar] [CrossRef] [PubMed]
- Snapper, C.M. Distinct Immunologic Properties of Soluble Versus Particulate Antigens. Front. Immunol. 2018, 9, 598. [Google Scholar] [CrossRef]
- Mesa, C.; León, J.; Fernández, L.E. VSSP: A new adjuvant for the vaccination of immuno-compromised patients. Biotecnol. Apl. 2005, 22, 228–233. [Google Scholar]
- Lei, Q.; Wang, D.; Sun, K.; Wang, L.; Zhang, Y. Resistance Mechanisms of Anti-PD1/PDL1 Therapy in Solid Tumors. Front. Cell Dev. Biol. 2020, 8, 672. [Google Scholar] [CrossRef]
- Liu, J.; Chen, Z.; Li, Y.; Zhao, W.; Wu, J.; Zhang, Z. PD-1/PD-L1 Checkpoint Inhibitors in Tumor Immunotherapy. Front. Pharmacol. 2021, 12, 731798. [Google Scholar] [CrossRef]
- Honess, P. A brief history of primate research: Global health improvements and ethical challenges. Arch. Med. Biomed. Res. 2016, 2, 151. [Google Scholar] [CrossRef]
- Morera-Díaz, Y.; Gavilondo, J.V.; Bequet-Romero, M.; Ramírez, J.S.; Hernández-Bernal, F.; Selman-Housein, K.H.; Perez, L.; Ayala-Ávila, M. Specific active immunotherapy with the HEBERSaVax VEGF-based cancer vaccine: From bench to bedside. Semin. Oncol. 2018, 45, 68–74. [Google Scholar] [CrossRef]
- Gavilondo, J.V.; Hernández-Bernal, F.; Ayala-Ávila, M.; de la Torre, A.V.; de la Torre, J.; Morera-Díaz, Y.; Bequet-Romero, M.; Sánchez, J.; Valenzuela, C.M.; Martín, Y.; et al. Specific active immunotherapy with a VEGF vaccine in patients with advanced solid tumors. results of the CENTAURO antigen dose escalation phase I clinical trial. Vaccine 2014, 32, 2241–2250. [Google Scholar] [CrossRef] [PubMed]
- Carrodeguas, R.A.O.; Monteagudo, G.L.; Chaviano, P.P.G.; Montané, I.Á.; Saldívar, E.E.S.; Lambert, L.L.; Sosa, K.C.; Pino, R.B.; Mustelier, P.B.; Rodríguez, E.V.; et al. Safety and effectiveness of CIMAvax-EGF administered in community polyclinics. Front. Oncol. 2023, 13, 1287902. [Google Scholar] [CrossRef] [PubMed]
- Caballero, I.; Aira, L.E.; Lavastida, A.; Popa, X.; Rivero, J.; González, J.; Mesa, M.; González, N.; Coba, K.; Lorenzo-Luaces, P.; et al. Safety and Immunogenicity of a Human Epidermal Growth Factor Receptor 1 (HER1)-Based Vaccine in Prostate Castration-Resistant Carcinoma Patients: A Dose-Escalation Phase I Study Trial. Front. Pharmacol. 2017, 8, 263. [Google Scholar] [CrossRef] [PubMed]
- Maeng, H.M.; Moore, B.N.; Bagheri, H.; Steinberg, S.M.; Inglefield, J.; Dunham, K.; Wei, W.Z.; Morris, J.C.; Terabe, M.; England, L.C.; et al. Phase I Clinical Trial of an Autologous Dendritic Cell Vaccine Against HER2 Shows Safety and Preliminary Clinical Efficacy. Front. Oncol. 2021, 11, 789078. [Google Scholar] [CrossRef]
- Oliva-Hernández, R. Considerations of preclinical toxicological studies for vaccines. VacciMonitor 2019, 28, 48–51. [Google Scholar]
- Algaissi, A.; Alamer, E.; Jeraiby, M.; Alomaish, A.; Elrhima, O.; Qumayi, S.; Qasir, N.A.; Areeshi, H.; Masmali, A.; Alhazmi, A. Evaluation of hematological parameters and thrombocytopenia following Pfizer-BioNTech (BNT162b2) SARS-CoV-2 vaccination. Saudi Med. J. 2022, 43, 567–571. [Google Scholar] [CrossRef]
- Li, W.; Chang, N.; Li, L. Heterogeneity and Function of Kupffer Cells in Liver Injury. Front. Immunol. 2022, 13, 940867. [Google Scholar] [CrossRef]
- Nguyen-Lefebvre, A.T.; Horuzsko, A. Kupffer Cell Metabolism and Function. J. Enzymol. Metab. 2015, 1, 101. [Google Scholar]
- Segatori, V.I.; Otero, L.L.; Fernandez, L.E.; Gomez, D.E.; Alonso, D.F.; Gabri, M.R. Antitumor protection by NGcGM3/VSSP vaccine against transfected B16 mouse melanoma cells overexpressing N-glycolylated gangliosides. In Vivo 2012, 26, 609–617. [Google Scholar]
- Zhang, P.; Wang, Y.; Miao, Q.; Chen, Y. The therapeutic potential of PD-1/PD-L1 pathway on immune-related diseases: Based on the innate and adaptive immune components. Biomed. Pharmacother. 2023, 167, 115569. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameter | Physiological Range (Unit) | GROUPS | DAY 0 (Pre-Vaccination) | DAY 56 (Post-Vaccination) |
|---|---|---|---|---|
| White blood cells | 5.2–12.5 (103/μL) | I | 10.35 ± 2.105 a | 15.28 ± 4.484 a |
| II | 8.4 ± 0.966 a | 14.63 ± 2.265 a | ||
| III | 11.3 ± 1.386 a | 13.33 ± 2.281 a | ||
| Red blood cells | 5.3–6.8 (106/μL) | I | 7.41 ± 0.312 a | 6.58 ± 0.5 a |
| II | 6.64 ± 0.212 a | 6.74 ± 0.512 a | ||
| III | 6.98 ± 0.137 a | 6.96 ± 0.441 a | ||
| Hemoglobin | 9.8–14 (g/dL) | I | 12.58 ± 0.78 a | 11.15 ± 0.975 a |
| II | 11 ± 0.216 a | 11.9 ± 1.122 a | ||
| III | 11.93 ± 0.499 a | 12.05 ± 0.794 a | ||
| Hematocrit | 34–43 (%) | I | 43.9 ± 2.483 a | 39.13 ± 3.198 a |
| II | 39.8 ± 1.087 a | 40.78 ± 3.556 a | ||
| III | 41.58 ± 1.597 a | 41.88 ± 2.947 a | ||
| Mean Corpuscular volume | 60–69 (fL) | I | 59.23 ± 2.141 a | 59.45 ± 1.723 a |
| II | 58.83 ± 0.538 a | 60.5 ± 2.04 a | ||
| III | 59.58 ± 1.352 a | 60.18 ± 1.47 a | ||
| Mean Corpuscular Hemoglobin | 20–23 (Pg) | I | 16.95 ± 0.592 a | 16.95 ± 0.436 a |
| II | 16.58 ± 0.403 a | 17.68 ± 0.826 a | ||
| III | 17.1 ± 0.469 a | 17.35 ± 0.493 a | ||
| Mean Corpuscular Hemoglobin Concentration | 31–35 (g/dL) | I | 28.65 ± 0.289 a | 28.45 ± 0.238 a |
| II | 28.15 ± 0.686 a | 29.18 ± 0.457 a | ||
| III | 28.68 ± 0.17 a | 28.78 ± 0.33 a | ||
| Platelet count | 158–650 (103/μL) | I | 172 ± 78.59 a | 515 ± 151.30 a |
| II | 261 ± 71.58 a | 354.25 ± 77.779 a | ||
| III | 330.25 ± 87.724 a | 286.75 ± 67.465 a | ||
| Neutrophils percentage | 25–75 (%) | I | 58.75 ± 9.535 a | 57.25 ± 8.342 a |
| II | 75.5 ± 7.853 a | 55.5 ± 13.304 a | ||
| III | 70.5 ± 10.017 a | 50.25 ± 9.5 a | ||
| Lymphocytes percentage | 30–85 (%) | I | 38.25 ± 10.996 a | 39.5 ± 9.037 a |
| II | 22.5 ± 7.047 a | 41.5 ± 13.304 a | ||
| III | 28.75 ± 10.34 a | 46.5 ± 9.678 a | ||
| Monocytes percentage | 1–4 (%) | I | 1.25 ± 2.5 a | 0.75 ± 0.957 a |
| II | 1 ± 0.816 a | 0.75 ± 0.957 a | ||
| III | 0.5 ± 10 a | 0.5 ± 0.577 a | ||
| Alanine aminotransferase | 10–45 (U/L) | I | 18.52 ± 6.414 a | 27.56 ± 8.241 a |
| II | 20.13 ± 6.072 a | 28.15 ± 5.663 a | ||
| III | 18.38 ± 6.363 a | 37.63 ± 6.694 a | ||
| Aspartate aminotransferase | 10–120 (U/L) | I | 7.44 ± 2.912 a | 22.9 ± 7.506 a |
| II | 19.69 ± 18.632 a | 21.73 ± 1.985 a | ||
| III | 12.25 ± 8.012 a | 17.94 ± 2.492 a | ||
| Creatinine | 0.5–2.5 (mg/dL) | I | 0.9 ± 0.648 a | 2.37 ± 0.287 a |
| II | 0.63 ± 0.574 a | 2.15 ± 0.287 a | ||
| III | 0.3 ± 0.600 a | 1.52 ± 1.166 a | ||
| Total Protein | 5.3–8.4 (g/dL) | I | 6.94 ± 1.763 a | 7.38 ± 0.506 a |
| II | 6.32 ± 0.517 a | 8.05 ± 1.414 a | ||
| III | 6.78 ± 0.524 a | 8.07 ± 0.760 a | ||
| Total bilirubin | 1.5–2.5 (mmol/L) | I | 1.04 ± 0.39 a | 0.76 ± 0.376 a |
| II | 0.86 ± 0.430 a | 0.68 ± 0.364 a | ||
| III | 1.2 ± 0.534 a | 0.57 ± 0.216 a | ||
| Albumin | 2.7–5 (g/dL) | I | 3.99 ± 0.568 a | 4.41 ± 0.812 a |
| II | 3.2 ± 0.520 a | 4.17 ± 0.483 a | ||
| III | 2.82 ± 0.314 a | 3.78 ± 0.706 a |
| PARAMETER | Physiological Range (Unit) | DAY 0 (Pre-Vaccination) | DAY 85 (Post-Vaccination) |
|---|---|---|---|
| White blood cells | 3.1–9.6 (103/μL) | 5.15 ± 0.07 a | 6.77 ± 2.19 a |
| Red blood cells | 5.10–7.9 (106/μL) | 6.22 ± 0.13 a | 6.29 ± 0.14 a |
| Hemoglobin | 11.00–19.04 (g/dL) | 12.30 ± 0.44 a | 12.30 ± 0.40 a |
| Hematocrit | 33.1–49.67 (%) | 42.43 ± 1.27 a | 42.83 ± 1.06 a |
| Mean Corpuscular volume | 52.02–76.06 (fL) | 68.33 ± 2.52 a | 68.33 ± 3.22 a |
| Mean Corpuscular Hemoglobin | 17.1–24.2 (Pg) | 19.77 ± 0.80 a | 19.57 ± 1.00 a |
| Mean Corpuscular Hemoglobin Concentration | 23.6–33.9 (g/dL) | 29.00 ± 0.10 a | 28.7 ± 0.17 a |
| Platelet count | 281–672 (103/μL) | 375 ± 33.18 a | 364.33 ± 9.07 a |
| Alanine aminotransferase | 1–56 (U/L) | 11 ± 2.65 a | 5.93 ± 3.25 a |
| Aspartate aminotransferase | 10.3–59.8 (U/L) | 31 ± 9.17 a | 33.27 ± 15.66 a |
| Creatinine | 0.1–1.10 (mg/dL) | 0.30 ± 0.05 a | 0.34 ± 0.44 a |
| Alkaline phosphatase | 60–850.4 (U/L) | 588.33 ± 233.33 a | 390 ± 152.59 a |
| Glucose | 56.81–99.09 (mmol/L) | 81.08 ± 0.457 a | 83.24 ± 0.272 a |
| Total bilirubin | 0.1–2.30 (mg/dL) | 3.58 ± 0.414 a | 1.70 ± 0.458 a |
| Direct bilirubin | 0.1–0.80 (mg/dL) | 0.80 ± 0.173 a | 0.77 ± 0.153 a |
| Triglycerides | 9–213 (mg/dL) | 175.10 a | 178.91 a |
| Phosphorus | 1.10–6.60 (mg/dL) | 2.287 ± 0.38 a | 1.72 ± 0.13 a |
| Urea | 7.50–28 (mg/dL) | 9.05 ± 2.69 a | 10.03 ± 0.35 a |
| Gama-glutamyl transferase | 1–199 (U/L) | 65.67 ± 24.21 a | 76 ± 36.06 a |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Canaán-Haden, C.; Sánchez-Ramírez, J.; Martínez-Castillo, R.; Bequet-Romero, M.; Puente-Pérez, P.; Gonzalez-Moya, I.; Rodríguez-Álvarez, Y.; Ayala-Ávila, M.; Castro-Velazco, J.; Cabanillas-Bernal, O.; et al. Immunogenicity and Safety Profile of Two Adjuvanted-PD-L1-Based Vaccine Candidates in Mice, Rats, Rabbits, and Cynomolgus Monkeys. Vaccines 2025, 13, 296. https://doi.org/10.3390/vaccines13030296
Canaán-Haden C, Sánchez-Ramírez J, Martínez-Castillo R, Bequet-Romero M, Puente-Pérez P, Gonzalez-Moya I, Rodríguez-Álvarez Y, Ayala-Ávila M, Castro-Velazco J, Cabanillas-Bernal O, et al. Immunogenicity and Safety Profile of Two Adjuvanted-PD-L1-Based Vaccine Candidates in Mice, Rats, Rabbits, and Cynomolgus Monkeys. Vaccines. 2025; 13(3):296. https://doi.org/10.3390/vaccines13030296
Chicago/Turabian StyleCanaán-Haden, Camila, Javier Sánchez-Ramírez, Rafael Martínez-Castillo, Mónica Bequet-Romero, Pedro Puente-Pérez, Isabel Gonzalez-Moya, Yunier Rodríguez-Álvarez, Marta Ayala-Ávila, Jorge Castro-Velazco, Olivia Cabanillas-Bernal, and et al. 2025. "Immunogenicity and Safety Profile of Two Adjuvanted-PD-L1-Based Vaccine Candidates in Mice, Rats, Rabbits, and Cynomolgus Monkeys" Vaccines 13, no. 3: 296. https://doi.org/10.3390/vaccines13030296
APA StyleCanaán-Haden, C., Sánchez-Ramírez, J., Martínez-Castillo, R., Bequet-Romero, M., Puente-Pérez, P., Gonzalez-Moya, I., Rodríguez-Álvarez, Y., Ayala-Ávila, M., Castro-Velazco, J., Cabanillas-Bernal, O., De-León-Nava, M. A., Licea-Navarro, A. F., & Morera-Díaz, Y. (2025). Immunogenicity and Safety Profile of Two Adjuvanted-PD-L1-Based Vaccine Candidates in Mice, Rats, Rabbits, and Cynomolgus Monkeys. Vaccines, 13(3), 296. https://doi.org/10.3390/vaccines13030296