
Construction of a Triple-Gene Deletion Mutant of Orf Virus and Evaluation of Its Safety, Immunogenicity and Protective Efficacy

 , , ,
, , , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Cells and Viruses
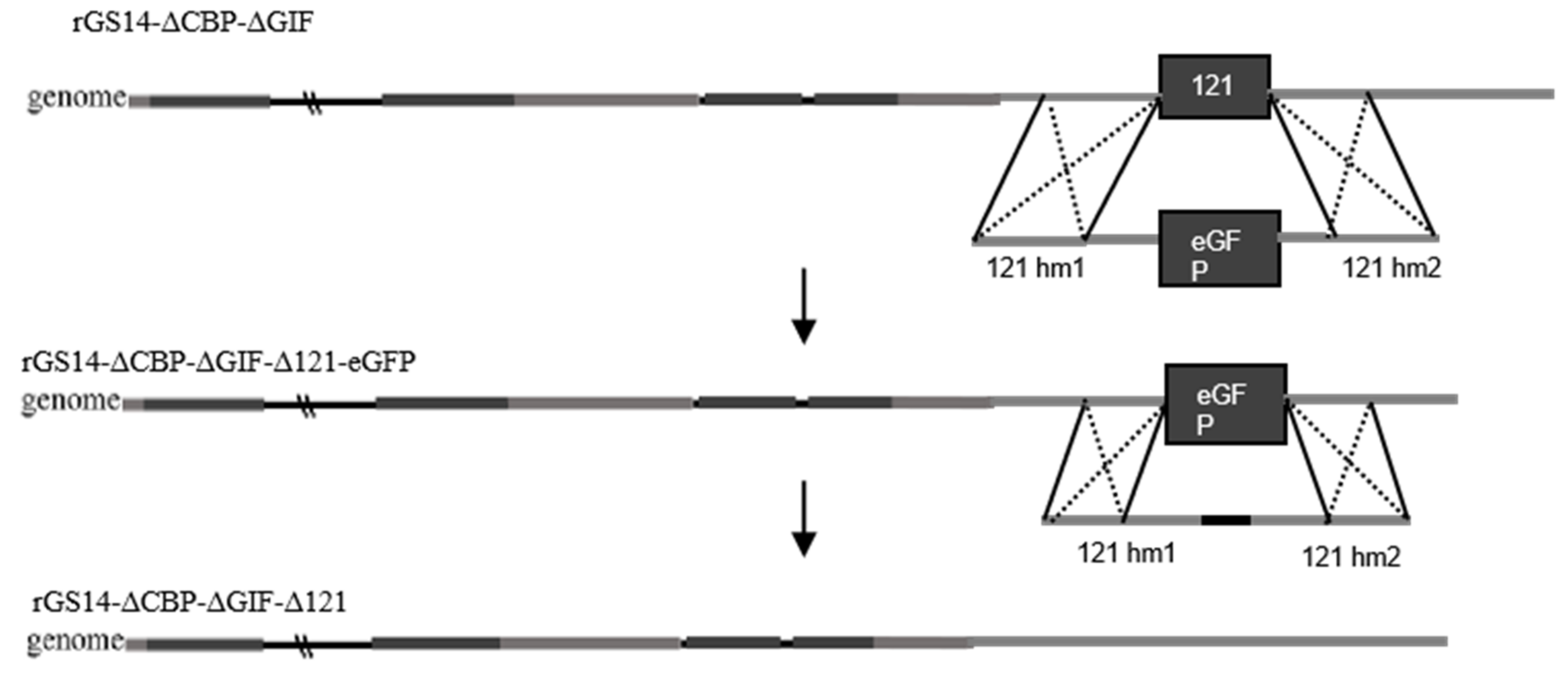
2.2. Construction of rGS14ΔCBPΔGIFΔ121

2.3. Animals
2.4. Virus Titration and Growth Curve with TCID50 Titration
2.5. Safety Test
2.6. Flow Cytometry Analysis
2.7. Cytokines Detection and Analysis
2.8. Comparison of Immune Protection of rGS14ΔCBPΔGIFΔ121
2.9. Statistical Analysis
3. Result
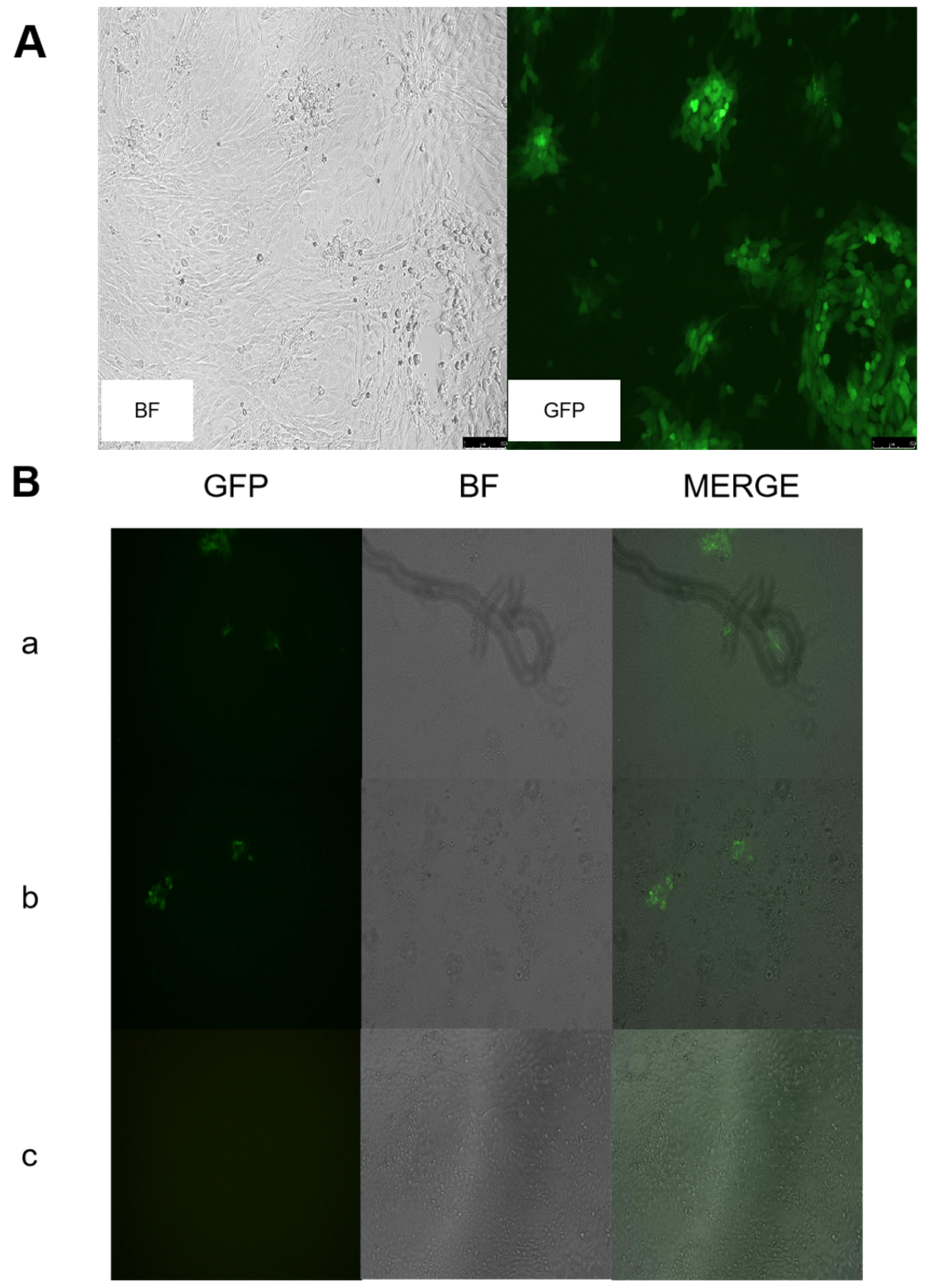
3.1. Construction of Recombinant Virus rGS14ΔCBPΔGIFΔ121
3.2. In Vitro Characterization of the Recombinant Virus
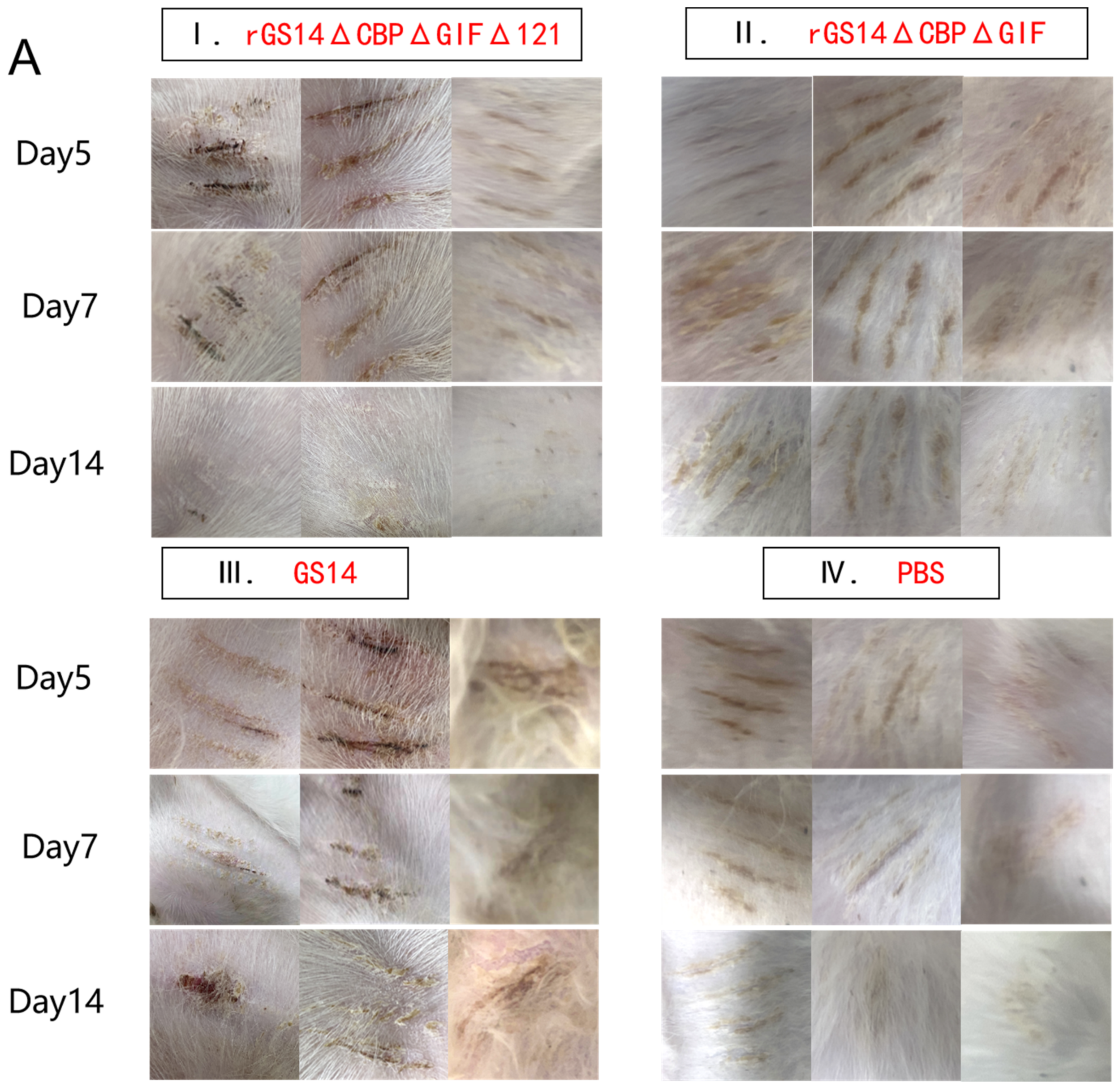
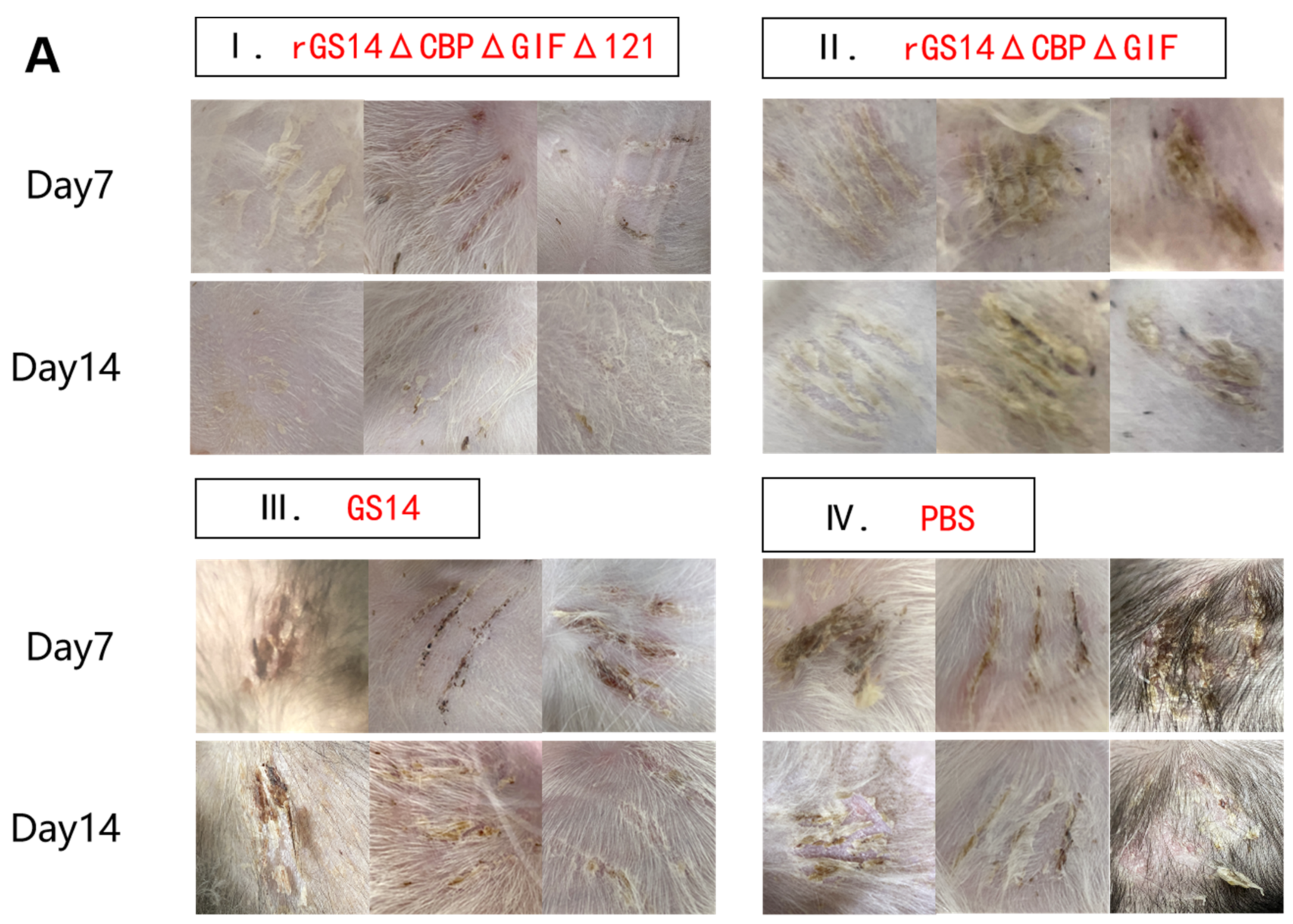
3.3. rGS14ΔCBPΔGIFΔ121 Group < rGS14ΔCBPΔGIF Group < GS14 Group in Terms of Virulence
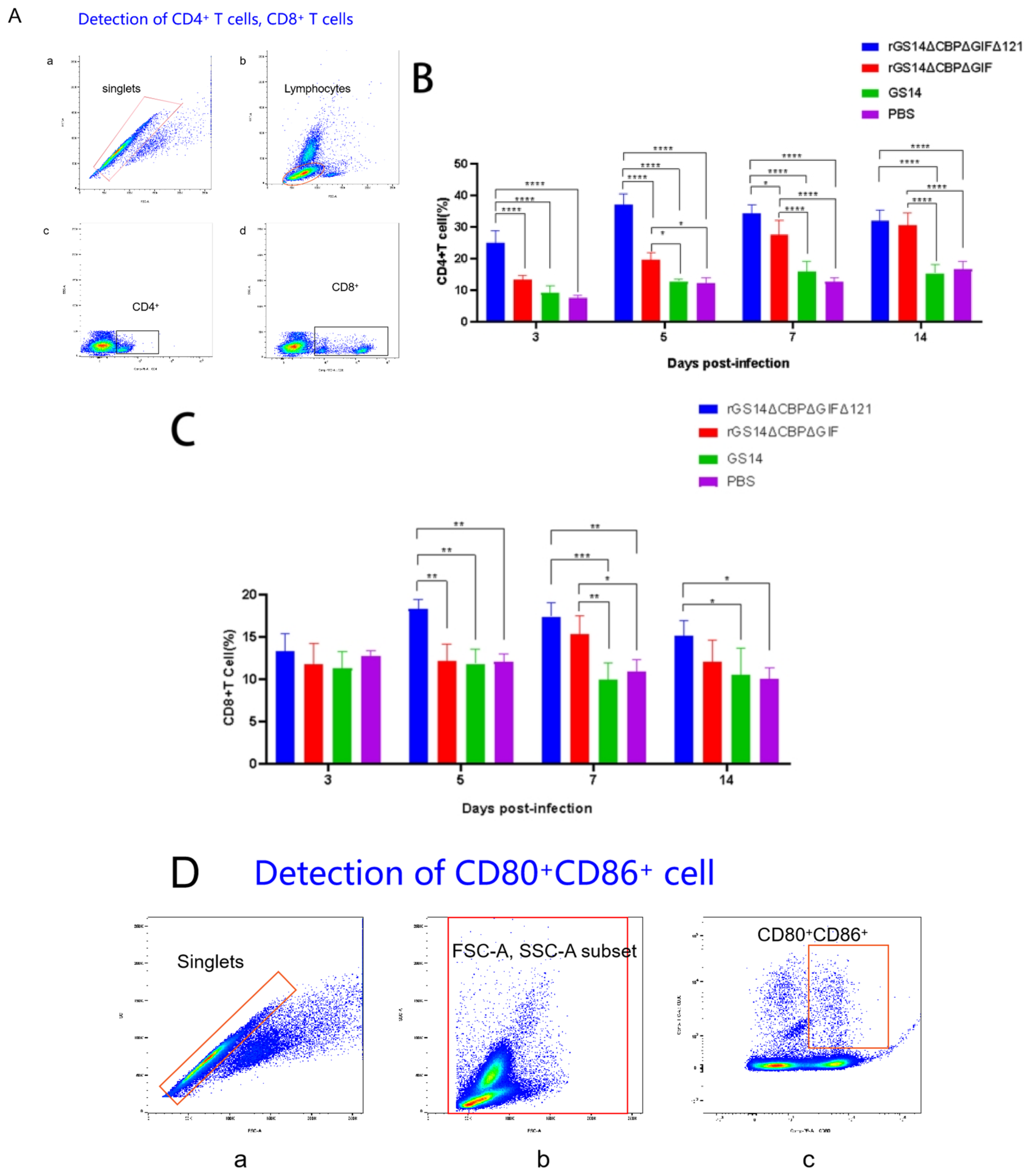
3.4. rGS14ΔCBPΔGIFΔ121 Causing High Levels of Expression of CD4+ Cells, CD8+ Cells and CD80+CD86+ Cells
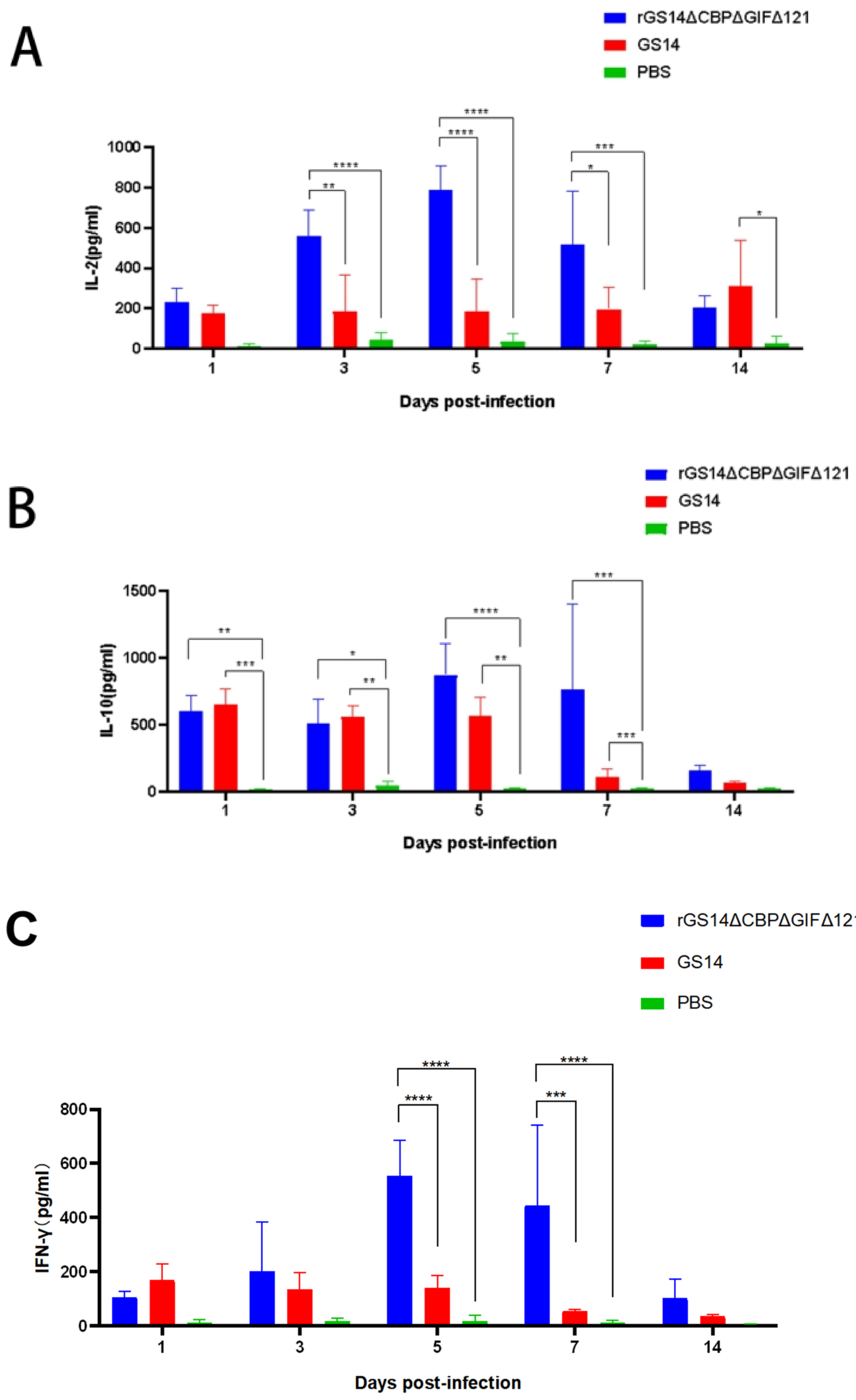
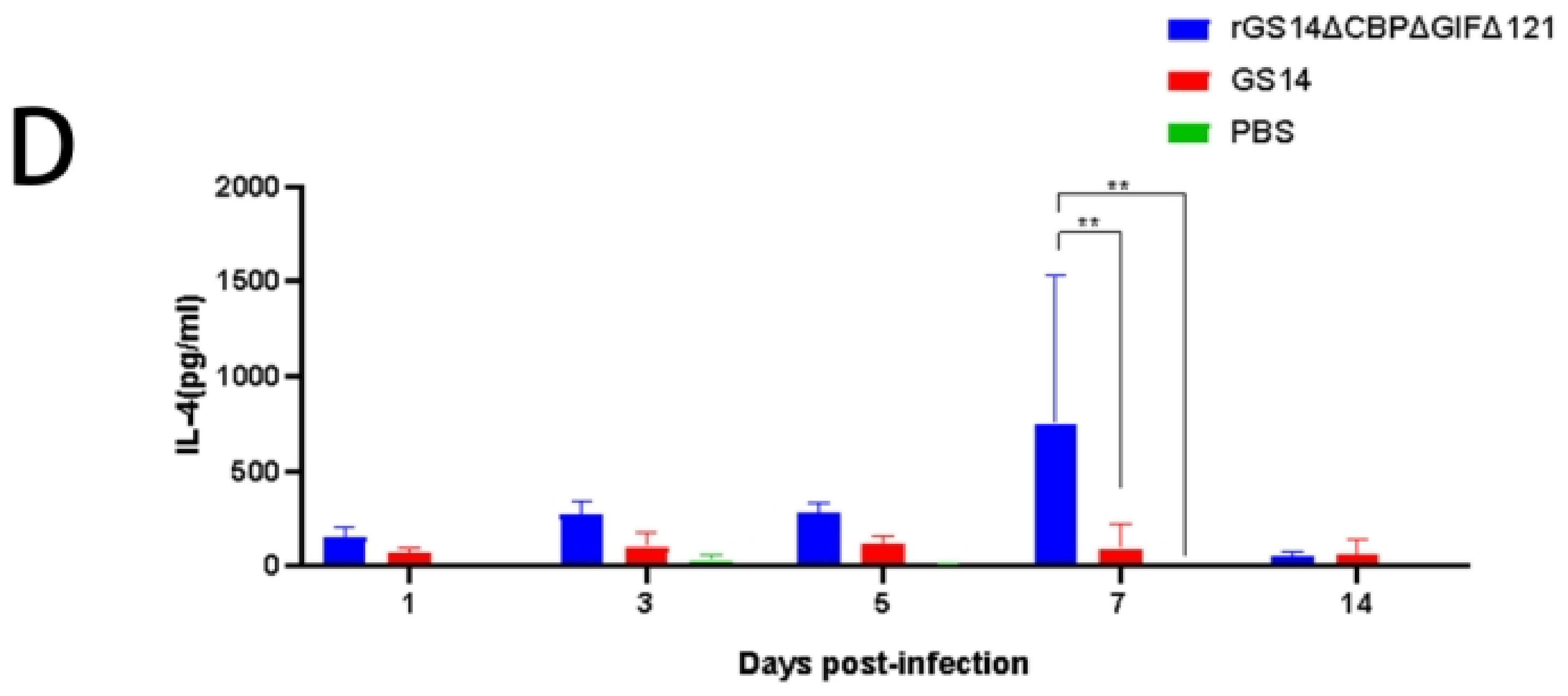
3.5. rGS14ΔCBPΔGIFΔ121 Induces High Levels of IL-2, IL-10 and IFN-γ Expression
3.6. rGS14-TrypMut > rGS14-DoubMut > GS14-w t > PBS in Terms of Protective Efficacy
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Fleming, S.B.; McCaughan, C.; Lateef, Z.; Dunn, A.; Wise, L.M.; Real, N.C.; Mercer, A.A. Deletion of the Chemokine Binding Protein Gene from the Parapoxvirus Orf Virus Reduces Virulence and Pathogenesis in Sheep. Front. Microbiol. 2017, 8, 46. [Google Scholar] [CrossRef]
- Deane, D.; Ueda, N.; Wise, L.M.; Wood, A.R.; Percival, A.; Jepson, C.; Inglis, N.F.; Fleming, S.B.; Mercer, A.A.; McInnes, C.J. Conservation and variation of the parapoxvirus GM-CSF-inhibitory factor (GIF) proteins. J. Gen. Virol. 2009, 90, 970–977. [Google Scholar] [CrossRef]
- Zhu, Z.; Qu, G.; Du, J.; Wang, C.; Chen, Y.; Shen, Z.; Zhou, Z.; Yin, C.; Chen, X. Construction and characterization of a contagious ecthyma virus double-gene deletion strain and evaluation of its potential as a live-attenuated vaccine in goat. Front. Immunol. 2022, 13, 961287. [Google Scholar] [CrossRef] [PubMed]
- Kassa, T. Erratum: Culex quinquefasciatus Egg Membrane Alteration and Ovicidal Activity of Cipadessa baccifera (Roth) Plant Extracts Compared to Synthetic Insect Growth Regulators [Corrigendum]. Res. Rep. Trop. Med. 2020, 11, 1. [Google Scholar] [CrossRef]
- Andreani, J.; Fongue, J.; Bou Khalil, J.Y.; David, L.; Mougari, S.; Le Bideau, M.; Abrahão, J.; Berbis, P.; La Scola, B. Human Infection with Orf Virus and Description of Its Whole Genome, France, 2017. Emerg. Infect. Dis. 2019, 25, 2197–2204. [Google Scholar] [CrossRef]
- Tryland, M.; Beckmen, K.B.; Burek-Huntington, K.A.; Breines, E.M.; Klein, J. Orf virus infection in Alaskan mountain goats, Dall’s sheep, muskoxen, caribou and Sitka black-tailed deer. Acta Vet. Scand. 2018, 60, 12. [Google Scholar] [CrossRef] [PubMed]
- Hosamani, M.; Scagliarini, A.; Bhanuprakash, V.; McInnes, C.J.; Singh, R.K. Orf: An update on current research and future perspectives. Expert Rev. Anti-Infect. Ther. 2009, 7, 879–893. [Google Scholar] [CrossRef] [PubMed]
- Paiba, G.A.; Thomas, D.R.; Morgan, K.L.; Bennett, M.; Salmon, R.L.; Chalmers, R.; Kench, S.M.; Coleman, T.J.; Meadows, D.; Morgan-Capner, P.; et al. Orf (contagious pustular dermatitis) in farmworkers: Prevalence and risk factors in three areas of England. Vet. Rec. 1999, 145, 7–11. [Google Scholar] [CrossRef]
- Feng, Q.; Li, L.; Du, G.; Liu, Y.; Liu, X.; Wu, J.; Shang, Y. Proteomic analyses reveal that Orf virus induces the activation and maturation of mouse bone marrow-derived dendritic cells. Res. Vet. Sci. 2020, 132, 563–573. [Google Scholar] [CrossRef] [PubMed]
- Jia, H.; Zhan, L.; Wang, X.; He, X.; Chen, G.; Zhang, Y.; Feng, Y.; Wei, Y.; Zhang, Y.; Jing, Z. Transcriptome analysis of sheep oral mucosa response to Orf virus infection. PLoS ONE 2017, 12, e0186681. [Google Scholar] [CrossRef]
- Karki, M.; Venkatesan, G.; Kumar, A.; Pandey, A.B. Genetic analysis of two viroceptor genes of orf virus. Arch. Virol. 2022, 167, 1577–1582. [Google Scholar] [CrossRef]
- Deane, D.; McInnes, C.J.; Percival, A.; Wood, A.; Thomson, J.; Lear, A.; Gilray, J.; Fleming, S.; Mercer, A.; Haig, D. Orf virus encodes a novel secreted protein inhibitor of granulocyte-macrophage colony-stimulating factor and interleukin-2. J. Virol. 2000, 74, 1313–1320. [Google Scholar] [CrossRef]
- Zhang, K.; Liu, Y.; Kong, H.; Shang, Y.; Liu, X. Comparison and phylogenetic analysis based on the B2L gene of orf virus from goats and sheep in China during 2009–2011. Arch. Virol. 2014, 159, 1475–1479. [Google Scholar] [CrossRef] [PubMed]
- Yogisharadhya, R.; Kumar, A.; Bhanuprakash, V.; Shivachandra, S.B. Evaluation of a recombinant major envelope protein (F1L) based indirect- ELISA for sero-diagnosis of orf in sheep and goats. J. Virol. Methods 2018, 261, 112–120. [Google Scholar] [CrossRef]
- Seet, B.T.; McCaughan, C.A.; Handel, T.M.; Mercer, A.; Brunetti, C.; McFadden, G.; Fleming, S.B. Analysis of an orf virus chemokine-binding protein: Shifting ligand specificities among a family of poxvirus viroceptors. Proc. Natl. Acad. Sci. USA 2003, 100, 15137–15142. [Google Scholar] [CrossRef]
- Fleming, S.B.; McCaughan, C.A.; Andrews, A.E.; Nash, A.D.; Mercer, A.A. A homolog of interleukin-10 is encoded by the poxvirus orf virus. J. Virol. 1997, 71, 4857–4861. [Google Scholar] [CrossRef]
- Mercer, A.A.; Fraser, K.; Barns, G.; Robinson, A.J. The structure and cloning of orf virus DNA. Virology 1987, 157, 1–12. [Google Scholar] [CrossRef]
- Fleming, S.B.; Wise, L.M.; Mercer, A.A. Molecular genetic analysis of orf virus: A poxvirus that has adapted to skin. Viruses 2015, 7, 1505–1539. [Google Scholar] [CrossRef] [PubMed]
- Diel, D.G.; Luo, S.; Delhon, G.; Peng, Y.; Flores, E.F.; Rock, D.L. Orf virus ORFV121 encodes a novel inhibitor of NF-kappaB that contributes to virus virulence. J. Virol. 2011, 85, 2037–2049. [Google Scholar] [CrossRef] [PubMed]
- Martins, M.; Joshi, L.R.; Rodrigues, F.S.; Anziliero, D.; Frandoloso, R.; Kutish, G.F.; Rock, D.L.; Weiblen, R.; Flores, E.F.; Diel, D.G. Immunogenicity of ORFV-based vectors expressing the rabies virus glycoprotein in livestock species. Virology 2017, 511, 229–239. [Google Scholar] [CrossRef]
- Rziha, H.; Henkel, M.; Cottone, R.; Bauer, B.; Auge, U.; Götz, F.; Pfaff, E.; Röttgen, M.; Dehio, C.; Büttner, M. Generation of recombinant parapoxviruses: Non-essential genes suitable for insertion and expression of foreign genes. J. Biotechnol. 2000, 83, 137–145. [Google Scholar] [CrossRef] [PubMed]
- Rziha, H.J.; Büttner, M.; Müller, M.; Salomon, F.; Reguzova, A.; Laible, D.; Amann, R. Genomic Characterization of Orf Virus Strain D1701-V (Parapoxvirus) and Development of Novel Sites for Multiple Transgene Expression. Viruses 2019, 11, 127. [Google Scholar] [CrossRef]
- Lacasta, D.; Reina, R.; Ruiz de Arcaute, M.; Ferrer, L.M.; Benito, A.A.; Tejedor, M.T.; Echeverria, I.; Ruiz, H.; Martinez Cardenas, S.; Windsor, P.A. Effect of a Topical Formulation on Infective Viral Load in Lambs Naturally Infected with Orf Virus. Vet. Med. 2021, 12, 149–158. [Google Scholar] [CrossRef]
- Zhao, K.; He, W.; Gao, W.; Lu, H.; Han, T.; Li, J.; Zhang, X.; Zhang, B.; Wang, G.; Su, G.; et al. Orf virus DNA vaccines expressing ORFV 011 and ORFV 059 chimeric protein enhances immunogenicity. Virol. J. 2011, 8, 562. [Google Scholar] [CrossRef]
- Mercer, A.A.; Wise, L.M.; Scagliarini, A.; McInnes, C.J.; Büttner, M.; Rziha, H.J.; McCaughan, C.A.; Fleming, S.B.; Ueda, N.; Nettleton, P.F. Vascular endothelial growth factors encoded by Orf virus show surprising sequence variation but have a conserved, functionally relevant structure. J. Gen. Virol. 2002, 83, 2845–2855. [Google Scholar] [CrossRef]
- Martins, M.; Rodrigues, F.S.; Joshi, L.R.; Jardim, J.C.; Flores, M.M.; Weiblen, R.; Flores, E.F.; Diel, D.G. Orf virus ORFV112, ORFV117 and ORFV127 contribute to ORFV IA82 virulence in sheep. Vet. Microbiol. 2021, 257, 109066. [Google Scholar] [CrossRef] [PubMed]
- Kumar, N.; Wadhwa, A.; Chaubey, K.K.; Singh, S.V.; Gupta, S.; Sharma, S.; Sharma, D.K.; Singh, M.K.; Mishra, A.K. Isolation and phylogenetic analysis of an orf virus from sheep in Makhdoom, India. Virus Genes 2014, 48, 312–319. [Google Scholar] [CrossRef]
- Martins, M.; Cargnelutti, J.F.; Weiblen, R.; Flores, E.F. Pathogenesis in lambs and sequence analysis of putative virulence genes of Brazilian orf virus isolates. Vet. Microbiol. 2014, 174, 69–77. [Google Scholar] [CrossRef]
- Da Costa, R.A.; Cargnelutti, J.F.; Schild, C.O.; Flores, E.F.; Riet-Correa, F.; Giannitti, F. Outbreak of contagious ecthyma caused by Orf virus (Parapoxvirus ovis) in a vaccinated sheep flock in Uruguay. Braz. J. Microbiol. 2019, 50, 565–569. [Google Scholar] [CrossRef] [PubMed]
- Bukar, A.M.; Jesse, F.F.A.; Abdullah, C.A.C.; Noordin, M.M.; Lawan, Z.; Mangga, H.K.; Balakrishnan, K.N.; Azmi, M.M. Immunomodulatory Strategies for Parapoxvirus: Current Status and Future Approaches for the Development of Vaccines against Orf Virus Infection. Vaccines 2021, 9, 1341. [Google Scholar] [CrossRef]
- Buddle, B.M.; Pulford, H.D. Effect of passively-acquired antibodies and vaccination on the immune response to contagious ecthyma virus. Vet. Microbiol. 1984, 9, 515–522. [Google Scholar] [CrossRef]
- Pye, D. Vaccination of sheep with cell culture grown orf virus. Aust. Vet. J. 1990, 67, 182–186. [Google Scholar] [CrossRef] [PubMed]
- Lee, N.H.; Lee, J.A.; Park, S.Y.; Song, C.S.; Choi, I.S.; Lee, J.B. A review of vaccine development and research for industry animals in Korea. Clin. Exp. Vaccine Res. 2012, 1, 18–34. [Google Scholar] [CrossRef] [PubMed]
- Gladue, D.P.; Ramirez-Medina, E.; Vuono, E.; Silva, E.; Rai, A.; Pruitt, S.; Espinoza, N.; Velazquez-Salinas, L.; Borca, M.V. Deletion of the A137R Gene from the Pandemic Strain of African Swine Fever Virus Attenuates the Strain and Offers Protection against the Virulent Pandemic Virus. J. Virol. 2021, 95, e0113921. [Google Scholar] [CrossRef]
- Struzik, J.; Szulc-Dąbrowska, L. NF-κB as an Important Factor in Optimizing Poxvirus-Based Vaccines against Viral Infections. Pathogens 2020, 9, 1001. [Google Scholar] [CrossRef]
- Lateef, Z.; Baird, M.A.; Wise, L.M.; Young, S.; Mercer, A.A.; Fleming, S.B. The chemokine-binding protein encoded by the poxvirus orf virus inhibits recruitment of dendritic cells to sites of skin inflammation and migration to peripheral lymph nodes. Cell. Microbiol. 2010, 12, 665–676. [Google Scholar] [CrossRef]
- McFadden, G.; Moyer, R. Parapoxvirus (Orf Virus) IL-10 Homolog; Arizona State University: Tempe, AZ, USA, 2014. [Google Scholar]
- Pasparakis, M. Regulation of tissue homeostasis by NF-kappaB signalling: Implications for inflammatory diseases. Nat. Rev. Immunol. 2009, 9, 778–788. [Google Scholar] [CrossRef] [PubMed]
- Weisfelner, M.E.; Gottlieb, A.B. The role of apoptosis in human epidermal keratinocytes. J. Drugs Dermatol. 2003, 2, 385–391. [Google Scholar]











Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shen, Z.; Liu, B.; Zhu, Z.; Du, J.; Zhou, Z.; Pan, C.; Chen, Y.; Yin, C.; Luo, Y.; Li, H.; et al. Construction of a Triple-Gene Deletion Mutant of Orf Virus and Evaluation of Its Safety, Immunogenicity and Protective Efficacy. Vaccines 2023, 11, 909. https://doi.org/10.3390/vaccines11050909
Shen Z, Liu B, Zhu Z, Du J, Zhou Z, Pan C, Chen Y, Yin C, Luo Y, Li H, et al. Construction of a Triple-Gene Deletion Mutant of Orf Virus and Evaluation of Its Safety, Immunogenicity and Protective Efficacy. Vaccines. 2023; 11(5):909. https://doi.org/10.3390/vaccines11050909
Chicago/Turabian StyleShen, Zhanning, Bo Liu, Zhen Zhu, Jige Du, Zhiyu Zhou, Chenfan Pan, Yong Chen, Chunsheng Yin, Yufeng Luo, Huanrong Li, and et al. 2023. "Construction of a Triple-Gene Deletion Mutant of Orf Virus and Evaluation of Its Safety, Immunogenicity and Protective Efficacy" Vaccines 11, no. 5: 909. https://doi.org/10.3390/vaccines11050909
APA StyleShen, Z., Liu, B., Zhu, Z., Du, J., Zhou, Z., Pan, C., Chen, Y., Yin, C., Luo, Y., Li, H., & Chen, X. (2023). Construction of a Triple-Gene Deletion Mutant of Orf Virus and Evaluation of Its Safety, Immunogenicity and Protective Efficacy. Vaccines, 11(5), 909. https://doi.org/10.3390/vaccines11050909