
Maternal and Infant Histo-Blood Group Antigen (HBGA) Profiles and Their Influence on Oral Rotavirus Vaccine (RotarixTM) Immunogenicity among Infants in Zambia
 , , , , , ,
, , , , , ,
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Design and Participants
2.2. Laboratory Testing
2.2.1. Determination of the Infant ABO and Lewis HBGA Phenotypes in Saliva
2.2.2. Determination of the Infant FUT2 Genotypes
2.2.3. Measurement of Rotavirus-Specific IgA
2.3. Statistical Analysis
3. Results
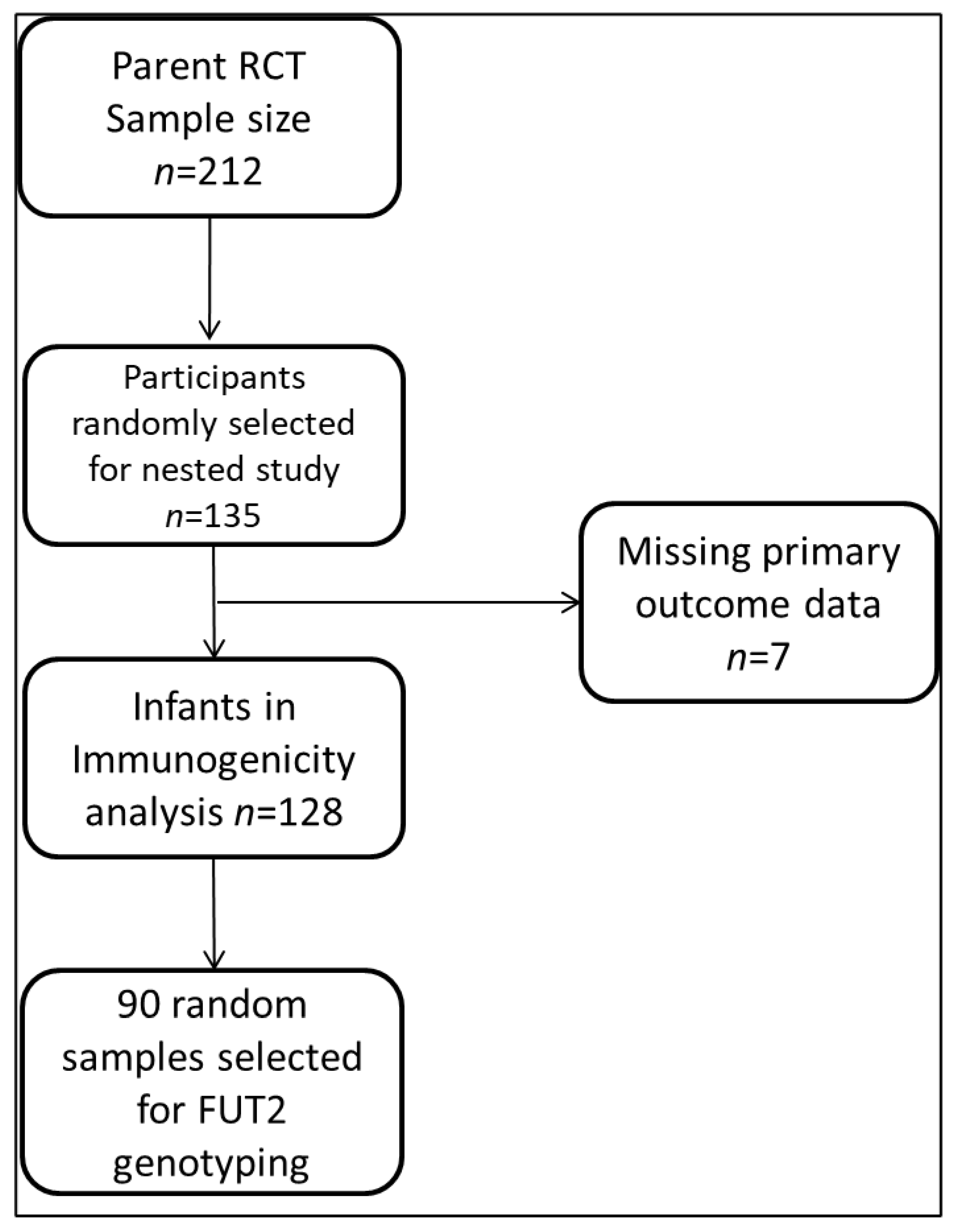
3.1. Participant and Sampling Flow Chart
3.2. Study Population Characteristics and Overall Seroconversion Frequency
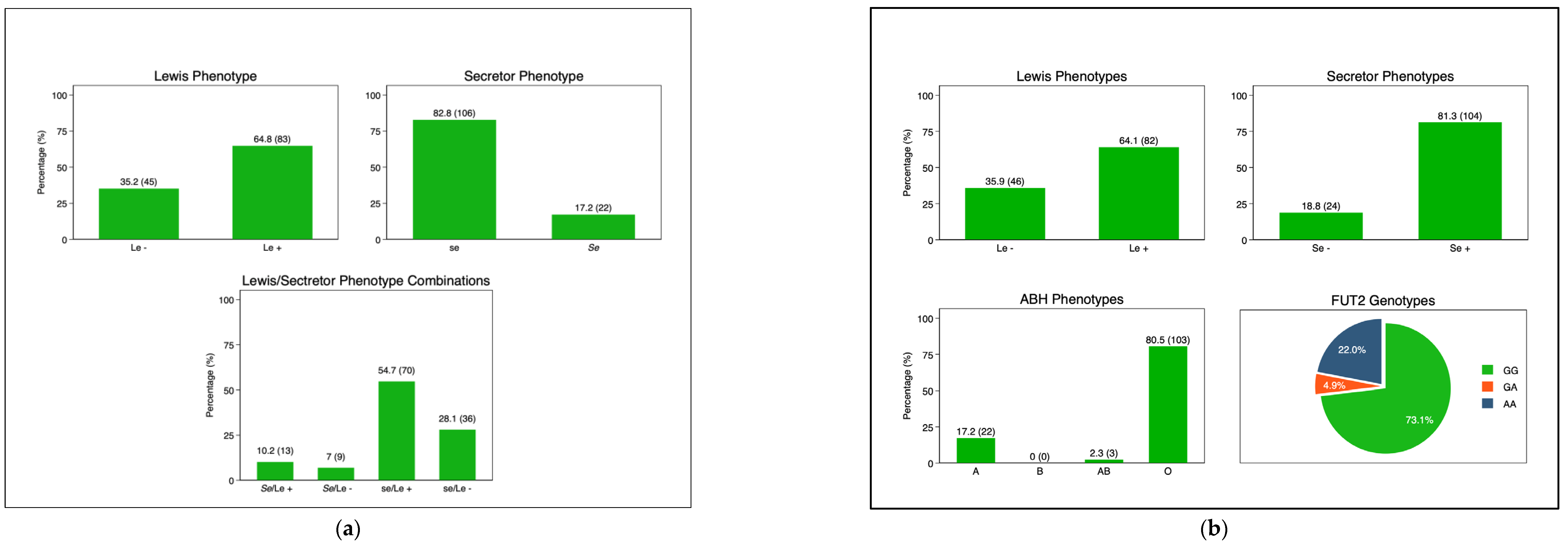
3.3. Mother and Infant HBGA Profiles
3.4. Maternal and Infant HBGA and RV-IgA Immunogenicity
3.5. Maternal and Infant HBGA and RV-IgA Immunogenicity 3 Months Post-Dose-3
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lanata, C.F.; Fischer-walker, C.L.; Olascoaga, A.C.; Torres, C.X.; Aryee, M.J. Global Causes of Diarrheal Disease Mortality in Children, 5 Years of Age: A Systematic Review. PLoS ONE 2013, 8, e72788. [Google Scholar] [CrossRef] [PubMed]
- Abbafati, C.; Abbas, K.M.; Abbasi-Kangevari, M.; Abd-Allah, F.; Abdelalim, A.; Abdollahi, M. Global burden of 369 diseases and injuries in 204 countries and territories, 1990–2019: A systematic analysis for the Global Burden of Disease Study 2019. Lancet 2020, 396, 1204–1222. [Google Scholar]
- Chilengi, R.; Rudd, C.; Bolton, C.; Guffey, B.; Masumbu, P.K.; Stringer, J. Successes, Challenges and Lessons Learned in Accelerating Introduction of Rotavirus Immunisation in Zambia. World J. Vaccines 2015, 5, 43–53. [Google Scholar] [CrossRef]
- Burnett, E.; Jonesteller, C.L.; Tate, J.E.; Yen, C.; Parashar, U.D. Global Impact of Rotavirus Vaccination on Childhood Hospitalizations and Mortality from Diarrhea. J. Infect. Dis. 2017, 215, 1666–1672. [Google Scholar] [CrossRef]
- Church, J.A.; Parker, E.P.; Kirkpatrick, B.D.; Grassly, N.C.; Prendergast, A.J. Interventions to Improve Oral Vaccine Performance in developing countries: A Systematic Review and Meta-analysis Protocol Information. Lancet Infect. Dis. 2017, 19, 203–214. [Google Scholar] [CrossRef]
- Meta-analysis, C.A.; Mwila-kazimbaya, K.; Bosomprah, S.; Simuyandi, M.; Chisenga, C.C.; Munsaka, S. Efficacy and Effectiveness of Rotavirus Vaccine on Incidence of Diarrhoea among Children: A Meta-analysis. Pediatr. Infect. Dis. Open Access 2018, 3, 4. [Google Scholar]
- Velasquez, D.E.; Parashar, U.; Jiang, B. Decreased performance of live attenuated, oral rotavirus vaccines in low-income settings: Causes and contributing factors. Expert Rev. Vaccines 2018, 17, 145–161. [Google Scholar] [CrossRef]
- Mwape, I.; Bosomprah, S.; Mwaba, J.; Mwila-kazimbaya, K.; Laban, N.M.; Chisenga, C.C.; Sijumbila, G.; Simuyandi, M.; Chilengi, R. Immunogenicity of rotavirus vaccine (Rotarix TM) in infants with environmental enteric dysfunction. PLoS ONE 2017, 12, e0187761. [Google Scholar] [CrossRef]
- Mwila-kazimbaya, K.; Garcia, M.P.; Bosomprah, S.; Laban, M.; Chisenga, C.C.; Permar, S.R.; Simuyandi, M.; Munsaka, S.; Chilengi, R. Effect of innate antiviral glycoproteins in breast milk on seroconversion to rotavirus vaccine (Rotarix) in children in Lusaka, Zambia. PLoS ONE 2017, 12, e0189351. [Google Scholar] [CrossRef]
- Chilengi, R.; Simuyandi, M.; Beach, L.; Mwila, K.; Becker, S. Association of Maternal Immunity with Rotavirus Vaccine Immunogenicity in Zambian Infants. PLoS ONE 2016, 11, e0150100. [Google Scholar] [CrossRef]
- Xu, S.; Liu, Y.; Tan, M.; Zhong, W.; Zhao, D.; Jiang, X.; Kennedy, M.A. Molecular basis of P [6] and P [8] major human rotavirus VP8 * domain interactions with histo-blood group antigens. BioRxiv 2019, 512301. [Google Scholar]
- Armah, G.E.; Cortese, M.M.; Dennis, F.E.; Yu, Y.; Morrow, A.L.; Mcneal, M.M.; Lewis, K.D.C.; Awuni, D.A.; Armachie, J.; Parashar, U.D. Rotavirus Vaccine Take in Infants Is Associated With Secretor Status. J. Infect. Dis. 2019, 219, 746–749. [Google Scholar] [CrossRef] [PubMed]
- Pollock, L.; Bennett, A.; Jere, K.C.; Dube, Q.; Mandolo, J.; Bar-Zeev, N.; Heyderman, R.S.; Cunliffe, N.A.; Iturriza-Gomara, M. Nonsecretor Histo-blood Group Antigen Phenotype Is Associated With Reduced Risk of Clinical Rotavirus Vaccine Failure in Malawian Infants. Clin Infect Dis. 2019, 69, 1313–1319. [Google Scholar] [CrossRef] [PubMed]
- Bucardo, F.; Nordgren, J.; Reyes, Y.; Gonzalez, F.; Sharma, S.; Svensson, L. The Lewis A phenotype is a restriction factor for Rotateq and Rotarix vaccine-take in Nicaraguan children. Sci. Rep. 2018, 8, 1502. [Google Scholar] [CrossRef]
- Lee, B.; Dickson, D.M.; DeCamp, A.C.; Ross Colgate, E.; Diehl, S.A.; Uddin, M.I.; Sharmin, S.; Islam, S.; Bhuiyan, T.R.; Alam, M.; et al. Histo–Blood group antigen phenotype determines susceptibility to genotype-specific rotavirus infections and impacts measures of rotavirus vaccine efficacy. J. Infect. Dis. 2018, 217, 1399–1407. [Google Scholar] [CrossRef]
- Nordgren, J.; Sharma, S.; Bucardo, F.; Nasir, W.; Günaydin, G.; Ouermi, D.; Nitiema, L.W.; Becker-Dreps, S.; Simpore, J.; Hammarström, L.; et al. Both lewis and secretor status mediate susceptibility to rotavirus infections in a rotavirus genotype-dependent manner. Clin. Infect. Dis. 2014, 59, 1567–1573. [Google Scholar] [CrossRef]
- Liu, Y.; Huang, P.; Tan, M.; Liu, Y.; Biesiada, J.; Meller, J.; Castello, A.A.; Jiang, B.; Jiang, X. Rotavirus VP8*: Phylogeny, Host Range, and Interaction with Histo-Blood Group Antigens. J. Virol. 2012, 86, 9899–9910. [Google Scholar] [CrossRef]
- Abegaz, S.B. Human ABO Blood Groups and Their Associations with Different Diseases. BioMed Res. Int. 2021, 2021, 6629060. [Google Scholar] [CrossRef]
- Daniels, G. Human Blood Groups, 2nd ed.; John Wiley and Sons: Hoboken, NJ, USA, 2016; pp. 1–23. Available online: https://onlinelibrary.wiley.com/doi/book/10.1002/9781118493595 (accessed on 23 May 2023).
- Jiang, X.; Huang, P.; Zhong, W.; Tan, M.; Farkas, T.; Morrow, A.L.; Newburg, D.S.; Ruiz-Palacios, G.M.; Pickering, L.K. Human Milk Contains Elements That Block Binding of Noroviruses to Human Histo—Blood Group Antigens in Saliva. J. Artic. 2004, 190, 3039. [Google Scholar] [CrossRef]
- Williams, F.B.; Kader, A.; Colgate, E.R.; Dickson, D.M.; Carmolli, M.; Uddin, M.I.; Sharmin, S.; Islam, S.; Bhuiyan, T.R.; Alam, M.; et al. Maternal Secretor Status Affects Oral Rotavirus Vaccine Response in Breastfed Infants in Bangladesh. J. Infect. Dis. 2020, 224, 1147–1151. [Google Scholar] [CrossRef]
- Colston, J.M.; Francois, R.; Pisanic, N.; Yori, P.P.; Mccormick, B.J.J.; Olortegui, M.P.; Gazi, A.; Svensen, E.; Ahmed, M.M.M.; Mduma, E.; et al. Effects of Child and Maternal Histo-Blood Group Antigen Status on Symptomatic and Asymptomatic Enteric Infections in Early Childhood. J. Infect. Dis. 2019, 220, 151–162. [Google Scholar] [CrossRef] [PubMed]
- Parker, E.P.K.; Ramani, S.; Lopman, B.A.; Church, J.A. Causes of impaired oral vaccine efficacy in developing countries. Futur. Microbiol. 2018, 13, 97–118. [Google Scholar] [CrossRef] [PubMed]
- Laban, N.M.; Bosomprah, S.; Simuyandi, M.; Chibuye, M.; Chauwa, A.; Chirwa-Chobe, M.; Sukwa, N.; Chipeta, C.; Velu, R.; Njekwa, K.; et al. Evaluation of ROTARIX ® Booster Dose Vaccination at 9 Months for Safety and Enhanced Anti-Rotavirus Immunity in Zambian Children: A Randomised Controlled Trial. Vaccines 2023, 11, 346. [Google Scholar] [CrossRef] [PubMed]
- Armah, G.E.; Sow, S.O.; Breiman, R.F.; Dallas, M.J.; Tapia, M.D.; Feikin, D.R.; Binka, F.N.; Steele, A.D.; Laserson, K.F.; Ansah, N.A.; et al. Efficacy of pentavalent rotavirus vaccine against severe rotavirus gastroenteritis in infants in developing countries in sub-Saharan Africa: A randomised, double-blind, placebo-controlled trial. Lancet 2010, 376, 606–614. [Google Scholar] [CrossRef] [PubMed]
- Ferrer-Admetlla, A.; Sikora, M.; Laayouni, H.; Esteve, A.; Roubinet, F.; Blancher, A.; Calafell, F.; Bertranpetit, J.; Casals, F. A natural history of FUT2 polymorphism in humans. Mol. Biol. Evol. 2009, 26, 1993–2003. [Google Scholar] [CrossRef] [PubMed]
- Sun, X.; Guo, N.; Li, J.; Yan, X.; He, Z.; Li, D.; Jin, M.; Xie, G.; Pang, L.; Zhang, Q.; et al. Rotavirus infection and histo-blood group antigens in the children hospitalized with diarrhoea in China. Clin. Microbiol. Infect. 2016, 22, 740.e1–740.e3. [Google Scholar] [CrossRef] [PubMed]
- De Mattos, L.C. Structural diversity and biological importance of ABO, H, Lewis and secretor histo-blood group carbohydrates. Rev. Bras. Hematol. Hemoter. 2016, 38, 331–340. [Google Scholar] [CrossRef]
- Payne, D.C.; Currier, R.L.; Staat, M.A.; Sahni, L.C.; Selvarangan, R.; Halasa, N.B.; Englund, J.A.; Weinberg, G.A.; Boom, J.A.; Szilagyi, P.G.; et al. Epidemiologic association between FUT2 secretor status and severe rotavirus gastroenteritis in children in the United States. JAMA Pediatr. 2015, 169, 1040–1045. [Google Scholar] [CrossRef]
- Heggelund, J.E.; Varrot, A.; Imberty, A. Histo-blood group antigens as mediators of infections. Curr. Opin. Struct. Biol. 2017, 44, 190–200. [Google Scholar] [CrossRef]
- Jiang, X.; Liu, Y.; Tan, M. Histo-blood group antigens as receptors for rotavirus, new understanding on rotavirus epidemiology and vaccine strategy Rotavirus host receptor and vaccine strategy Histo-blood group antigens as receptors for rotavirus, new understanding on rotavirus ep. Nat. Publ. Gr. 2019, 1751. [Google Scholar] [CrossRef]
- Kazi, A.M.; Cortese, M.M.; Yu, Y.; Lopman, B.; Morrow, A.L.; Fleming, J.A.; McNeal, M.M.; Steele, A.D.; Parashar, U.D.; Zaidi, A.K.M.; et al. Secretor and salivary ABO blood group antigen status predict rotavirus vaccine take in infants. J. Infect. Dis. 2017, 215, 786–789. [Google Scholar] [CrossRef]
- Yang, T.A.; Hou, J.Y.; Huang, Y.C.; Chen, C.J. Genetic Susceptibility to Rotavirus Gastroenteritis and Vaccine Effectiveness in Taiwanese Children. Sci. Rep. 2017, 7, 6412. [Google Scholar] [CrossRef]
- Harris, V.; Ali, A.; Fuentes, S.; Korpela, K.; Kazi, M.; Tate, J.; Parashar, U.; Wiersinga, W.J.; Giaquinto, C.; de Weerth, C.; et al. Rotavirus vaccine response correlates with the infant gut microbiota composition in Pakistan. Gut Microbes 2018, 9, 93–101. [Google Scholar] [CrossRef]
- Sharma, S.; Nordgren, J. Effect of infant and maternal secretor status on rotavirus vaccine take—An overview. Viruses 2021, 13, 1144. [Google Scholar] [CrossRef]
- Böhm, R.; Fleming, F.E.; Maggioni, A.; Dang, V.T.; Holloway, G.; Coulson, B.S.; von Itzstein, M.; Haselhorst, T. Revisiting the role of histo-blood group antigens in rotavirus host-cell invasion. Nat. Commun. 2015, 6, 5907. [Google Scholar] [CrossRef]
- Huang, P.; Xia, M.; Tan, M.; Zhong, W.; Wei, C.; Wang, L.; Morrow, A.; Jiang, X. Spike Protein VP8* of Human Rotavirus Recognizes Histo-Blood Group Antigens in a Type-Specific Manner. J. Virol. 2012, 86, 4833–4843. [Google Scholar] [CrossRef]
- Cooling, L. Blood Groups in Infection and Host Susceptibility. Clin. Microbiol. Rev. 2015, 28, 801–870. [Google Scholar] [CrossRef] [PubMed]
- Van Trang, N.; Vu, H.T.; Le, N.T.; Huang, P.; Jiang, X.; Anh, D.D. Association between norovirus and rotavirus infection and histo-blood group antigen types in Vietnamese children. J. Clin. Microbiol. 2014, 52, 1366–1374. [Google Scholar] [CrossRef]
- Bekdas, M.; Demircioglu, F.; Goksugur, S.B.; Kucukbayrak, B. Is there a relationship between rotavirus-positive gastroenteritis and ABO blood groups? A retrospective cohort study. Arch. Argent. Pediatr. 2014, 112, 345–348. [Google Scholar] [PubMed]
- Mihala, G.; Ware, R.S.; Cheung, C.; Lambert, S.B.; Sly, P.D.; Whiley, D.M.; Grimwood, K. Intestinal shedding of enteric agents in histo-blood group antigen-secretor children in an Australian community-based birth cohort study. J. Pediatr. Neonatal Individ. Med. 2022, 11, e110116. [Google Scholar]
- Ramani, S.; Giri, S. Influence of histo blood group antigen expression on susceptibility to enteric viruses and vaccines. Curr. Opin. Infect. Dis. 2019, 32, 445–452. [Google Scholar] [CrossRef] [PubMed]
- Ayouni, S.; Sdiri-Loulizi, K.; de Rougemont, A.; Estienney, M.; Ambert-Balay, K.; Aho, S.; Hamami, S.; Aouni, M.; Neji-Guediche, M.; Pothier, P.; et al. Rotavirus P [8] infections in persons with secretor and nonsecretor phenotypes, Tunisia. Emerg. Infect. Dis. 2015, 21, 2055–2058. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Seroconverted | ||||
|---|---|---|---|---|
| Mother–Infant Pairs (N = 128) | No (n = 91, 71.1%) | Yes (n = 37, 28.9%) | p-Value | |
| n (% of total) | n (%) | n (%) | ||
| Infants’ Characteristics | ||||
| Age (Weeks) | ||||
| Median (IQR) | 6 (6–6) | 6 (6–6) | 6 (6–6) | 0.442 |
| Mean (SD) | 6 (0.6) | 6 (0.6) | 5.9 (0.7) | |
| Sex | ||||
| Male | 69 (53.9) | 51 (73.9) | 18 (26.0) | 0.447 |
| Female | 59 (46.1) | 40 (67.7) | 19 (32.2) | |
| Feeding | ||||
| Exclusively breastfeeding | 122 (95.3) | 86 (70.4) | 36 (29.5) | 0.672 |
| Mixed feeding | 6 (4.7) | 5 (83.3) | 1 (16.6) | |
| Birthweight (kg) | ||||
| <2.5 | 5 (3.9) | 3 (60.0) | 2 (40.0) | 0.626 |
| ≥2.5 | 123 (96.1) | 88 (71.5) | 35 (28.4) | |
| HIV Exposure | ||||
| Not exposed | 89 (69.5) | 62 (69.6) | 27 (30.3) | 0.590 |
| Exposed | 39 (30.5) | 28 (73.6) | 10 (26.3) | |
| Nutritional Status | ||||
| Stunted | ||||
| No (HAZ ≥ −2) | 107 (83.6) | 78 (72.8) | 29 (27.1) | 0.310 |
| Yes (HAZ < −2) | 21 (16.4) | 13 (61.9) | 8 (38.0) | |
| Wasting | ||||
| No (WAZ ≥ −2) | 119 (93.0) | 86 (72.2) | 33 (27.7) | 0.281 |
| Yes (WAZ < −2) | 9 (7.0) | 5 (55.5) | 4 (44.4) | |
| Mothers’ Characteristics | ||||
| Age (years) | ||||
| <20 | 20 (15.6) | 15 (75.0) | 5 (25.0) | 0.080 |
| 20–24 | 45 (35.2) | 37 (82.2) | 8 (17.7) | |
| 25–29 | 34 (26.6) | 19 (55.8) | 15 (44.1) | |
| ≥30 | 29 (22.7) | 20 (68.9) | 9 (31.0) | |
| Highest Education Level | ||||
| None | 6 (4.7) | 4 (66.7) | 2 (33.3) | 0.470 * |
| Primary | 40 (31.3) | 25 (62.5) | 15 (37.5) | |
| Secondary | 81 (63.3) | 61 (75.3) | 20 (24.6) | |
| Tertiary | 1 (0.8) | 1 (100.0) | 0 (0.0) | |
| Water Source | ||||
| Piped into house/yard | 45 (35.2) | 33 (75.0) | 12 (25.0) | 0.882 |
| Protected well | 5 (3.9) | 4 (80.0) | 1 (20.0) | |
| Public borehole/tap and pipe | 78 (60.9) | 54 (80.0) | 24 (20.0) | |
| Shared Toilet Facility | ||||
| No | 24 (18.8) | 17 (70.8) | 7 (29.1) | 0.975 |
| Yes | 104 (81.3) | 74 (71.1) | 30 (28.8) | |
| Type of Toilet Facility | ||||
| Flushing toilet | 26 (20.3) | 17 (65.4) | 9 (34.6) | 0.476 |
| Pit latrine | 102 (79.7) | 74 (72.6) | 28 (27.5) | |
| Characteristics | Number of Mother–Infant Pairs (% of Total) | GMTs (95% CI) | ANOVA p-Value | Seroconversion (n = 37, 28.9%) | Chi-Square p-Value | Crude Odds Ratio (95% CI) | p-Value |
|---|---|---|---|---|---|---|---|
| n (%) | |||||||
| Infant | |||||||
| Infant HBGA Phenotype | |||||||
| A | 22 (17.2) | 2.5 (0.9, 6.8) | 0.874 | 7 (31.8) | 0.929 | ref | |
| AB | 3 (2.3) | 1.6 (0, 270.6) | 1 (33.3) | 1.1 (0.1, 13.9) | 0.958 | ||
| O | 103 (80.5) | 1.9 (1.2, 3) | 29 (28.2) | 0.8 (0.3, 2.3) | 0.731 | ||
| Infant Lewis Phenotype | |||||||
| Le− (Le a−b−) | 46 (35.9) | 2.6 (1.3, 5.2) | 0.332 | 14 (30.4) | 0.775 | ref | |
| Le+ (Le a+b−, Le a−b+, or Le a+b+) | 82 (64.1) | 1.7 (1.1, 2.8) | 23 (28.2) | 0.9 (0.4, 2) | 0.775 | ||
| Secretor Phenotype | |||||||
| Non-secretor (se) | 24 (18.8) | 1.3 (0.6, 2.8) | 0.279 | 5 (20.8) | 0.24 | ref | |
| Secretor Phenotype (Se) | 104 (81.3) | 2.2 (1.4, 3.5) | 32 (30.8) | 1.7 (0.6, 4.9) | 0.337 | ||
| Infant FUT2 Genotype * | |||||||
| Homozygous secretor (GG) | 60 (46.9) | 1.4 (0.8, 2.5) | 0.093 | 15 (25.0) | 0.289 | ref | |
| Heterozygous secretor (GA) | 4 (3.1) | 5.6 (0, 1426.5) | 2 (50.0) | 3 (0.4, 23.2) | 0.292 | ||
| Non-secretor (AA) | 18 (14.1) | 4.9 (1.5, 16.3) | 7 (38.9) | 1.9 (0.6, 5.8) | 0.255 | ||
| Missing | 46 (35.9) | 2 (1, 3.8) | 13 (28.3) | - | - | ||
| Mother | |||||||
| Lewis Phenotype | |||||||
| Le− (Le a−b−) | 45 (35.2) | 1.6 (0.9, 2.8) | 0.358 | 13 (28.9) | 0.997 | ref | |
| Le+ (Le a+b−, Le a−b+, or Le a+b+) | 83 (64.8) | 2.3 (1.4, 3.9) | 24 (28.9) | 1.0 (0.4, 2.2) | 0.997 | ||
| Secretor Phenotype | |||||||
| Non-secretor (se) | 106 (82.8) | 2 (1.3, 3.1) | 0.85 | 32 (30.2) | 0.336 | ref | |
| Secretor Phenotype (Se) | 22 (17.2) | 1.8 (0.7, 5.2) | 5 (22.7) | 0.7 (0.2, 2.0) | 0.484 |
| Characteristics | V12 GMTs | ANOVA, p-Value | GMT Ratio (95% CI) | p-Value |
|---|---|---|---|---|
| GMT (95% CI) | ||||
| Infant | ||||
| Infant ABO Phenotype | ||||
| A | 5.02 (4.14, 6.07) | 0.002 | ref | |
| AB | 5.28 (1.86, 15) | 0.59 (0.10, 3.47) | 0.560 | |
| O | 3.7 (3.35, 4.08) | 0.36 (0.09, 1.41) | 0.140 | |
| Infant Lewis Phenotype | ||||
| Le− (Le a−b−) | 3.57 (3.03, 4.22) | 0.015 | ref | |
| Le+ (Le a+b−, Le a−b+, or Le a+b+) | 4.17 (3.75, 4.63) | 0.83 (0.31, 2.23) | 0.705 | |
| Secretor Phenotype | ||||
| Non-secretor (se) | 2.89 (2.26, 3.71) | <0.001 | ref | |
| Secretor phenotype (Se) | 4.14 (3.78, 4.54) | 1.94 (0.59, 6.4) | 0.276 | |
| Infant FUT2 Genotype | ||||
| Secretor (GG)/(GA) | 3.95 (3.45, 4.52) | 0.063 | ref | |
| Non-secretor (AA) | 3.24 (2.44, 4.31) | 1.66 (0.96, 2.83) | 0.543 | |
| Mother | ||||
| Lewis phenotype | ||||
| Le− (Le a−b−) | 4.02 (3.52, 4.58) | 0.521 | ref | |
| Le+ (Le a+b−, Le a−b+, or Le a+b+) | 3.95 (3.51, 4.44) | 1.09 (0.41, 2.88) | 0.863 | |
| Secretor Phenotype | ||||
| Non-secretor (se) | 4.08 (3.72, 4.48) | 0.368 | ref | |
| Secretor Phenotype (Se) | 3.45 (2.64, 4.51) | 0.83 (0.25, 2.70) | 0.751 | |
| Treatment Arm | ||||
| Control (MR) | 4.08 (3.56, 4.67) | 0.260 | ref | |
| Intervention (ROTARIX® + MR) | 3.88 (3.44, 4.37) | 1.39 (0.55, 3.49) | 0.479 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chauwa, A.; Bosomprah, S.; Laban, N.M.; Phiri, B.; Chibuye, M.; Chilyabanyama, O.N.; Munsaka, S.; Simuyandi, M.; Mwape, I.; Mubanga, C.; et al. Maternal and Infant Histo-Blood Group Antigen (HBGA) Profiles and Their Influence on Oral Rotavirus Vaccine (RotarixTM) Immunogenicity among Infants in Zambia. Vaccines 2023, 11, 1303. https://doi.org/10.3390/vaccines11081303
Chauwa A, Bosomprah S, Laban NM, Phiri B, Chibuye M, Chilyabanyama ON, Munsaka S, Simuyandi M, Mwape I, Mubanga C, et al. Maternal and Infant Histo-Blood Group Antigen (HBGA) Profiles and Their Influence on Oral Rotavirus Vaccine (RotarixTM) Immunogenicity among Infants in Zambia. Vaccines. 2023; 11(8):1303. https://doi.org/10.3390/vaccines11081303
Chicago/Turabian StyleChauwa, Adriace, Samuel Bosomprah, Natasha Makabilo Laban, Bernard Phiri, Mwelwa Chibuye, Obvious Nchimunya Chilyabanyama, Sody Munsaka, Michelo Simuyandi, Innocent Mwape, Cynthia Mubanga, and et al. 2023. "Maternal and Infant Histo-Blood Group Antigen (HBGA) Profiles and Their Influence on Oral Rotavirus Vaccine (RotarixTM) Immunogenicity among Infants in Zambia" Vaccines 11, no. 8: 1303. https://doi.org/10.3390/vaccines11081303
APA StyleChauwa, A., Bosomprah, S., Laban, N. M., Phiri, B., Chibuye, M., Chilyabanyama, O. N., Munsaka, S., Simuyandi, M., Mwape, I., Mubanga, C., Chobe, M. C., Chisenga, C., & Chilengi, R. (2023). Maternal and Infant Histo-Blood Group Antigen (HBGA) Profiles and Their Influence on Oral Rotavirus Vaccine (RotarixTM) Immunogenicity among Infants in Zambia. Vaccines, 11(8), 1303. https://doi.org/10.3390/vaccines11081303