


Updated Considerations for the Immunopharmacological Aspects of the “Talented mRNA Vaccines”

 ,
,  , ,
, ,  , and
, and
Abstract
1. Introduction: An Overview of RNA-Based Drugs
2. Methods
3. mRNA-Based Vaccine Platform
4. Pharmacokinetic Considerations of mRNA Vaccines
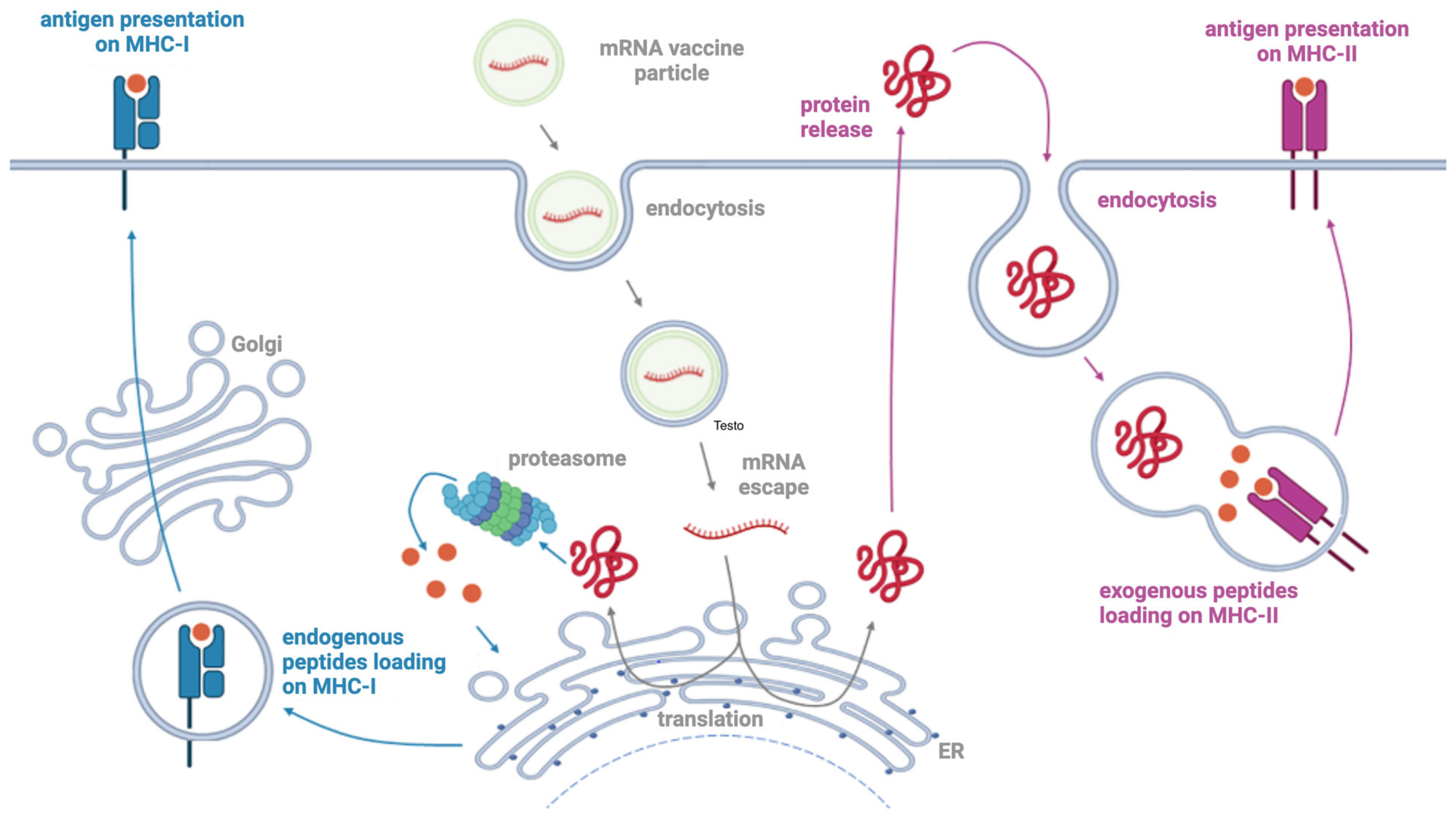
5. Mechanism of Action of mRNA Vaccines: Antigen Presentation
5.1. Innate Immunity
5.2. Adaptive Immunity
5.3. Mucosal Immunity
5.4. mRNA- vs. DNA-Based Vaccines
6. Safety Profile of mRNA Vaccines: Evidence from Clinical Trial Data and Post-Market Surveillance
6.1. Data from Post-Marketing Studies
6.2. Adverse Events of Special Interest
6.3. Care of Special Populations
6.3.1. Children and Adolescents
6.3.2. Elderly
6.3.3. Pregnancy
7. Relevant Clinical Trials on mRNA Vaccines
Phase 2/3 Trials
8. Challenges and Remarks
8.1. Lesson from Pfizer-BioNTech Comirnaty and the Moderna Spikevax
8.2. Perspectives
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Ezkurdia, I.; Juan, D.; Rodriguez, J.M.; Frankish, A.; Diekhans, M.; Harrow, J.; Vazquez, J.; Valencia, A.; Tress, M.L. Multiple Evidence Strands Suggest That There May Be as Few as 19,000 Human Protein-Coding Genes. Hum. Mol. Genet. 2014, 23, 5866–5878. [Google Scholar] [CrossRef] [PubMed]
- Hopkins, A.L.; Groom, C.R. The Druggable Genome. Nat. Rev. Drug Discov. 2002, 1, 727–730. [Google Scholar] [CrossRef] [PubMed]
- Russ, A.P.; Lampel, S. The Druggable Genome: An Update. Drug Discov. Today 2005, 10, 1607–1610. [Google Scholar] [CrossRef] [PubMed]
- Damase, T.R.; Sukhovershin, R.; Boada, C.; Taraballi, F.; Pettigrew, R.I.; Cooke, J.P. The Limitless Future of RNA Therapeutics. Front. Bioeng. Biotechnol. 2021, 9, 628137. [Google Scholar] [CrossRef]
- Qin, S.; Tang, X.; Chen, Y.; Chen, K.; Fan, N.; Xiao, W.; Zheng, Q.; Li, G.; Teng, Y.; Wu, M.; et al. MRNA-Based Therapeutics: Powerful and Versatile Tools to Combat Diseases. Signal Transduct. Target. Ther. 2022, 7, 166. [Google Scholar] [CrossRef]
- Roberts, T.C.; Langer, R.; Wood, M.J.A. Advances in Oligonucleotide Drug Delivery. Nat. Rev. Drug Discov. 2020, 19, 673–694. [Google Scholar] [CrossRef]
- Egli, M.; Manoharan, M. Chemistry, Structure and Function of Approved Oligonucleotide Therapeutics. Nucleic Acids Res. 2023, 51, 2529–2573. [Google Scholar] [CrossRef]
- Ranasinghe, P.; Addison, M.L.; Dear, J.W.; Webb, D.J. Small Interfering RNA: Discovery, Pharmacology and Clinical Development—An Introductory Review. Br. J. Pharmacol. 2022. [Google Scholar] [CrossRef]
- Loganathan, T.; Doss C, G.P. Non-Coding RNAs in Human Health and Disease: Potential Function as Biomarkers and Therapeutic Targets. Funct. Integr. Genom. 2023, 23, 33. [Google Scholar] [CrossRef]
- Vandghanooni, S.; Eskandani, M.; Barar, J.; Omidi, Y. Bispecific Therapeutic Aptamers for Targeted Therapy of Cancer: A Review on Cellular Perspective. J. Mol. Med. 2018, 96, 885–902. [Google Scholar] [CrossRef]
- Yuhan, J.; Zhu, L.; Zhu, L.; Huang, K.; He, X.; Xu, W. Cell-Specific Aptamers as Potential Drugs in Therapeutic Applications: A Review of Current Progress. J. Control. Release 2022, 346, 405–420. [Google Scholar] [CrossRef] [PubMed]
- Curreri, A.; Sankholkar, D.; Mitragotri, S.; Zhao, Z. RNA Therapeutics in the Clinic. Bioeng. Transl. Med. 2023, 8, e10374. [Google Scholar] [CrossRef] [PubMed]
- Chaudhary, N.; Weissman, D.; Whitehead, K.A. MRNA Vaccines for Infectious Diseases: Principles, Delivery and Clinical Translation. Nat. Rev. Drug Discov. 2021, 20, 817–838. [Google Scholar] [CrossRef] [PubMed]
- Barbier, A.J.; Jiang, A.Y.; Zhang, P.; Wooster, R.; Anderson, D.G. The Clinical Progress of MRNA Vaccines and Immunotherapies. Nat. Biotechnol. 2022, 40, 840–854. [Google Scholar] [CrossRef]
- Pardi, N.; Hogan, M.J.; Porter, F.W.; Weissman, D. MRNA Vaccines—A New Era in Vaccinology. Nat. Rev. Drug Discov. 2018, 17, 261–279. [Google Scholar] [CrossRef]
- Alberer, M.; Gnad-Vogt, U.; Hong, H.S.; Mehr, K.T.; Backert, L.; Finak, G.; Gottardo, R.; Bica, M.A.; Garofano, A.; Koch, S.D.; et al. Safety and Immunogenicity of a MRNA Rabies Vaccine in Healthy Adults: An Open-Label, Non-Randomised, Prospective, First-in-Human Phase 1 Clinical Trial. Lancet 2017, 390, 1511–1520. [Google Scholar] [CrossRef]
- Chahal, J.S.; Khan, O.F.; Cooper, C.L.; McPartlan, J.S.; Tsosie, J.K.; Tilley, L.D.; Sidik, S.M.; Lourido, S.; Langer, R.; Bavari, S.; et al. Dendrimer-RNA Nanoparticles Generate Protective Immunity against Lethal Ebola, H1N1 Influenza, and Toxoplasma Gondii Challenges with a Single Dose. Proc. Natl. Acad. Sci. USA 2016, 113, E4133–E4142. [Google Scholar] [CrossRef]
- Kose, N.; Fox, J.M.; Sapparapu, G.; Bombardi, R.; Tennekoon, R.N.; de Silva, A.D.; Elbashir, S.M.; Theisen, M.A.; Humphris-Narayanan, E.; Ciaramella, G.; et al. A Lipid-Encapsulated MRNA Encoding a Potently Neutralizing Human Monoclonal Antibody Protects against Chikungunya Infection. Sci. Immunol. 2019, 4, eaaw6647. [Google Scholar] [CrossRef]
- Magini, D.; Giovani, C.; Mangiavacchi, S.; Maccari, S.; Cecchi, R.; Ulmer, J.B.; De Gregorio, E.; Geall, A.J.; Brazzoli, M.; Bertholet, S. Self-Amplifying MRNA Vaccines Expressing Multiple Conserved Influenza Antigens Confer Protection against Homologous and Heterosubtypic Viral Challenge. PLoS ONE 2016, 11, e0161193. [Google Scholar] [CrossRef]
- Richner, J.M.; Himansu, S.; Dowd, K.A.; Butler, S.L.; Salazar, V.; Fox, J.M.; Julander, J.G.; Tang, W.W.; Shresta, S.; Pierson, T.C.; et al. Modified MRNA Vaccines Protect against Zika Virus Infection. Cell 2017, 168, 1114–1125.e10. [Google Scholar] [CrossRef]
- Baeza Garcia, A.; Siu, E.; Sun, T.; Exler, V.; Brito, L.; Hekele, A.; Otten, G.; Augustijn, K.; Janse, C.J.; Ulmer, J.B.; et al. Neutralization of the Plasmodium-Encoded MIF Ortholog Confers Protective Immunity against Malaria Infection. Nat. Commun. 2018, 9, 2714. [Google Scholar] [CrossRef] [PubMed]
- Maruggi, G.; Chiarot, E.; Giovani, C.; Buccato, S.; Bonacci, S.; Frigimelica, E.; Margarit, I.; Geall, A.; Bensi, G.; Maione, D. Immunogenicity and Protective Efficacy Induced by Self-Amplifying MRNA Vaccines Encoding Bacterial Antigens. Vaccine 2017, 35, 361–368. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Chen, J.; Xu, Q. Current Developments and Challenges of MRNA Vaccines. Annu. Rev. Biomed. Eng. 2022, 24, 85–109. [Google Scholar] [CrossRef]
- Miao, L.; Zhang, Y.; Huang, L. MRNA Vaccine for Cancer Immunotherapy. Mol. Cancer 2021, 20, 41. [Google Scholar] [CrossRef] [PubMed]
- He, Q.; Gao, H.; Tan, D.; Zhang, H.; Wang, J. MRNA Cancer Vaccines: Advances, Trends and Challenges. Acta Pharm. Sin. B 2022, 12, 2969–2989. [Google Scholar] [CrossRef] [PubMed]
- Liao, J.-Y.; Zhang, S. Safety and Efficacy of Personalized Cancer Vaccines in Combination with Immune Checkpoint Inhibitors in Cancer Treatment. Front. Oncol. 2021, 11, 663264. [Google Scholar] [CrossRef]
- Fiedler, K.; Lazzaro, S.; Lutz, J.; Rauch, S.; Heidenreich, R. MRNA Cancer Vaccines. Curr. Strateg. Cancer Gene Ther. 2016, 209, 61–85. [Google Scholar] [CrossRef]
- Beck, J.D.; Reidenbach, D.; Salomon, N.; Sahin, U.; Türeci, Ö.; Vormehr, M.; Kranz, L.M. MRNA Therapeutics in Cancer Immunotherapy. Mol. Cancer 2021, 20, 69. [Google Scholar] [CrossRef]
- Cafri, G.; Gartner, J.J.; Zaks, T.; Hopson, K.; Levin, N.; Paria, B.C.; Parkhurst, M.R.; Yossef, R.; Lowery, F.J.; Jafferji, M.S.; et al. MRNA Vaccine–Induced Neoantigen-Specific T Cell Immunity in Patients with Gastrointestinal Cancer. J. Clin. Investig. 2020, 130, 5976–5988. [Google Scholar] [CrossRef]
- Sahin, U.; Oehm, P.; Derhovanessian, E.; Jabulowsky, R.A.; Vormehr, M.; Gold, M.; Maurus, D.; Schwarck-Kokarakis, D.; Kuhn, A.N.; Omokoko, T.; et al. An RNA Vaccine Drives Immunity in Checkpoint-Inhibitor-Treated Melanoma. Nature 2020, 585, 107–112. [Google Scholar] [CrossRef]
- Schumacher, T.N.; Schreiber, R.D. Neoantigens in Cancer Immunotherapy. Science 2015, 348, 69–74. [Google Scholar] [CrossRef] [PubMed]
- Lorentzen, C.L.; Haanen, J.B.; Met, Ö.; Svane, I.M. Clinical Advances and Ongoing Trials of MRNA Vaccines for Cancer Treatment. Lancet Oncol. 2022, 23, e450–e458. [Google Scholar] [CrossRef] [PubMed]
- Roesler, E.; Weiss, R.; Weinberger, E.E.; Fruehwirth, A.; Stoecklinger, A.; Mostböck, S.; Ferreira, F.; Thalhamer, J.; Scheiblhofer, S. Immunize and Disappear—Safety-Optimized MRNA Vaccination with a Panel of 29 Allergens. J. Allergy Clin. Immunol. 2009, 124, 1070–1077.e11. [Google Scholar] [CrossRef]
- Weiss, R.; Scheiblhofer, S.; Roesler, E.; Ferreira, F.; Thalhamer, J. Prophylactic MRNA Vaccination against Allergy. Curr. Opin. Allergy Clin. Immunol. 2010, 10, 567–574. [Google Scholar] [CrossRef]
- Sahin, U.; Karikó, K.; Türeci, Ö. MRNA-Based Therapeutics—Developing a New Class of Drugs. Nat. Rev. Drug Discov. 2014, 13, 759–780. [Google Scholar] [CrossRef] [PubMed]
- Lee, K.; Kim, M.; Seo, Y.; Lee, H. Development of MRNA Vaccines and Their Prophylactic and Therapeutic Applications. Nano Res. 2018, 11, 5173–5192. [Google Scholar] [CrossRef]
- Li, M.; Wang, Z.; Xie, C.; Xia, X. Advances in MRNA Vaccines. In International Review of Cell and Molecular Biology; Academic Press: Cambridge, MA, USA, 2022; Volume 372, pp. 295–316. [Google Scholar]
- Wu, Z.; Li, T. Nanoparticle-Mediated Cytoplasmic Delivery of Messenger RNA Vaccines: Challenges and Future Perspectives. Pharm. Res. 2021, 38, 473–478. [Google Scholar] [CrossRef] [PubMed]
- Son, S.; Lee, K. Development of MRNA Vaccines/Therapeutics and Their Delivery System. Mol. Cells 2023, 46, 41–47. [Google Scholar] [CrossRef]
- Mirtaleb, M.S.; Falak, R.; Heshmatnia, J.; Bakhshandeh, B.; Taheri, R.A.; Soleimanjahi, H.; Zolfaghari Emameh, R. An Insight Overview on COVID-19 MRNA Vaccines: Advantageous, Pharmacology, Mechanism of Action, and Prospective Considerations. Int. Immunopharmacol. 2023, 117, 109934. [Google Scholar] [CrossRef]
- Herkt, M.; Thum, T. Pharmacokinetics and Proceedings in Clinical Application of Nucleic Acid Therapeutics. Mol. Ther. 2021, 29, 521–539. [Google Scholar] [CrossRef]
- Vervaeke, P.; Borgos, S.E.; Sanders, N.N.; Combes, F. Regulatory Guidelines and Preclinical Tools to Study the Biodistribution of RNA Therapeutics. Adv. Drug Deliv. Rev. 2022, 184, 114236. [Google Scholar] [CrossRef] [PubMed]
- Marlowe, J.L.; Akopian, V.; Karmali, P.; Kornbrust, D.; Lockridge, J.; Semple, S. Recommendations of the Oligonucleotide Safety Working Group’s Formulated Oligonucleotide Subcommittee for the Safety Assessment of Formulated Oligonucleotide-Based Therapeutics. Nucleic Acid. Ther. 2017, 27, 183–196. [Google Scholar] [CrossRef] [PubMed]
- Bortolotti, D.; Gentili, V.; Rizzo, S.; Schiuma, G.; Beltrami, S.; Strazzabosco, G.; Fernandez, M.; Caccuri, F.; Caruso, A.; Rizzo, R. TLR3 and TLR7 RNA Sensor Activation during SARS-CoV-2 Infection. Microorganisms 2021, 9, 1820. [Google Scholar] [CrossRef]
- Fu, L.; Gilham, D.; Stotz, S.C.; Sarsons, C.D.; Rakai, B.D.; Tsujikawa, L.M.; Wasiak, S.; Johansson, J.O.; Sweeney, M.; Wong, N.C.W.; et al. Dual Mechanism: Epigenetic Inhibitor Apabetalone Reduces SARS-CoV-2 Delta and Omicron Variant Spike Binding and Attenuates SARS-CoV-2 RNA Induced Inflammation. Int. Immunopharmacol. 2023, 117, 109929. [Google Scholar] [CrossRef] [PubMed]
- Tregoning, J.S.; Stirling, D.C.; Wang, Z.; Flight, K.E.; Brown, J.C.; Blakney, A.K.; McKay, P.F.; Cunliffe, R.F.; Murugaiah, V.; Fox, C.B.; et al. Formulation, Inflammation, and RNA Sensing Impact the Immunogenicity of Self-Amplifying RNA Vaccines. Mol. Ther. Nucleic Acids 2023, 31, 29–42. [Google Scholar] [CrossRef] [PubMed]
- Mulligan, M.J.; Lyke, K.E.; Kitchin, N.; Absalon, J.; Gurtman, A.; Lockhart, S.; Neuzil, K.; Raabe, V.; Bailey, R.; Swanson, K.A.; et al. Phase I/II Study of COVID-19 RNA Vaccine BNT162b1 in Adults. Nature 2020, 586, 589–593. [Google Scholar] [CrossRef]
- Sahin, U.; Muik, A.; Derhovanessian, E.; Vogler, I.; Kranz, L.M.; Vormehr, M.; Baum, A.; Pascal, K.; Quandt, J.; Maurus, D.; et al. COVID-19 Vaccine BNT162b1 Elicits Human Antibody and TH1 T Cell Responses. Nature 2020, 586, 594–599. [Google Scholar] [CrossRef]
- Jackson, L.A.; Anderson, E.J.; Rouphael, N.G.; Roberts, P.C.; Makhene, M.; Coler, R.N.; McCullough, M.P.; Chappell, J.D.; Denison, M.R.; Stevens, L.J.; et al. An MRNA Vaccine against SARS-CoV-2—Preliminary Report. N. Engl. J. Med. 2020, 383, 1920–1931. [Google Scholar] [CrossRef]
- Grogan, S.; Preuss, C.V. Pharmacokinetics; StatPearls Publishing: Treasure Island, FL, USA, 2023. [Google Scholar]
- Malangu, N. Introductory Chapter: Linkages between Pharmacokinetics and Adverse Effects of Drugs. In Pharmacokinetics and Adverse Effects of Drugs—Mechanisms and Risks Factors; InTechopen: London, UK, 2018. [Google Scholar]
- Plitnick, L.M. Global Regulatory Guidelines for Vaccines. In Nonclinical Development of Novel Biologics, Biosimilars, Vaccines and Specialty Biologics; Elsevier: Amsterdam, The Netherlands, 2013; pp. 225–241. [Google Scholar]
- Ingle, R.G.; Fang, W.-J. An Overview of the Stability and Delivery Challenges of Commercial Nucleic Acid Therapeutics. Pharmaceutics 2023, 15, 1158. [Google Scholar] [CrossRef]
- Hou, X.; Zaks, T.; Langer, R.; Dong, Y. Lipid Nanoparticles for MRNA Delivery. Nat. Rev. Mater. 2021, 6, 1078–1094. [Google Scholar] [CrossRef]
- Naasani, I. Establishing the Pharmacokinetics of Genetic Vaccines Is Essential for Maximising Their Safety and Efficacy. Clin. Pharmacokinet. 2022, 61, 921–927. [Google Scholar] [CrossRef] [PubMed]
- Cosentino, M.; Marino, F. Understanding the Pharmacology of COVID-19 MRNA Vaccines: Playing Dice with the Spike? Int. J. Mol. Sci. 2022, 23, 10881. [Google Scholar] [CrossRef] [PubMed]
- Kozak, M.; Hu, J. The Integrated Consideration of Vaccine Platforms, Adjuvants, and Delivery Routes for Successful Vaccine Development. Vaccines 2023, 11, 695. [Google Scholar] [CrossRef]
- Zuckerman, J.N. The Importance of Injecting Vaccines into Muscle. BMJ 2000, 321, 1237–1238. [Google Scholar] [CrossRef]
- Kim, J.; De Jesus, O. Medication Routes of Administration; StatPearls Publishing: Treasure Island, FL, USA, 2023. [Google Scholar]
- Nitika; Wei, J.; Hui, A.-M. The Delivery of MRNA Vaccines for Therapeutics. Life 2022, 12, 1254. [Google Scholar] [CrossRef] [PubMed]
- Baldwin, J.; Piplani, S.; Sakala, I.G.; Honda-Okubo, Y.; Li, L.; Petrovsky, N. Rapid Development of Analytical Methods for Evaluating Pandemic Vaccines: A COVID-19 Perspective. Bioanalysis 2021, 13, 1805–1826. [Google Scholar] [CrossRef]
- Pardi, N.; Tuyishime, S.; Muramatsu, H.; Kariko, K.; Mui, B.L.; Tam, Y.K.; Madden, T.D.; Hope, M.J.; Weissman, D. Expression Kinetics of Nucleoside-Modified MRNA Delivered in Lipid Nanoparticles to Mice by Various Routes. J. Control. Release 2015, 217, 345–351. [Google Scholar] [CrossRef]
- Fertig, T.E.; Chitoiu, L.; Marta, D.S.; Ionescu, V.-S.; Cismasiu, V.B.; Radu, E.; Angheluta, G.; Dobre, M.; Serbanescu, A.; Hinescu, M.E.; et al. Vaccine MRNA Can Be Detected in Blood at 15 Days Post-Vaccination. Biomedicines 2022, 10, 1538. [Google Scholar] [CrossRef]
- Ogata, A.F.; Cheng, C.-A.; Desjardins, M.; Senussi, Y.; Sherman, A.C.; Powell, M.; Novack, L.; Von, S.; Li, X.; Baden, L.R.; et al. Circulating Severe Acute Respiratory Syndrome Coronavirus 2 (SARS-CoV-2) Vaccine Antigen Detected in the Plasma of MRNA-1273 Vaccine Recipients. Clin. Infect. Dis. 2022, 74, 715–718. [Google Scholar] [CrossRef]
- Castruita, J.A.S.; Schneider, U.V.; Mollerup, S.; Leineweber, T.D.; Weis, N.; Bukh, J.; Pedersen, M.S.; Westh, H. SARS-CoV-2 Spike mRNA Vaccine Sequences Circulate in Blood up to 28 Days after COVID-19 Vaccination. Apmis 2023, 131, 128–132. [Google Scholar] [CrossRef]
- Yoon, B.K.; Oh, T.G.; Bu, S.; Seo, K.J.; Kwon, S.H.; Lee, J.Y.; Kim, Y.; Kim, J.; Ahn, H.-S.; Fang, S. The Peripheral Immune Landscape in a Patient with Myocarditis after the Administration of BNT162b2 MRNA Vaccine. Mol. Cells 2022, 45, 738–748. [Google Scholar] [CrossRef]
- Verbeke, R.; Hogan, M.J.; Loré, K.; Pardi, N. Innate Immune Mechanisms of MRNA Vaccines. Immunity 2022, 55, 1993–2005. [Google Scholar] [CrossRef] [PubMed]
- Tahtinen, S.; Tong, A.-J.; Himmels, P.; Oh, J.; Paler-Martinez, A.; Kim, L.; Wichner, S.; Oei, Y.; McCarron, M.J.; Freund, E.C.; et al. IL-1 and IL-1ra Are Key Regulators of the Inflammatory Response to RNA Vaccines. Nat. Immunol. 2022, 23, 532–542. [Google Scholar] [CrossRef] [PubMed]
- Dilliard, S.A.; Cheng, Q.; Siegwart, D.J. On the Mechanism of Tissue-Specific MRNA Delivery by Selective Organ Targeting Nanoparticles. Proc. Natl. Acad. Sci. USA 2021, 118, e2109256118. [Google Scholar] [CrossRef]
- Lenart, K.; Hellgren, F.; Ols, S.; Yan, X.; Cagigi, A.; Cerveira, R.A.; Winge, I.; Hanczak, J.; Mueller, S.O.; Jasny, E.; et al. A Third Dose of the Unmodified COVID-19 MRNA Vaccine CVnCoV Enhances Quality and Quantity of Immune Responses. Mol. Ther. Methods Clin. Dev. 2022, 27, 309–323. [Google Scholar] [CrossRef] [PubMed]
- Karikó, K.; Buckstein, M.; Ni, H.; Weissman, D. Suppression of RNA Recognition by Toll-like Receptors: The Impact of Nucleoside Modification and the Evolutionary Origin of RNA. Immunity 2005, 23, 165–175. [Google Scholar] [CrossRef]
- Li, C.; Lee, A.; Grigoryan, L.; Arunachalam, P.S.; Scott, M.K.D.; Trisal, M.; Wimmers, F.; Sanyal, M.; Weidenbacher, P.A.; Feng, Y.; et al. Mechanisms of Innate and Adaptive Immunity to the Pfizer-BioNTech BNT162b2 Vaccine. Nat. Immunol. 2022, 23, 543–555. [Google Scholar] [CrossRef]
- Salleh, M.Z.; Norazmi, M.N.; Deris, Z.Z. Immunogenicity Mechanism of MRNA Vaccines and Their Limitations in Promoting Adaptive Protection against SARS-CoV-2. PeerJ 2022, 10, e13083. [Google Scholar] [CrossRef]
- Sette, A.; Crotty, S. Adaptive Immunity to SARS-CoV-2 and COVID-19. Cell 2021, 184, 861–880. [Google Scholar] [CrossRef]
- Vogel, A.B.; Kanevsky, I.; Che, Y.; Swanson, K.A.; Muik, A.; Vormehr, M.; Kranz, L.M.; Walzer, K.C.; Hein, S.; Güler, A.; et al. BNT162b Vaccines Protect Rhesus Macaques from SARS-CoV-2. Nature 2021, 592, 283–289. [Google Scholar] [CrossRef]
- Nakayama, T.; Sawada, A.; Ito, T. Comparison of cytokine production in mice inoculated with messenger RNA vaccines BNT162b2 and mRNA-1273. Microbiol. Immunol. 2023, 67, 120–128. [Google Scholar] [CrossRef] [PubMed]
- Corbett, K.S.; Flynn, B.; Foulds, K.E.; Francica, J.R.; Boyoglu-Barnum, S.; Werner, A.P.; Flach, B.; O’Connell, S.; Bock, K.W.; Minai, M.; et al. Evaluation of the MRNA-1273 Vaccine against SARS-CoV-2 in Nonhuman Primates. N. Engl. J. Med. 2020, 383, 1544–1555. [Google Scholar] [CrossRef] [PubMed]
- Baden, L.R.; El Sahly, H.M.; Essink, B.; Kotloff, K.; Frey, S.; Novak, R.; Diemert, D.; Spector, S.A.; Rouphael, N.; Creech, C.B.; et al. Efficacy and Safety of the MRNA-1273 SARS-CoV-2 Vaccine. N. Engl. J. Med. 2021, 384, 403–416. [Google Scholar] [CrossRef] [PubMed]
- Andrews, N.; Stowe, J.; Kirsebom, F.; Toffa, S.; Rickeard, T.; Gallagher, E.; Gower, C.; Kall, M.; Groves, N.; O’Connell, A.-M.; et al. Covid-19 Vaccine Effectiveness against the Omicron (B.1.1.529) Variant. N. Engl. J. Med. 2022, 386, 1532–1546. [Google Scholar] [CrossRef] [PubMed]
- Fenizia, C.; Cetin, I.; Mileto, D.; Vanetti, C.; Saulle, I.; Di Giminiani, M.; Saresella, M.; Parisi, F.; Trabattoni, D.; Clerici, M.; et al. Pregnant Women Develop a Specific Immunological Long-Lived Memory against SARS-COV-2. Front. Immunol. 2022, 13, 827889. [Google Scholar] [CrossRef]
- Pulliam, J.R.C.; van Schalkwyk, C.; Govender, N.; von Gottberg, A.; Cohen, C.; Groome, M.J.; Dushoff, J.; Mlisana, K.; Moultrie, H. Increased Risk of SARS-CoV-2 Reinfection Associated with Emergence of Omicron in South Africa. Science 2022, 376, eabn4947. [Google Scholar] [CrossRef]
- Garcia-Beltran, W.F.; St. Denis, K.J.; Hoelzemer, A.; Lam, E.C.; Nitido, A.D.; Sheehan, M.L.; Berrios, C.; Ofoman, O.; Chang, C.C.; Hauser, B.M.; et al. MRNA-Based COVID-19 Vaccine Boosters Induce Neutralizing Immunity against SARS-CoV-2 Omicron Variant. Cell 2022, 185, 457–466.e4. [Google Scholar] [CrossRef]
- Mileto, D.; Micheli, V.; Fenizia, C.; Cutrera, M.; Gagliardi, G.; Mancon, A.; Bracchitta, F.; De Silvestri, A.; Rizzardini, G.; Lombardi, A.; et al. Reduced Neutralization of SARS-CoV-2 Omicron Variant by BNT162b2 Vaccinees’ Sera: A Preliminary Evaluation. Emerg. Microbes Infect. 2022, 11, 790–792. [Google Scholar] [CrossRef]
- Mileto, D.; Fenizia, C.; Cutrera, M.; Gagliardi, G.; Gigantiello, A.; De Silvestri, A.; Rizzo, A.; Mancon, A.; Bianchi, M.; De Poli, F.; et al. SARS-CoV-2 MRNA Vaccine BNT162b2 Triggers a Consistent Cross-Variant Humoral and Cellular Response. Emerg. Microbes Infect. 2021, 10, 2235–2243. [Google Scholar] [CrossRef]
- Hvidt, A.K.; Baerends, E.A.M.; Søgaard, O.S.; Stærke, N.B.; Raben, D.; Reekie, J.; Nielsen, H.; Johansen, I.S.; Wiese, L.; Benfield, T.L.; et al. Comparison of Vaccine-Induced Antibody Neutralization against SARS-CoV-2 Variants of Concern Following Primary and Booster Doses of COVID-19 Vaccines. Front. Med. 2022, 9, 994160. [Google Scholar] [CrossRef]
- Laczkó, D.; Hogan, M.J.; Toulmin, S.A.; Hicks, P.; Lederer, K.; Gaudette, B.T.; Castaño, D.; Amanat, F.; Muramatsu, H.; Oguin, T.H.; et al. A Single Immunization with Nucleoside-Modified MRNA Vaccines Elicits Strong Cellular and Humoral Immune Responses against SARS-CoV-2 in Mice. Immunity 2020, 53, 724–732.e7. [Google Scholar] [CrossRef] [PubMed]
- Garziano, M.; Utyro, O.; Poliseno, M.; Santantonio, T.A.; Saulle, I.; Strizzi, S.; Lo Caputo, S.; Clerici, M.; Introini, A.; Biasin, M. Natural SARS-CoV-2 Infection Affects Neutralizing Activity in Saliva of Vaccinees. Front. Immunol. 2022, 13, 820250. [Google Scholar] [CrossRef] [PubMed]
- Pilapitiya, D.; Wheatley, A.K.; Tan, H.-X. Mucosal Vaccines for SARS-CoV-2: Triumph of Hope over Experience. EBioMedicine 2023, 92, 104585. [Google Scholar] [CrossRef] [PubMed]
- Azzi, L.; Dalla Gasperina, D.; Veronesi, G.; Shallak, M.; Maurino, V.; Baj, A.; Gianfagna, F.; Cavallo, P.; Dentali, F.; Tettamanti, L.; et al. Mucosal Immune Response after the Booster Dose of the BNT162b2 COVID-19 Vaccine. EBioMedicine 2023, 88, 104435. [Google Scholar] [CrossRef] [PubMed]
- Sunagar, R.; Singh, A.; Kumar, S. SARS-CoV-2: Immunity, Challenges with Current Vaccines, and a Novel Perspective on Mucosal Vaccines. Vaccines 2023, 11, 849. [Google Scholar] [CrossRef] [PubMed]
- Tang, J.; Zeng, C.; Cox, T.M.; Li, C.; Son, Y.M.; Cheon, I.S.; Wu, Y.; Behl, S.; Taylor, J.J.; Chakaraborty, R.; et al. Respiratory Mucosal Immunity against SARS-CoV-2 after MRNA Vaccination. Sci. Immunol. 2022, 7, eadd4853. [Google Scholar] [CrossRef]
- Houston, S. SARS-CoV-2 Mucosal Vaccine. Nat. Immunol. 2023, 24, 1. [Google Scholar] [CrossRef]
- Jackson, N.A.C.; Kester, K.E.; Casimiro, D.; Gurunathan, S.; DeRosa, F. The Promise of MRNA Vaccines: A Biotech and Industrial Perspective. NPJ Vaccines 2020, 5, 11. [Google Scholar] [CrossRef]
- Iavarone, C.; O’hagan, D.T.; Yu, D.; Delahaye, N.F.; Ulmer, J.B. Mechanism of Action of MRNA-Based Vaccines. Expert. Rev. Vaccines 2017, 16, 871–881. [Google Scholar] [CrossRef]
- Xu, S.; Yang, K.; Li, R.; Zhang, L. MRNA Vaccine Era—Mechanisms, Drug Platform and Clinical Prospection. Int. J. Mol. Sci. 2020, 21, 6582. [Google Scholar] [CrossRef]
- Gote, V.; Bolla, P.K.; Kommineni, N.; Butreddy, A.; Nukala, P.K.; Palakurthi, S.S.; Khan, W. A Comprehensive Review of MRNA Vaccines. Int. J. Mol. Sci. 2023, 24, 2700. [Google Scholar] [CrossRef] [PubMed]
- Polack, F.P.; Thomas, S.J.; Kitchin, N.; Absalon, J.; Gurtman, A.; Lockhart, S.; Perez, J.L.; Pérez Marc, G.; Moreira, E.D.; Zerbini, C.; et al. Safety and Efficacy of the BNT162b2 MRNA Covid-19 Vaccine. N. Engl. J. Med. 2020, 383, 2603–2615. [Google Scholar] [CrossRef] [PubMed]
- Theofilopoulos, A.N.; Baccala, R.; Beutler, B.; Kono, D.H. Type I interferons (α/β) in immunity and autoimmunity. Annu. Rev. Immunol. 2005, 23, 307–335. [Google Scholar] [CrossRef] [PubMed]
- Kannemeier, C.; Shibamiya, A.; Nakazawa, F.; Trusheim, H.; Ruppert, C.; Markart, P.; Song, Y.; Tzima, E.; Kennerknecht, E.; Niepmann, M.; et al. Extracellular RNA Constitutes a Natural Procoagulant Cofactor in Blood Coagulation. Proc. Natl. Acad. Sci. USA 2007, 104, 6388–6393. [Google Scholar] [CrossRef] [PubMed]
- Walsh, E.E.; Frenck, R.W.; Falsey, A.R.; Kitchin, N.; Absalon, J.; Gurtman, A.; Lockhart, S.; Neuzil, K.; Mulligan, M.J.; Bailey, R.; et al. Safety and Immunogenicity of Two RNA-Based COVID-19 Vaccine Candidates. N. Engl. J. Med. 2020, 383, 2439–2450. [Google Scholar] [CrossRef] [PubMed]
- Mahase, E. COVID-19: Pfizer and BioNTech Submit Vaccine for US Authorisation. BMJ 2020, 371, m4552. [Google Scholar] [CrossRef]
- Shimabukuro, T.T.; Cole, M.; Su, J.R. Reports of Anaphylaxis After Receipt of MRNA COVID-19 Vaccines in the US—December 14, 2020-January 18, 2021. JAMA 2021, 325, 1101. [Google Scholar] [CrossRef]
- McNeil, M.M.; Weintraub, E.S.; Duffy, J.; Sukumaran, L.; Jacobsen, S.J.; Klein, N.P.; Hambidge, S.J.; Lee, G.M.; Jackson, L.A.; Irving, S.A.; et al. Risk of Anaphylaxis after Vaccination in Children and Adults. J. Allergy Clin. Immunol. 2016, 137, 868–878. [Google Scholar] [CrossRef]
- Besin, G.; Milton, J.; Sabnis, S.; Howell, R.; Mihai, C.; Burke, K.; Benenato, K.E.; Stanton, M.; Smith, P.; Senn, J.; et al. Accelerated Blood Clearance of Lipid Nanoparticles Entails a Biphasic Humoral Response of B-1 Followed by B-2 Lymphocytes to Distinct Antigenic Moieties. Immunohorizons 2019, 3, 282–293. [Google Scholar] [CrossRef]
- Kozma, G.T.; Shimizu, T.; Ishida, T.; Szebeni, J. Anti-PEG Antibodies: Properties, Formation, Testing and Role in Adverse Immune Reactions to PEGylated Nano-Biopharmaceuticals. Adv. Drug Deliv. Rev. 2020, 154–155, 163–175. [Google Scholar] [CrossRef]
- Manolis, A.S.; Manolis, T.A.; Manolis, A.A.; Melita, H. Cardiovascular Implications and Complications of the Coronavirus Disease-2019 Pandemic: A World Upside Down. Curr. Opin. Cardiol. 2021, 36, 241–251. [Google Scholar] [CrossRef] [PubMed]
- Tarawneh, O.; Tarawneh, H. Immune Thrombocytopenia in a 22-year-old Post COVID-19 Vaccine. Am. J. Hematol. 2021, 96, E133–E134. [Google Scholar] [CrossRef] [PubMed]
- Carli, G.; Nichele, I.; Ruggeri, M.; Barra, S.; Tosetto, A. Deep Vein Thrombosis (DVT) Occurring Shortly after the Second Dose of MRNA SARS-CoV-2 Vaccine. Intern. Emerg. Med. 2021, 16, 803–804. [Google Scholar] [CrossRef] [PubMed]
- Bhan, C.; Bheesham, N.; Shakuntulla, F.; Sharma, M.; Sun, C.; Weinstein, M. An Unusual Presentation of Acute Deep Vein Thrombosis after the Moderna COVID-19 Vaccine—A Case Report. Ann. Transl. Med. 2021, 9, 1605. [Google Scholar] [CrossRef]
- Malayala, S.V.; Mohan, G.; Vasireddy, D.; Atluri, P. Purpuric Rash and Thrombocytopenia After the MRNA-1273 (Moderna) COVID-19 Vaccine. Cureus 2021, 13, e14099. [Google Scholar] [CrossRef]
- Terrell, D.R.; Beebe, L.A.; Vesely, S.K.; Neas, B.R.; Segal, J.B.; George, J.N. The Incidence of Immune Thrombocytopenic Purpura in Children and Adults: A Critical Review of Published Reports. Am. J. Hematol. 2010, 85, 174–180. [Google Scholar] [CrossRef]
- Brazete, C.; Aguiar, A.; Furtado, I.; Duarte, R. Thrombotic Events and COVID-19 Vaccines. Int. J. Tuberc. Lung Dis. 2021, 25, 701–707. [Google Scholar] [CrossRef]
- Montano, D. Frequency and Associations of Adverse Reactions of COVID-19 Vaccines Reported to Pharmacovigilance Systems in the European Union and the United States. Front. Public Health 2022, 9, 756633. [Google Scholar] [CrossRef]
- Lai, F.T.T.; Li, X.; Peng, K.; Huang, L.; Ip, P.; Tong, X.; Chui, C.S.L.; Wan, E.Y.F.; Wong, C.K.H.; Chan, E.W.Y.; et al. Carditis After COVID-19 Vaccination with a Messenger RNA Vaccine and an Inactivated Virus Vaccine. Ann. Intern. Med. 2022, 175, 362–370. [Google Scholar] [CrossRef]
- Patone, M.; Mei, X.W.; Handunnetthi, L.; Dixon, S.; Zaccardi, F.; Shankar-Hari, M.; Watkinson, P.; Khunti, K.; Harnden, A.; Coupland, C.A.C.; et al. Risks of Myocarditis, Pericarditis, and Cardiac Arrhythmias Associated with COVID-19 Vaccination or SARS-CoV-2 Infection. Nat. Med. 2022, 28, 410–422. [Google Scholar] [CrossRef]
- Husby, A.; Hansen, J.V.; Fosbøl, E.; Thiesson, E.M.; Madsen, M.; Thomsen, R.W.; Sørensen, H.T.; Andersen, M.; Wohlfahrt, J.; Gislason, G.; et al. SARS-CoV-2 Vaccination and Myocarditis or Myopericarditis: Population Based Cohort Study. BMJ 2021, 375, e068665. [Google Scholar] [CrossRef]
- Karlstad, Ø.; Hovi, P.; Husby, A.; Härkänen, T.; Selmer, R.M.; Pihlström, N.; Hansen, J.V.; Nohynek, H.; Gunnes, N.; Sundström, A.; et al. SARS-CoV-2 Vaccination and Myocarditis in a Nordic Cohort Study of 23 Million Residents. JAMA Cardiol. 2022, 7, 600. [Google Scholar] [CrossRef]
- Mevorach, D.; Anis, E.; Cedar, N.; Bromberg, M.; Haas, E.J.; Nadir, E.; Olsha-Castell, S.; Arad, D.; Hasin, T.; Levi, N.; et al. Myocarditis after BNT162b2 MRNA Vaccine against COVID-19 in Israel. N. Engl. J. Med. 2021, 385, 2140–2149. [Google Scholar] [CrossRef]
- Witberg, G.; Barda, N.; Hoss, S.; Richter, I.; Wiessman, M.; Aviv, Y.; Grinberg, T.; Auster, O.; Dagan, N.; Balicer, R.D.; et al. Myocarditis after Covid-19 Vaccination in a Large Health Care Organization. N. Engl. J. Med. 2021, 385, 2132–2139. [Google Scholar] [CrossRef]
- Montgomery, J.; Ryan, M.; Engler, R.; Hoffman, D.; McClenathan, B.; Collins, L.; Loran, D.; Hrncir, D.; Herring, K.; Platzer, M.; et al. Myocarditis Following Immunization with MRNA COVID-19 Vaccines in Members of the US Military. JAMA Cardiol. 2021, 6, 1202. [Google Scholar] [CrossRef] [PubMed]
- Oster, M.E.; Shay, D.K.; Su, J.R.; Gee, J.; Creech, C.B.; Broder, K.R.; Edwards, K.; Soslow, J.H.; Dendy, J.M.; Schlaudecker, E.; et al. Myocarditis Cases Reported After MRNA-Based COVID-19 Vaccination in the US From December 2020 to August 2021. JAMA 2022, 327, 331. [Google Scholar] [CrossRef] [PubMed]
- Krug, A.; Stevenson, J.; Høeg, T.B. BNT162b2 Vaccine-Associated Myo/Pericarditis in Adolescents: A Stratified Risk-Benefit Analysis. Eur. J. Clin. Investig. 2022, 52, e13759. [Google Scholar] [CrossRef] [PubMed]
- Wong, H.-L.; Hu, M.; Zhou, C.K.; Lloyd, P.C.; Amend, K.L.; Beachler, D.C.; Secora, A.; McMahill-Walraven, C.N.; Lu, Y.; Wu, Y.; et al. Risk of Myocarditis and Pericarditis after the COVID-19 MRNA Vaccination in the USA: A Cohort Study in Claims Databases. Lancet 2022, 399, 2191–2199. [Google Scholar] [CrossRef]
- Ling, R.R.; Ramanathan, K.; Tan, F.L.; Tai, B.C.; Somani, J.; Fisher, D.; MacLaren, G. Myopericarditis Following COVID-19 Vaccination and Non-COVID-19 Vaccination: A Systematic Review and Meta-Analysis. Lancet Respir. Med. 2022, 10, 679–688. [Google Scholar] [CrossRef]
- Nave, L.; Margalit, I.; Tau, N.; Cohen, I.; Yelin, D.; Lienert, F.; Yahav, D. Immunogenicity and Safety of Modified Vaccinia Ankara (MVA) Vaccine—A Systematic Review and Meta-Analysis of Randomized Controlled Trials. Vaccines 2023, 11, 1410. [Google Scholar] [CrossRef]
- Tian, F.; Yang, R.; Chen, Z. Safety and Efficacy of COVID-19 Vaccines in Children and Adolescents: A Systematic Review of Randomized Controlled Trials. J. Med. Virol. 2022, 94, 4644–4653. [Google Scholar] [CrossRef]
- Frenck, R.W.; Klein, N.P.; Kitchin, N.; Gurtman, A.; Absalon, J.; Lockhart, S.; Perez, J.L.; Walter, E.B.; Senders, S.; Bailey, R.; et al. Safety, Immunogenicity, and Efficacy of the BNT162b2 COVID-19 Vaccine in Adolescents. N. Engl. J. Med. 2021, 385, 239–250. [Google Scholar] [CrossRef]
- Walter, E.B.; Talaat, K.R.; Sabharwal, C.; Gurtman, A.; Lockhart, S.; Paulsen, G.C.; Barnett, E.D.; Muñoz, F.M.; Maldonado, Y.; Pahud, B.A.; et al. Evaluation of the BNT162b2 COVID-19 Vaccine in Children 5 to 11 Years of Age. N. Engl. J. Med. 2022, 386, 35–46. [Google Scholar] [CrossRef]
- Ali, K.; Berman, G.; Zhou, H.; Deng, W.; Faughnan, V.; Coronado-Voges, M.; Ding, B.; Dooley, J.; Girard, B.; Hillebrand, W.; et al. Evaluation of MRNA-1273 SARS-CoV-2 Vaccine in Adolescents. N. Engl. J. Med. 2021, 385, 2241–2251. [Google Scholar] [CrossRef]
- Watanabe, A.; Kani, R.; Iwagami, M.; Takagi, H.; Yasuhara, J.; Kuno, T. Assessment of Efficacy and Safety of MRNA COVID-19 Vaccines in Children Aged 5 to 11 Years. JAMA Pediatr. 2023, 177, 384. [Google Scholar] [CrossRef]
- Zhang, L.; Jiang, L.; Tian, T.; Li, W.; Pan, Y.; Wang, Y. Efficacy and Safety of COVID-19 Vaccination in Older Adults: A Systematic Review and Meta-Analysis. Vaccines 2022, 11, 33. [Google Scholar] [CrossRef]
- Wong, H.-L.; Tworkoski, E.; Ke Zhou, C.; Hu, M.; Thompson, D.; Lufkin, B.; Do, R.; Feinberg, L.; Chillarige, Y.; Dimova, R.; et al. Surveillance of COVID-19 Vaccine Safety among Elderly Persons Aged 65 Years and Older. Vaccine 2023, 41, 532–539. [Google Scholar] [CrossRef] [PubMed]
- Ellington, S.; Olson, C.K. Safety of MRNA COVID-19 Vaccines during Pregnancy. Lancet Infect. Dis. 2022, 22, 1514–1515. [Google Scholar] [CrossRef]
- Sadarangani, M.; Soe, P.; Shulha, H.P.; Valiquette, L.; Vanderkooi, O.G.; Kellner, J.D.; Muller, M.P.; Top, K.A.; Isenor, J.E.; McGeer, A.; et al. Safety of COVID-19 Vaccines in Pregnancy: A Canadian National Vaccine Safety (CANVAS) Network Cohort Study. Lancet Infect. Dis. 2022, 22, 1553–1564. [Google Scholar] [CrossRef] [PubMed]
- Lipkind, H.S.; Vazquez-Benitez, G.; DeSilva, M.; Vesco, K.K.; Ackerman-Banks, C.; Zhu, J.; Boyce, T.G.; Daley, M.F.; Fuller, C.C.; Getahun, D.; et al. Receipt of COVID-19 Vaccine During Pregnancy and Preterm or Small-for-Gestational-Age at Birth—Eight Integrated Health Care Organizations, United States, December 15, 2020–July 22, 2021. MMWR Morb. Mortal. Wkly. Rep. 2022, 71, 26–30. [Google Scholar] [CrossRef] [PubMed]
- Santi Laurini, G.; Montanaro, N.; Motola, D. Safety of COVID-19 Vaccines in Pregnancy: A VAERS Based Analysis. Eur. J. Clin. Pharmacol. 2023, 79, 657–661. [Google Scholar] [CrossRef] [PubMed]
- Prasad, S.; Kalafat, E.; Blakeway, H.; Townsend, R.; O’Brien, P.; Morris, E.; Draycott, T.; Thangaratinam, S.; Le Doare, K.; Ladhani, S.; et al. Systematic Review and Meta-Analysis of the Effectiveness and Perinatal Outcomes of COVID-19 Vaccination in Pregnancy. Nat. Commun. 2022, 13, 2414. [Google Scholar] [CrossRef] [PubMed]
- Lin, M.J.; Svensson-Arvelund, J.; Lubitz, G.S.; Marabelle, A.; Melero, I.; Brown, B.D.; Brody, J.D. Cancer Vaccines: The next Immunotherapy Frontier. Nat. Cancer 2022, 3, 911–926. [Google Scholar] [CrossRef] [PubMed]
- Ljungman, P.; Hakki, M.; Boeckh, M. Cytomegalovirus in Hematopoietic Stem Cell Transplant Recipients. Hematol. Oncol. Clin. North. Am. 2011, 25, 151–169. [Google Scholar] [CrossRef]
- Pass, R.F.; Duliegè, A.; Boppana, S.; Sekulovich, R.; Percell, S.; Britt, W.; Burke, R.L. A Subunit Cytomegalovirus Vaccine Based on Recombinant Envelope Glycoprotein B and a New Adjuvant. J. Infect. Dis. 1999, 180, 970–975. [Google Scholar] [CrossRef]
- Bernstein, D.I.; Munoz, F.M.; Callahan, S.T.; Rupp, R.; Wootton, S.H.; Edwards, K.M.; Turley, C.B.; Stanberry, L.R.; Patel, S.M.; Mcneal, M.M.; et al. Safety and Efficacy of a Cytomegalovirus Glycoprotein B (GB) Vaccine in Adolescent Girls: A Randomized Clinical Trial. Vaccine 2016, 34, 313–319. [Google Scholar] [CrossRef]
- Gomes, A.C.; Baraniak, I.A.; Lankina, A.; Moulder, Z.; Holenya, P.; Atkinson, C.; Tang, G.; Mahungu, T.; Kern, F.; Griffiths, P.D.; et al. The Cytomegalovirus GB/MF59 Vaccine Candidate Induces Antibodies against an Antigenic Domain Controlling Cell-to-Cell Spread. Nat. Commun. 2023, 14, 1041. [Google Scholar] [CrossRef]
- Somes, M.P.; Turner, R.M.; Dwyer, L.J.; Newall, A.T. Estimating the Annual Attack Rate of Seasonal Influenza among Unvaccinated Individuals: A Systematic Review and Meta-Analysis. Vaccine 2018, 36, 3199–3207. [Google Scholar] [CrossRef]
- Tyrrell, C.S.B.; Allen, J.L.Y.; Gkrania-Klotsas, E. Influenza: Epidemiology and Hospital Management. Medicine 2021, 49, 797–804. [Google Scholar] [CrossRef]
- Jones-Gray, E.; Robinson, E.J.; Kucharski, A.J.; Fox, A.; Sullivan, S.G. Does Repeated Influenza Vaccination Attenuate Effectiveness? A Systematic Review and Meta-Analysis. Lancet Respir. Med. 2023, 11, 27–44. [Google Scholar] [CrossRef]
- Li, Y.; Wang, X.; Blau, D.M.; Caballero, M.T.; Feikin, D.R.; Gill, C.J.; Madhi, S.A.; Omer, S.B.; Simões, E.A.F.; Campbell, H.; et al. Global, Regional, and National Disease Burden Estimates of Acute Lower Respiratory Infections Due to Respiratory Syncytial Virus in Children Younger than 5 Years in 2019: A Systematic Analysis. Lancet 2022, 399, 2047–2064. [Google Scholar] [CrossRef]
- Jenkins, V.A.; Hoet, B.; Hochrein, H.; De Moerlooze, L. The Quest for a Respiratory Syncytial Virus Vaccine for Older Adults: Thinking beyond the F Protein. Vaccines 2023, 11, 382. [Google Scholar] [CrossRef] [PubMed]
- Xia, Y.; Zhang, X.; Zhang, L.; Fu, C. Efficacy, Effectiveness, and Safety of Herpes Zoster Vaccine in the Immunocompetent and Immunocompromised Subjects: A Systematic Review and Network Meta-Analysis. Front. Immunol. 2022, 13, 978203. [Google Scholar] [CrossRef]
- Mbinta, J.F.; Nguyen, B.P.; Awuni, P.M.A.; Paynter, J.; Simpson, C.R. Post-Licensure Zoster Vaccine Effectiveness against Herpes Zoster and Postherpetic Neuralgia in Older Adults: A Systematic Review and Meta-Analysis. Lancet Healthy Longev. 2022, 3, e263–e275. [Google Scholar] [CrossRef] [PubMed]
- Martins, M.M.; Alves da Cunha, A.J.L.; Robaina, J.R.; Raymundo, C.E.; Barbosa, A.P.; de Andrade Medronho, R. Fetal, Neonatal, and Infant Outcomes Associated with Maternal Zika Virus Infection during Pregnancy: A Systematic Review and Meta-Analysis. PLoS ONE 2021, 16, e0246643. [Google Scholar] [CrossRef] [PubMed]
- Bergman, H.; Buckley, B.S.; Villanueva, G.; Petkovic, J.; Garritty, C.; Lutje, V.; Riveros-Balta, A.X.; Low, N.; Henschke, N. Comparison of Different Human Papillomavirus (HPV) Vaccine Types and Dose Schedules for Prevention of HPV-Related Disease in Females and Males. Cochrane Database Syst. Rev. 2019, 2019, CD013479. [Google Scholar] [CrossRef]
- Hanna, N.; Heffes-Doon, A.; Lin, X.; Manzano De Mejia, C.; Botros, B.; Gurzenda, E.; Nayak, A. Detection of Messenger RNA COVID-19 Vaccines in Human Breast Milk. JAMA Pediatr. 2022, 176, 1268. [Google Scholar] [CrossRef]
- Ndeupen, S.; Qin, Z.; Jacobsen, S.; Bouteau, A.; Estanbouli, H.; Igyártó, B.Z. The MRNA-LNP Platform’s Lipid Nanoparticle Component Used in Preclinical Vaccine Studies Is Highly Inflammatory. iScience 2021, 24, 103479. [Google Scholar] [CrossRef]
- Schieffelin, J.S.; Norton, E.B.; Kolls, J.K. What Should Define a SARS-CoV-2 “Breakthrough” Infection? J. Clin. Investig. 2021, 131, e151186. [Google Scholar] [CrossRef]
- Beatty, G.L.; Gladney, W.L. Immune Escape Mechanisms as a Guide for Cancer Immunotherapy. Clin. Cancer Res. 2015, 21, 687–692. [Google Scholar] [CrossRef]
- Chemaitelly, H.; Ayoub, H.H.; AlMukdad, S.; Coyle, P.; Tang, P.; Yassine, H.M.; Al-Khatib, H.A.; Smatti, M.K.; Hasan, M.R.; Al-Kanaani, Z.; et al. Duration of MRNA Vaccine Protection against SARS-CoV-2 Omicron BA.1 and BA.2 Subvariants in Qatar. Nat. Commun. 2022, 13, 3082. [Google Scholar] [CrossRef] [PubMed]
- Agrati, C.; Di Cosimo, S.; Fenoglio, D.; Apolone, G.; Ciceri, F.; Ciliberto, G.; Baldanti, F.; Costantini, M.; Giannarelli, D.; Ippolito, G.; et al. COVID-19 Vaccination in Fragile Patients: Current Evidence and an Harmonized Transdisease Trial. Front. Immunol. 2021, 12, 704110. [Google Scholar] [CrossRef] [PubMed]
- Vanetti, C.; Lampasona, V.; Stracuzzi, M.; Fenizia, C.; Biasin, M.; Saulle, I.; Limanaqi, F.; Abdelsalam, A.; Loretelli, C.; Paradiso, L.; et al. The Immunological Profile of SARS-CoV-2 Infection in Children Is Linked to Clinical Severity and Age. Int. J. Mol. Sci. 2023, 24, 6779. [Google Scholar] [CrossRef] [PubMed]
- Vanetti, C.; Trabattoni, D.; Stracuzzi, M.; Amendola, A.; Fappani, C.; Rubinacci, V.; Fenizia, C.; Gianolio, L.; Biasin, M.; Dighera, A.; et al. Immunological Characterization of HIV and SARS-CoV-2 Coinfected Young Individuals. Cells 2021, 10, 3187. [Google Scholar] [CrossRef]
- Fenizia, C.; Moltrasio, C.; Ottobrini, L.; Utyro, O.; Genovese, G.; Vanetti, C.; Trabattoni, D.; Marzano, A.V. SARS-CoV-2 Vaccination Effectiveness in Rituximab-Treated Patients Affected by Pemphigus Vulgaris. J. Investig. Dermatol. 2023, 143, 1601–1604. [Google Scholar] [CrossRef]
- Brenner, S.; JACOB, F.; Meselson, M. An Unstable Intermediate Carrying Information from Genes to Ribosomes for Protein Synthesis. Nature 1961, 190, 576–581. [Google Scholar] [CrossRef]
- Gurdon, J.B.; Lane, C.D.; Woodland, H.R.; Marbaix, G. Use of Frog Eggs and Oocytes for the Study of Messenger RNA and Its Translation in Living Cells. Nature 1971, 233, 177–182. [Google Scholar] [CrossRef]
- Malone, R.W.; Felgner, P.L.; Verma, I.M. Cationic Liposome-Mediated RNA Transfection. Proc. Natl. Acad. Sci. USA 1989, 86, 6077–6081. [Google Scholar] [CrossRef]
- Conry, R.M.; LoBuglio, A.F.; Wright, M.; Sumerel, L.; Pike, M.J.; Johanning, F.; Benjamin, R.; Lu, D.; Curiel, D.T. Characterization of a Messenger RNA Polynucleotide Vaccine Vector. Cancer Res. 1995, 55, 1397–1400. [Google Scholar]
- Wolff, J.A.; Malone, R.W.; Williams, P.; Chong, W.; Acsadi, G.; Jani, A.; Felgner, P.L. Direct Gene Transfer into Mouse Muscle in Vivo. Science 1990, 247, 1465–1468. [Google Scholar] [CrossRef]
- Heiser, A.; Coleman, D.; Dannull, J.; Yancey, D.; Maurice, M.A.; Lallas, C.D.; Dahm, P.; Niedzwiecki, D.; Gilboa, E.; Vieweg, J. Autologous Dendritic Cells Transfected with Prostate-Specific Antigen RNA Stimulate CTL Responses against Metastatic Prostate Tumors. J. Clin. Investig. 2002, 109, 409–417. [Google Scholar] [CrossRef] [PubMed]
- Kormann, M.S.D.; Hasenpusch, G.; Aneja, M.K.; Nica, G.; Flemmer, A.W.; Herber-Jonat, S.; Huppmann, M.; Mays, L.E.; Illenyi, M.; Schams, A.; et al. Expression of Therapeutic Proteins after Delivery of Chemically Modified MRNA in Mice. Nat. Biotechnol. 2011, 29, 154–157. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Disease | Completed | Active, Not Recruiting | Recruiting | Not yet Recruiting | Terminated/Unknown | Total | |
|---|---|---|---|---|---|---|---|
| Cancer | Advanced solid | 1 | 2 | 1 | 4 | ||
| AML | 1 | 1 | |||||
| Breast | 1 | 1 | |||||
| EBV related | 2 | 2 | |||||
| Gastrointestinal | 1 | 1 | 2 | 4 | |||
| Glioblastoma | 3 | 1 | 1 | 1 | 6 | ||
| Melanoma | 2 | 2 | 1 | 1 | 6 | ||
| NSCLC | 2 | 2 | |||||
| Prostate | 1 | 1 | |||||
| other | 1 | 2 | 2 | 5 | |||
| Viral infections | Cytomegalovirus | 2 | 1 | 3 | 1 | 7 | |
| Epstein–Barr | 1 | 1 | |||||
| Hepatitis B | 1 | 1 | |||||
| Herpes simplex 2 | 1 | 1 | |||||
| Varicella zoster | 2 | 2 | |||||
| HIV | 1 | 3 | 1 | 5 | |||
| Influenza | 2 | 4 | 8 | 14 | |||
| Metapneumovirus | 2 | 2 | |||||
| Nipah | 1 | 1 | |||||
| Papilloma | 2 | 2 | |||||
| Rabies | 2 | 2 | |||||
| Respiratory Syncytial | 2 | 4 | 6 | ||||
| Zika | 2 | 1 | 3 | ||||
| Influenza + SARS-CoV-2 | 1 | 1 | |||||
| Plasmodium falciparum | 1 | 1 | |||||
| Mycobacterium tuberculosis | 1 | 2 | 3 | ||||
| Total excluding SARS-CoV-2 | 23 | 16 | 31 | 9 | 5 | 84 | |
| Viral infections by SARS-CoV-2 | 72 | 87 | 116 | 35 | 22 | 332 | |
| Total | 95 | 103 | 147 | 44 | 27 | 416 |
| Disease | Active st. mRNA /Active Studies | Active st. mRNA/Vaccine Studies | Active st. mRNA/Active Vaccine st. | |
|---|---|---|---|---|
| Cancer | Advanced solid | 0.1% | 10.5% | 14.8% |
| EBV related | 9.1% | 50% | 100% | |
| Gastrointestinal | <0.1% | 1.3% | 3.6% | |
| Glioblastoma | 0.5% | 3.4% | 6.8% | |
| Melanoma | 0.4% | 1.7% | 10.8% | |
| Viral infections | Cytomegalovirus | 5.5% | 12.5% | 29.4% |
| Epstein–Barr | 0.8% | 7.1% | 16.7% | |
| Herpes simplex 2 | 4.0% | 8.3% | 2.6% | |
| Varicella zoster | 2.2% | 1.0% | 4.4% | |
| HIV | 0.2% | 0.6% | 3.3% | |
| Influenza | 4.0% | 0.8% | 8.4% | |
| Nipah | 50.0% | 33.3% | 50% | |
| Papilloma | 1.6% | 1.6% | 5% | |
| Respiratory Syncytial | 7.7% | 4.6% | 14.0% | |
| Zika | 6.7% | 3.7% | 33.3% | |
| Influenza + SARS-CoV-2 | 1.4% | 5.9% | 8.3% | |
| Plasmodium falciparum | 1.3% | 0.6% | 3.2% | |
| Mycobacterium tuberculosis | 0.7% | 1.4% | 6.5% | |
| Viral infections by SARS-CoV-2 | 7.0% | 17.7% | 26.1% |
| Disease | Phase 3 RCTs | Phase 2 RCTs | |||
|---|---|---|---|---|---|
| Cancer | Gastrointestinal | 0 | 1 | NCT05456165 | |
| Melanoma | 0 | 2 | NCT03897881; NCT04526899 | ||
| Viral infections | Cytomegalovirus | 1 | NCT05085366 | 2 | NCT04232280; NCT05683457 |
| Varicella zoster | 0 | 1 | NCT05703607 | ||
| Influenza | 3 | NCT05415462; NCT05540522; NCT05566639 | 1 | NCT05606965 | |
| Papilloma | 0 | 1 | NCT04534205 | ||
| Respiratory Syncytial | 2 | NCT05127434; NCT05330975 (phase 2/3) | 0 | ||
| Zika | 0 | 1 | NCT04917861 | ||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Perrotta, C.; Fenizia, C.; Carnovale, C.; Pozzi, M.; Trabattoni, D.; Cervia, D.; Clementi, E. Updated Considerations for the Immunopharmacological Aspects of the “Talented mRNA Vaccines”. Vaccines 2023, 11, 1481. https://doi.org/10.3390/vaccines11091481
Perrotta C, Fenizia C, Carnovale C, Pozzi M, Trabattoni D, Cervia D, Clementi E. Updated Considerations for the Immunopharmacological Aspects of the “Talented mRNA Vaccines”. Vaccines. 2023; 11(9):1481. https://doi.org/10.3390/vaccines11091481
Chicago/Turabian StylePerrotta, Cristiana, Claudio Fenizia, Carla Carnovale, Marco Pozzi, Daria Trabattoni, Davide Cervia, and Emilio Clementi. 2023. "Updated Considerations for the Immunopharmacological Aspects of the “Talented mRNA Vaccines”" Vaccines 11, no. 9: 1481. https://doi.org/10.3390/vaccines11091481
APA StylePerrotta, C., Fenizia, C., Carnovale, C., Pozzi, M., Trabattoni, D., Cervia, D., & Clementi, E. (2023). Updated Considerations for the Immunopharmacological Aspects of the “Talented mRNA Vaccines”. Vaccines, 11(9), 1481. https://doi.org/10.3390/vaccines11091481