
Safety and Immunogenicity of Inactivated Bacillus subtilis Spores as a Heterologous Antibody Booster for COVID-19 Vaccines

 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Trial Objectives, Participants and Oversight
2.2. Trial Procedures
2.3. Safety
2.4. Statistical Analysis
3. Results
3.1. Characteristics of the Participants
3.2. Safety
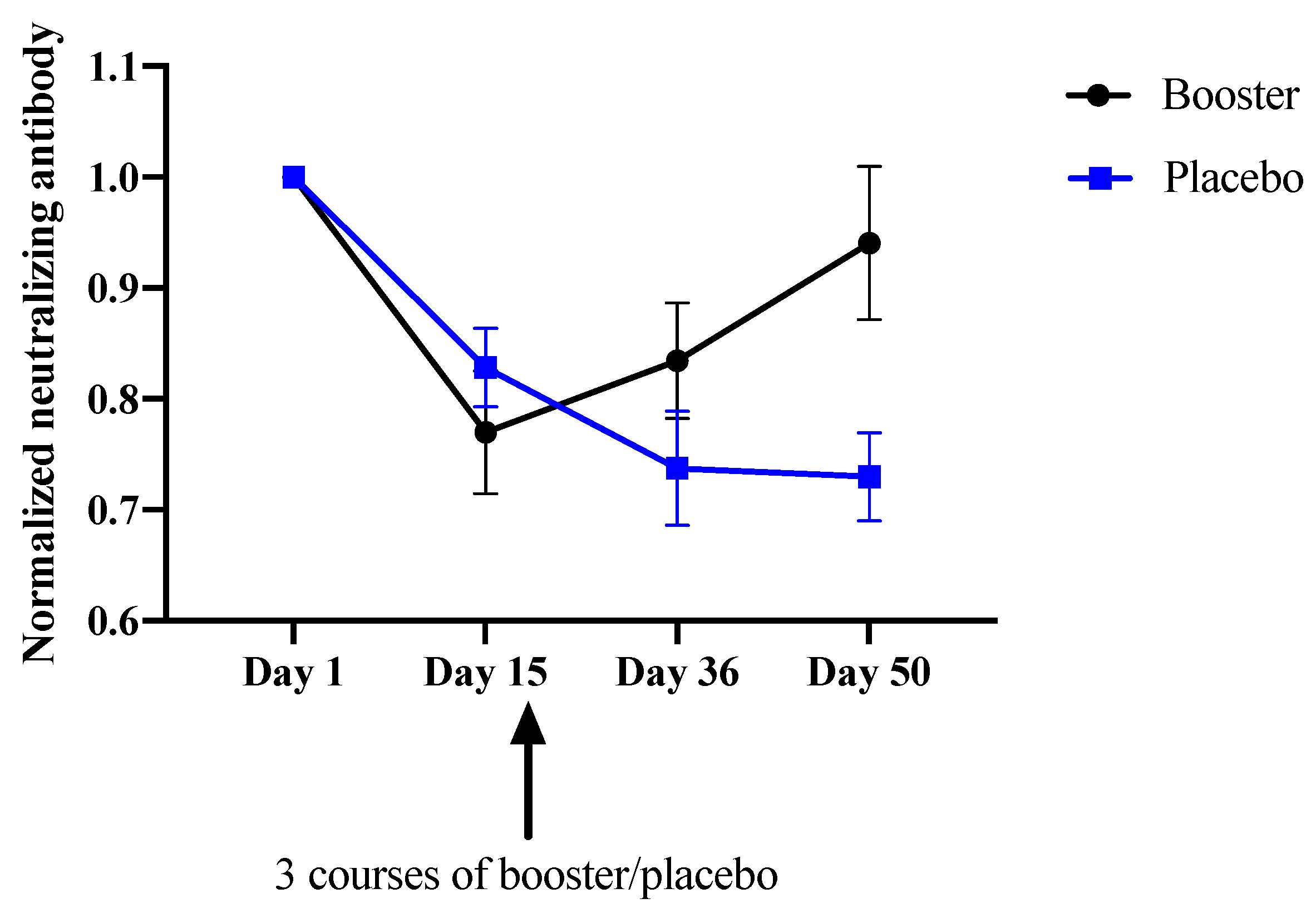
3.3. Immunoreactivity
4. Discussion
5. Patents
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- WHO Coronavirus (COVID-19) Dashboard. Available online: https://covid19.who.int (accessed on 11 May 2022).
- COVID-19 Vaccine Tracker and Landscape. Available online: https://www.who.int/publications/m/item/draft-landscape-of-covid-19-candidate-vaccines (accessed on 11 May 2022).
- Rosenberg, E.S.; Dorabawila, V.; Easton, D.; Bauer, U.E.; Kumar, J.; Hoen, R.; Hoefer, D.; Wu, M.; Lutterloh, E.; Conroy, M.B.; et al. Covid-19 Vaccine Effectiveness in New York State. New Engl. J. Med. 2022, 386, 116–127. [Google Scholar] [CrossRef] [PubMed]
- CDC. COVID Data Tracker. Centers for Disease Control and Prevention. 28 March 2020. Available online: https://covid.cdc.gov/covid-data-tracker (accessed on 12 May 2022).
- Choi, A.; Koch, M.; Wu, K.; Chu, L.; Ma, L.; Hill, A.; Nunna, N.; Huang, W.; Oestreicher, J.; Colpitts, T.; et al. Safety and immunogenicity of SARS-CoV-2 variant mRNA vaccine boosters in healthy adults: An interim analysis. Nat. Med. 2021, 27, 11. [Google Scholar] [CrossRef] [PubMed]
- Feleszko, W.; Lewulis, P.; Czarnecki, A.; Waszkiewicz, P. Flattening the Curve of COVID-19 Vaccine Rejection—An International Overview. Vaccines 2021, 9, 44. [Google Scholar] [CrossRef] [PubMed]
- Bednarczyk, R.A. Examining the ‘why’ of vaccine hesitancy. Health Psychol. 2018, 37, 316–317. [Google Scholar] [CrossRef]
- Hornsey, M.J.; Harris, E.A.; Fielding, K.S. The psychological roots of anti-vaccination attitudes: A 24-nation investigation. Health Psychol. 2018, 37, 307–315. [Google Scholar] [CrossRef]
- Kersten, G.; Hirschberg, H. Needle-free vaccine delivery. Expert Opin. Drug Deliv. 2007, 4, 459–474. [Google Scholar] [CrossRef]
- Hong, H.A.; Huang, J.-M.; Khaneja, R.; Hiep, L.V.; Urdaci, M.C.; Cutting, S.M. The safety of Bacillus subtilis and Bacillus indicus as food probiotics. J. Appl. Microbiol. 2008, 105, 510–520. [Google Scholar] [CrossRef]
- Potocki, W.; Negri, A.; Peszyńska-Sularz, G.; Hinc, K.; Obuchowski, M.; Iwanicki, A. IL-1 Fragment Modulates Immune Response Elicited by Recombinant Bacillus subtilis Spores Presenting an Antigen/Adjuvant Chimeric Protein. Mol. Biotechnol. 2018, 60, 810–819. [Google Scholar] [CrossRef] [Green Version]
- Amuguni, H.; Tzipori, S. Bacillus subtilis. Hum. Vaccines Immunother. 2012, 8, 979–986. [Google Scholar] [CrossRef] [Green Version]
- Barnes, A.G.C.; Cerovic, V.; Hobson, P.S.; Klavinskis, L.S. Bacillus subtilis spores: A novel microparticle adjuvant which can instruct a balanced Th1 and Th2 immune response to specific antigen. Eur. J. Immunol. 2007, 37, 1538–1547. [Google Scholar] [CrossRef]
- Paccez, J.D.; Luiz, W.B.; Sbrogio-Almeida, M.E.; Ferreira, R.C.C.; Schumann, W.; Ferreira, L.C.S. Stable episomal expression system under control of a stress inducible promoter enhances the immunogenicity of Bacillus subtilis as a vector for antigen delivery. Vaccine 2006, 24, 2935–2943. [Google Scholar] [CrossRef] [PubMed]
- Song, M.; Hong, H.A.; Huang, J.M.; Colenutt, C.; Khang, D.D.; Nguyen, T.V.A.; Park, S.M.; Shim, B.S.; Song, H.H.; Cheon, I.S.; et al. Killed Bacillus subtilis spores as a mucosal adjuvant for an H5N1 vaccine. Vaccine 2012, 30, 3266–3277. [Google Scholar] [CrossRef] [PubMed]
- WHO: An Unwavering Voice for Vaccine Equity. Available online: https://www.who.int/nepal/news/detail/17-05-2022-who--an-unwavering-voice-for-vaccine-equity (accessed on 14 June 2022).
- Uddin, M.N.; Roni, M.A. Challenges of Storage and Stability of mRNA-Based COVID-19 Vaccines. Vaccines 2021, 9, 1033. [Google Scholar] [CrossRef] [PubMed]
- Lin, D.Y.; Gu, Y.; Wheeler, B.; Young, H.; Holloway, S.; Sunny, S.K.; Moore, Z.; Zeng, D. Effectiveness of Covid-19 Vaccines over a 9-Month Period in North Carolina. New Engl. J. Med. 2022, 386, 933–941. [Google Scholar] [CrossRef] [PubMed]
- Thomas, S.J.; Moreira, E.D., Jr.; Kitchin, N.; Absalon, J.; Gurtman, A.; Lockhart, S.; Perez, J.L.; Pérez Marc, G.; Polack, F.P.; Zerbini, C.; et al. Safety and Efficacy of the BNT162b2 mRNA Covid-19 Vaccine through 6 Months. New Engl. J. Med. 2021, 385, 1761–1773. [Google Scholar] [CrossRef]
- Sung, J.C.C.; Liu, Y.; Wu, K.C.; Choi, M.C.; Ma, C.H.Y.; Lin, J.; He, E.I.C.; Leung, D.Y.M.; Sze, E.T.P.; Hamied, Y.K.; et al. Expression of SARS-CoV-2 Spike Protein Receptor Binding Domain on Recombinant, B. subtilis on Spore Surface: A Potential COVID-19 Oral Vaccine Candidate. Vaccines 2022, 10, 2. [Google Scholar] [CrossRef]
- Walsh, E.E.; Frenck, R.W., Jr.; Falsey, A.R.; Kitchin, N.; Absalon, J.; Gurtman, A.; Lockhart, S.; Neuzil, K.; Mulligan, M.J.; Bailey, R.; et al. Safety and Immunogenicity of Two RNA-Based Covid-19 Vaccine Candidates. New Engl. J. Med. 2020, 383, 2439–2450. [Google Scholar] [CrossRef]
- Xia, S.; Zhang, Y.; Wang, Y.; Wang, H.; Yang, Y.; Gao, G.F.; Tan, W.; Wu, G.; Xu, M.; Lou, Z.; et al. Safety and immunogenicity of an inactivated SARS-CoV-2 vaccine, BBIBP-CorV: A randomised, double-blind, placebo-controlled, phase 1/2 trial. Lancet Infect. Dis. 2021, 21, 39–51. [Google Scholar] [CrossRef]
- Yan, R.; Zhang, Y.; Li, Y.; Xia, L.; Guo, Y.; Zhou, Q. Structural basis for the recognition of SARS-CoV-2 by full-length human ACE2. Science 2020, 367, 1444–1448. [Google Scholar] [CrossRef] [Green Version]
- Setlow, P. Spore germination. Curr. Opin. Microbiol. 2003, 6, 550–556. [Google Scholar] [CrossRef]
- Setlow, P. Spores of Bacillus subtilis: Their resistance to and killing by radiation, heat and chemicals. J. Appl. Microbiol. 2006, 101, 514–525. [Google Scholar] [CrossRef] [PubMed]
- Matula, Z.; Gönczi, M.; Bekő, G.; Kádár, B.; Ajzner, É.; Uher, F.; Vályi-Nagy, I. Antibody and T Cell Responses against SARS-CoV-2 Elicited by the Third Dose of BBIBP-CorV (Sinopharm) and BNT162b2 (Pfizer-BioNTech) Vaccines Using a Homologous or Heterologous Booster Vaccination Strategy. Vaccines 2022, 10, 539. [Google Scholar] [CrossRef] [PubMed]
- Wu, A.H.B.; Nguyen, E.D.; Ong, C.M.; Yun, C.; Lynch, K.L. Rate of Serum SARS-CoV-2 Antibody Decline for Two mRNA Vaccines. J. Appl. Lab. Med. 2022, 7, 625–627. [Google Scholar] [CrossRef]
- Sughayer, M.A.; Souan, L.; Alhowr, M.A.; Al Rimawi, D.; Siag, M.; Albadr, S.; Owdeh, M.; Al Atrash, T. Comparison of the effectiveness and duration of anti-RBD SARS-CoV-2 IgG antibody response between different types of vaccines: Implications for vaccine strategies. Vaccine 2022, 40, 2841–2847. [Google Scholar] [CrossRef]
- Harwood, C.R. Bacillus subtilis and its relatives: Molecular biological and industrial workhorses. Trends Biotechnol. 1992, 10, 247–256. [Google Scholar] [CrossRef]
- Yu, J.; Collins, N.D.; Mercado, N.B.; McMahan, K.; Chandrashekar, A.; Liu, J.; Anioke, T.; Chang, A.; Giffin, V.M.; Hope, D.L.; et al. Protective Efficacy of Gastrointestinal SARS-CoV-2 Delivery against Intranasal and Intratracheal SARS-CoV-2 Challenge in Rhesus Macaques. J. Virol. 2022, 96, e0159921. [Google Scholar] [CrossRef]
- Maeda, N.; Nakamura, R.; Hirose, Y.; Murosaki, S.; Yamamoto, Y.; Kase, T.; Yoshikai, Y. Oral administration of heat-killed Lactobacillus plantarum L-137 enhances protection against influenza virus infection by stimulation of type I interferon production in mice. Int. Immunopharmacol. 2009, 9, 1122–1125. [Google Scholar] [CrossRef]
- Rhee, K.-J.; Sethupathi, P.; Driks, A.; Lanning, D.K.; Knight, K.L. Role of commensal bacteria in development of gut-associated lymphoid tissues and preimmune antibody repertoire. J. Immunol. Baltim. Md. 1950 2004, 172, 1118–1124. [Google Scholar] [CrossRef] [Green Version]
- Severson, K.M.; Mallozzi, M.; Driks, A.; Knight, K.L. B Cell Development in GALT: Role of Bacterial Superantigen-Like Molecules. J. Immunol. 2010, 184, 6782–6789. [Google Scholar] [CrossRef] [Green Version]
- Lanning, D.; Sethupathi, P.; Rhee, K.-J.; Zhai, S.-K.; Knight, K.-L. Intestinal Microflora and Diversification of the Rabbit Antibody Repertoire. J. Immunol. 2000, 165, 2012–2019. [Google Scholar] [CrossRef]
- Hooper, L.V.; Wong, M.H.; Thelin, A.; Hansson, L.; Falk, P.G.; Gordon, J.I. Molecular analysis of commensal host-microbial relationships in the intestine. Science 2001, 291, 881–884. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, N.-K.; Kim, W.-S.; Paik, H.-D. Bacillus strains as human probiotics: Characterization, safety, microbiome, and probiotic carrier. Food Sci. Biotechnol. 2019, 28, 1297–1305. [Google Scholar] [CrossRef] [PubMed]
- Rhayat, L.; Maresca, M.; Nicoletti, C.; Perrier, J.; Brinch, K.S.; Christian, S.; Devillard, E.; Eckhardt, E. Effect of Bacillus subtilis Strains on Intestinal Barrier Function and Inflammatory Response. Front. Immunol. 2019, 10, 564. Available online: https://www.frontiersin.org/article/10.3389/fimmu.2019.00564 (accessed on 15 June 2022).
- Khodavirdipour, A.; Chamanrokh, P.; Alikhani, M.Y.; Alikhani, M.S. Potential of Bacillus subtilis Against SARS-CoV-2—A Sustainable Drug Development Perspective. Front. Microbiol. 2022, 13, 718786. [Google Scholar] [CrossRef]
- Shahbaz, F.F.; Martins, R.S.; Umair, A.; Ukrani, R.D.; Jabeen, K.; Sohail, M.R.; Khan, E. A Review of Coronaviruses Associated with Kawasaki Disease: Possible Implications for Pathogenesis of the Multisystem Inflammatory Syndrome Associated With COVID-19. Clin. Med. Insights Pediatr. 2022, 16, 11795565221075320. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
| Treatment | Previous Vaccination Received | Level of Neutralizing Antibody (μg/mL) | |||
|---|---|---|---|---|---|
| Day 1 | Day 15 | Day 36 | Day 50 | ||
| Booster #1 | BNT162b2 | 1.030 | 0.907 | 0.900 | 0.929 |
| Booster #2 | BNT162b2 | 0.731 | 0.682 | 0.687 | 0.678 |
| Booster #3 | BNT162b2 | 0.537 | 0.576 | 0.601 | 0.566 |
| Booster #4 | BBIBP | 0.119 | 0.082 | 0.099 | 0.115 |
| Booster #5 | BBIBP | 0.045 | 0.030 | 0.030 | 0.034 |
| Booster #6 | BBIBP | 0.077 | 0.047 | 0.062 | 0.063 |
| Booster #7 | BBIBP | 0.047 | 0.030 | 0.030 | 0.066 |
| Booster #8 | BBIBP | 0.071 | 0.043 | 0.049 | 0.049 |
| Booster #9 | BBIBP | 0.036 | 0.030 | 0.034 | 0.034 |
| Placebo #1 | BNT162b2 | 0.825 | 0.719 | 0.698 | 0.608 |
| Placebo #2 | BNT162b2 | 0.542 | 0.498 | 0.459 | 0.461 |
| Placebo #3 | BBIBP | 0.284 | 0.233 | 0.248 | 0.246 |
| Placebo #4 | BBIBP | 0.040 | 0.034 | 0.032 | 0.030 |
| Placebo #5 | BBIBP | 0.103 | 0.080 | 0.055 | 0.067 |
| Placebo #6 | BBIBP | 0.492 | 0.448 | 0.320 | 0.334 |
| Placebo #7 | BBIBP | 0.108 | 0.070 | 0.066 | 0.062 |
| Treatment | Previous Vaccination Received | Day 15 | Day 36 | Day 50 |
|---|---|---|---|---|
| Booster #1 | BNT162b2 | 0.881 | 0.874 | 0.902 |
| Booster #2 | BNT162b2 | 0.933 | 0.940 | 0.927 |
| Booster #3 | BNT162b2 | 1.073 | 1.119 | 1.054 |
| Booster #4 | BBIBP | 0.689 | 0.832 | 0.966 |
| Booster #5 | BBIBP | 0.667 | 0.667 | 0.756 |
| Booster #6 | BBIBP | 0.610 | 0.805 | 0.818 |
| Booster #7 | BBIBP | 0.638 | 0.638 | 1.404 |
| Booster #8 | BBIBP | 0.606 | 0.690 | 0.690 |
| Booster #9 | BBIBP | 0.833 | 0.944 | 0.944 |
| Placebo #1 | BNT162b2 | 0.872 | 0.846 | 0.737 |
| Placebo #2 | BNT162b2 | 0.919 | 0.847 | 0.851 |
| Placebo #3 | BBIBP | 0.820 | 0.873 | 0.866 |
| Placebo #4 | BBIBP | 0.850 | 0.800 | 0.750 |
| Placebo #5 | BBIBP | 0.777 | 0.534 | 0.650 |
| Placebo #6 | BBIBP | 0.911 | 0.650 | 0.679 |
| Placebo #7 | BBIBP | 0.648 | 0.611 | 0.574 |
| Previous Vaccination Received | Day 15 | Day 36 | Day 50 | |
|---|---|---|---|---|
| Booster Group | BNT162b2 | 0.962 ± 0.099 * | 0.978 ± 0.127 | 0.961 ± 0.081 |
| BBIBP | 0.674 ± 0.084 * | 0.801 ± 0.098 | 0.930 ± 0.256 | |
| Overall | 0.770 ± 0.166 | 0.834 ± 0.156 | 0.940 ± 0.207 | |
| Placebo Group | BNT162b2 | 0.895 ± 0.033 | 0.846 ± 0.001 | 0.794 ± 0.080 |
| BBIBP | 0.801 ± 0.098 | 0.694 ± 0.139 | 0.704 ± 0.110 | |
| Overall | 0.828 ± 0.094 | 0.737 ± 0.136 | 0.730 ± 0.105 |
| Source of Variation | % of Total Variation | p Value | Significance |
|---|---|---|---|
| Time × Treatment | 9.807 | 0.0108 | * |
| Time | 27.68 | 0.0001 | *** |
| Treatment | 3.673 | 0.1861 | not significant |
| Subject | 26.60 | 0.0127 | * |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sung, J.C.-C.; Lai, N.C.-Y.; Wu, K.-C.; Choi, M.-C.; Ma, C.H.-Y.; Lin, J.; Kuok, C.-N.; Leong, W.-L.; Lam, W.-K.; Hamied, Y.K.; et al. Safety and Immunogenicity of Inactivated Bacillus subtilis Spores as a Heterologous Antibody Booster for COVID-19 Vaccines. Vaccines 2022, 10, 1014. https://doi.org/10.3390/vaccines10071014
Sung JC-C, Lai NC-Y, Wu K-C, Choi M-C, Ma CH-Y, Lin J, Kuok C-N, Leong W-L, Lam W-K, Hamied YK, et al. Safety and Immunogenicity of Inactivated Bacillus subtilis Spores as a Heterologous Antibody Booster for COVID-19 Vaccines. Vaccines. 2022; 10(7):1014. https://doi.org/10.3390/vaccines10071014
Chicago/Turabian StyleSung, Johnny Chun-Chau, Nelson Cheuk-Yin Lai, Kam-Chau Wu, Man-Chung Choi, Chloe Ho-Yi Ma, Jayman Lin, Cheong-Nang Kuok, Wai-Leng Leong, Weng-Kei Lam, Yusuf Khwaja Hamied, and et al. 2022. "Safety and Immunogenicity of Inactivated Bacillus subtilis Spores as a Heterologous Antibody Booster for COVID-19 Vaccines" Vaccines 10, no. 7: 1014. https://doi.org/10.3390/vaccines10071014
APA StyleSung, J. C.-C., Lai, N. C.-Y., Wu, K.-C., Choi, M.-C., Ma, C. H.-Y., Lin, J., Kuok, C.-N., Leong, W.-L., Lam, W.-K., Hamied, Y. K., Lam, D. M.-K., Sze, E. T.-P., & Kwong, K. W.-Y. (2022). Safety and Immunogenicity of Inactivated Bacillus subtilis Spores as a Heterologous Antibody Booster for COVID-19 Vaccines. Vaccines, 10(7), 1014. https://doi.org/10.3390/vaccines10071014