
Self−Assembling Nanovaccine Fused with Flagellin Enhances Protective Effect against Foot−and−Mouth Disease Virus


 and
and 
Abstract
:1. Introduction
2. Materials and Methods
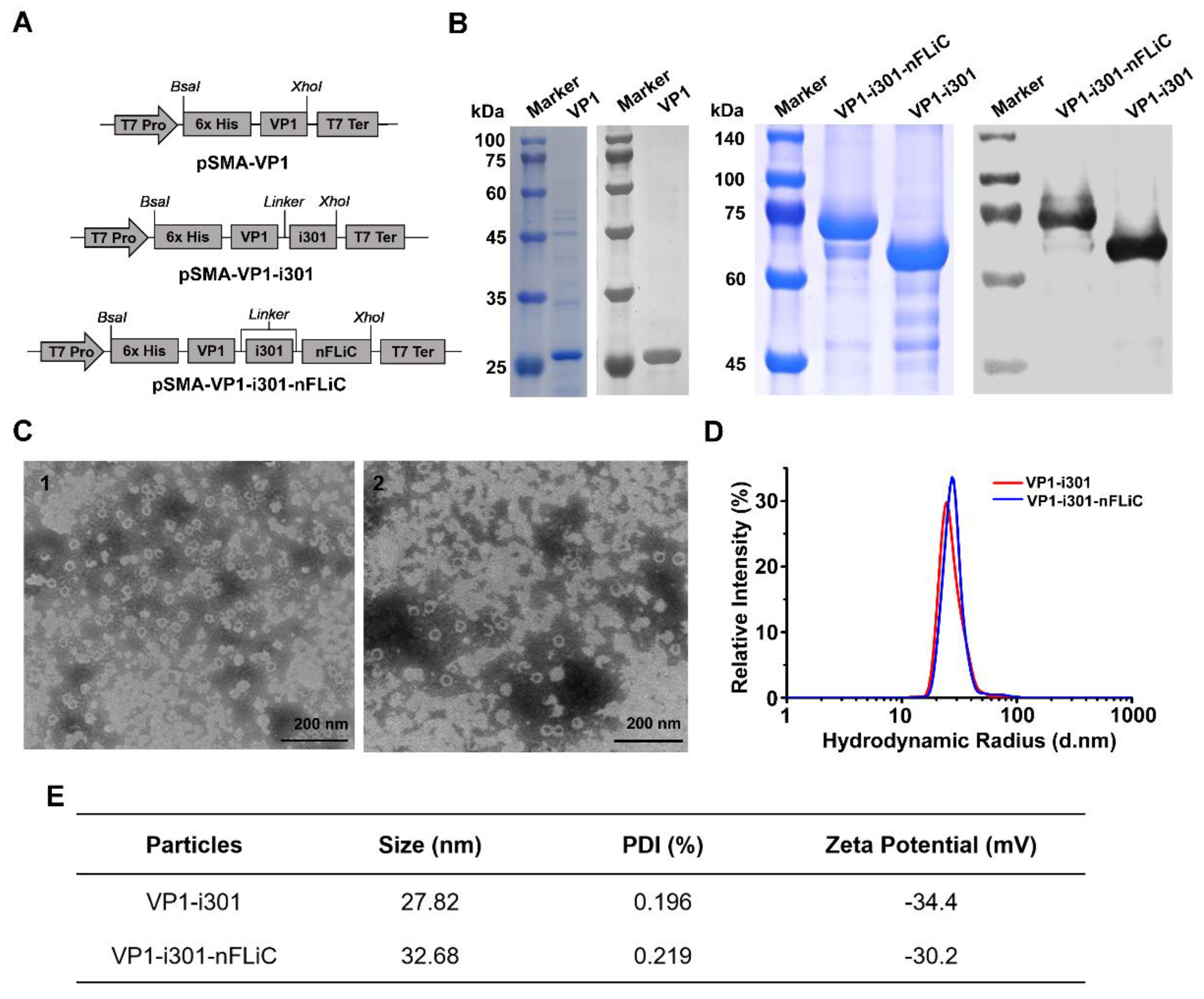
2.1. Construction of Recombinant Positive Plasmids
2.2. Expression and Purification of Recombinant Proteins
2.3. Physicochemical Conditions and Properties of NPs
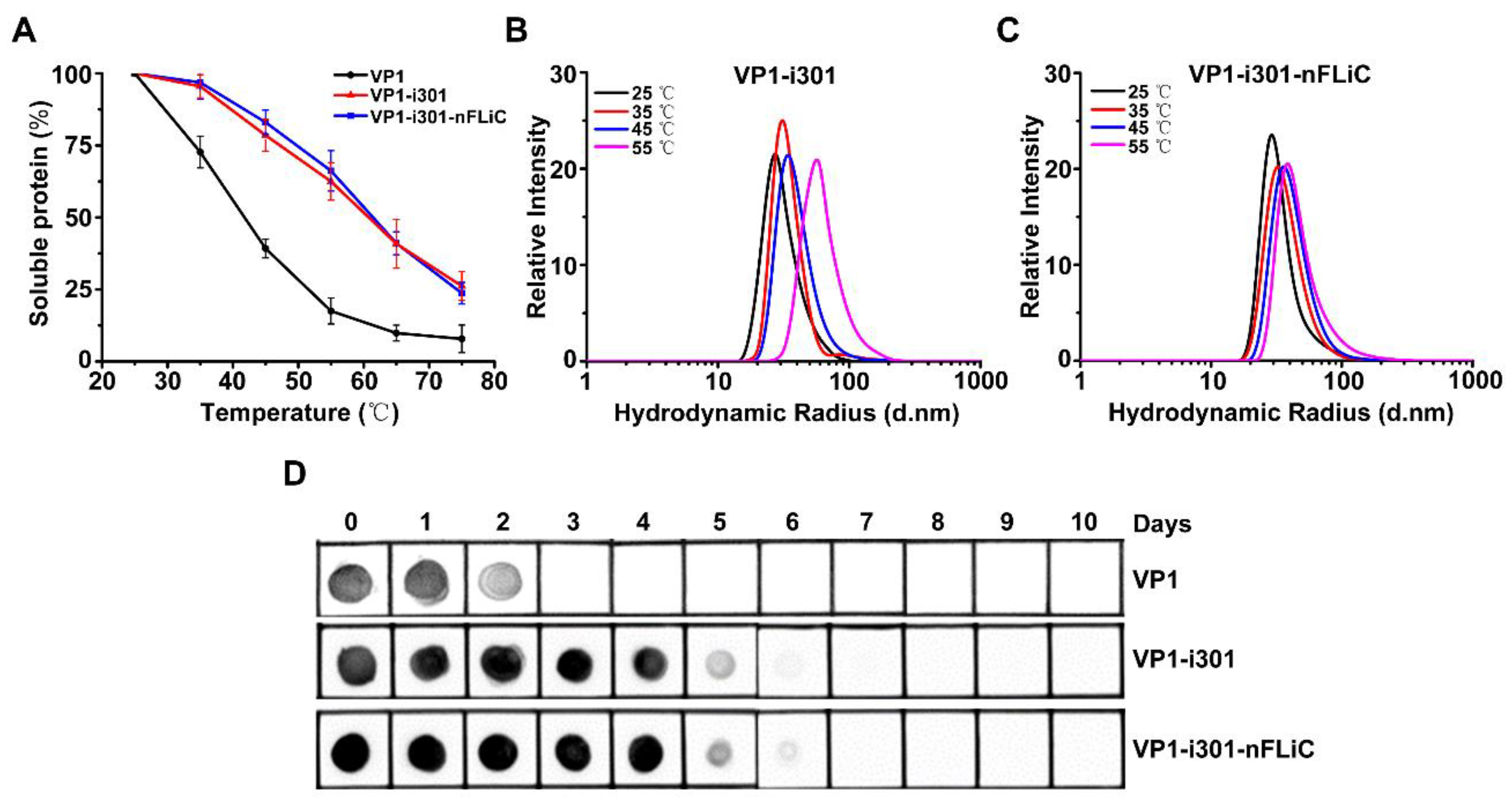
2.4. Thermal Stability Detection of NPs
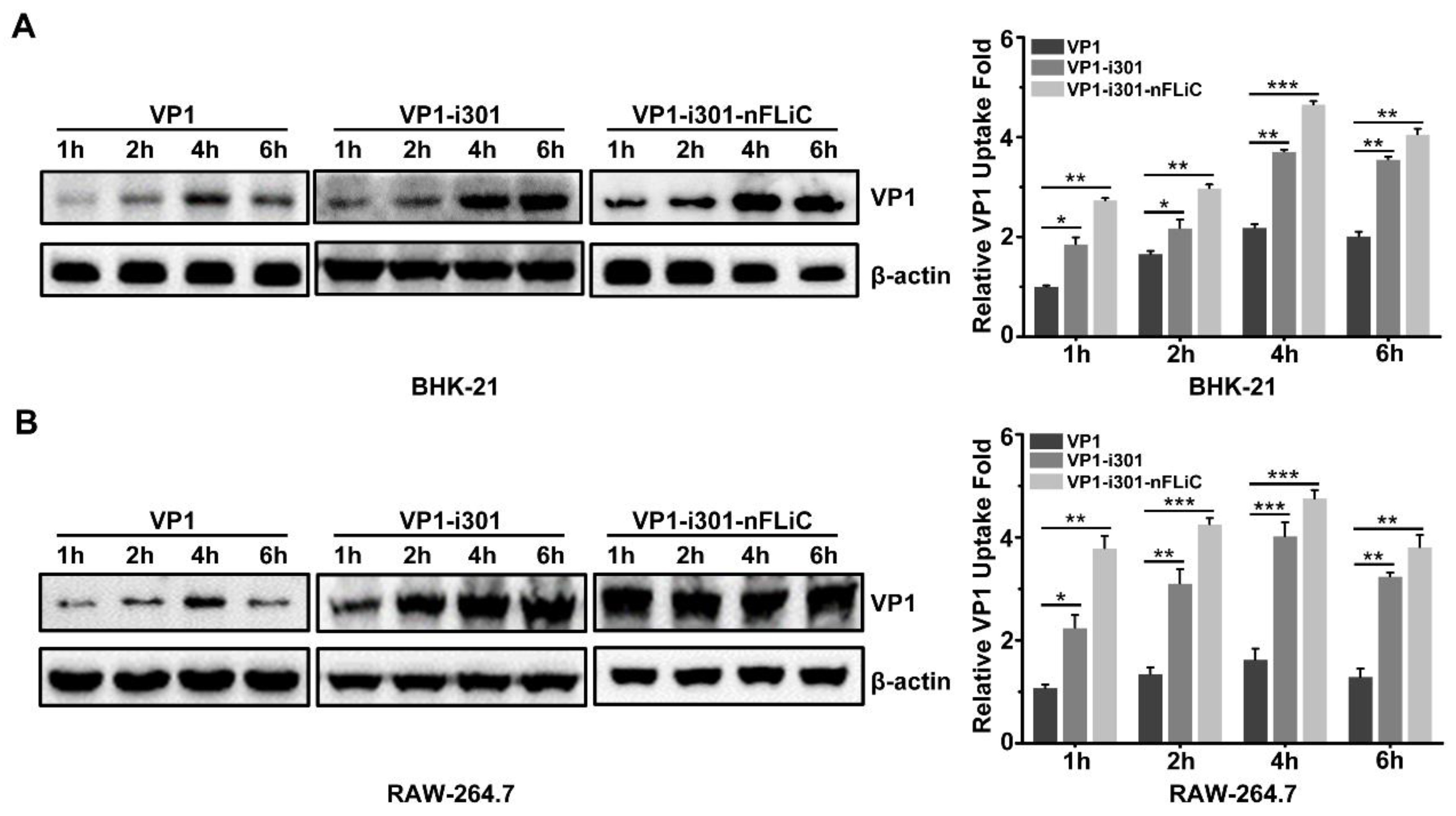
2.5. Cellular Uptake Assay
2.6. Separation and Generation of Bone−Marrow−Derived Cells (BMDCs)
2.7. TLR5 Pathway Activation Assay
2.8. Expression of Marker Genes and Cytokines by BMDCs
2.9. Animal Immunization and Challenge Trial
2.10. T lymphocyte Proliferation Response
2.11. Statistical Analysis
3. Results
3.1. Expression and Characterization of NPs
3.2. Thermal Stability Analysis of NPs
3.3. Cellular Uptake with NPs
3.4. NPs Induce TLR5−Mediated Inflammation in BMDCs
3.5. NPs Stimulated Cytokine Production in BMDCs
3.6. Enhanced Immune Response Induced by Nanovaccines in Guinea Pigs
3.7. Protective Efficacy against FMDV Challenge Stimulated by Nanovaccines
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Poonsuk, K.; Gimenez−Lirola, L.; Zimmerman, J.J. A review of foot−and−mouth disease virus (FMDV) testing in livestock with an emphasis on the use of alternative diagnostic specimens. Anim. Health Res. Rev. 2018, 19, 100–112. [Google Scholar] [CrossRef] [PubMed]
- Brown, F. Foot−and−mouth disease—One of the remaining great plagues. Proc. R. Soc. Lond. B Biol. Sci. 1986, 229, 215–226. [Google Scholar] [PubMed]
- Cox, J. Foot and mouth disease. Br. J. Gen. Pract. 2001, 51, 417. [Google Scholar] [PubMed]
- Frenkel, H.S. Research on foot−and−mouth disease. III. The cultivation of the virus on a practical scale in explantations of bovine tongue epithelium. Am. J. Vet. Res. 1951, 12, 187–190. [Google Scholar]
- Grubman, M.J.; Baxt, B. Foot−and−mouth disease. Clin. Microbiol. Rev. 2004, 17, 465–493. [Google Scholar] [CrossRef]
- Knight−Jones, T.J.; Rushton, J. The economic impacts of foot and mouth disease—What are they, how big are they and where do they occur? Prev. Vet. Med. 2013, 112, 161–173. [Google Scholar] [CrossRef]
- Condy, J.B.; Hedger, R.S.; Hamblin, C.; Barnett, I.T. The duration of the foot−and−mouth disease virus carrier state in African buffalo (i) in the individual animal and (ii) in a free−living herd. Comp. Immunol. Microbiol. Infect. Dis. 1985, 8, 259–265. [Google Scholar] [CrossRef]
- Li, P.; Lu, Z.; Bai, X.; Li, D.; Sun, P.; Bao, H.; Fu, Y.; Cao, Y.; Chen, Y.; Xie, B.; et al. Evaluation of a 3A−truncated foot−and−mouth disease virus in pigs for its potential as a marker vaccine. Vet. Res. 2014, 45, 51. [Google Scholar] [CrossRef]
- Karch, C.P.; Burkhard, P. Vaccine technologies: From whole organisms to rationally designed protein assemblies. Biochem. Pharmacol. 2016, 120, 1–14. [Google Scholar] [CrossRef]
- Skwarczynski, M.; Toth, I. Recent advances in peptide−based subunit nanovaccines. Nanomedicine 2014, 9, 2657–2669. [Google Scholar] [CrossRef]
- Negahdaripour, M.; Golkar, N.; Hajighahramani, N.; Kianpour, S.; Nezafat, N.; Ghasemi, Y. Harnessing self−assembled peptide nanoparticles in epitope vaccine design. Biotechnol. Adv. 2017, 35, 575–596. [Google Scholar] [CrossRef] [PubMed]
- Yang, F.; Mariz, F.C.; Zhao, X.; Spagnoli, G.; Ottonello, S.; Muller, M. Broad Neutralization Responses against Oncogenic Human Papillomaviruses Induced by a Minor Capsid L2 Polytope Genetically Incorporated into Bacterial Ferritin Nanoparticles. Front. Immunol. 2020, 11, 606569. [Google Scholar] [CrossRef]
- Rae, C.S.; Khor, I.W.; Wang, Q.; Destito, G.; Gonzalez, M.J.; Singh, P.; Thomas, D.M.; Estrada, M.N.; Powell, E.; Finn, M.G.; et al. Systemic trafficking of plant virus nanoparticles in mice via the oral route. Virology 2005, 343, 224–235. [Google Scholar] [CrossRef] [PubMed]
- Lewis, J.D.; Destito, G.; Zijlstra, A.; Gonzalez, M.J.; Quigley, J.P.; Manchester, M.; Stuhlmann, H. Viral nanoparticles as tools for intravital vascular imaging. Nat. Med. 2006, 12, 354–360. [Google Scholar] [CrossRef]
- Cai, Y.; Wang, Y.; Xu, H.; Cao, C.; Zhu, R.; Tang, X.; Zhang, T.; Pan, Y. Positive magnetic resonance angiography using ultrafine ferritin−based iron oxide nanoparticles. Nanoscale 2019, 11, 2644–2654. [Google Scholar] [CrossRef]
- Zhen, Z.; Tang, W.; Zhang, W.; Xie, J. Folic acid conjugated ferritins as photosensitizer carriers for photodynamic therapy. Nanoscale 2015, 7, 10330–10333. [Google Scholar] [CrossRef] [PubMed]
- Suthiwangcharoen, N.; Li, T.; Li, K.; Thompson, P.; You, S.J.; Wang, Q. M13 Bacteriophage−Polymer Nanoassemblies as Drug Delivery Vehicles. Nano Res. 2011, 4, 483–493. [Google Scholar] [CrossRef]
- Sun, X.; Li, W.; Zhang, X.; Qi, M.; Zhang, Z.; Zhang, X.E.; Cui, Z. In Vivo Targeting and Imaging of Atherosclerosis Using Multifunctional Virus−like Particles of Simian Virus 40. Nano Lett. 2016, 16, 6164–6171. [Google Scholar] [CrossRef]
- Clarke, B.E.; Newton, S.E.; Carroll, A.R.; Francis, M.J.; Appleyard, G.; Syred, A.D.; Highfield, P.E.; Rowlands, D.J.; Brown, F. Improved immunogenicity of a peptide epitope after fusion to hepatitis B core protein. Nature 1987, 330, 381–384. [Google Scholar] [CrossRef]
- Zhao, L.; Seth, A.; Wibowo, N.; Zhao, C.X.; Mitter, N.; Yu, C.; Middelberg, A.P. Nanoparticle vaccines. Vaccine 2014, 32, 327–337. [Google Scholar] [CrossRef]
- Paavonen, J.; Jenkins, D.; Bosch, F.X.; Naud, P.; Salmeron, J.; Wheeler, C.M.; Chow, S.N.; Apter, D.L.; Kitchener, H.C.; Castellsague, X.; et al. Efficacy of a prophylactic adjuvanted bivalent L1 virus−like−particle vaccine against infection with human papillomavirus types 16 and 18 in young women: An interim analysis of a phase III double−blind, randomised controlled trial. Lancet 2007, 369, 2161–2170. [Google Scholar] [CrossRef]
- Michel, M.L.; Tiollais, P. Hepatitis B vaccines: Protective efficacy and therapeutic potential. Pathol. Biol. 2010, 58, 288–295. [Google Scholar] [CrossRef]
- Guo, H.C.; Sun, S.Q.; Jin, Y.; Yang, S.L.; Wei, Y.Q.; Sun, D.H.; Yin, S.H.; Ma, J.W.; Liu, Z.X.; Guo, J.H.; et al. Foot−and−mouth disease virus−like particles produced by a SUMO fusion protein system in Escherichia coli induce potent protective immune responses in guinea pigs, swine and cattle. Vet. Res. 2013, 44, 48. [Google Scholar] [CrossRef]
- Pasick, J. Application of DIVA vaccines and their companion diagnostic tests to foreign animal disease eradication. Anim. Health Res. Rev. 2004, 5, 257–262. [Google Scholar] [CrossRef]
- Mignaqui, A.C.; Ruiz, V.; Durocher, Y.; Wigdorovitz, A. Advances in novel vaccines for foot and mouth disease: Focus on recombinant empty capsids. Crit. Rev. Biotechnol. 2019, 39, 306–320. [Google Scholar] [CrossRef]
- Teng, Z.; Sun, S.; Luo, X.; Zhang, Z.; Seo, H.; Xu, X.; Huang, J.; Dong, H.; Mu, S.; Du, P.; et al. Bi−functional gold nanocages enhance specific immunological responses of foot−and−mouth disease virus−like particles vaccine as a carrier and adjuvant. Nanomedicine 2021, 33, 102358. [Google Scholar] [CrossRef]
- Du, P.; Liu, R.; Sun, S.; Dong, H.; Zhao, R.; Tang, R.; Dai, J.; Yin, H.; Luo, J.; Liu, Z.; et al. Biomineralization improves the thermostability of foot−and−mouth disease virus−like particles and the protective immune response induced. Nanoscale 2019, 11, 22748–22761. [Google Scholar] [CrossRef]
- Hsia, Y.; Bale, J.B.; Gonen, S.; Shi, D.; Sheffler, W.; Fong, K.K.; Nattermann, U.; Xu, C.; Huang, P.S.; Ravichandran, R.; et al. Corrigendum: Design of a hypersTable 60−subunit protein icosahedron. Nature 2016, 540, 150. [Google Scholar] [CrossRef]
- Gutjahr, A.; Phelip, C.; Coolen, A.L.; Monge, C.; Boisgard, A.S.; Paul, S.; Verrier, B. Biodegradable Polymeric Nanoparticles−Based Vaccine Adjuvants for Lymph Nodes Targeting. Vaccines 2016, 4, 34. [Google Scholar] [CrossRef]
- Kumar, S.; Anselmo, A.C.; Banerjee, A.; Zakrewsky, M.; Mitragotri, S. Shape and size−dependent immune response to antigen−carrying nanoparticles. J. Control Release 2015, 220 Pt A, 141–148. [Google Scholar] [CrossRef]
- Stano, A.; Nembrini, C.; Swartz, M.A.; Hubbell, J.A.; Simeoni, E. Nanoparticle size influences the magnitude and quality of mucosal immune responses after intranasal immunization. Vaccine 2012, 30, 7541–7546. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.N.; Lazarovits, J.; Poon, W.; Ouyang, B.; Nguyen, L.N.M.; Kingston, B.R.; Chan, W.C.W. Nanoparticle Size Influences Antigen Retention and Presentation in Lymph Node Follicles for Humoral Immunity. Nano Lett. 2019, 19, 7226–7235. [Google Scholar] [CrossRef] [PubMed]
- Fan, C.; Han, J.; Liu, X.; Zhang, F.; Long, Y.; Xie, Q. Modulation of hypoxia−inducible factor−1alpha/cyclo−oxygenase−2 pathway associated with attenuation of intestinal mucosa inflammatory damage by Acanthopanax senticosus polysaccharides in lipopolysaccharide−challenged piglets. Br. J. Nutr. 2019, 122, 666–675. [Google Scholar] [CrossRef] [PubMed]
- Bo, R.; Zheng, S.; Xing, J.; Luo, L.; Niu, Y.; Huang, Y.; Liu, Z.; Hu, Y.; Liu, J.; Wu, Y.; et al. The immunological activity of Lycium barbarum polysaccharides liposome in vitro and adjuvanticity against PCV2 in vivo. Int. J. Biol. Macromol. 2016, 85, 294–301. [Google Scholar]
- Zhang, C.; Wang, B.; Wang, M. GM−CSF and IL−2 as adjuvant enhance the immune effect of protein vaccine against foot−and−mouth disease. Virol. J. 2011, 8, 7. [Google Scholar]
- Xu, X.; Du, L.; Fan, B.; Sun, B.; Zhou, J.; Guo, R.; Yu, Z.; Shi, D.; He, K.; Li, B. A flagellin−adjuvanted inactivated porcine epidemic diarrhea virus (PEDV) vaccine provides enhanced immune protection against PEDV challenge in piglets. Arch. Virol. 2020, 165, 1299–1309. [Google Scholar]
- Yonekura, K.; Maki−Yonekura, S.; Namba, K. Complete atomic model of the bacterial flagellar filament by electron cryomicroscopy. Nature 2003, 424, 643–650. [Google Scholar] [CrossRef] [PubMed]
- Cui, B.; Liu, X.; Fang, Y.; Zhou, P.; Zhang, Y.; Wang, Y. Flagellin as a vaccine adjuvant. Expert. Rev. Vaccines 2018, 17, 335–349. [Google Scholar] [CrossRef]
- Khani, M.H.; Bagheri, M.; Zahmatkesh, A.; Moradi Bidhendi, S. Immunostimulatory effects of truncated and full−length flagellin recombinant proteins. Microb. Pathog. 2019, 127, 190–197. [Google Scholar] [CrossRef]
- Panda, A.K. Induction of anti−tumor immunity and T−cell responses using nanodelivery systems engrafting TLR−5 ligand. Expert. Rev. Vaccines 2011, 10, 155–157. [Google Scholar] [CrossRef]
- Mizel, S.B.; Honko, A.N.; Moors, M.A.; Smith, P.S.; West, A.P. Induction of macrophage nitric oxide production by Gram−negative flagellin involves signaling via heteromeric Toll−like receptor 5/Toll−like receptor 4 complexes. J. Immunol. 2003, 170, 6217–6223. [Google Scholar]
- Akira, S.; Takeda, K. Toll−like receptor signalling. Nat. Rev. Immunol. 2004, 4, 499–511. [Google Scholar] [PubMed]
- Song, W.S.; Jeon, Y.J.; Namgung, B.; Hong, M.; Yoon, S.I. A conserved TLR5 binding and activation hot spot on flagellin. Sci. Rep. 2017, 7, 40878. [Google Scholar] [PubMed]
- Smith, K.D.; Andersen−Nissen, E.; Hayashi, F.; Strobe, K.; Bergman, M.A.; Barrett, S.L.; Cookson, B.T.; Aderem, A. Toll−like receptor 5 recognizes a conserved site on flagellin required for protofilament formation and bacterial motility. Nat. Immunol. 2003, 4, 1247–1253. [Google Scholar] [PubMed]
- Doan, T.D.; Wang, H.Y.; Ke, G.M.; Cheng, L.T. N−terminus of Flagellin Fused to an Antigen Improves Vaccine Efficacy against Pasteurella multocida Infection in Chickens. Vaccines 2020, 8, 283. [Google Scholar] [CrossRef] [PubMed]
- Eaves−Pyles, T.D.; Wong, H.R.; Odoms, K.; Pyles, R.B. Salmonella flagellin−dependent proinflammatory responses are localized to the conserved amino and carboxyl regions of the protein. J. Immunol. 2001, 167, 7009–7016. [Google Scholar] [PubMed]
- Zheng, H.; He, J.; Guo, J.; Jin, Y.; Yang, F.; Lv, L.; Liu, X. Genetic characterization of a new pandemic Southeast Asia topotype strain of serotype O foot−and−mouth disease virus isolated in China during 2010. Virus Genes 2012, 44, 80–88. [Google Scholar]
- Bruun, T.U.J.; Andersson, A.C.; Draper, S.J.; Howarth, M. Engineering a Rugged Nanoscaffold to Enhance Plug−and−Display Vaccination. ACS Nano 2018, 12, 8855–8866. [Google Scholar] [CrossRef]
- Dong, H.; Lu, Y.; Zhang, Y.; Mu, S.; Wang, N.; Du, P.; Zhi, X.; Wen, X.; Wang, X.; Sun, S.; et al. A Heat−Induced Mutation on VP1 of Foot−and−Mouth Disease Virus Serotype O Enhanced Capsid Stability and Immunogenicity. J. Virol. 2021, 95, e0017721. [Google Scholar] [CrossRef]
- Liu, F.; Wu, X.; Li, L.; Ge, S.; Liu, Z.; Wang, Z. Virus−like particles: Promising platforms with characteristics of DIVA for veterinary vaccine design. Comp. Immunol. Microbiol. Infect. Dis. 2013, 36, 343–352. [Google Scholar]
- Ellis, T.M. New DIVA vaccine for pigs. Vet. J. 2016, 213, 16–17. [Google Scholar] [CrossRef] [PubMed]
- Hardham, J.M.; Krug, P.; Pacheco, J.M.; Thompson, J.; Dominowski, P.; Moulin, V.; Gay, C.G.; Rodriguez, L.L.; Rieder, E. Novel Foot−and−Mouth Disease Vaccine Platform: Formulations for Safe and DIVA−Compatible FMD Vaccines with Improved Potency. Front. Vet. Sci. 2020, 7, 554305. [Google Scholar] [CrossRef] [PubMed]
- Bearson, B.L.; Bearson, S.M.D.; Brunelle, B.W.; Bayles, D.O.; Lee, I.S.; Kich, J.D. Salmonella DIVA vaccine reduces disease, colonization and shedding due to virulent S. Typhimurium infection in swine. J. Med. Microbiol. 2017, 66, 651–661. [Google Scholar] [CrossRef] [PubMed]
- Pavlov, A.V.; Rybakov, S.S.; Ivaniushchenkov, V.N.; Chepurkin, A.V.; Petrov, V.N.; Driagalin, N.N.; Burdov, A.N. Protection from foot−and−mouth disease virus in naturally−susceptible animals by a linear polymer of a synthetic peptide. Bioorg. Khim. 1991, 17, 953–963. [Google Scholar]
- Lin, G.J.; Deng, M.C.; Chen, Z.W.; Liu, T.Y.; Wu, C.W.; Cheng, C.Y.; Chien, M.S.; Huang, C. Yeast expressed classical swine fever E2 subunit vaccine candidate provides complete protection against lethal challenge infection and prevents horizontal virus transmission. Vaccine 2012, 30, 2336–2341. [Google Scholar] [CrossRef]
- Gao, F.; Jiang, Y.; Li, G.; Li, L.; Zhang, Y.; Yu, L.; Zheng, H.; Tong, W.; Zhou, Y.; Liu, C.; et al. Evaluation of immune efficacy of recombinant PRRSV vectored vaccine rPRRSV−E2 in piglets with maternal derived antibodies. Vet. Microbiol. 2020, 248, 108833. [Google Scholar]
- Kardani, K.; Bolhassani, A.; Shahbazi, S. Prime−boost vaccine strategy against viral infections: Mechanisms and benefits. Vaccine 2016, 34, 413–423. [Google Scholar]
- Shen, H.; Ackerman, A.L.; Cody, V.; Giodini, A.; Hinson, E.R.; Cresswell, P.; Edelson, R.L.; Saltzman, W.M.; Hanlon, D.J. Enhanced and prolonged cross−presentation following endosomal escape of exogenous antigens encapsulated in biodegradable nanoparticles. Immunology 2006, 117, 78–88. [Google Scholar] [CrossRef]
- Kim, Y.S.; Son, A.; Kim, J.; Kwon, S.B.; Kim, M.H.; Kim, P.; Kim, J.; Byun, Y.H.; Sung, J.; Lee, J.; et al. Chaperna−Mediated Assembly of Ferritin−Based Middle East Respiratory Syndrome−Coronavirus Nanoparticles. Front. Immunol. 2018, 9, 1093. [Google Scholar] [CrossRef]
- King, N.P.; Lai, Y.T. Practical approaches to designing novel protein assemblies. Curr. Opin. Struct. Biol. 2013, 23, 632–638. [Google Scholar]
- Salgado, E.N.; Radford, R.J.; Tezcan, F.A. Metal−directed protein self−assembly. Acc. Chem. Res. 2010, 43, 661–672. [Google Scholar] [CrossRef] [PubMed]
- Ma, X.; Zou, F.; Yu, F.; Li, R.; Yuan, Y.; Zhang, Y.; Zhang, X.; Deng, J.; Chen, T.; Song, Z.; et al. Nanoparticle Vaccines Based on the Receptor Binding Domain (RBD) and Heptad Repeat (HR) of SARS−CoV−2 Elicit Robust Protective Immune Responses. Immunity 2020, 53, 1315–1330.e9. [Google Scholar] [CrossRef] [PubMed]
- Dhakal, S.; Renukaradhya, G.J. Nanoparticle−based vaccine development and evaluation against viral infections in pigs. Vet. Res. 2019, 50, 90. [Google Scholar] [CrossRef]
- Wang, D.; Zhang, S.; Zou, Y.; Yu, W.; Jiang, Y.; Zhan, Y.; Wang, N.; Dong, Y.; Yang, Y. Structure−Based Design of Porcine Circovirus Type 2 Chimeric VLPs (cVLPs) Displays Foreign Peptides on the Capsid Surface. Front. Cell Infect. Microbiol. 2018, 8, 232. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.H.; Xu, H.L.; Han, G.W.; Tao, L.N.; Lu, Y.; Zheng, S.Y.; Fang, W.H.; He, F. A self−assembling nanoparticle: Implications for the development of thermostable vaccine candidates. Int. J. Biol. Macromol. 2021, 183, 2162–2173. [Google Scholar] [CrossRef]
- Kang, Y.F.; Zhang, X.; Yu, X.H.; Zheng, Q.B.; Liu, Z.; Li, J.P.; Sun, C.; Kong, X.W.; Zhu, Q.Y.; Chen, H.W.; et al. Immunization with a Self−Assembled Nanoparticle Vaccine Elicits Potent Neutralizing Antibody Responses against EBV Infection. Nano Letters 2021, 21, 2476–2486. [Google Scholar] [CrossRef]
- Burman, A.; Clark, S.; Abrescia, N.G.; Fry, E.E.; Stuart, D.I.; Jackson, T. Specificity of the VP1 GH loop of Foot−and−Mouth Disease virus for alphav integrins. J. Virol. 2006, 80, 9798–9810. [Google Scholar] [CrossRef]
- Mahdy, S.E.; Liu, S.; Su, L.; Zhang, X.; Chen, H.; Pei, X.; Wang, C. Expression of the VP1 protein of FMDV integrated chromosomally with mutant Listeria monocytogenes strain induced both humoral and cellular immune responses. Appl. Microbiol. Biotechnol. 2019, 103, 1919–1929. [Google Scholar] [CrossRef]
- Wu, P.; Yin, X.; Liu, Q.; Wu, W.; Chen, C. Recombinant T7 Phage with FMDV AKT−III Strain VP1 Protein is a Potential FMDV Vaccine. Biotechnol. Lett. 2021, 43, 35–41. [Google Scholar] [CrossRef]
- Akache, B.; Stark, F.C.; Agbayani, G.; Renner, T.M.; McCluskie, M.J. Adjuvants: Engineering Protective Immune Responses in Human and Veterinary Vaccines. Methods Mol. Biol. 2022, 2412, 179–231. [Google Scholar]
- Liu, Z.H.; Xu, H.L.; Han, G.W.; Tao, L.N.; Lu, Y.; Zheng, S.Y.; Fang, W.H.; He, F. Self−Assembling Nanovaccine Enhances Protective Efficacy against CSFV in Pigs. Front. Immunol. 2021, 12, 689187. [Google Scholar] [CrossRef] [PubMed]
- Li, B.; Du, L.; Yu, Z.; Sun, B.; Xu, X.; Fan, B.; Guo, R.; Yuan, W.; He, K. Poly (d,l−lactide−co−glycolide) nanoparticle−entrapped vaccine induces a protective immune response against porcine epidemic diarrhea virus infection in piglets. Vaccine 2017, 35, 7010–7017. [Google Scholar] [CrossRef] [PubMed]
- Lycke, N. Recent progress in mucosal vaccine development: Potential and limitations. Nat. Rev. Immunol. 2012, 12, 592–605. [Google Scholar] [CrossRef]
- Yang, F.; Wang, F.; Guo, Y.; Zhou, Q.; Wang, Y.; Yin, Y.; Sun, S. Enhanced capacity of antigen presentation of HBc−VLP−pulsed RAW264.7 cells revealed by proteomics analysis. J. Proteome Res. 2008, 7, 4898–4903. [Google Scholar] [CrossRef] [PubMed]
- Kang, Y.F.; Sun, C.; Zhuang, Z.; Yuan, R.Y.; Zheng, Q.; Li, J.P.; Zhou, P.P.; Chen, X.C.; Liu, Z.; Zhang, X.; et al. Rapid Development of SARS−CoV−2 Spike Protein Receptor−Binding Domain Self−Assembled Nanoparticle Vaccine Candidates. ACS Nano 2021, 15, 2738–2752. [Google Scholar] [CrossRef] [PubMed]
- Kanekiyo, M.; Wei, C.J.; Yassine, H.M.; McTamney, P.M.; Boyington, J.C.; Whittle, J.R.; Rao, S.S.; Kong, W.P.; Wang, L.; Nabel, G.J. Self−assembling influenza nanoparticle vaccines elicit broadly neutralizing H1N1 antibodies. Nature 2013, 499, 102–106. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Sequence |
|---|---|
| β−actin F | GCTGTCCCTGTATGCCTCT |
| β−actin R | TTGATGTCACGCACGATTT |
| MHC II F: | CTGTCTGGATGCTTCCTGAGTTT |
| MHC II R: | TCAGCTATGTTTTGCAGTCCACC |
| CD80 F | CCCCAGAAGACCCTCCTGATAG |
| CD80 R | CGAAGGTAAGGCTGTTGTTTG |
| CD86 F | GCCGTGCCCATTTACAAAGGCTCAA |
| CD86 R | TGTTACATTCTGAGCCAGTTTTATT |
| TNF−α F | GGCAGGTCTACTTTGGAGTCATTGC |
| TNF−α R | ACATTCGAGGCTCCAGTGAATTCGG |
| IL−6 F | TCCAGTTGCCTTCTTGGGAC |
| IL−6 R | GTGTAATTAAGCCTCCGACTTG |
| IFN−γ F | TCAGCTGATCCTTTGGACCC |
| IFN−γ R | CTCAGAGCTAGGCCGCAGG |
| Gene | Sequence |
|---|---|
| GAPDH F | TGCCCCTATGTTCGTGATGG |
| GAPDH R | TGTGGTCATGAGTCCCTCCA |
| TNF−α F | CGCTCACACTCAGATCAGCTT |
| TNF−α R | GACGGTATGGGTGAGAAGCA |
| IFN−γ F | AATGACGAGCATGTCCAGCG |
| IFN−γ R | CTCTCCGGCTCTGAAACAGC |
| IL−12p 70 F | GTCACAAAGGAGGCGAGGTT |
| IL−12p 70 R | AGCAGGTGAAACGTCCAGAA |
| Guinea Pigs | PBS | Inactivated FMDV | VP1 | VP1−i301 | VP1−i301−nFLiC |
|---|---|---|---|---|---|
| 1 | None | Partial | None | Full | Full |
| 2 | None | Full | Full | Full | Full |
| 3 | None | Full | None | None | Full |
| 4 | None | Full | Partial | Full | Partial |
| 5 | None | Full | Partial | Partial | Full |
| 6 | None | Full | Full | Full | Full |
| Rate of protection (%) | 0 | 83.3 | 33.3 | 66.7 | 66.7 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pei, C.; Dong, H.; Teng, Z.; Wei, S.; Zhang, Y.; Yin, S.; Tang, J.; Sun, S.; Guo, H. Self−Assembling Nanovaccine Fused with Flagellin Enhances Protective Effect against Foot−and−Mouth Disease Virus. Vaccines 2023, 11, 1675. https://doi.org/10.3390/vaccines11111675
Pei C, Dong H, Teng Z, Wei S, Zhang Y, Yin S, Tang J, Sun S, Guo H. Self−Assembling Nanovaccine Fused with Flagellin Enhances Protective Effect against Foot−and−Mouth Disease Virus. Vaccines. 2023; 11(11):1675. https://doi.org/10.3390/vaccines11111675
Chicago/Turabian StylePei, Chenchen, Hu Dong, Zhidong Teng, Sumin Wei, Yun Zhang, Shuanghui Yin, Jianli Tang, Shiqi Sun, and Huichen Guo. 2023. "Self−Assembling Nanovaccine Fused with Flagellin Enhances Protective Effect against Foot−and−Mouth Disease Virus" Vaccines 11, no. 11: 1675. https://doi.org/10.3390/vaccines11111675
APA StylePei, C., Dong, H., Teng, Z., Wei, S., Zhang, Y., Yin, S., Tang, J., Sun, S., & Guo, H. (2023). Self−Assembling Nanovaccine Fused with Flagellin Enhances Protective Effect against Foot−and−Mouth Disease Virus. Vaccines, 11(11), 1675. https://doi.org/10.3390/vaccines11111675