
Cross-Protection against Acute Staphylococcus aureus Lung Infection in Mice by a D-Glutamate Auxotrophic Vaccine Candidate

 , , and
, , and
Abstract
1. Introduction
2. Materials and Methods
2.1. Bacterial Strains and Culture Conditions
2.2. Ethics Statement
2.3. Inoculation with S. aureus 132 ΔmurI Δdat and Mice Sampling
2.4. Acute Lung Infection with Virulent S. aureus
2.5. Elisa
2.6. Statistical Analysis
3. Results
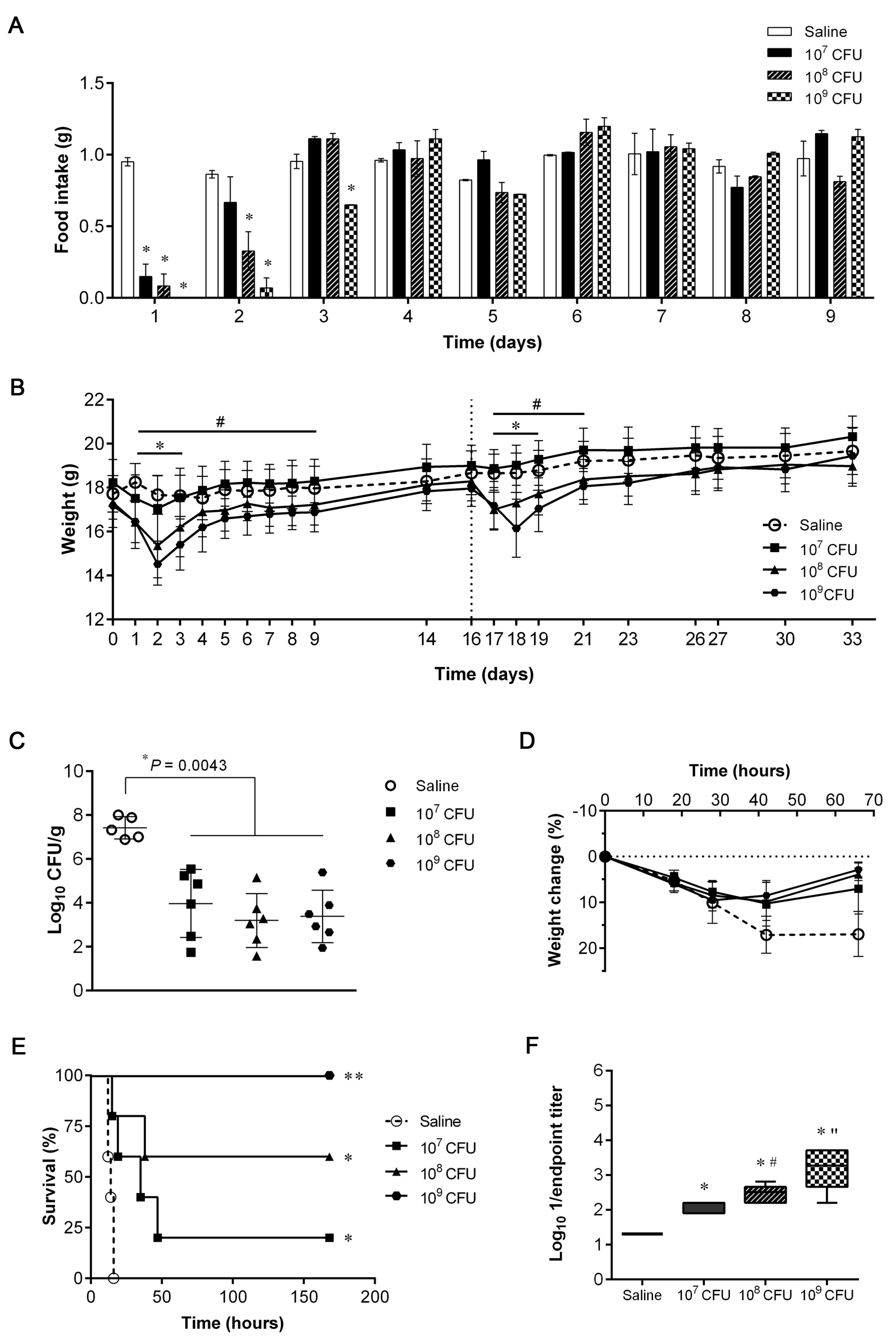
3.1. Dosage Escalation for Safety Assessment and Evaluation of Protective Efficacy
3.2. Humoral Immune Responses after IN Immunization with S. aureus 132 ΔmurI Δdat
3.3. Protective Efficacy Conferred by IN Immunization with S. aureus 132 ΔmurI Δdat against Acute Lung Infection Caused by Heterologous S. aureus
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lowy, F.D. Staphylococcus aureus infections. N. Engl. J. Med. 1998, 339, 520–532. [Google Scholar] [CrossRef] [PubMed]
- Tong, S.Y.; Davis, J.S.; Eichenberger, E.; Holland, T.L.; Fowler, V.G., Jr. Staphylococcus aureus infections: Epidemiology, pathophysiology, clinical manifestations, and management. Clin. Microbiol. Rev. 2015, 28, 603–661. [Google Scholar] [CrossRef]
- Farver, C.F.; Zander, D.S. Molecular Pathology: The Molecular Basis of Human Disease; Coleman, W.B., Tsongalis, G.J., Eds.; Elsevier, Inc.: Amsterdam, The Netherlands, 2009; Chapter 18; pp. 305–364. [Google Scholar]
- Clark, S.B.; Hicks, M.A. Staphylococcal Pneumonia. In StatPearls [Internet]; StatPearls Publishing: Treasure Island, FL, USA, 2022. [Google Scholar]
- Guillet, Y.; Issartel, B.; Vanhems, P.; Fournet, J.C.; Lina, G.; Bes, M.; Vandenesch, F.; Piémont, Y.; Brousse, N.; Floret, D.; et al. Association between Staphylococcus aureus strains carrying gene for Panton-Valentine leukocidin and highly lethal necrotising pneumonia in young immunocompetent patients. Lancet 2002, 359, 753–759. [Google Scholar] [CrossRef] [PubMed]
- Chambers, H.F.; Deleo, F.R. Waves of resistance: Staphylococcus aureus in the antibiotic era. Nat. Rev. Microbiol. 2009, 7, 629–641. [Google Scholar] [CrossRef] [PubMed]
- Clegg, J.; Soldaini, E.; McLoughlin, R.M.; Rittenhouse, S.; Bagnoli, F.; Phogat, S. Staphylococcus aureus Vaccine Research and Development: The Past, Present and Future, Including Novel Therapeutic Strategies. Front Immunol. 2021, 12, 705360. [Google Scholar] [CrossRef]
- Cabral, M.P.; García, P.; Beceiro, A.; Rumbo, C.; Pérez, A.; Moscoso, M.; Bou, G. Design of live attenuated bacterial vaccines based on D-glutamate auxotrophy. Nat. Commun. 2017, 8, 15480. [Google Scholar] [CrossRef]
- Vergara-Irigaray, M.; Valle, J.; Merino, N.; Latasa, C.; García, B.; Ruiz de Los Mozos, I.; Solano, C.; Toledo-Arana, A.; Penadés, J.R.; Lasa, I. Relevant role of fibronectin-binding proteins in Staphylococcus aureus biofilm-associated foreign-body infections. Infect. Immun. 2009, 77, 3978–3991. [Google Scholar] [CrossRef]
- Baba, T.; Bae, T.; Schneewind, O.; Takeuchi, F.; Hiramatsu, K. Genome sequence of Staphylococcus aureus strain Newman and comparative analysis of staphylococcal genomes: Polymorphism and evolution of two major pathogenicity islands. J. Bacteriol. 2008, 190, 300–310. [Google Scholar] [CrossRef]
- Diep, B.A.; Gill, S.R.; Chang, R.F.; Phan, T.H.; Chen, J.H.; Davidson, M.G.; Lin, F.; Lin, J.; Carleton, H.A.; Mongodin, E.F.; et al. Complete genome sequence of USA300, an epidemic clone of community-acquired meticillin-resistant Staphylococcus aureus. Lancet 2006, 367, 731–739. [Google Scholar] [CrossRef]
- Fluit, A.C.; Carpaij, N.; Majoor, E.A.; Weinstein, R.A.; Aroutcheva, A.; Rice, T.W.; Bonten, M.J.; Willems, R.J. Comparison of an ST80 MRSA strain from the USA with European ST80 strains. J. Antimicrob. Chemother. 2015, 70, 664–669. [Google Scholar] [CrossRef]
- GBD 2019 Antimicrobial Resistance Collaborators. Global mortality associated with 33 bacterial pathogens in 2019: A systematic analysis for the Global Burden of Disease Study 2019. Lancet 2022, 400, 2221–2248. [Google Scholar] [CrossRef]
- Antimicrobial Resistance Collaborators. Global burden of bacterial antimicrobial resistance in 2019: A systematic analysis. Lancet 2022, 399, 629–655. [Google Scholar] [CrossRef]
- Bubeck, W.J.; Schneewind, O. Vaccine protection against Staphylococcus aureus pneumonia. J. Exp. Med. 2008, 205, 287–294. [Google Scholar] [CrossRef]
- Spaulding, A.R.; Salgado-Pabón, W.; Merriman, J.A.; Stach, C.S.; Ji, Y.; Gillman, A.N.; Peterson, M.L.; Schlievert, P.M. Vaccination against Staphylococcus aureus pneumonia. J. Infect. Dis. 2014, 209, 1955–1962. [Google Scholar] [CrossRef]
- Wacker, M.; Wang, L.; Kowarik, M.; Dowd, M.; Lipowsky, G.; Faridmoayer, A.; Shields, K.; Park, S.; Alaimo, C.; Kelley, K.A.; et al. Prevention of Staphylococcus aureus infections by glycoprotein vaccines synthesized in Escherichia coli. J. Infect. Dis. 2014, 209, 1551–1561. [Google Scholar] [CrossRef]
- Choi, S.J.; Kim, M.H.; Jeon, J.; Kim, O.Y.; Choi, Y.; Seo, J.; Hong, S.W.; Lee, W.H.; Jeon, S.G.; Gho, Y.S.; et al. Active Immunization with Extracellular Vesicles Derived from Staphylococcus aureus Effectively Protects against Staphylococcal Lung Infections, Mainly via Th1 Cell-Mediated Immunity. PLoS ONE 2015, 10, e0136021. [Google Scholar] [CrossRef]
- Liu, Q.; Du, X.; Hong, X.; Li, T.; Zheng, B.; He, L.; Wang, Y.; Otto, M.; Li, M. Targeting surface protein SasX by active and passive vaccination to reduce Staphylococcus aureus colonization and infection. Infect. Immun. 2015, 83, 2168–2174. [Google Scholar] [CrossRef]
- Yang, L.Y.; Zhou, H.; Yang, Y.; Tong, Y.N.; Peng, L.S.; Zhu, B.H.; Diao, W.B.; Zeng, H.; Sun, H.W.; Zou, Q.M. Protective effects of a nanoemulsion adjuvant vaccine (2C-Staph/NE) administered intranasally against invasive Staphylococcus aureus pneumonia. RSC Adv. 2018, 8, 9996–10008. [Google Scholar] [CrossRef]
- Fan, X.; Li, N.; Xu, M.; Yang, D.; Wang, B. Intrapulmonary Vaccination Induces Long-lasting and Effective Pulmonary Immunity Against Staphylococcus aureus Pneumonia. J. Infect. Dis. 2021, 224, 903–913. [Google Scholar] [CrossRef]
- Kyd, J.M.; Foxwell, A.R.; Cripps, A.W. Mucosal immunity in the lung and upper airway. Vaccine 2001, 19, 2527–2533. [Google Scholar] [CrossRef]
- Lycke, N. Recent progress in mucosal vaccine development: Potential and limitations. Nat. Rev. Immunol. 2012, 12, 592–605. [Google Scholar] [CrossRef] [PubMed]
- Karauzum, H.; Datta, S.K. Adaptive Immunity Against Staphylococcus aureus. Curr. Top. Microbiol. Immunol. 2017, 409, 419–439. [Google Scholar] [CrossRef] [PubMed]
- Lin, L.; Ibrahim, A.S.; Xu, X.; Farber, J.M.; Avanesian, V.; Baquir, B.; Fu, Y.; French, S.W.; Edwards, J.E., Jr.; Spellberg, B. Th1-Th17 cells mediate protective adaptive immunity against Staphylococcus aureus and Candida albicans infection in mice. PLoS Pathog. 2009, 5, e1000703. [Google Scholar] [CrossRef] [PubMed]
- Narita, K.; Hu, D.L.; Mori, F.; Wakabayashi, K.; Iwakura, Y.; Nakane, A. Role of interleukin-17A in cell-mediated protection against Staphylococcus aureus infection in mice immunized with the fibrinogen-binding domain of clumping factor A. Infect. Immun. 2010, 78, 4234–4242. [Google Scholar] [CrossRef] [PubMed]
- Cho, J.S.; Pietras, E.M.; Garcia, N.C.; Ramos, R.I.; Farzam, D.M.; Monroe, H.R.; Magorien, J.E.; Blauvelt, A.; Kolls, J.K.; Cheung, A.L.; et al. IL-17 is essential for host defense against cutaneous Staphylococcus aureus infection in mice. J. Clin. Invest. 2010, 120, 1762–1773. [Google Scholar] [CrossRef]
- Montgomery, C.P.; Daniels, M.; Zhao, F.; Alegre, M.L.; Chong, A.S.; Daum, R.S. Protective immunity against recurrent Staphylococcus aureus skin infection requires antibody and interleukin-17A. Infect. Immun. 2014, 82, 2125–2134. [Google Scholar] [CrossRef]
- Mancini, F.; Monaci, E.; Lofano, G.; Torre, A.; Bacconi, M.; Tavarini, S.; Sammicheli, C.; Arcidiacono, L.; Galletti, B.; Laera, D.; et al. One Dose of Staphylococcus aureus 4C-Staph Vaccine Formulated with a Novel TLR7-Dependent Adjuvant Rapidly Protects Mice through Antibodies, Effector CD4+ T Cells, and IL-17A. PLoS ONE 2016, 11, e0147767. [Google Scholar] [CrossRef]
- Yang, L.; Cai, C.; Feng, Q.; Shi, Y.; Zuo, Q.; Yang, H.; Jing, H.; Wei, C.; Zhuang, Y.; Zou, Q.; et al. Protective efficacy of the chimeric Staphylococcus aureus vaccine candidate IC in sepsis and pneumonia models. Sci. Rep. 2016, 11, 20929. [Google Scholar] [CrossRef]
- Bröker, B.M.; Mrochen, D.; Péton, V. The T Cell Response to Staphylococcus aureus. Pathogens 2016, 17, 31. [Google Scholar] [CrossRef]
- Cheng, P.; Liu, T.; Zhou, W.Y.; Zhuang, Y.; Peng, L.S.; Zhang, J.Y.; Yin, Z.N.; Mao, X.H.; Guo, G.; Shi, Y.; et al. Role of gamma-delta T cells in host response against Staphylococcus aureus-induced pneumonia. BMC Immunol. 2012, 13, 38. [Google Scholar] [CrossRef]
- Kudva, A.; Scheller, E.V.; Robinson, K.M.; Crowe, C.R.; Choi, S.M.; Slight, S.R.; Khader, S.A.; Dubin, P.J.; Enelow, R.I.; Kolls, J.K.; et al. Influenza A inhibits Th17-mediated host defense against bacterial pneumonia in mice. J. Immunol. 2011, 186, 1666–1674. [Google Scholar] [CrossRef]
- Braverman, J.; Monk, I.R.; Ge, C.; Westall, G.P.; Stinear, T.P.; Wakim, L.M. Staphylococcus aureus specific lung resident memory CD4+ Th1 cells attenuate the severity of influenza virus induced secondary bacterial pneumonia. Mucosal Immunol. 2022, 15, 783–796. [Google Scholar] [CrossRef]
- Christensen, D.; Mortensen, R.; Rosenkrands, I.; Dietrich, J.; Andersen, P. Vaccine-induced Th17 cells are established as resident memory cells in the lung and promote local IgA responses. Mucosal Immunol. 2016, 10, 260–270. [Google Scholar] [CrossRef]
- Bubeck, W.J.; Patel, R.J.; Schneewind, O. Surface proteins and exotoxins are required for the pathogenesis of Staphylococcus aureus pneumonia. Infect. Immun. 2007, 75, 1040–1044. [Google Scholar] [CrossRef]
- Labandeira-Rey, M.; Couzon, F.; Boisset, S.; Brown, E.L.; Bes, M.; Benito, Y.; Barbu, E.M.; Vazquez, V.; Höök, M.; Etienne, J.; et al. Staphylococcus aureus Panton-Valentine leukocidin causes necrotizing pneumonia. Science 2007, 315, 1130–1133. [Google Scholar] [CrossRef]
- Owen, A.M.; Fults, J.B.; Patil, N.K.; Hernandez, A.; Bohannon, J.K. TLR Agonists as Mediators of Trained Immunity: Mechanistic Insight and Immunotherapeutic Potential to Combat Infection. Front Immunol. 2021, 18, 622614. [Google Scholar] [CrossRef]
- Kim, T.H.; Kim, D.; Gautam, A.; Lee, H.; Kwak, M.H.; Park, M.C.; Park, S.; Wu, G.; Lee, B.L.; Lee, Y.; et al. CpG-DNA exerts antibacterial effects by protecting immune cells and producing bacteria-reactive antibodies. Sci. Rep. 2018, 2, 16236. [Google Scholar] [CrossRef]
- Bagnoli, F.; Fontana, M.R.; Soldaini, E.; Mishra, R.P.; Fiaschi, L.; Cartocci, E.; Nardi-Dei, V.; Ruggiero, P.; Nosari, S.; De Falco, M.G.; et al. Vaccine composition formulated with a novel TLR7-dependent adjuvant induces high and broad protection against Staphylococcus aureus. Proc. Natl. Acad. Sci. USA 2015, 24, 3680–3685. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Strain | Relevant Features | Reference |
|---|---|---|
| 132 | MRSA clinical isolate, PVL-positive | [9] |
| 132 ΔmurI Δdat | 132 derivative, MurI–, Dat–deficient | [8] |
| 132 Δspa | 132 derivative, SpaA (protein A)–deficient | [9] |
| Newman | MSSA, isolated from human infection | [10] |
| FPR3757 (USA300LAC) | CA-MRSA, isolated from wrist abscess, USA300 clone ST8, PVL-positive | [11] |
| S1475 | CA-MRSA, isolate from necrotizing pneumonia, European clone ST80, PVL-positive | [12] |
| Sa07365 | MSSA, isolated from staphylococcal pneumonia and co-infection with influenza A virus | Laboratory Collection, CHUAC |
| Time (h) Post-Inoculation | |||
|---|---|---|---|
| Dose (CFUs/Mouse) | 24 | 48 | 72 |
| 107 | 4.24 ± 0.33 (4/4) 1 | 2.30 ± 0.20 (3/4) | 0.00 ± 0.00 (0/4) |
| 108 | 5.78 ± 0.34 (4/4) | 3.62 ± 0.47 (4/4) | 0.00 ± 0.00 (0/4) |
| 109 | 7.07 ± 0.29 (4/4) | 4.27 ± 0.41 (4/4) | 2.57 ± 0.31 (3/4) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
García, P.; Cabral, M.P.; Beceiro, A.; Moscoso, M.; Bou, G. Cross-Protection against Acute Staphylococcus aureus Lung Infection in Mice by a D-Glutamate Auxotrophic Vaccine Candidate. Vaccines 2023, 11, 210. https://doi.org/10.3390/vaccines11020210
García P, Cabral MP, Beceiro A, Moscoso M, Bou G. Cross-Protection against Acute Staphylococcus aureus Lung Infection in Mice by a D-Glutamate Auxotrophic Vaccine Candidate. Vaccines. 2023; 11(2):210. https://doi.org/10.3390/vaccines11020210
Chicago/Turabian StyleGarcía, Patricia, Maria P. Cabral, Alejandro Beceiro, Miriam Moscoso, and Germán Bou. 2023. "Cross-Protection against Acute Staphylococcus aureus Lung Infection in Mice by a D-Glutamate Auxotrophic Vaccine Candidate" Vaccines 11, no. 2: 210. https://doi.org/10.3390/vaccines11020210
APA StyleGarcía, P., Cabral, M. P., Beceiro, A., Moscoso, M., & Bou, G. (2023). Cross-Protection against Acute Staphylococcus aureus Lung Infection in Mice by a D-Glutamate Auxotrophic Vaccine Candidate. Vaccines, 11(2), 210. https://doi.org/10.3390/vaccines11020210