
Pathological Evaluation of Porcine Circovirus 2d (PCV2d) Strain and Comparative Evaluation of PCV2d and PCV2b Inactivated Vaccines against PCV2d Infection in a Specific Pathogen-Free (SPF) Yucatan Miniature Pig Model

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
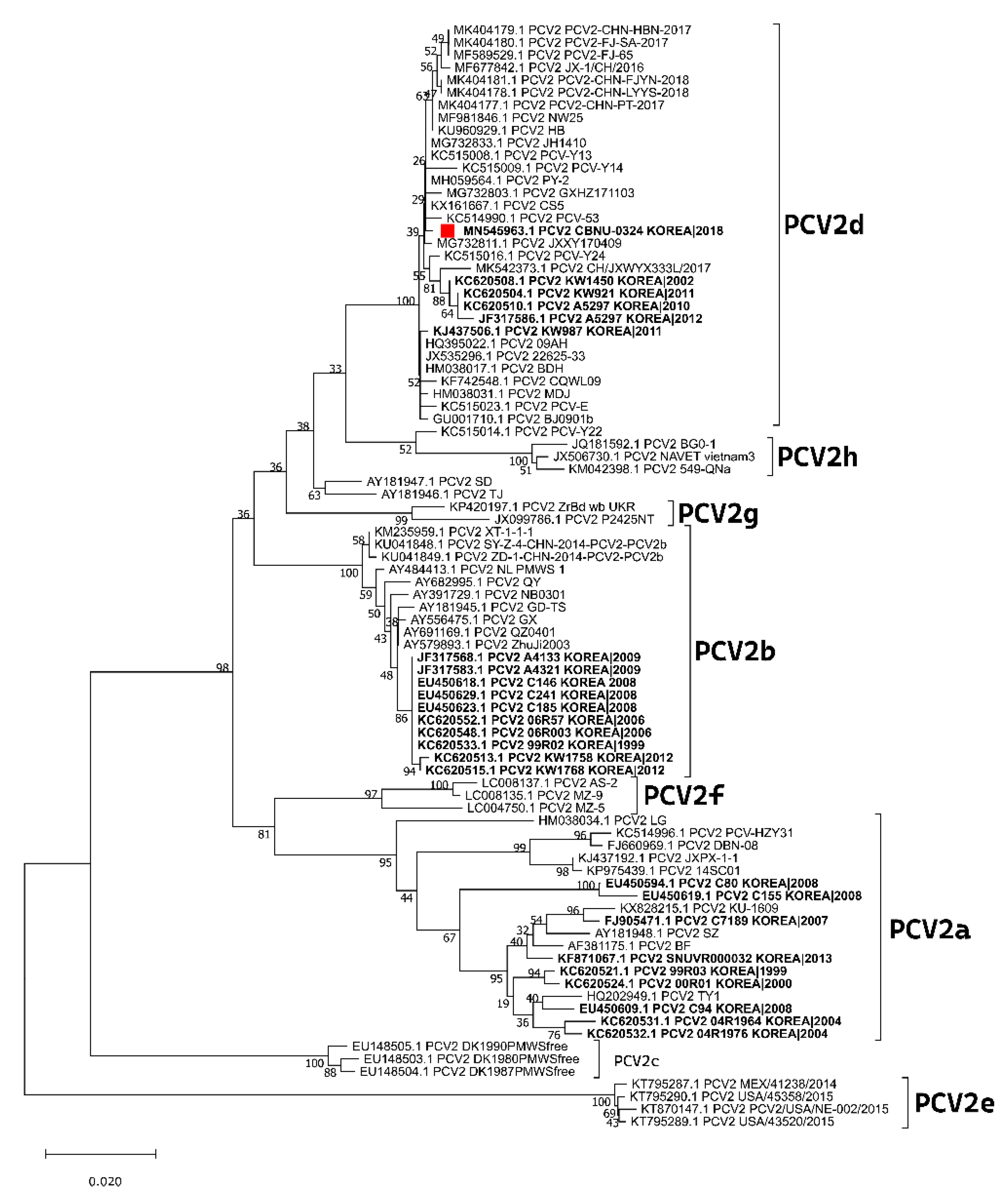
2.1. Cells and Virus Isolation
2.2. Vaccine Preparation
2.3. Animals
2.4. Experimental Design
2.5. Clinical Evaluation
2.6. Serology and Measurement of Neutralizing Antibodies against PCV2
2.7. Quantitative Real-Time PCR
2.8. Necropsy and Macroscopic Lesions
2.9. Microscopic Lesions and Immunohistochemistry
2.10. Statistical Analysis
3. Results
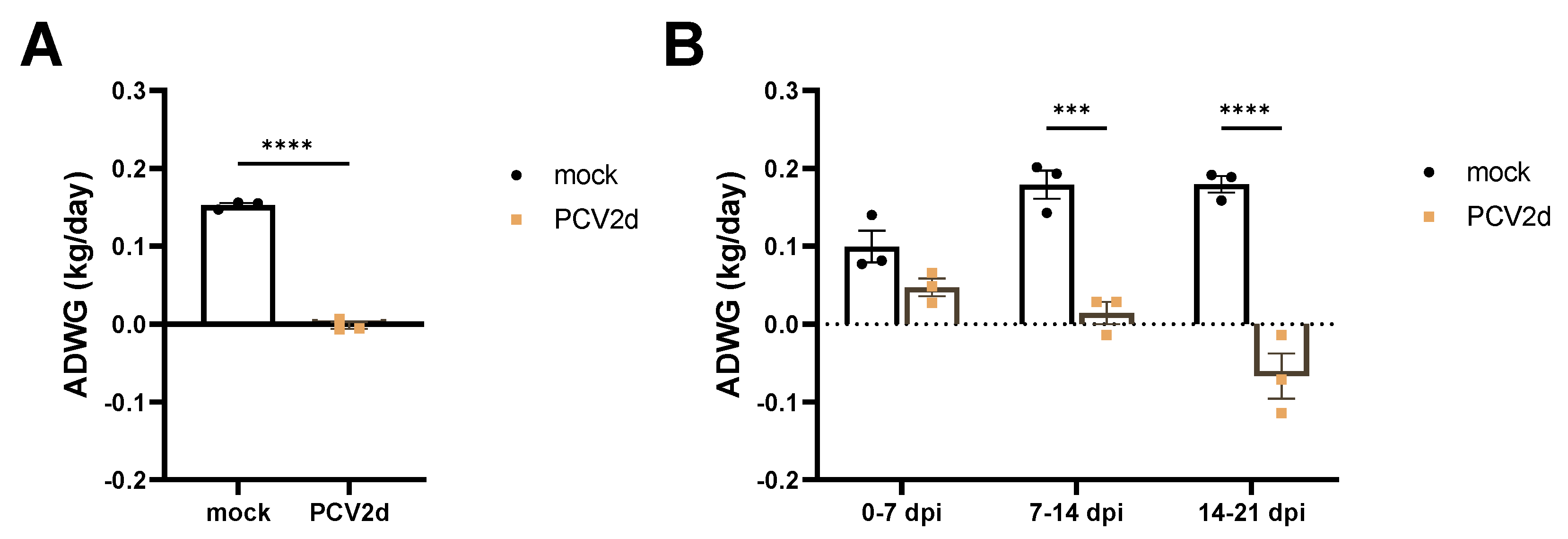
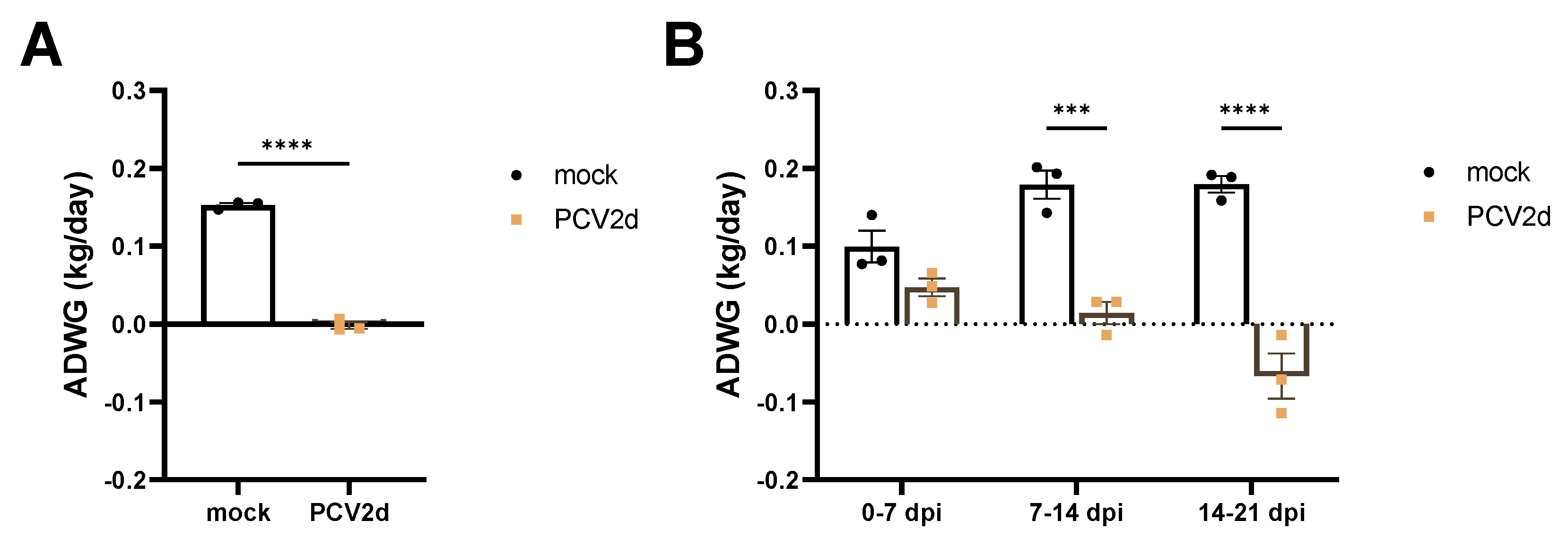
3.1. Experiment 1: Clinical Evaluation and Growth Performance
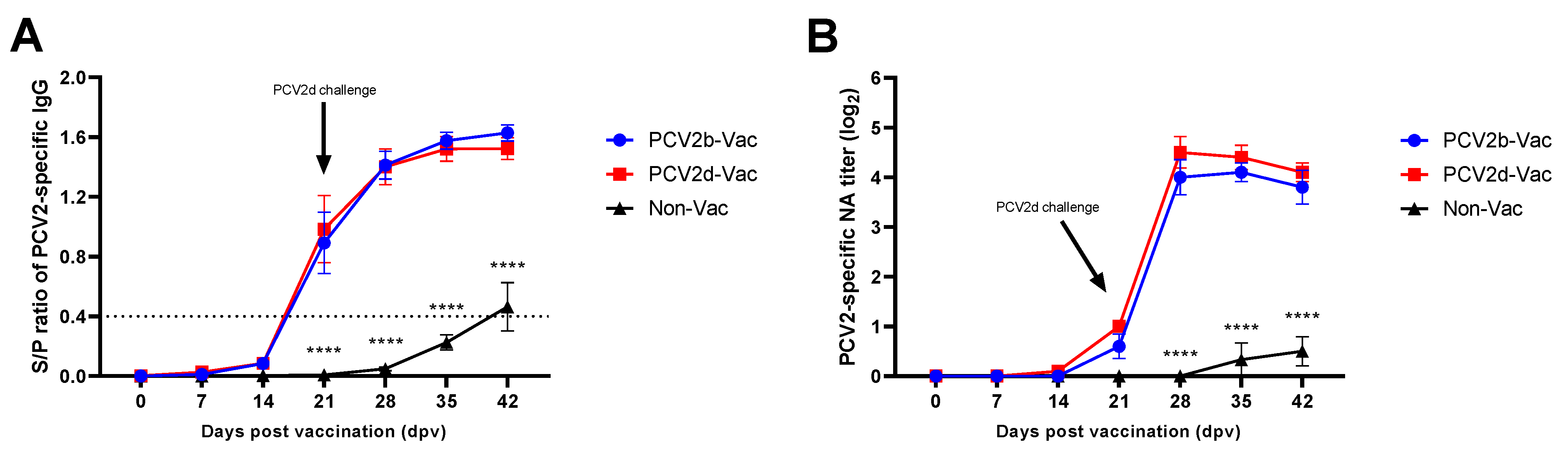
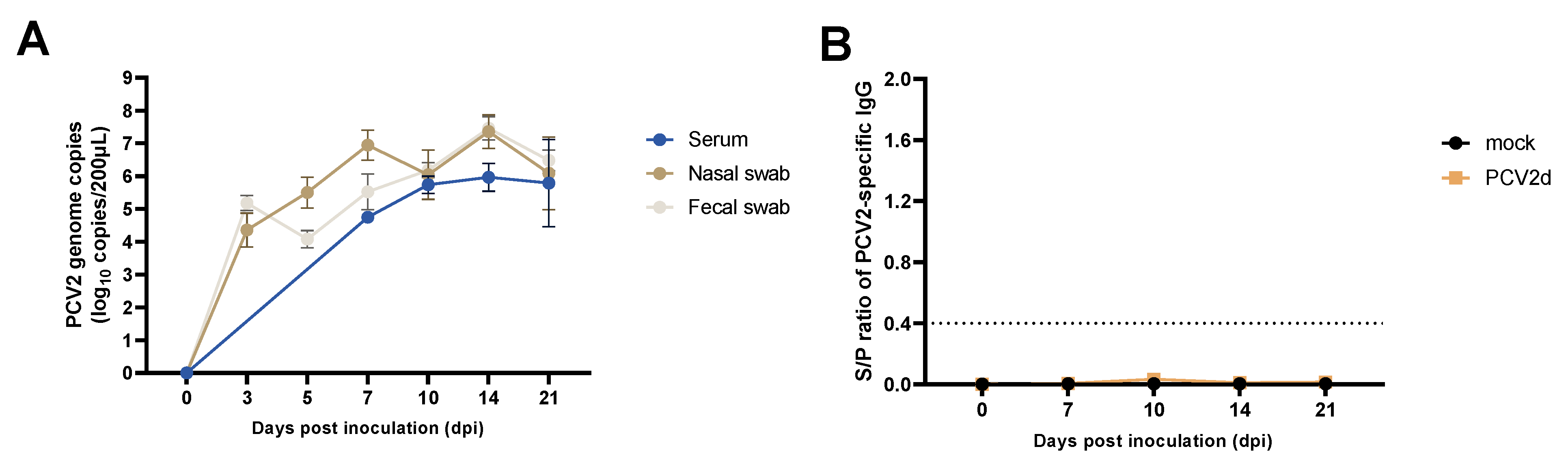
3.2. Experiment 1: PCV2d Viremia, Viral Shedding, and PCV2-Specific IgG
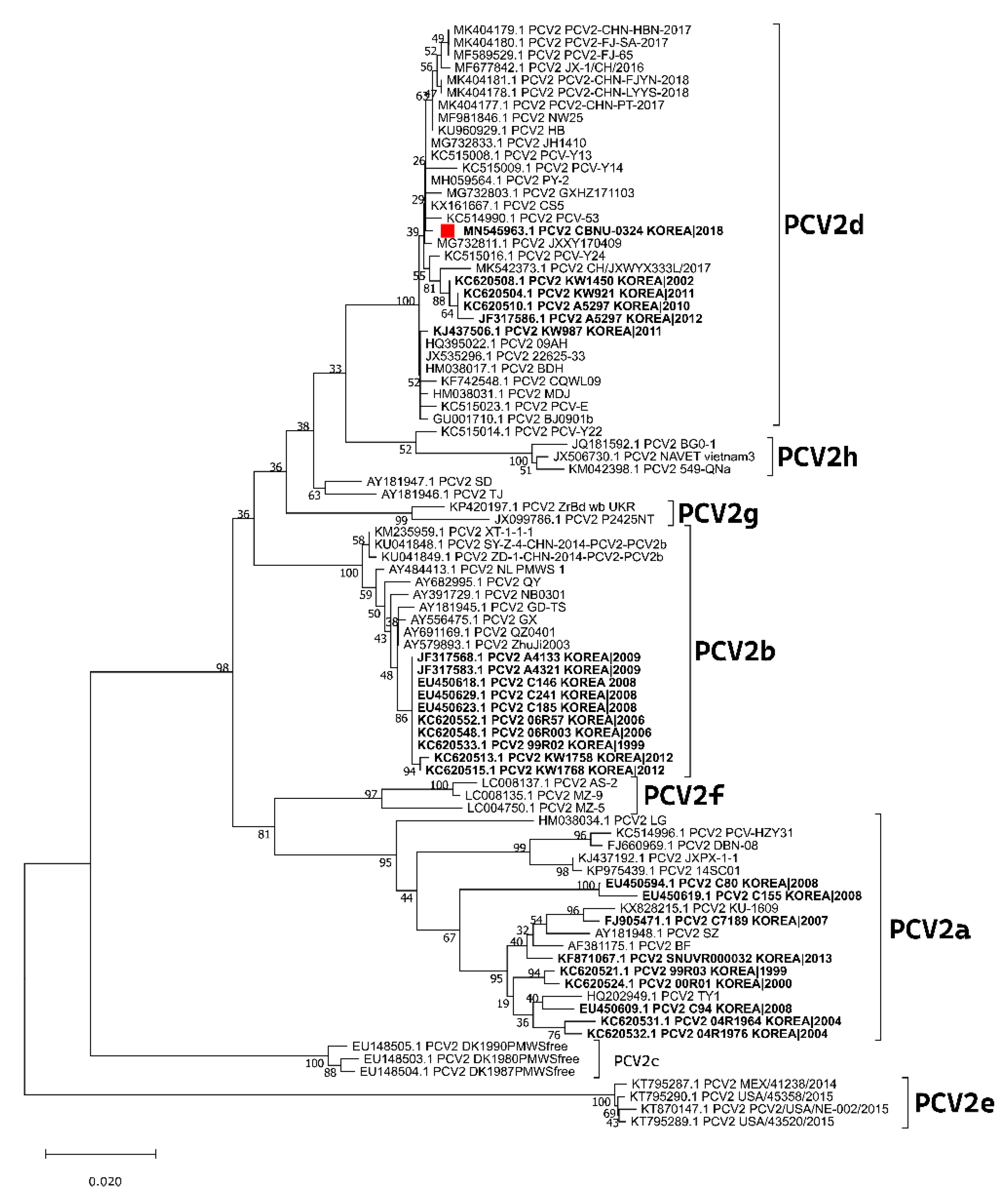
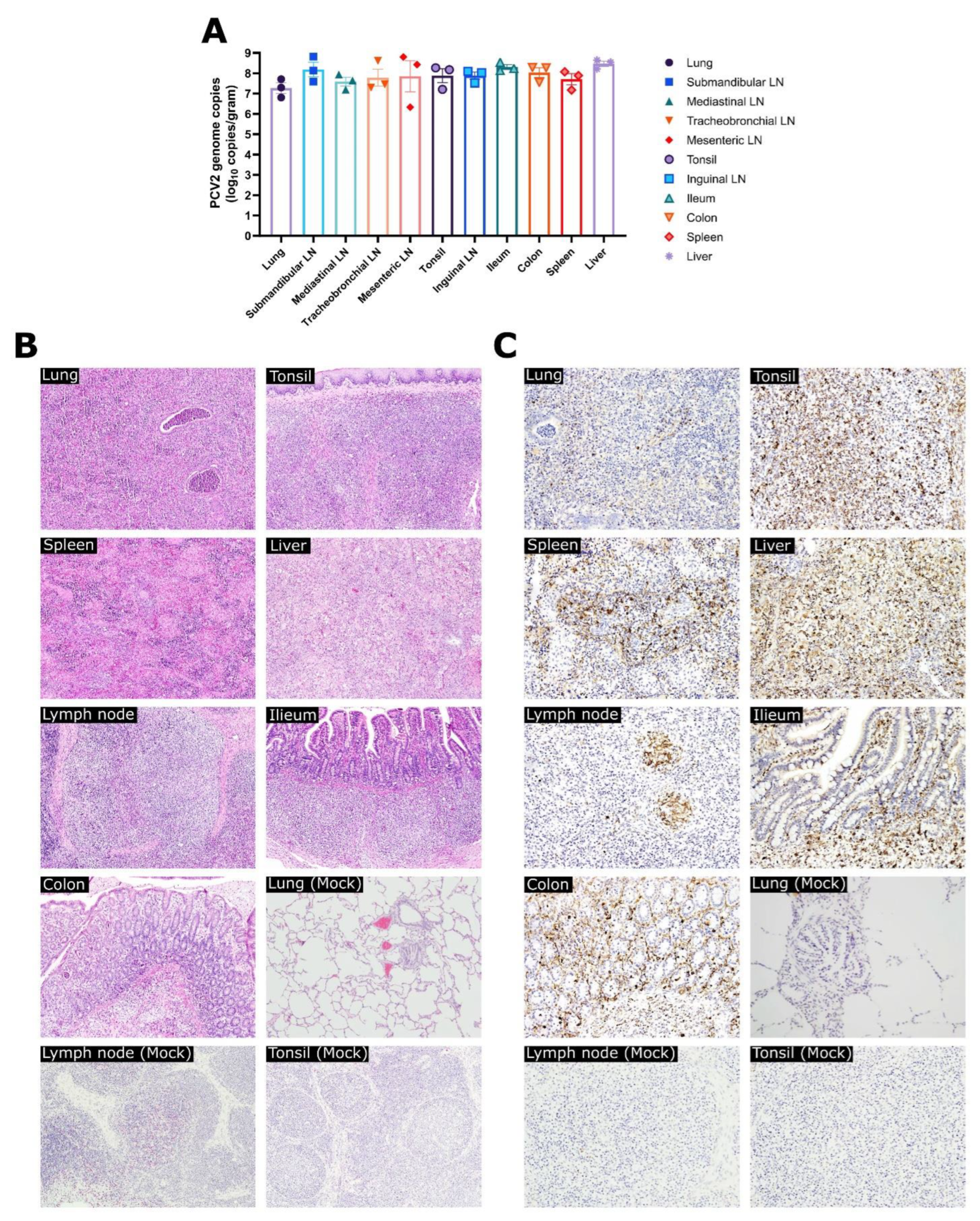
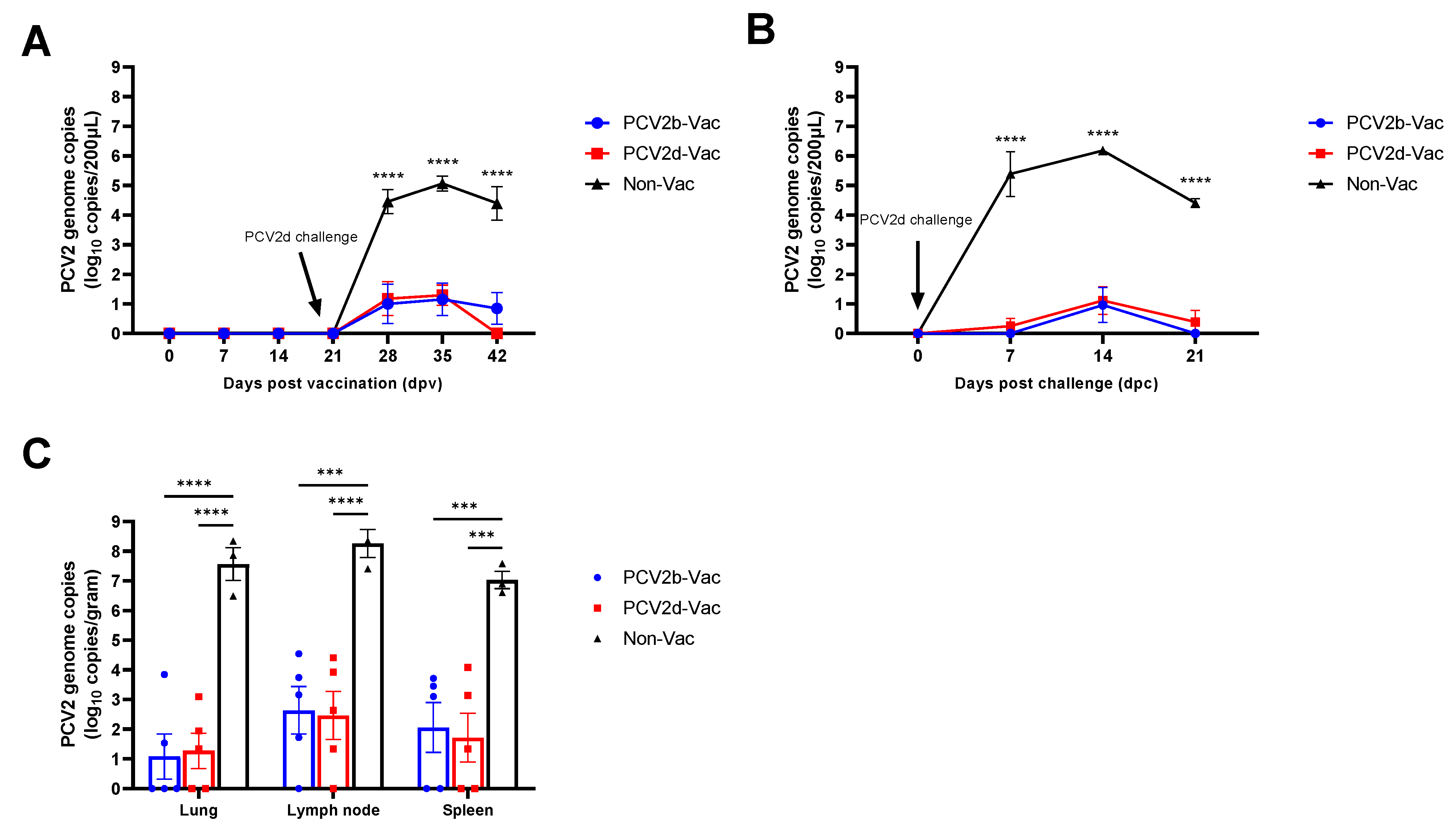
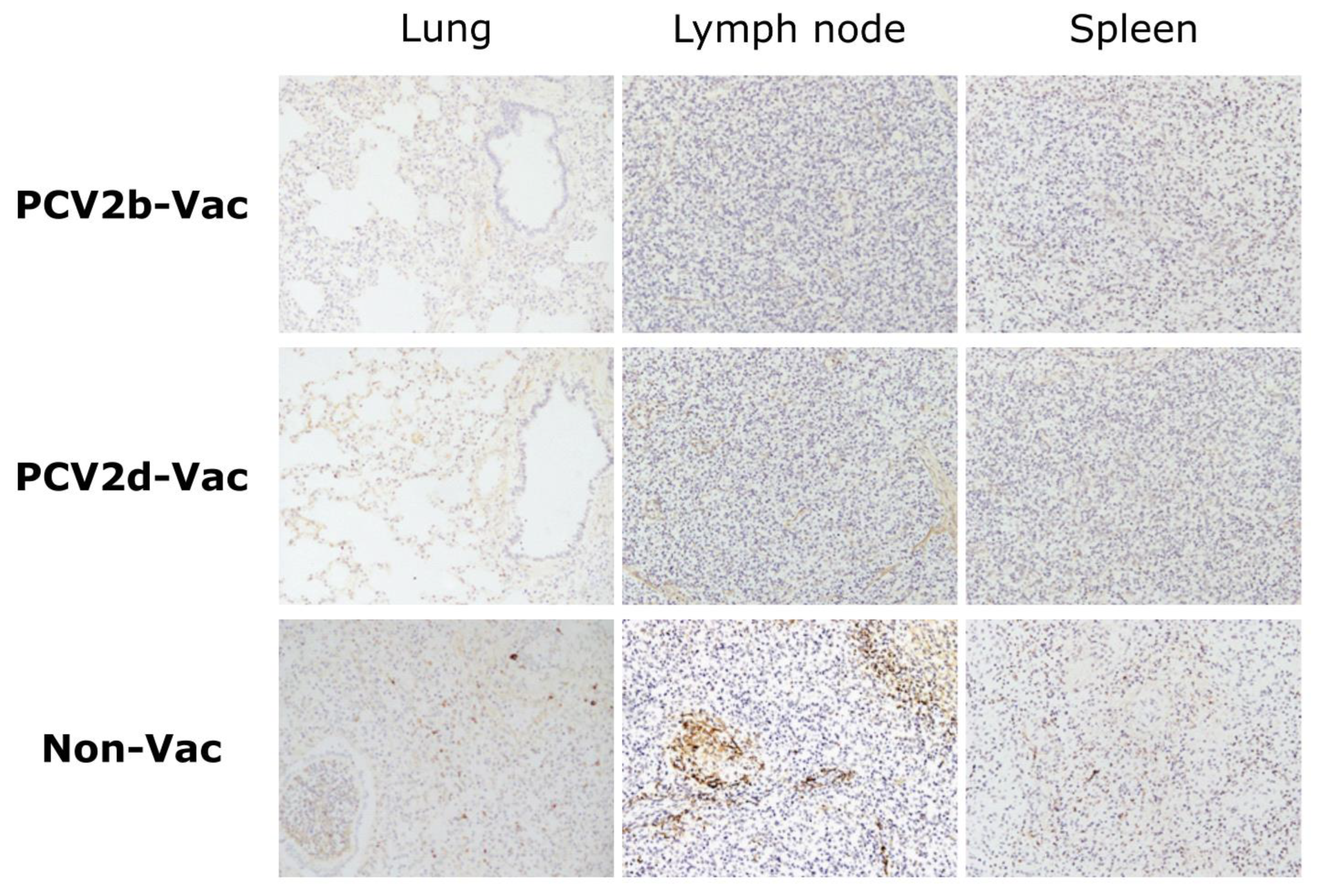
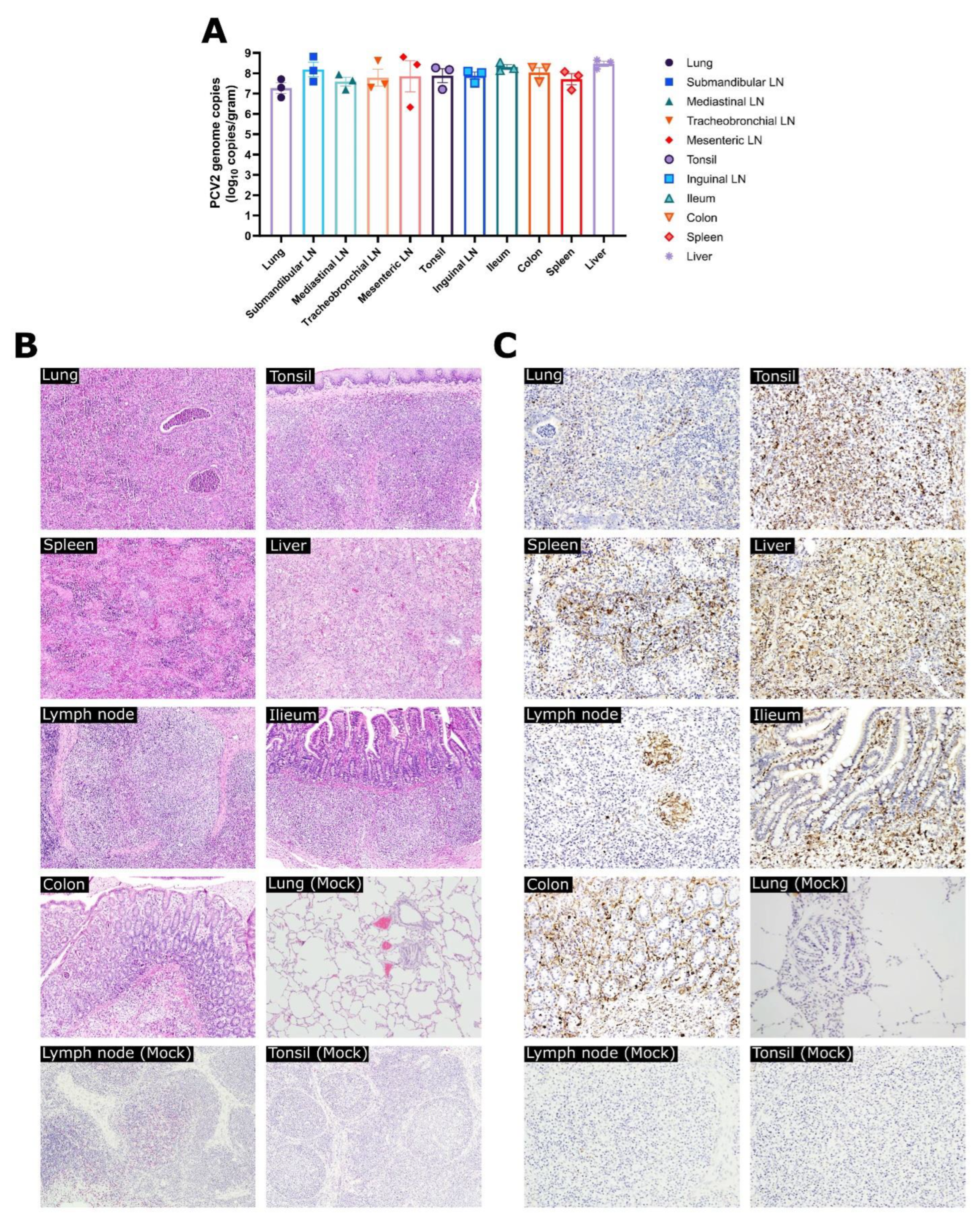
3.3. Experiment 1: Viral Loads, Pathological Evaluation, and Detection of PCV2 Antigen in Tissues
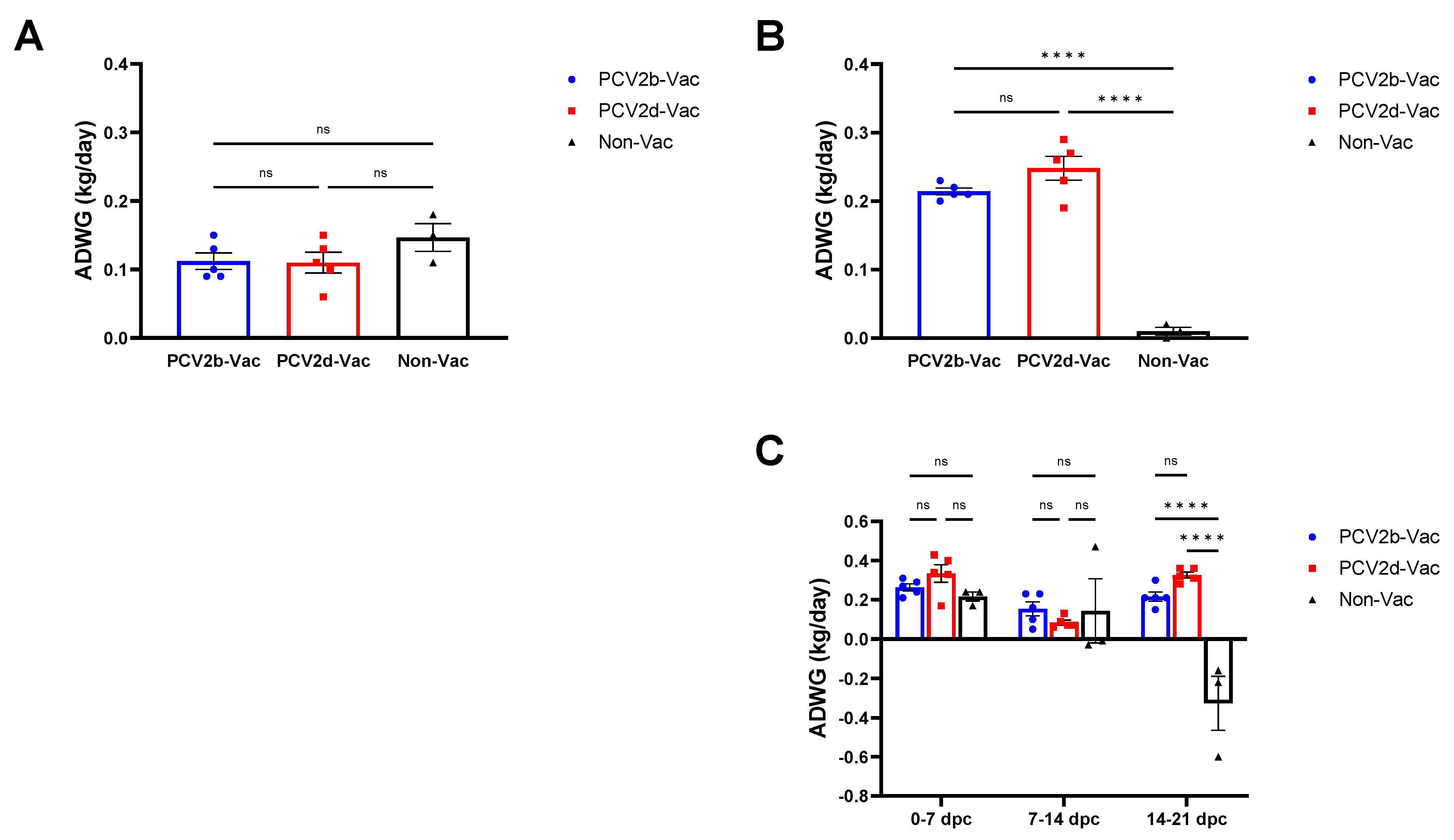
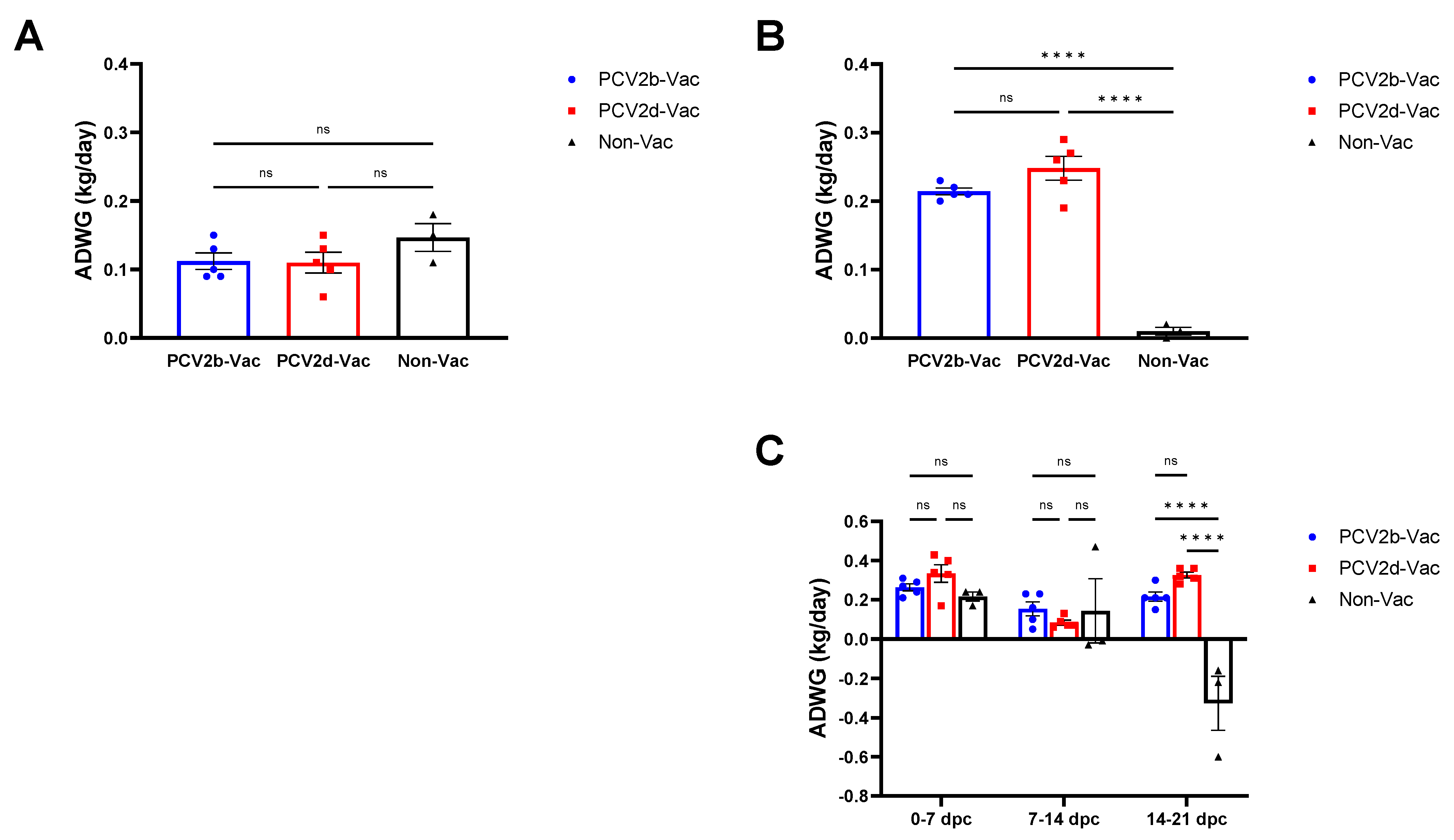
3.4. Experiment 2: Clinical Evaluation and Growth Performance
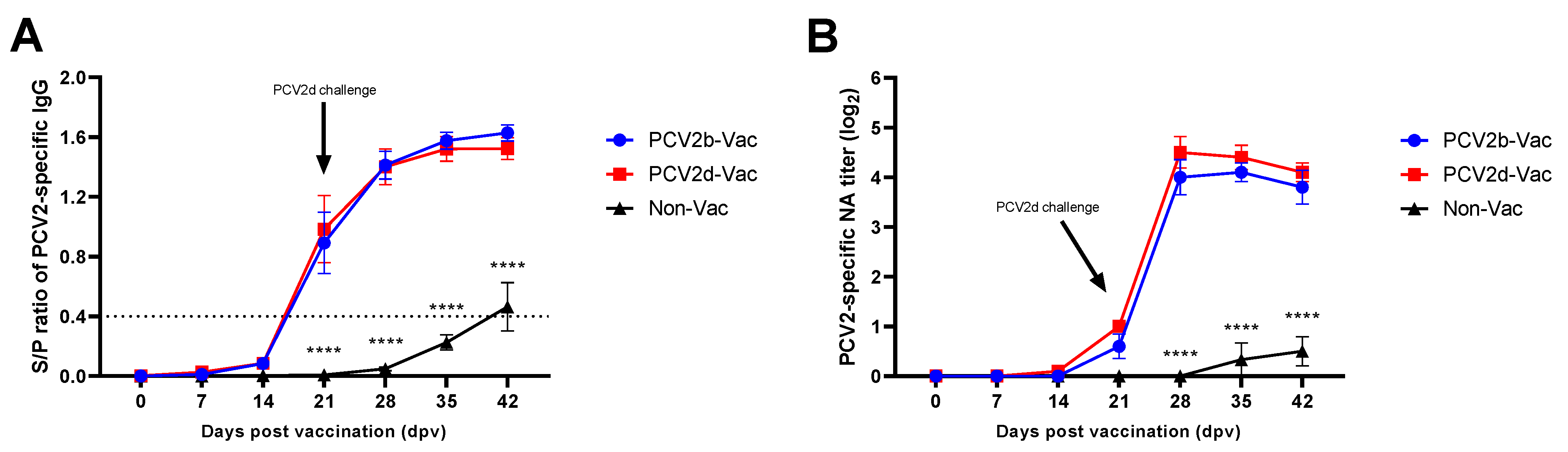
3.5. Experiment 2: Serological Evaluation
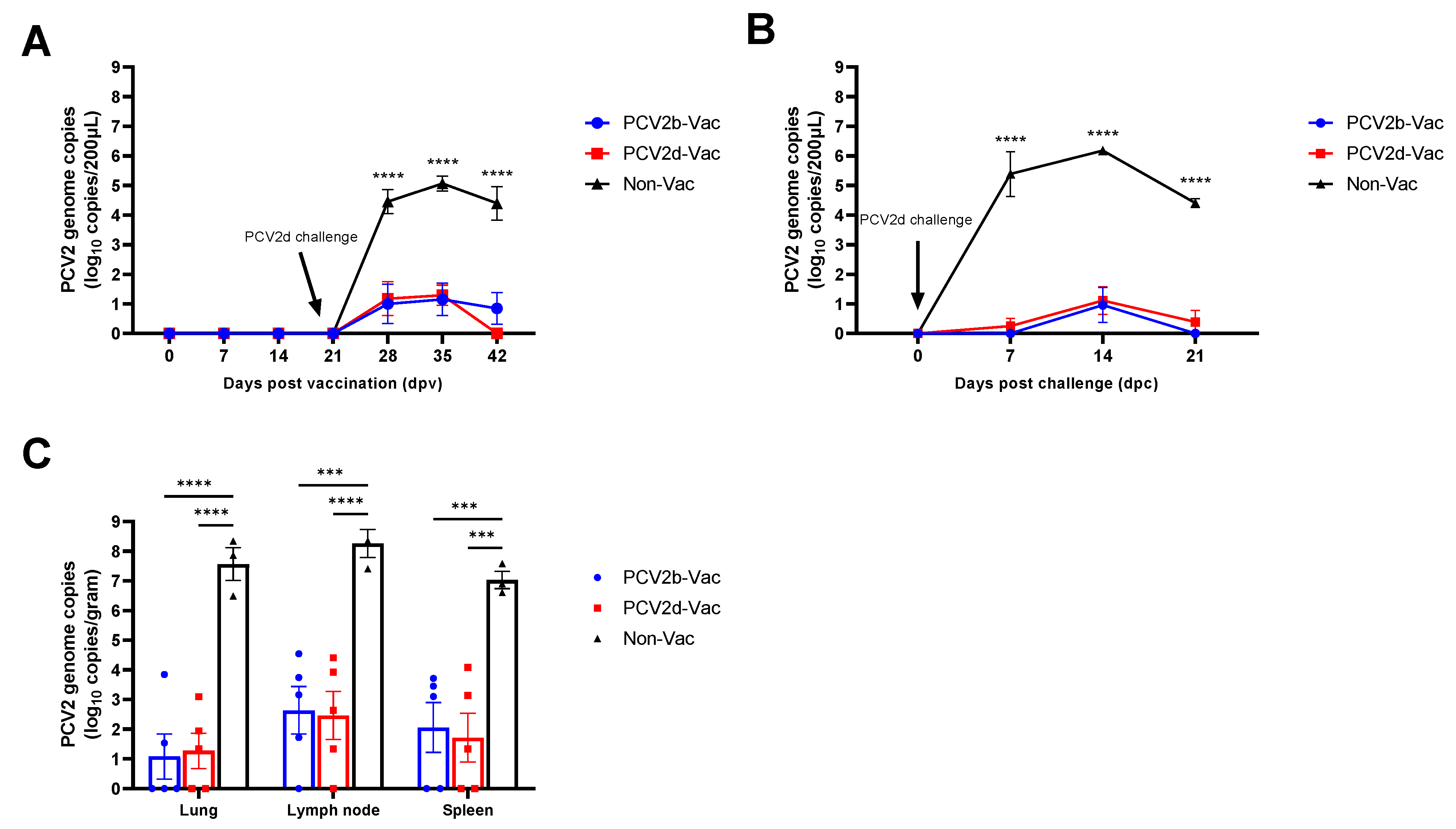
3.6. Experiment 2: Viremia, Viral Shedding, and Viral Load in Tissues
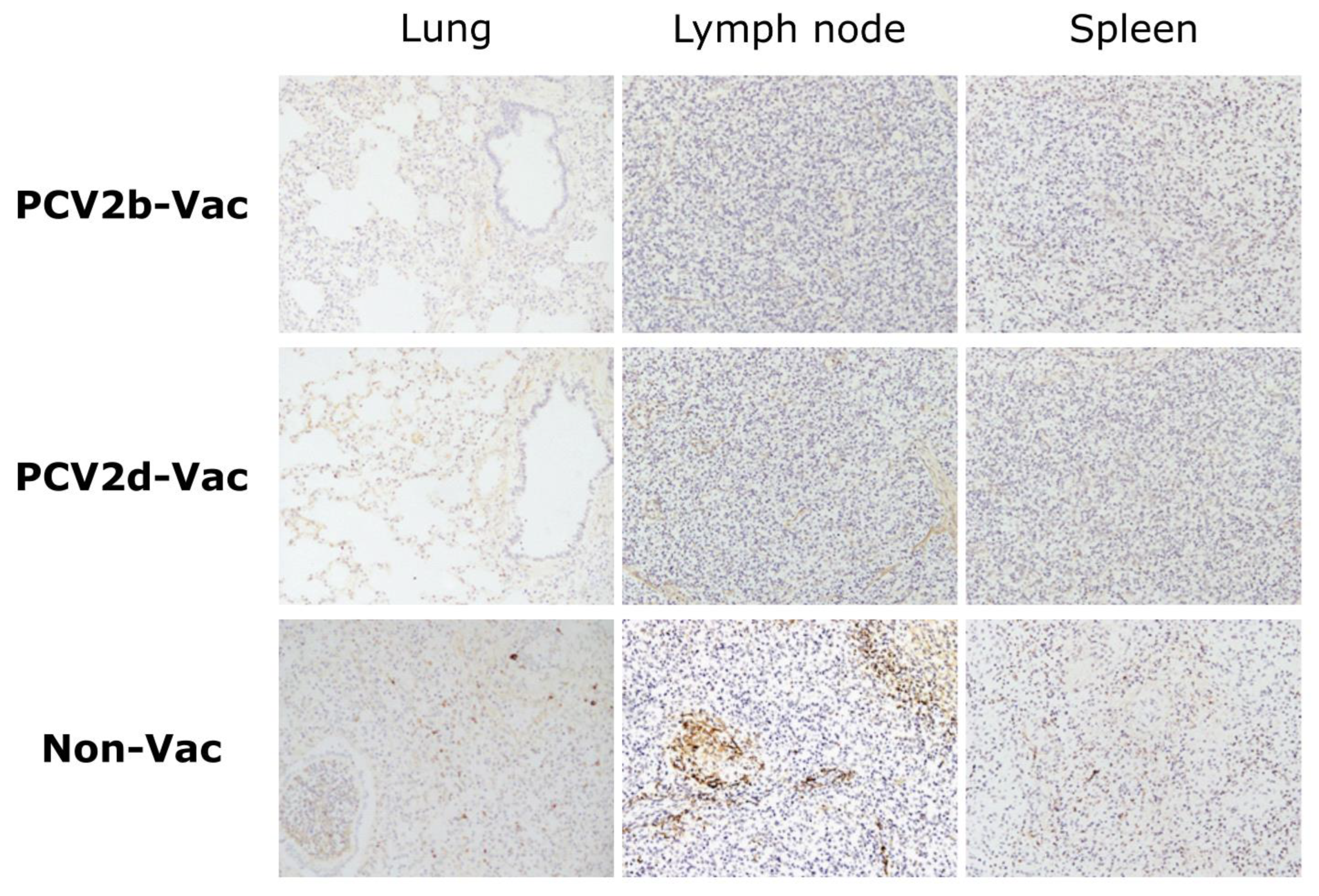
3.7. Experiment 2: Histopathological Lesions and PCV2 Antigens in Lymphoid Tissues
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Chae, C. A review of porcine circovirus 2-associated syndromes and diseases. Vet. J. 2005, 169, 326–336. [Google Scholar] [CrossRef] [PubMed]
- Jensen, T.K.; Vigre, H.; Svensmark, B.; Bille-Hansen, V. Distinction between porcine circovirus type 2 enteritis and porcine proliferative enteropathy caused by Lawsonia intracellularis. J. Comp. Pathol. 2006, 135, 176–182. [Google Scholar] [CrossRef] [PubMed]
- Seo, H.W.; Park, C.; Kang, I.; Choi, K.; Jeong, J.; Park, S.-J.; Chae, C. Genetic and antigenic characterization of a newly emerging porcine circovirus type 2b mutant first isolated in cases of vaccine failure in Korea. Arch. Virol. 2014, 159, 3107–3111. [Google Scholar] [CrossRef]
- Harding, J.C.S.; Clark, E.G. Recognizing and diagnosing postweaning multisystemic wasting syndrome (PMWS). J. Swine Health Prod. 1997, 5, 201–203. [Google Scholar]
- Karuppannan, A.K.; Opriessnig, T. Porcine Circovirus Type 2 (PCV2) Vaccines in the Context of Current Molecular Epidemiology. Viruses 2017, 9, 99. [Google Scholar] [CrossRef] [PubMed]
- Franzo, G.; Segales, J. Porcine circovirus 2 (PCV-2) genotype update and proposal of a new genotyping methodology. PLoS ONE 2018, 13, e0208585. [Google Scholar] [CrossRef]
- Segalés, J. Porcine circovirus type 2 (PCV2) infections: Clinical signs, pathology and laboratory diagnosis. Virus Res. 2012, 164, 10–19. [Google Scholar] [CrossRef]
- Dupont, K.; Nielsen, E.; Baekbo, P.; Larsen, L. Genomic analysis of PCV2 isolates from Danish archives and a current PMWS case—Control study supports a shift in genotypes with time. Vet. Microbiol. 2008, 128, 56–64. [Google Scholar] [CrossRef]
- Guo, L.J.; Lu, Y.H.; Wei, Y.W.; Huang, L.P.; Liu, C.M. Porcine circovirus type 2 (PCV2): Genetic variation and newly emerging genotypes in China. Virol. J. 2010, 7, 273. [Google Scholar] [CrossRef]
- Xiao, C.-T.; Halbur, P.G.; Opriessnig, T. Global molecular genetic analysis of porcine circovirus type 2 (PCV2) sequences confirms the presence of four main PCV2 genotypes and reveals a rapid increase of PCV2d. J. Gen. Virol. 2015, 96, 1830–1841. [Google Scholar] [CrossRef]
- Opriessnig, T.; Gerber, P.F.; Xiao, C.T.; Mogler, M.; Halbur, P.G. A commercial vaccine based on PCV2a and an experimental vaccine based on a variant mPCV2b are both effective in protecting pigs against challenge with a 2013 US variant mPCV2b strain. Vaccine 2014, 32, 230–237. [Google Scholar] [CrossRef] [PubMed]
- Davies, B.; Wang, X.; Dvorak, C.M.; Marthaler, D.; Murtaugh, M.P. Diagnostic phylogenetics reveals a new porcine circovirus 2 cluster. Virus Res. 2016, 217, 32–37. [Google Scholar] [CrossRef]
- Harmon, K.M.; Gauger, P.C.; Zhang, J.; Pineyro, P.E.; Dunn, D.D.; Chriswell, A.J. Whole-Genome Sequences of Novel Porcine Circovirus Type 2 Viruses Detected in Swine from Mexico and the United States. Genome Announc. 2015, 3, e01315-15. [Google Scholar] [CrossRef] [PubMed]
- Bao, F.; Mi, S.; Luo, Q.; Guo, H.; Tu, C.; Zhu, G.; Gong, W. Retrospective study of porcine circovirus type 2 infection reveals a novel genotype PCV 2f. Transbound. Emerg. Dis. 2018, 65, 432–440. [Google Scholar] [CrossRef]
- Cheung, A.K.; Lager, K.M.; Kohutyuk, O.I.; Vincent, A.L.; Henry, S.C.; Baker, R.B.; Rowland, R.R.; Dunham, A.G. Detection of two porcine circovirus type 2 genotypic groups in United States swine herds. Arch. Virol. 2007, 152, 1035–1044. [Google Scholar] [CrossRef] [PubMed]
- Kwon, T.; Lee, D.U.; Yoo, S.J.; Je, S.H.; Shin, J.Y.; Lyoo, Y.S. Genotypic diversity of porcine circovirus type 2 (PCV2) and genotype shift to PCV2d in Korean pig population. Virus Res. 2017, 228, 24–29. [Google Scholar] [CrossRef]
- Kim, S.C.; Nazki, S.; Kwon, S.; Juhng, J.H.; Mun, K.H.; Jeon, D.Y.; Jeong, C.G.; Khatun, A.; Kang, S.J.; Kim, W.I. The prevalence and genetic characteristics of porcine circovirus type 2 and 3 in Korea. BMC Vet. Res. 2018, 14, 294. [Google Scholar] [CrossRef]
- Ellis, J. Porcine Circovirus A Historical Perspective. Vet. Pathol. 2014, 51, 315–327. [Google Scholar] [CrossRef]
- Thangthamniyom, N.; Sangthong, P.; Poolperm, P.; Thanantong, N.; Boonsoongnern, A.; Hansoongnern, P.; Semkum, P.; Petcharat, N.; Lekcharoensuk, P. Genetic diversity of porcine circovirus type 2 (PCV2) in Thailand during 2009-2015. Vet. Microbiol. 2017, 208, 239–246. [Google Scholar] [CrossRef]
- Guo, L.J.; Fu, Y.J.; Huang, L.P.; Wang, Y.P.; Wei, Y.W.; Wu, H.L.; Liu, C.M. A commercial PCV2a-based vaccine is effective in protection from experimental challenge of PCV2 mutant with two amino acids elongation in capsid protein. Vaccine 2015, 33, 3752–3757. [Google Scholar] [CrossRef]
- Paladino, E.S.; Gabardo, M.D.P.; Lunardi, P.N.; Mores, N.; Guedes, R.M.C. Anatomopathological pneumonic aspects associated with highly pathogenic Pasteurella multocida in finishing pigs. Pesqui. Veterinária Bras. 2017, 37, 1091–1100. [Google Scholar] [CrossRef] [Green Version]
- Opriessnig, T.; Gerber, P.F.; Xiao, C.T.; Halbur, P.G.; Matzinger, S.R.; Meng, X.J. Commercial PCV2a-based vaccines are effective in protecting naturally PCV2b-infected finisher pigs against experimental challenge with a 2012 mutant PCV2. Vaccine 2014, 32, 4342–4348. [Google Scholar] [CrossRef] [PubMed]
- Opriessnig, T.; Xiao, C.T.; Halbur, P.G.; Gerber, P.F.; Matzinger, S.R.; Meng, X.J. A commercial porcine circovirus (PCV) type 2a-based vaccine reduces PCV2d viremia and shedding and prevents PCV2d transmission to naive pigs under experimental conditions. Vaccine 2017, 35, 248–254. [Google Scholar] [CrossRef]
- Park, K.H.; Oh, T.; Yang, S.; Cho, H.; Kang, I.; Chae, C. Evaluation of a porcine circovirus type 2a (PCV2a) vaccine efficacy against experimental PCV2a, PCV2b, and PCV2d challenge. Vet. Microbiol. 2019, 231, 87–92. [Google Scholar] [CrossRef] [PubMed]
- Ellis, J.; Hassard, L.; Clark, E.; Harding, J.; Allan, G.; Willson, P.; Strokappe, J.; Martin, K.; McNeilly, F.; Meehan, B. Isolation of circovirus from lesions of pigs with postweaning multisystemic wasting syndrome. Can. Vet. J. 1998, 39, 44. [Google Scholar]
- Gauger, P.C. Characterization of Porcine Circovirus Type 2a and 2b Infection and Lesions in Gnotobiotic Pigs; Iowa State University: Ames, IA, USA, 2008. [Google Scholar]
- Lager, K.; Gauger, P.; Vincent, A.; Opriessnig, T.; Kehrli, M.; Cheung, A. Mortality in pigs given porcine circovirus type 2 subgroup 1 and 2 viruses derived from DNA clones. Vet. Rec. 2007, 161, 428. [Google Scholar] [CrossRef]
- Meurens, F.; Summerfield, A.; Nauwynck, H.; Saif, L.; Gerdts, V. The pig: A model for human infectious diseases. Trends Microbiol. 2012, 20, 50–57. [Google Scholar] [CrossRef]
- Azevedo, M.; Yuan, L.; Pouly, S.; Gonzales, A.; Jeong, K.; Nguyen, T.; Saif, L. Cytokine responses in gnotobiotic pigs after infection with virulent or attenuated human rotavirus. J. Virol. 2006, 80, 372–382. [Google Scholar] [CrossRef]
- Svendsen, O. The minipig in toxicology. Exp. Toxicol. Pathol. 2006, 57, 335–339. [Google Scholar] [CrossRef]
- Kim, H.; Kim, J.-K.; Song, H.; Choi, J.; Shim, B.; Kang, B.; Moon, H.; Yeom, M.; Kim, S.-H.; Song, D. Preliminary study about sublingual administration of bacteria-expressed pandemic H1N1 influenza vaccine in miniature pigs. J. Microbiol. 2014, 52, 794–800. [Google Scholar] [CrossRef]
- Seo, S.H.; Hoffmann, E.; Webster, R.G. Lethal H5N1 influenza viruses escape host anti-viral cytokine responses. Nat. Med. 2002, 8, 950. [Google Scholar] [CrossRef] [PubMed]
- Seo, D.J.; Jung, D.; Jung, S.; Ha, S.K.; Ha, S.D.; Choi, I.S.; Myoung, J.; Choi, C. Experimental miniature piglet model for the infection of human norovirus GII. J. Med. Virol. 2018, 90, 655–662. [Google Scholar] [CrossRef] [PubMed]
- Jung, S.; Seo, D.J.; Yeo, D.; Wang, Z.; Min, A.; Zhao, Z.; Song, M.; Choi, I.-S.; Myoung, J.; Choi, C. Experimental infection of hepatitis E virus induces pancreatic necroptosis in miniature pigs. Sci. Rep. 2020, 10, 12022. [Google Scholar] [CrossRef] [PubMed]
- Allan, G.M.; McNeilly, F.; Kennedy, S.; Daft, B.; Clarke, E.G.; Ellis, J.A.; Haines, D.M.; Meehan, B.M.; Adair, B.M. Isolation of porcine circovirus-like viruses from pigs with a wasting disease in the USA and Europe. J. Vet. Diagn. Investig. 1998, 10, 3–10. [Google Scholar] [CrossRef] [PubMed]
- Pogranichnyy, R.M.; Yoon, K.J.; Harms, P.A.; Swenson, S.L.; Zimmerman, J.J.; Sorden, S.D. Characterization of immune response of young pigs to porcine circovirus type 2 infection. Viral. Immunol. 2000, 13, 143–153. [Google Scholar] [CrossRef] [PubMed]
- Polo, J.; Opriessnig, T.; O’Neill, K.C.; Rodriguez, C.; Russell, L.E.; Campbell, J.M.; Crenshaw, J.; Segales, J.; Pujols, J. Neutralizing antibodies against porcine circovirus type 2 in liquid pooled plasma contribute to the biosafety of commercially manufactured spray-dried porcine plasma. J. Anim. Sci. 2013, 91, 2192–2198. [Google Scholar] [CrossRef] [PubMed]
- Seo, H.W.; Han, K.; Oh, Y.; Park, C.; Chae, C. Efficacy of a reformulated inactivated chimeric PCV1-2 vaccine based on clinical, virological, pathological and immunological examination under field conditions. Vaccine 2012, 30, 6671–6677. [Google Scholar] [CrossRef]
- Lee, S.-I.; Jeong, C.-G.; Nazki, S.; Aganja, R.P.; Kim, S.-C.; Khatun, A.; Oh, Y.; Noh, S.-H.; Lee, S.-M.; Kim, W.-I. Protective immunity induced by concurrent intradermal injection of porcine circovirus type 2 and Mycoplasma hyopneumoniae inactivated vaccines in pigs. Vaccine 2021, 39, 6691–6699. [Google Scholar] [CrossRef]
- Opriessnig, T.; Prickett, J.R.; Madson, D.M.; Shen, H.G.; Juhan, N.M.; Pogranichniy, R.R.; Meng, X.J.; Halbur, P.G. Porcine circovirus type 2 (PCV2)-infection and re-inoculation with homologous or heterologous strains: Virological, serological, pathological and clinical effects in growing pigs. Vet Res 2010, 41, 31. [Google Scholar] [CrossRef] [PubMed]
- Opriessnig, T.; Xiao, C.T.; Gerber, P.F.; Halbur, P.G. Emergence of a novel mutant PCV2b variant associated with clinical PCVAD in two vaccinated pig farms in the U.S. concurrently infected with PPV2. Vet. Microbiol. 2013, 163, 177–183. [Google Scholar] [CrossRef]
- Brunborg, I.M.; Moldal, T.; Jonassen, C.M. Quantitation of porcine circovirus type 2 isolated from serum/plasma and tissue samples of healthy pigs and pigs with postweaning multisystemic wasting syndrome using a TaqMan-based real-time PCR. J. Virol. Methods 2004, 122, 171–178. [Google Scholar] [CrossRef]
- Opriessnig, T.; Thacker, E.L.; Yu, S.; Fenaux, M.; Meng, X.J.; Halbur, P.G. Experimental reproduction of postweaning multisystemic wasting syndrome in pigs by dual infection with Mycoplasma hyopneumoniae and porcine circovirus type 2. Vet. Pathol. 2004, 41, 624–640. [Google Scholar] [CrossRef] [Green Version]
- Opriessnig, T.; Ramamoorthy, S.; Madson, D.M.; Patterson, A.R.; Pal, N.; Carman, S.; Meng, X.J.; Halbur, P.G. Differences in virulence among porcine circovirus type 2 isolates are unrelated to cluster type 2a or 2b and prior infection provides heterologous protection. J. Gen. Virol. 2008, 89, 2482–2491. [Google Scholar] [CrossRef] [PubMed]
- Opriessnig, T.; Karuppannan, A.K.; Halbur, P.G.; Calvert, J.G.; Nitzel, G.P.; Matzinger, S.R.; Meng, X.-J. Porcine circovirus type 2a or 2b based experimental vaccines provide protection against PCV2d/porcine parvovirus 2 co-challenge. Vaccine 2020, 38, 1975–1981. [Google Scholar] [CrossRef] [PubMed]
- Opriessnig, T.; Patterson, A.; Elsener, J.; Meng, X.; Halbur, P. Influence of maternal antibodies on efficacy of porcine circovirus type 2 (PCV2) vaccination to protect pigs from experimental infection with PCV2. Clin. Vaccine Immunol. 2008, 15, 397–401. [Google Scholar] [CrossRef] [PubMed]
- Fort, M.; Sibila, M.; Allepuz, A.; Mateu, E.; Roerink, F.; Segalés, J. Porcine circovirus type 2 (PCV2) vaccination of conventional pigs prevents viremia against PCV2 isolates of different genotypes and geographic origins. Vaccine 2008, 26, 1063–1071. [Google Scholar] [CrossRef]
- Opriessnig, T.; Madson, D.; Prickett, J.; Kuhar, D.; Lunney, J.; Elsener, J.; Halbur, P. Effect of porcine circovirus type 2 (PCV2) vaccination on porcine reproductive and respiratory syndrome virus (PRRSV) and PCV2 coinfection. Vet. Microbiol. 2008, 131, 103–114. [Google Scholar] [CrossRef]
- Rose, N.; Andraud, M.; Bigault, L.; Jestin, A.; Grasland, B. A commercial PCV2a-based vaccine significantly reduces PCV2b transmission in experimental conditions. Vaccine 2016, 34, 3738–3745. [Google Scholar] [CrossRef]
- Kolyvushko, O.; Rakibuzzaman, A.; Pillatzki, A.; Webb, B.; Ramamoorthy, S. Efficacy of a Commercial PCV2a Vaccine with a Two-Dose Regimen Against PCV2d. Vet. Sci. 2019, 6, 61. [Google Scholar] [CrossRef] [PubMed]
- Gan, Y.; Xie, X.; Zhang, L.; Xiong, Q.; Shao, G.; Feng, Z. Establishment of a model of Mycoplasma hyopneumoniae infection using Bama miniature pigs. Food Prod. Process. Nutr. 2020, 2, 19. [Google Scholar] [CrossRef]
- Kim, H.; Song, K.D.; Kim, H.J.; Park, W.; Kim, J.; Lee, T.; Shin, D.-H.; Kwak, W.; Kwon, Y.-J.; Sung, S. Exploring the genetic signature of body size in Yucatan miniature pig. PLoS ONE 2015, 10, e0121732. [Google Scholar] [CrossRef]
- Smith, D.M.; Martens, G.W.; Ho, C.S.; Asbury, J.M. DNA sequence based typing of swine leukocyte antigens in Yucatan miniature pigs. Xenotransplantation 2005, 12, 481–488. [Google Scholar] [CrossRef] [PubMed]
- Panepinto, L.; Phillips, R. The Yucatan miniature pig: Characterization and utilization in biomedical research. Lab. Anim. Sci. 1986, 36, 344–347. [Google Scholar] [PubMed]
- Van Der Windt, D.J.; Bottino, R.; Kumar, G.; Wijkstrom, M.; Hara, H.; Ezzelarab, M.; Ekser, B.; Phelps, C.; Murase, N.; Casu, A. Clinical islet xenotransplantation: How close are we? Diabetes 2012, 61, 3046–3055. [Google Scholar] [CrossRef]
- Bassols, A.; Costa, C.; Eckersall, P.D.; Osada, J.; Sabria, J.; Tibau, J. The pig as an animal model for human pathologies: A proteomics perspective. PROTEOMICS–Clin. Appl. 2014, 8, 715–731. [Google Scholar] [CrossRef] [PubMed]
- Bolin, S.R.; Stoffregen, W.C.; Nayar, G.P.; Hamel, A.L. Postweaning multisystemic wasting syndrome induced after experimental inoculation of cesarean-derived, colostrum-deprived piglets with type 2 porcine circovirus. J. Vet. Diagn. Investig. 2001, 13, 185–194. [Google Scholar] [CrossRef] [PubMed]
- Okuda, Y.; Ono, M.; Yazawa, S.; Shibata, I. Experimental reproduction of postweaning multisystemic wasting syndrome in cesarean-derived, colostrum-deprived piglets inoculated with porcine circovirus type 2 (PCV2): Investigation of quantitative PCV2 distribution and antibody responses. J. Vet. Diagn. Investig. 2003, 15, 107–114. [Google Scholar] [CrossRef]
- Harms, P.; Sorden, S.; Halbur, P.; Bolin, S.; Lager, K.; Morozov, I.; Paul, P. Experimental reproduction of severe disease in CD/CD pigs concurrently infected with type 2 porcine circovirus and porcine reproductive and respiratory syndrome virus. Vet. Pathol. 2001, 38, 528–539. [Google Scholar] [CrossRef]
- Allan, G.; Kennedy, S.; McNeilly, F.; Foster, J.; Ellis, J.; Krakowka, S.; Meehan, B.; Adair, B. Experimental reproduction of severe wasting disease by co-infection of pigs with porcine circovirus and porcine parvovirus. J. Comp. Pathol. 1999, 121, 1–11. [Google Scholar] [CrossRef]
- Krakowka, S.; Ellis, J.; Meehan, B.; Kennedy, S.; McNeilly, F.; Allan, G. Viral wasting syndrome of swine: Experimental reproduction of postweaning multisystemic wasting syndrome in gnotobiotic swine by coinfection with porcine circovirus 2 and porcine parvovirus. Vet. Pathol. 2000, 37, 254–263. [Google Scholar] [CrossRef]
- Fort, M.; Olvera, A.; Sibila, M.; Segalés, J.; Mateu, E. Detection of neutralizing antibodies in postweaning multisystemic wasting syndrome (PMWS)-affected and non-PMWS-affected pigs. Vet. Microbiol. 2007, 125, 244–255. [Google Scholar] [CrossRef] [PubMed]
- Huan, C.; Fan, M.; Cheng, Q.; Wang, X.; Gao, Q.; Wang, W.; Gao, S.; Liu, X. Evaluation of the efficacy and cross-protective immunity of live-attenuated chimeric PCV1-2b vaccine against PCV2b and PCV2d subtype challenge in pigs. Front. Microbiol. 2018, 9, 455. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matzinger, S.R.; Opriessnig, T.; Xiao, C.-T.; Catanzaro, N.; Beach, N.M.; Slade, D.E.; Nitzel, G.P.; Meng, X.-J. A chimeric virus created by DNA shuffling of the capsid genes of different subtypes of porcine circovirus type 2 (PCV2) in the backbone of the non-pathogenic PCV1 induces protective immunity against the predominant PCV2b and the emerging PCV2d in pigs. Virology 2016, 498, 82–93. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer | Nucleotide Sequence (5′-3′) | Product Size | Purpose |
|---|---|---|---|
| F-PCV2C | GCT GGC TGA ACT TTT GAA AGT | 1767 bp | PCV2 detection and sequencing |
| R-PCV2C | AAA TTT CTG ACA AAC GTT ACA | ||
| PCV2ab 2NF | GGT TGG AAG TAA TCA ATA GTG GA | 277 bp | PCV2a-specific |
| PCV2a 2NR | GGG GAA CCA ACA AAA TCT C | ||
| PCV2ab 2NF | GGT TGG AAG TAA TCA ATA GTG GA | 277 bp | PCV2b-specific |
| PCV2b 2NR | GGG GCT CAA ACC CCC GCT C | ||
| PCV2d 2NF | GGT TGG AAG TAA TCG ATT GTC CT | 343 bp | PCV2d-specific |
| PCV2d 2NR | TCA GAA CGC CCT CCT GGA AT |
| Score | 0 Points | 1 Point | 2 Points | 3 Points |
|---|---|---|---|---|
| Lymphoid depletion | Not present | Mild, with loss of overall cellularity | Moderate | Severe, with loss of lymphoid follicle structure |
| Histiocytic-to-granulomatous inflammation | Not present | Mild | Moderate | Severe, with replacement of follicles |
| PCV2 antigen * | Not present | Less than 10% | 10–50% | More than 50% |
| PCV2b-Vac | PCV2d-Vac | Non-Vac | ||
|---|---|---|---|---|
| Lung macroscopic lesion | Dorsal right lobe | 2.4 ± 1.5 a | 1.2 ± 0.6 a | 18.3 ± 0.6 b |
| Dorsal left lobe | 3.0 ± 1.0 a | 1.6 ± 0.5 a | 17.7 ± 1.5 b | |
| Accessory lobe | 1.1 ± 0.5 | 0.8 ± 0.3 | 2.7 ± 0.6 | |
| Ventral right lobe | 3.2 ± 0.8 a | 2.2 ± 0.4 a | 21.3 ± 2.1 b | |
| Ventral left lobe | 2.8 ± 0.8 a | 2.2 ± 1.1 a | 19.3 ± 3.5 b | |
| Average of sum | 12.5 ± 3.7 a | 8.0 ± 1.5 b | 79.3 ± 6.8 c | |
| Lymphoid depletion | Lung | 0.2 ± 0.4 a | 0.0 ± 0.0 a | 2.0 ± 0.0 b |
| Lymph node | 0.8 ± 0.4 a | 0.8 ± 0.4 a | 2.7 ± 0.6 b | |
| Spleen | 0.0 ± 0.0 a | 0.0 ± 0.0a | 2.3 ± 0.6 b | |
| Histiocytic-to-granulomatous inflammation | Lung | 0.8 ± 0.4 a | 0.6 ± 0.5 a | 2.7 ± 0.6 b |
| Lymph node | 0.0 ± 0.0 a | 0.0 ± 0.0 a | 2.7 ± 0.6 b | |
| Spleen | 0.0 ± 0.0 a | 0.0 ± 0.0 a | 2.3 ± 0.6 b | |
| PCV2 antigen * | Lung | 0.0 ± 0.0 a | 0.0 ± 0.0 a | 1.3 ± 0.6 b |
| Lymph node | 0.0 ± 0.0 a | 0.0 ± 0.0 a | 2.0 ± 0.0 b | |
| Spleen | 0.0 ± 0.0 a | 0.0 ± 0.0 a | 1.3 ± 0.6 b |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Noh, Y.-H.; Kim, S.-C.; Jeong, C.-G.; Lee, S.-C.; Lee, D.-U.; Yoon, I.-J.; Kim, W.-I. Pathological Evaluation of Porcine Circovirus 2d (PCV2d) Strain and Comparative Evaluation of PCV2d and PCV2b Inactivated Vaccines against PCV2d Infection in a Specific Pathogen-Free (SPF) Yucatan Miniature Pig Model. Vaccines 2022, 10, 1469. https://doi.org/10.3390/vaccines10091469
Noh Y-H, Kim S-C, Jeong C-G, Lee S-C, Lee D-U, Yoon I-J, Kim W-I. Pathological Evaluation of Porcine Circovirus 2d (PCV2d) Strain and Comparative Evaluation of PCV2d and PCV2b Inactivated Vaccines against PCV2d Infection in a Specific Pathogen-Free (SPF) Yucatan Miniature Pig Model. Vaccines. 2022; 10(9):1469. https://doi.org/10.3390/vaccines10091469
Chicago/Turabian StyleNoh, Yun-Hee, Seung-Chai Kim, Chang-Gi Jeong, Seung-Chul Lee, Dong-Uk Lee, In-Joong Yoon, and Won-Il Kim. 2022. "Pathological Evaluation of Porcine Circovirus 2d (PCV2d) Strain and Comparative Evaluation of PCV2d and PCV2b Inactivated Vaccines against PCV2d Infection in a Specific Pathogen-Free (SPF) Yucatan Miniature Pig Model" Vaccines 10, no. 9: 1469. https://doi.org/10.3390/vaccines10091469
APA StyleNoh, Y.-H., Kim, S.-C., Jeong, C.-G., Lee, S.-C., Lee, D.-U., Yoon, I.-J., & Kim, W.-I. (2022). Pathological Evaluation of Porcine Circovirus 2d (PCV2d) Strain and Comparative Evaluation of PCV2d and PCV2b Inactivated Vaccines against PCV2d Infection in a Specific Pathogen-Free (SPF) Yucatan Miniature Pig Model. Vaccines, 10(9), 1469. https://doi.org/10.3390/vaccines10091469