
Safety Assessment of a Sublingual Vaccine Formulated with Poly(I:C) Adjuvant and Influenza HA Antigen in Mice and Macaque Monkeys: Comparison with Intranasal Vaccine
Abstract
1. Introduction
2. Materials and Methods
2.1. Reagents
2.2. Animals
2.3. Vaccination and Sampling
2.3.1. In Mice
2.3.2. In Macaque Monkeys
2.4. Blood Testing
2.5. RNA Isolation
2.6. Gene Expression Analyses Using Quantitative Reverse Transcription PCR (RT-qPCR)
2.7. Histological Examination
3. Results
3.1. Blood Testing
3.2. Gene Expression Analyses of Inflammation-Related Genes
3.2.1. In Mice
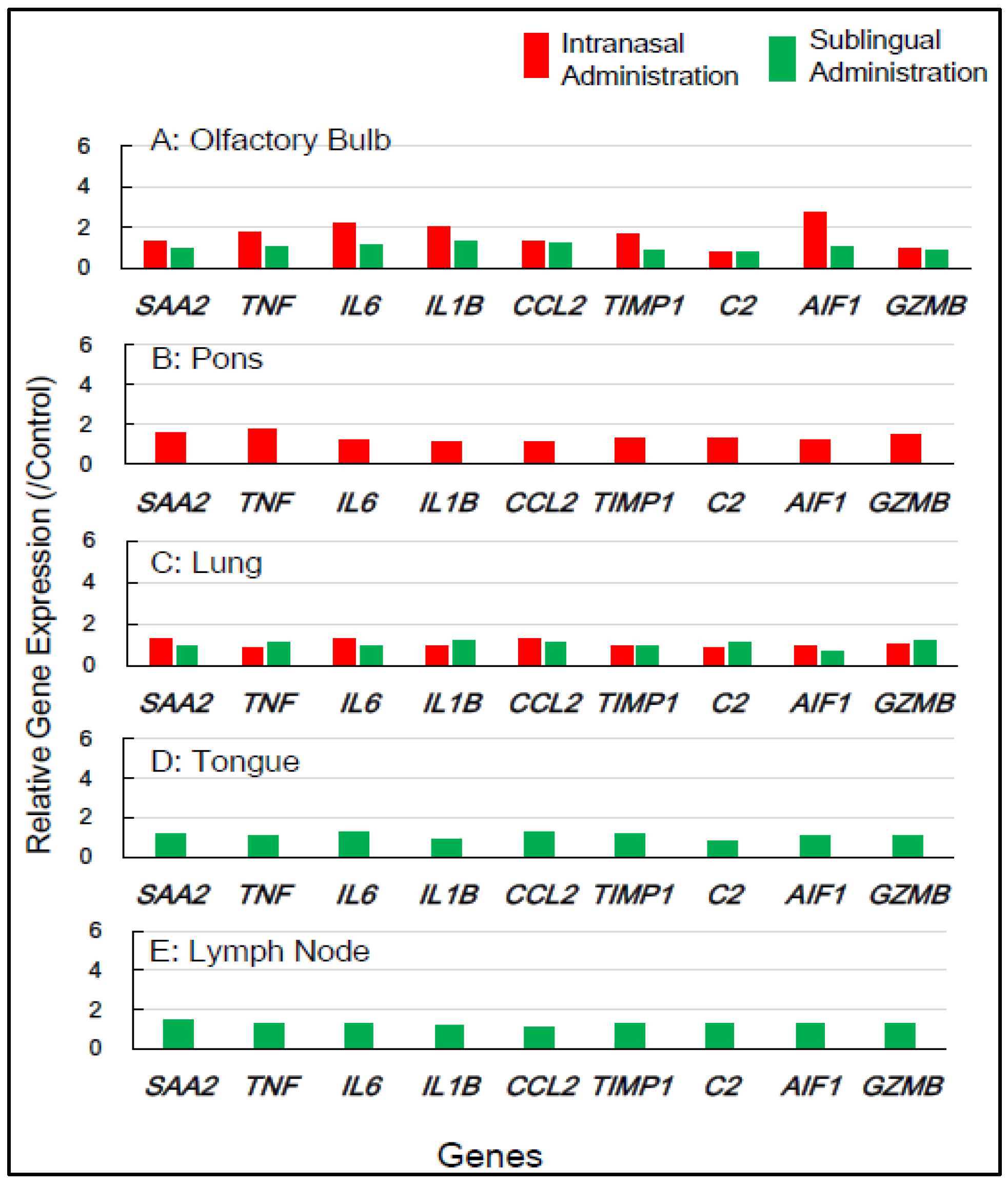
3.2.2. In Macaque Monkeys
3.3. Histological Examination
4. Discussions
4.1. Poly(I:C) Adjuvant
4.2. Safety Assessment of the Vaccine in Mice
4.3. Gene Expression Analyses for Vaccine Safety Evaluation
4.4. Use of PBWCs for Safety Assessment of Vaccines and/or Adjuvants
4.5. Safety of the Sublingual Poly(I:C)-Adjuvanted Vaccine
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Pushparajah, D.; Jimenez, S.; Wong, S.; Alattas, H.; Nafissi, N.; Slavcev, R.A. Advances in gene-based vaccine platforms to address the COVID-19 pandemic. Adv. Drug Deliv. Rev. 2021, 170, 113–141. [Google Scholar] [CrossRef] [PubMed]
- Soema, P.C.; van Riet, E.; Kersten, G.; Amorij, J.-P. Development of cross-protective influenza A vaccines based on cellular responses. Front. Immunol. 2015, 6, 237. [Google Scholar] [CrossRef] [PubMed]
- Dolgin, E. How protein-based COVID vaccines could change the pandemic. Nature 2021, 599, 359–360. [Google Scholar] [CrossRef] [PubMed]
- Pulendran, B.; Arunachalam, P.S.; O’hagan, D.T. Emerging concepts in the science of vaccine adjuvants. Nat. Rev. Drug Discov. 2021, 20, 454–475. [Google Scholar] [CrossRef]
- De Rosa, S.C.; Cohen, K.W.; Bonaparte, M.; Fu, B.; Garg, S.; Gerard, C.; Goepfert, P.A.; Huang, Y.; Larocque, D.; McElrath, M.J.; et al. Whole-blood cytokine secretion assay as a high-throughput alternative for assessing the cell-mediated immunity profile after two doses of an adjuvanted SARS-CoV-2 recombinant protein vaccine candidate. Clin. Transl. Immunol. 2022, 11, e1360. [Google Scholar] [CrossRef]
- Hafner, A.M.; Corthésy, B.; Merkle, H.P. Particulate formulations for the delivery of poly(I:C) as vaccine adjuvant. Adv. Drug Deliv. Rev. 2013, 65, 1386–1399. [Google Scholar] [CrossRef]
- Ainai, A.; van Riet, E.; Ito, R.; Ikeda, K.; Senchi, K.; Suzuki, T.; Tamura, S.I.; Asanuma, H.; Odagiri, T.; Tashiro, M.; et al. Human immune responses elicited by an intranasal inactivated H5 influenza vaccine. Microbiol. Immunol. 2020, 64, 313–325. [Google Scholar] [CrossRef]
- Mudgal, R.; Nehul, S.; Tomar, S. Prospects for mucosal vaccine: Shutting the door on SARS-CoV-2. Hum. Vaccines Immunother. 2020, 16, 2921–2931. [Google Scholar] [CrossRef]
- Ambrose, C.S.; Luke, C.; Coelingh, K. Current status of live attenuated influenza vaccine in the United States for seasonal and pandemic influenza. Influenza Other Respir. Viruses 2008, 2, 193–202. [Google Scholar] [CrossRef]
- Lemiale, F.; Kong, W.; Akyürek, L.M.; Ling, X.; Huang, Y.; Chakrabarti, B.K.; Eckhaus, M.; Nabel, G.J. Enhanced mucosal immunoglobulin A response of intranasal adenoviral vector human immunodeficiency virus vaccine and localization in the central nervous system. J. Virol. 2003, 77, 10078–10087. [Google Scholar] [CrossRef]
- Sasaki, E.; Momose, H.; Hiradate, Y.; Mizukami, T.; Hamaguchi, I. Establishment of a novel safety assessment method for vaccine adjuvant development. Vaccine 2018, 36, 7112–7118. [Google Scholar] [CrossRef] [PubMed]
- Yamamoto, T.; Tanji, M.; Mitsunaga, F.; Nakamura, S. SARS-CoV-2 sublingual vaccine with RBD antigen and poly(I:C) adjuvant: Preclinical study in cynomolgus macaques. Biol. Methods Protoc. 2023, 8, bpad017. [Google Scholar] [CrossRef] [PubMed]
- Yamamoto, T.; Mitsunaga, F.; Wasaki, K.; Kotani, A.; Tajima, K.; Tanji, M.; Nakamura, S. Mechanism Underlying the Immune Responses of Sublingual Vaccine for SARS-CoV-2 with RBD Antigen and Adjuvant, Poly(I:C) or AddaS03, in Non-human Primates. Arch. Microbiol. Immunol. 2023, 7, 150–164. [Google Scholar] [CrossRef]
- Yamamoto, T.; HIrano, M.; Mitsunaga, F.; Wasaki, K.; Kotani, A.; Tajima, K.; Nakamura, S. Molecular Events in Immune Responses to Sublingua Influenza Vaccine with Hemagglutinin Antigen and Poly(I:C) Adjuvant in Nonhuman Primates, Cynomolgus Macaques. Vaccines 2024, 12, 643. [Google Scholar] [CrossRef]
- Pillai, K.; Akhter, J.; Chua, T.C.; Morris, D.L. A formulation for in situ lysis of mucin secreted in pseudomyxoma peritonei. Int. J. Cancer 2014, 134, 478–486. [Google Scholar] [CrossRef]
- Suzuki, H.; Nagatake, T.; Nasu, A.; Lan, H.; Ikegami, K.; Setou, M.; Hamazaki, Y.; Kiyono, H.; Yagi, K.; Kondoh, M.; et al. Impaired airway mucociliary function reduces antigen-specific IgA immune response to immunization with a claudin-4-targeting nasal vaccine in mice. Sci. Rep. 2018, 8, 2904. [Google Scholar] [CrossRef]
- Ye, J.; Coulouris, G.; Zaretskaya, I.; Cutcutache, I.; Rozen, S.; Madden, T.L. Primer-BLAST: A Tool to Design Target-Specific Primers for Polymerase Chain Reaction. BMC Bioinform. 2012, 13, 134. [Google Scholar] [CrossRef]
- Gabrielsson, B.G.; Olofsson, L.E.; Sjögren, A.; Jernås, M.; Elander, A.; Lönn, M.; Rudemo, M.; Carlsson, L.M. Evaluation of Reference Genes for Studies of Gene Expression in Human Adipose Tissue. Obes. Res. 2005, 13, 649–652. [Google Scholar] [CrossRef]
- Sasaki, E.; Momose, H.; Hiradate, Y.; Furuhata, K.; Takai, M.; Asanuma, H.; Ishii, K.J.; Mizukami, T.; Hamaguchi, I. Modeling for influenza vaccines and adjuvants profile for safety prediction system using gene expression profiling and statistical tools. PLoS ONE 2018, 13, e0191896. [Google Scholar] [CrossRef]
- Sasaki, E.; Asanuma, H.; Momose, H.; Furuhata, K.; Mizukami, T.; Hamaguchi, I. Immunogenicity and Toxicity of Different Adjuvants Can Be Characterized by Profiling Lung Biomarker Genes After Nasal Immunization. Front. Immunol. 2020, 11, 2171. [Google Scholar] [CrossRef]
- van Loo, G.; Bertrand, M.J.M. Death by TNF: A road to inflammation. Nat. Rev. Immunol. 2023, 23, 289–303. [Google Scholar] [CrossRef] [PubMed]
- Tritto, E.; Muzzi, A.; Pesce, I.; Monaci, E.; Nuti, S.; Galli, G.; Wack, A.; Rappuoli, R.; Hussell, T.; De Gregorio, E. The acquired immune response to the mucosal adjuvant LTK63 imprints the mouse lung with a protective signature. J. Immunol. 2007, 179, 5346–5357. [Google Scholar] [CrossRef]
- Mulholland, B.S.; Forwood, M.R.; Morrison, N.A. Monocyte Chemoattractant Protein-1 (MCP-1/CCL2) Drives Activation of Bone Remodelling and Skeletal Metastasis. Curr. Osteoporos. Rep. 2019, 17, 538–547. [Google Scholar] [CrossRef] [PubMed]
- Yeh, C.F.; Chuang, T.-Y.; Lan, M.-Y.; Chin, Y.-C.; Wang, W.-H.; Lin, Y.-Y. Excessive Expression of Microglia/Macrophage and Proinflammatory Mediators in Olfactory Bulb and Olfactory Dysfunction After Stroke. In Vivo 2019, 33, 1893–1899. [Google Scholar] [CrossRef]
- Heneka, M.T.; Galea, E.; Gavriluyk, V.; Dumitrescu-Ozimek, L.; Daeschner, J.; O’Banion, M.K.; Weinberg, G.; Klockgether, T.; Feinstein, D.L. Noradrenergic depletion potentiates beta-amyloid-induced cortical inflammation: Implications for Alzheimer’s disease. J. Neurosci. 2002, 22, 2434–2442. [Google Scholar] [CrossRef]
- Liy, P.M.; Puzi, N.N.A.; Jose, S.; Vidyadaran, S. Nitric oxide modulation in neuroinflammation and the role of mesenchymal stem cells. Exp. Biol. Med. 2021, 246, 2399–2406. [Google Scholar] [CrossRef]
- Mitsunaga, F.; Nakamura, S. A Sensitive and Simple Method to Assess NK Cell Activity by RT-qPCR for Granzyme B Using Spleen and Blood. J. Biosci. Med. 2021, 9, 27–38. [Google Scholar] [CrossRef]
- Song, J.-H.; Nguyen, H.H.; Cuburu, N.; Horimoto, T.; Ko, S.-Y.; Park, S.-H.; Czerkinsky, C.; Kweon, M.-N. Sublingual vaccination with influenza virus protects mice against lethal viral infection. Proc. Natl. Acad. Sci. USA 2008, 105, 1644–1649. [Google Scholar] [CrossRef]
- Dotiwala, F.; Upadhyay, A.K. Next Generation Mucosal Vaccine Strategy for Respiratory Pathogens. Vaccines 2023, 11, 1585. [Google Scholar] [CrossRef]
- Mestas, J.; Hughes, C.C.W. Of Mice and Not Men: Differences between Mouse and Human Immunology. J. Immunol. 2004, 172, 2731–2738. [Google Scholar] [CrossRef]
- Longhi, M.P.; Trumpfheller, C.; Idoyaga, J.; Caskey, M.; Matos, I.; Kluger, C.; Salazar, A.M.; Colonna, M.; Steinman, R.M. Dendritic cells require a systemic type I interferon response to mature and induce CD4+ Th1 immunity with poly IC as adjuvant. J. Exp. Med. 2009, 206, 1589–1602. [Google Scholar] [CrossRef] [PubMed]
- Momose, H.; Mizukami, T.; Kuramitsu, M.; Takizawa, K.; Masumi, A.; Araki, K.; Furuhata, K.; Yamaguchi, K.; Hamaguchi, I. Establishment of a New Quality Control and Vaccine Safety Test for Influenza Vaccines and Adjuvants Using Gene Expression Profiling. PLoS ONE 2015, 10, e0124392. [Google Scholar] [CrossRef] [PubMed]
- Momose, H.; Sasaki, E.; Kuramitsu, M.; Hamaguchi, I.; Mizukami, T. Gene expression profiling toward the next generation safety control of influenza vaccines and adjuvants in Japan. Vaccine 2018, 36, 6449–6455. [Google Scholar] [CrossRef] [PubMed]
- Mutsch, M.; Zhou, W.; Rhodes, P.; Bopp, M.; Chen, R.T.; Linder, T.; Spyr, C.; Steffen, R. Use of the inactivated intranasal influenza vaccine and the risk of Bell’s palsy in Switzerland. N. Engl. J. Med. 2004, 350, 896–903. [Google Scholar] [CrossRef]
- Bertin, B.; Grenet, G.; Pizzoglio-Billaudaz, V.; Lepelley, M.; Atzenhoffer, M.; Vial, T. Vaccines and Bell’s palsy: A narrative review. Therapies 2023, 78, 279–292. [Google Scholar] [CrossRef]
- Toulgoat, F.; Sarrazin, J.; Benoudiba, F.; Pereon, Y.; Auffray-Calvier, E.; Daumas-Duport, B.; Lintia-Gaultier, A.; Desal, H. Facial nerve: From anatomy to pathology. Diagn. Interv. Imaging 2013, 94, 1033–1042. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Symbol | Gene Information Product; Description; Function [Reference] | Sample; Tissue/Site * | Time Point ** |
|---|---|---|---|
| Saa3 | Serum amyloid A 3; acute response protein [19,20] | OB, P, L, T, (S) LN | 1 d, 7 ds |
| Tnf | Tumor necrosis factor; inflammatory cytokine [21] | OB, P, L, T, (S) LN | 1 d, 7 ds |
| IL6 | Interleukin 6; immune–inflammatory response [22] | OB, P, L, T, (S) LN | 1 d, 7 ds |
| IL1b | Interleukin 1 beta; inflammatory cytokine | OB, P, L, T, (S) LN | 1 d, 7 ds |
| Ccl2 | C-C motif chemokine ligand 2(MCP1); chemokine [23] | OB, P, L, T, (S) LN | 1 d, 7 ds |
| Timp1 | Tissue inhibitor of metalloproteinase 1; tissue repairing protein [19,20] | OB, P, L, T, (S) LN | 1 d, 7 ds |
| C2 | Complement component 2; opsonic function; phagocytic cell activation [20] | OB, P, L, T, (S) LN | 1 d, 7 ds |
| Ifi47 | Interferon gamma inducible protein 47; pathogen defense protein [19,20] | OB, P, L, T, (S) LN | 1 d, 7 ds |
| Aif1 | Allograft inflammatory factor 1; microglial marker [24] | OB, P | 1 d, 7 ds |
| Omp | Olfactory marker protein; odor detection/signal transduction | OB | 1 d, 7 ds |
| Nos2 | Nitric oxide synthase 2, inducible, iNos; [25,26] | P | 1 d, 7 ds |
| Gzmb | Granzyme-B; NK cell protease; apoptosis induction [27] | L, (S) LN | 1 d, 7 ds |
| Gene Symbol | Gene Information Product; Description; Function [Reference] | Sample; Tissue/Site * | Time Point ** |
|---|---|---|---|
| SAA2 | Serum amyloid A 3; acute response protein [19,20] | OB, P, L, T, (S) LN | 7 ds |
| TNF | Tumor necrosis factor; inflammatory cytokine [21] | OB, P, L, T, (S) LN | 7 ds |
| IL6 | Interleukin 6; immune–inflammatory response [22] | OB, P, L, T, (S) LN | 7 ds |
| IL1B | Interleukin 1 beta; inflammatory cytokine | OB, P, L, T, (S) LN | 7 ds |
| CCL2 | C-C motif chemokine ligand 2 (MCP1); chemokine [23] | OB, P, L, T, (S) LN | 7 ds |
| TIMP | Tissue inhibitor of metalloproteinase 1; tissue repairing protein [19,20] | OB, P, L, T, (S) LN | 7 ds |
| C2 | Complement component 2; opsonic function; phagocytic cell activation [20] | OB, P, L, T, (S) LN | 7 ds |
| AIF1 | Allograft inflammatory factor 1; microglial marker [24] | OB, P, L, T, (S) LN | 7 ds |
| GZMB | Granzyme-B; NK cell protease; apoptosis induction [27] | OB, P, L, T, (S) LN | 7 ds |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yamamoto, T.; Mitsunaga, F.; Kotani, A.; Tajima, K.; Wasaki, K.; Nakamura, S. Safety Assessment of a Sublingual Vaccine Formulated with Poly(I:C) Adjuvant and Influenza HA Antigen in Mice and Macaque Monkeys: Comparison with Intranasal Vaccine. Vaccines 2025, 13, 261. https://doi.org/10.3390/vaccines13030261
Yamamoto T, Mitsunaga F, Kotani A, Tajima K, Wasaki K, Nakamura S. Safety Assessment of a Sublingual Vaccine Formulated with Poly(I:C) Adjuvant and Influenza HA Antigen in Mice and Macaque Monkeys: Comparison with Intranasal Vaccine. Vaccines. 2025; 13(3):261. https://doi.org/10.3390/vaccines13030261
Chicago/Turabian StyleYamamoto, Tetsuro, Fusako Mitsunaga, Atsushi Kotani, Kazuki Tajima, Kunihiko Wasaki, and Shin Nakamura. 2025. "Safety Assessment of a Sublingual Vaccine Formulated with Poly(I:C) Adjuvant and Influenza HA Antigen in Mice and Macaque Monkeys: Comparison with Intranasal Vaccine" Vaccines 13, no. 3: 261. https://doi.org/10.3390/vaccines13030261
APA StyleYamamoto, T., Mitsunaga, F., Kotani, A., Tajima, K., Wasaki, K., & Nakamura, S. (2025). Safety Assessment of a Sublingual Vaccine Formulated with Poly(I:C) Adjuvant and Influenza HA Antigen in Mice and Macaque Monkeys: Comparison with Intranasal Vaccine. Vaccines, 13(3), 261. https://doi.org/10.3390/vaccines13030261