
An Overview of Vaccine Adjuvants: Current Evidence and Future Perspectives



Abstract
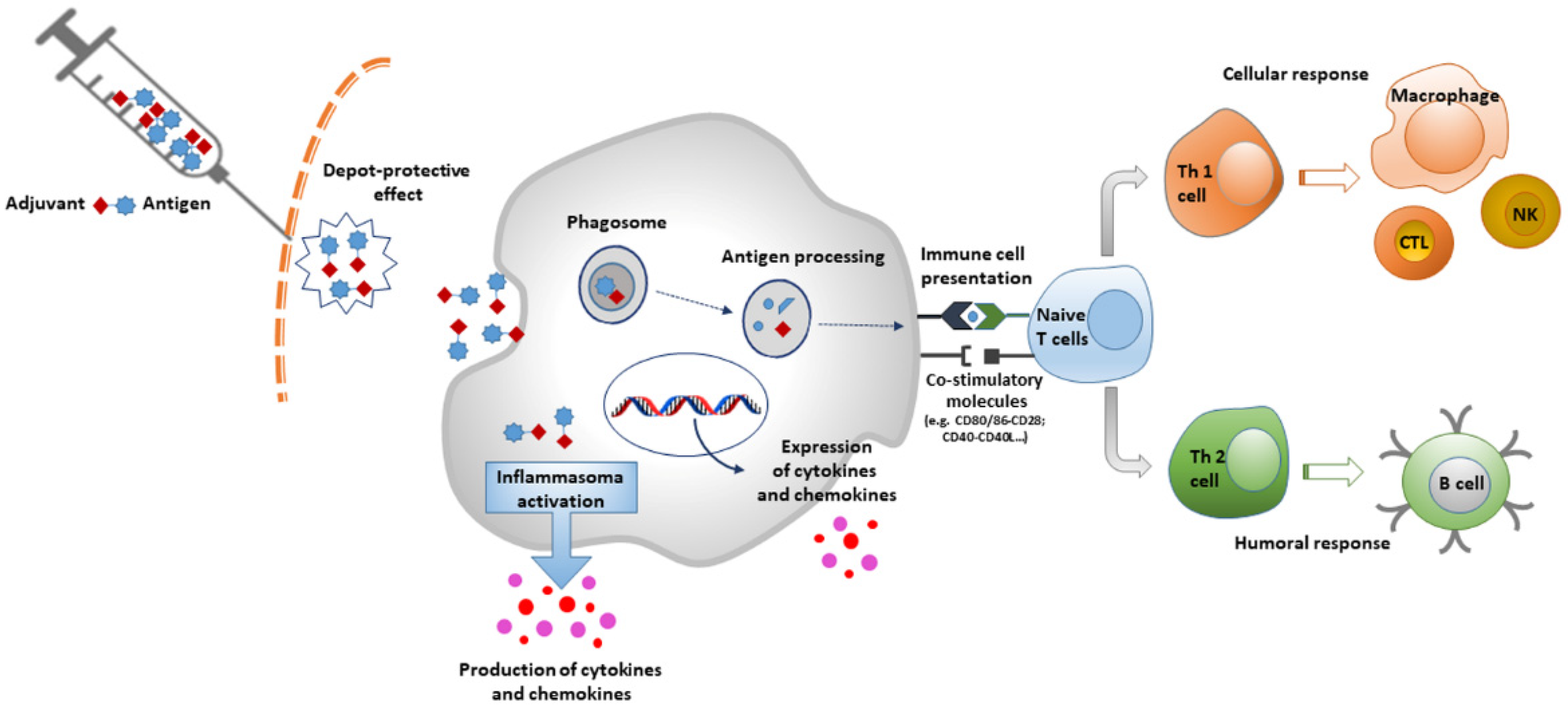
1. Introduction
2. Delivery Systems
2.1. Mineral Salts
Aluminium Salts
2.2. Emulsions
2.2.1. MF59
2.2.2. AS03
2.3. Microparticles
2.3.1. Virus-Like Particles
2.3.2. Virosomes
3. Immune Potentiators
3.1. TLR1/2 Agonists
TLR3 Agonists
3.2. TLR4 Agonists
3.3. TLR5 Agonists
3.4. TLR7/8 Agonists
3.5. TLR9 Agonists
4. Potential Side Effects of Adjuvants: The Asia Syndrome
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Centers for Disease Control and Prevention (CDC). Ten great public health achievements—Worldwide, 2001–2010. MMWR Morb. Mortal Wkly Rep. 2011, 60, 814–818. [Google Scholar]
- Bechini, A.; Boccalini, S.; Ninci, A.; Zanobini, P.; Sartor, G.; Bonaccorsi, G.; Grazzini, M.; Bonanni, P. Childhood vaccination coverage in Europe: Impact of different public health policies. Expert Rev. Vaccines 2019, 18, 693–701. [Google Scholar] [CrossRef]
- Di Pietro, A.; Visalli, G.; Antonuccio, G.M.; Facciolà, A. Today’s vaccination policies in Italy: The National Plan for Vaccine Prevention 2017–2019 and the Law 119/2017 on the mandatory vaccinations. Ann. Ig. 2019, 31 (Suppl. 1), 54–64. [Google Scholar]
- Vanderslott, S.; Marks, T. Charting mandatory childhood vaccination policies worldwide. Vaccine 2021, 39, 4054–4062. [Google Scholar] [CrossRef]
- Apostólico, J.S.; Lunardelli, V.A.; Coirada, F.C.; Boscardin, S.B.; Rosa, D.S. Adjuvants: Classification, Modus Operandi, and Licensing. J. Immunol. Res. 2016, 2016, 1459394. [Google Scholar] [CrossRef]
- Singh, M.; O’Hagan, D.T. Recent advances in vaccine adjuvants. Pharm. Res. 2002, 19, 715–728. [Google Scholar] [CrossRef]
- Pashine, A.; Valiante, N.M.; Ulmer, J.B. Targeting the innate immune response with improved vaccine adjuvants. Nat. Med. 2005, 11, S63–S68. [Google Scholar] [CrossRef]
- Shah, R.R.; Hassett, K.J.; Brito, L.A. Overview of Vaccine Adjuvants: Introduction, History, and Current Status. Methods Mol. Biol. 2017, 1494, 1–13. [Google Scholar]
- Olive, C. Pattern recognition receptors: Sentinels in innate immunity and targets of new vaccine adjuvants. Expert Rev. Vaccines 2012, 11, 237–256. [Google Scholar] [CrossRef]
- Crooke, S.N.; Ovsyannikova, I.G.; Poland, G.A.; Kennedy, R.B. Immunosenescence and human vaccine immune responses. Immun. Ageing 2019, 13, 25. [Google Scholar] [CrossRef]
- Skwarczynski, M.; Toth, I. Peptide-based synthetic vaccines. Chem. Sci. 2016, 7, 842–854. [Google Scholar] [CrossRef]
- Poland, G.A. Prevention of meningococcal disease: Current use of polysaccharide and conjugate vaccines. Clin. Infect. Dis. 2010, 50 (Suppl. 2), S45–S53. [Google Scholar] [CrossRef]
- European Medicinal Agency (EMA). Nuvaxoid. Summary of Product Characteristics. Available online: https://www.ema.europa.eu/en/documents/product-information/nuvaxovid-epar-product-information_en.pdf (accessed on 20 March 2022).
- Brito, L.A.; Malyala, P.; O’Hagan, D.T. Vaccine adjuvant formulations: A pharmaceutical perspective. Semin Immunol. 2013, 25, 130–145. [Google Scholar] [CrossRef]
- Harrison, E.A.; Wu, J.W. Vaccine confidence in the time of COVID-19. Eur. J. Epidemiol. 2020, 35, 325–330. [Google Scholar] [CrossRef]
- Palamenghi, L.; Barello, S.; Boccia, S.; Graffigna, G. Mistrust in biomedical research and vaccine hesitancy: The forefront challenge in the battle against COVID-19 in Italy. Eur. J. Epidemiol. 2020, 35, 785–788. [Google Scholar] [CrossRef]
- Xiao, X.; Wong, R.M. Vaccine hesitancy and perceived behavioral control: A meta-analysis. Vaccine 2020, 38, 5131–5138. [Google Scholar] [CrossRef]
- Salmon, D.A.; Dudley, M.Z.; Glanz, J.M.; Omer, S.B. Vaccine hesitancy: Causes, consequences, and a call to action. Vaccine 2015, 33 (Suppl. 4), D66–D71. [Google Scholar] [CrossRef]
- Facciolà, A.; Visalli, G.; Orlando, A.; Bertuccio, M.P.; Spataro, P.; Squeri, R.; Picerno, I.; Di Pietro, A. Vaccine hesitancy: An overview on parents’ opinions about vaccination and possible reasons of vaccine refusal. J. Public Health Res. 2019, 8, 1436. [Google Scholar] [CrossRef]
- Kreps, S.; Prasad, S.; Brownstein, J.S.; Hswen, Y.; Garibaldi, B.T.; Zhang, B.; Kriner, D.L. Factors Associated with US Adults’ Likelihood of Accepting COVID-19 Vaccination. JAMA Netw. Open 2020, 3, e2025594. [Google Scholar] [CrossRef]
- Centers for Disease Control and Prevention (CDC). Possible Side effects from Vaccines. Available online: https://www.cdc.gov/vaccines/vac-gen/side-effects.htm (accessed on 1 February 2022).
- World Health Organization (WHO). Side Effects of COVID-19 Vaccines. Available online: https://www.who.int/news-room/feature-stories/detail/side-effects-of-covid-19-vaccines (accessed on 1 February 2022).
- Glenny, A.; Pope, C.G.; Waddington, H.; Wallace, U. Immunological notes XVII to XXIV. J. Pathol. 1926, 29, 31–40. [Google Scholar] [CrossRef]
- Hassett, K.J.; Cousins, M.C.; Rabia, L.A.; Chadwick, C.M.; O’Hara, J.M.; Nandi, P.; Brey, R.N.; Mantis, N.J.; Carpenter, J.F.; Randolph, T.W. Stabilization of a recombinant ricin toxin A subunit vaccine through lyophilization. Eur. J. Pharm. Biopharm. 2013, 85, 279–286. [Google Scholar] [CrossRef] [PubMed]
- Agence francaise de sécurite sanitaire des aliments (Afssa). Evaluation des Risques Sanitaires Liés à L’exposition de la Population Française à l’aluminium. Eaux, Aliments, Produits de Santé. Available online: https://www.santepubliquefrance.fr/docs/evaluation-des-risques-sanitaires-lies-a-l-exposition-de-la-population-francaise-a-l-aluminium.-eaux-aliments-produits-de-sante (accessed on 9 February 2022).
- Gourier-Fréry, C.; Fréry, N. Aluminium. EMC Toxicol.-Pathol. 2004, 1, 79–95. [Google Scholar] [CrossRef]
- Agency for Toxic Substances and Disease Registry (ATDSR). Toxicological Profile for Aluminum. 2008. Available online: https://www.atsdr.cdc.gov/toxprofiles/tp22.pdf (accessed on 15 February 2022).
- Food and Drug Administration (FDA). Aluminum in Large and Small Volume Parenterals Used in Total Parenteral Nutrition. FDA, Code of Federal Regulations. 2012 [Title 21, 4, Sec. 201.323]. Available online: https://www.accessdata.fda.gov/scripts/cdrh/cfdocs/cfcfr/cfrsearch.cfm?fr=201.323 (accessed on 12 March 2022).
- Yokel, R.A.; Rhineheimer, S.S.; Sharma, P.; Elmore, D.; McNamara, P.J. Entry, Half-Life, and Desferrioxamine-Accelerated Clearance of Brain Aluminum after a Single 26Al Exposure. Toxicol. Sci. 2001, 64, 77–82. [Google Scholar] [CrossRef]
- Goullé, J.P.; Mahieu, L.; Anagnostides, J.G.; Bouige, D.; Saussereau, E.; Guerbet, M.; Lacroix, C. Profil métallique tissulaire par ICP-MS chez des sujets décédés. Ann. Toxicol. Anal. 2010, 22, 1–9. [Google Scholar] [CrossRef]
- Shi, S.; Zhu, H.; Xia, X.; Liang, Z.; Ma, X.; Sun, B. Vaccine adjuvants: Understanding the structure and mechanism of adjuvanticity. Vaccine 2019, 37, 3167–3178. [Google Scholar] [CrossRef]
- Caulfield, M.J.; Shi, L.; Wang, S.; Wang, B.; Tobery, T.W.; Mach, H.; Ahl, P.L.; Cannon, J.L.; Cook, J.C.; Heinrichs, J.H.; et al. Effect of alternative aluminum adjuvants on the absorption and immunogenicity of HPV16 L1 VLPs in mice. Hum. Vacc. 2014, 3, 139–145. [Google Scholar] [CrossRef]
- Hem, S.L.; Hogenesch, H. Relationship between physical and chemical properties of aluminum-containing adjuvants and immunopotentiation. Expert. Rev. Vaccines. 2007, 6, 685–698. [Google Scholar] [CrossRef]
- HogenEsch, H. Mechanisms of stimulation of the immune response by aluminum adjuvants. Vaccine 2002, 20 (Suppl. S3), S34–S39. [Google Scholar] [CrossRef]
- He, P.; Zou, Y.; Hu, Z. Advances in aluminum hydroxide-based adjuvant research and its mechanism. Hum. Vaccin. Immunother. 2015, 11, 477–488. [Google Scholar] [CrossRef]
- Reinke, S.; Thakur, A.; Gartlan, C.; Bezbradica, J.S.; Milicic, A. Inflammasome-Mediated Immunogenicity of Clinical and Experimental Vaccine Adjuvants. Vaccines 2020, 8, 554. [Google Scholar] [CrossRef]
- European Pharmacopeia. Monographs on Allergen Products (1063) and Vaccines for Human Use, (1697), 8th ed; Council of Europe: Strasbourg, France, 2014. [Google Scholar]
- Code of Federal Regulations 21, 7: Sec. 610.15 (Constituent Materials); Revised as of 1 April 2015; US Government Publication; Food and Drug Administration: Silver Spring, MD, USA, 2015.
- Flarend, R.E.; Hem, S.L.; White, J.L.; Elmore, D.; Suckow, M.A.; Rudy, A.C.; Dandashli, E.A. In vivo absorption of aluminium-containing vaccine adjuvants using 26Al. Vaccine 1997, 15, 1314–1318. [Google Scholar] [CrossRef]
- Talbot, H.K.; Rock, M.T.; Johnson, C.; Tussey, L.; Kavita, U.; Shanker, A.; Shaw, A.R.; Taylor, D.N. Immunopotentiation of trivalent influenza vaccine when given with VAX102, a recombinant influenza M2e vaccine fused to the TLR5 ligand flagellin. PLoS ONE 2010, 5, e14442. [Google Scholar] [CrossRef] [PubMed]
- Newton, D.; Talbot, R.J. Long-term retention of injected aluminium-26. Hum. Exp. Toxicol. 2012, 31, 1195–1198. [Google Scholar] [CrossRef] [PubMed]
- Cannata-Andía, J.B.; Fernández-Martín, J.L. The clinical impact of aluminium overload in renal failure. Nephrol. Dial. Transplant. 2002, 17 (Suppl. S2), 9–12. [Google Scholar] [CrossRef][Green Version]
- Malluche, H.H. Aluminium and bone disease in chronic renal failure. Nephrol. Dial. Transplant. 2002, 17 (Suppl. S2), 21–24. [Google Scholar] [CrossRef]
- Balasubramanyam, A.; Sailaja, N.; Mahboob, M.; Rahman, M.F.; Hussain, S.M.; Grover, P. In vitro mutagenicity assessment of aluminium oxide nanomaterials using the Salmonella/microsome assay. Toxicol In Vitro 2010, 24, 1871–1876. [Google Scholar] [CrossRef]
- Balasubramanyam, A.; Sailaja, N.; Mahboob, M.; Rahman, M.F.; Hussain, S.M.; Grover, P. In vivo genotoxicity assessment of aluminium oxide nanomaterials in rat peripheral blood cells using the comet assay and micronucleus test. Mutagenesis 2009, 24, 245–251. [Google Scholar] [CrossRef]
- Jalili, P.; Huet, S.; Burel, A.; Krause, B.C.; Fontana, C.; Chevance, S.; Gauffre, F.; Guichard, Y.; Lampen, A.; Laux, P.; et al. Genotoxic impact of aluminum-containing nanomaterials in human intestinal and hepatic cells. Toxicol. In Vitro 2022, 78, 105257. [Google Scholar] [CrossRef]
- Kim, Y.J.; Choi, H.S.; Song, M.K.; Youk, D.Y.; Kim, J.H.; Ryu, J.C. Genotoxicity of aluminum oxide (Al2O3) nanoparticle in mammalian cell lines. Mol. Cell. Toxicol. 2009, 5, 172–178. [Google Scholar]
- Villarini, M.; Gambelunghe, A.; Giustarini, D.; Ambrosini, M.V.; Fatigoni, C.; Rossi, R.; Dominici, L.; Levorato, S.; Muzi, G.; Piobbico, D.; et al. No evidence of DNA damage by co-exposure to extremely low frequency magnetic fields and aluminum on neuroblastoma cell lines. Mutat. Res. Genet. Toxicol. Environ. Mutagen. 2017, 823, 11–21. [Google Scholar] [CrossRef]
- Banasik, A.; Lankoff, A.; Piskulak, A.; Adamowska, K.; Lisowska, H.; Wojcik, A. Aluminium-induced micronuclei and apoptosis in human peripheral blood lymphocytes treated during different phases of the cell cycle. Environ. Toxicol. 2005, 20, 402–406. [Google Scholar] [CrossRef]
- Lima, P.D.L.; Leite, D.S.; Vasconcellos, M.C.; Cavalcanti, B.C.; Santos, R.A.; Costa-Lotufo, L.V.; Pessoa, C.; Moraes, M.O.; Burbano, R.R. Genotoxic effects of aluminium chloride in cultured human lymphocytes treated in different phases of cell cycle. Food Chem. Toxicol. 2007, 45, 1154–1159. [Google Scholar] [CrossRef] [PubMed]
- Pereira, S.; Cavalie, I.; Camilleri, V.; Gilbin, R.; Adam-Guillermin, C. Comparative genotoxicity of aluminium and cadmium in embryonic zebrafish cells. Mutat Res. 2013, 750, 19–26. [Google Scholar] [CrossRef] [PubMed]
- Türkez, H.; Yousef, M.I.; Geyikoglu, F. Propolis prevents aluminium-induced genetic and hepatic damages in rat liver. Food Chem. Toxicol. 2010, 48, 2741–2746. [Google Scholar] [CrossRef]
- Paz, L.N.F.; Moura, L.M.; Feio, D.C.A.; Cardoso, M.S.G.; Ximenes, W.L.O.; Montenegro, R.C.; Alves, A.P.N.; Burbano, R.R.; Lima, P.D.L. Evaluation of in vivo and in vitro toxicological and genotoxic potential of aluminum chloride. Chemosphere 2017, 175, 130–137. [Google Scholar] [CrossRef]
- Jenkinson, P. Critical review of the publications on the genotoxicology of aluminium salts: 1990–2018. Mutagenesis 2021, 36, 109–127. [Google Scholar] [CrossRef]
- Billiau, A.; Matthys, P. Modes of action of Freund’s adjuvants in experimental models of autoimmune diseases. J. Leukoc. Biol. 2001, 70, 849–860. [Google Scholar]
- Salk, J.E.; Laurent, A.M. The use of adjuvants in studies on influenza immunization. I. Measurements in monkeys of the dimensions of antigenicity of virus-mineral oil emulsions. J. Exp. Med. 1952, 95, 429–447. [Google Scholar] [CrossRef]
- Mussener, A.; Klareskog, L.; Lorentzen, J.C.; Kleinau, S. TNF-α dominates cytokine mRNA expression in lymphoid tissues of rats developing collagen- and oil-induced arthritis. Scand. J. Immunol. 1995, 42, 128–134. [Google Scholar] [CrossRef]
- Stills, H.F., Jr. Adjuvants and antibody production: Dispelling the myths associated with Freund’s complete and other adjuvants. ILAR J. 2005, 46, 280–293. [Google Scholar] [CrossRef]
- Miller, L.H.; Saul, A.; Mahanty, S. Revisiting Freund’s incomplete adjuvant for vaccines in the developing world. Trends Parasitol. 2005, 21, 412–414. [Google Scholar] [CrossRef] [PubMed]
- O’Hagan, D.T.; Ott, G.S.; Van Nest, G.; Rappuoli, R.; Del Giudice, G. The history of MF59 adjuvant: A phoenix that arose from the ashes. Expert Rev. Vaccines 2013, 12, 13–30. [Google Scholar] [CrossRef] [PubMed]
- Kommareddy, S.; Singh, M.; O’Hagan, D.T. Chapter 13—MF59: A safe and potent adjuvant for human use. In Immunopotentiators in Modern Vaccines, 2nd ed.; Schijns, V.E.J.C., O’Hagan, D.T., Eds.; Academic Press: Cambridge, MA, USA, 2017; pp. 249–263. [Google Scholar]
- O’Hagan, D.T.; Wack, A.; Podda, A. MF59 is a safe and potent vaccine adjuvant for flu vaccines in humans: What did we learn during its development? Clin. Pharmacol. Ther. 2007, 82, 740–744. [Google Scholar] [CrossRef]
- Vesikari, T.; Pellegrini, M.; Karvonen, A.; Growth, N.; Borkowski, A.; O’Hagan, D.T.; Podda, A. Enhanced immunogenicity of seasonal influenza vaccines in young children using MF59 adjuvant. Pediatr. Infect. Dis. J. 2009, 28, 563–571. [Google Scholar] [CrossRef]
- Vesikari, T.; Knuf, M.; Wutzler, P.; Karvonen, A.; Kieninger-Baum, D.; Schmitt, H.J.; Baehner, F.; Borkowski, A.; Tsai, T.F.; Clemens, R. Oil-in-water emulsion adjuvant with influenza vaccine in young children. N. Engl. J. Med. 2011, 365, 1406–1416. [Google Scholar] [CrossRef]
- Nakaya, H.I.; Clutterbuck, E.; Kazmin, D.; Wang, L.; Cortese, M.; Bosinger, S.E.; Patel, N.B.; Zak, D.E.; Aderem, A.; Dong, T.; et al. Systems biology of immunity to MF59-adjuvanted versus nonadjuvanted trivalent seasonal influenza vaccines in early childhood. Proc. Natl. Acad. Sci. USA 2016, 113, 1853–1858. [Google Scholar] [CrossRef]
- Heineman, T.C.; Clements-Mann, M.L.; Poland, G.A.; Jacobson, R.M.; Izu, A.E.; Sakamoto, D.; Eiden, J.; Van Nest, G.A.; Hsu, H.H. A randomized, controlled study in adults of the immunogenicity of a novel hepatitis B vaccine containing MF59 adjuvant. Vaccine 1999, 17, 2769–2778. [Google Scholar] [CrossRef]
- Dupuis, M.; McDonald, D.M.; Ott, G. Distribution of adjuvant MF59 and antigen gD2 after intramuscular injection in mice. Vaccine 1999, 18, 434–439. [Google Scholar] [CrossRef]
- Stephenson, I.; Bugarini, R.; Nicholson, K.G.; Podda, A.; Wood, J.M.; Zambon, M.C.; Katz, J.M. Cross-reactivity to highly pathogenic avian influenza H5N1 viruses after vaccination with nonadjuvanted and MF59-adjuvanted influenza A/Duck/Singapore/97 (H5N3) vaccine: A potential priming strategy. J. Infect. Dis. 2005, 191, 1210–1215. [Google Scholar] [CrossRef]
- Seubert, A.; Monaci, E.; Pizza, M.; O’Hagan, D.T.; Wack, A. The adjuvants aluminum hydroxide and MF59 induce monocyte and granulocyte chemoattractants and enhance monocyte differentiation toward dendritic cells. J. Immunol. 2008, 180, 5402–5412. [Google Scholar] [CrossRef]
- De Gregorio, E.; Caproni, E.; Ulmer, J.B. Vaccine adjuvants: Mode of action. Front. Immunol. 2013, 4, 214. [Google Scholar] [CrossRef] [PubMed]
- Mosca, F.; Tritto, E.; Muzzi, A.; Monaci, E.; Bagnoli, F.; Iavarone, C.; O’Hagan, D.; Rappuoli, R.; De Gregorio, E. Molecular and cellular signatures of human vaccine adjuvants. Proc. Natl. Acad. Sci. USA 2008, 105, 10501–10506. [Google Scholar] [CrossRef] [PubMed]
- Schultze, V.; D’Agosto, V.; Wack, A.; Novicki, D.; Zorn, J.; Hennig, R. Safety of MF59 adjuvant. Vaccine 2008, 26, 3209–3222. [Google Scholar] [CrossRef] [PubMed]
- Garcon, N.; Vaughn, D.W.; Didierlaurent, A.M. Development and evaluation of AS03, an adjuvant System containing a-tocopherol and squalene in an oil-in-water emulsion. Expert Rev. Vaccines 2012, 11, 349–366. [Google Scholar] [CrossRef]
- Moris, P.; van der Most, R.; Leroux-Roels, I.; Clement, F.; Dramé, M.; Hanon, E.; Leroux-Roels, G.G.; Van Mechelen, M. H5N1 influenza vaccine formulated with AS03 A induces strong cross-reactive and polyfunctional CD4 T-cell responses. J. Clin. Immunol. 2011, 31, 443–454. [Google Scholar] [CrossRef]
- Wu, Y.; Narum, D.L.; Fleury, S.; Jennings, G.; Yadava, A. Particle-based platforms for malaria vaccines. Vaccine 2015, 33, 7518–7524. [Google Scholar] [CrossRef]
- European Medicines Agency (EMA). Pandemrix-Adjuvanted H1N1 Inluenza Vaccine; 2017. Available online: https://www.ema.europa.eu/en/documents/other/pandemrix-package-leaflet_en.pdf (accessed on 30 March 2022).
- GlaxoSmithKline. Influenza A (H5N1) Virus Monovalent Vaccine, Adjuvanted; 2018. Available online: https://gskpro.com/content/dam/global/hcpportal/en_US/Prescribing_Information/Influenza/pdf/INFLUENZA-A-(H5N1)-VIRUS-MONOVALENT-VACCINE,-ADJUVANTED.PDF (accessed on 30 March 2022).
- Díez-Domingo, J.; Garces-Sanchez, M.; Baldò, J.M.; Planelles, M.V.; Ubeda, I.; JuBert, A.; Marés, J.; Moris, P.; Garcia-Corbeira, P.; Dramé, M.; et al. Immunogenicity and safety of H5N1 A/vietnam/1194/2004 (clade 1) AS03-adjuvanted prepandemic candidate influenza vaccines in children aged 3 to 9 years: A phase II, randomized, open, controlled study. Pediatric Infect. Dis. J. 2010, 29, e35–e46. [Google Scholar] [CrossRef]
- Morel, S.; Didierlaurent, A.; Bourguignon, P.; Delhaye, S.; Baras, B.; Jacob, V.; Planty, C.; Elouahabi, A.; Harvengt, P.; Carlsen, H.; et al. Adjuvant System AS03 containing α-tocopherol modulates innate immune response and leads to improved adaptive immunity. Vaccine 2011, 29, 2461–2473. [Google Scholar] [CrossRef]
- Martinez, A.C.; De La Cueva, I.S.; Boutet, P.; Abeele, C.V.; Smolenov, I.; Devaster, J.M. A phase 1, open-label safety and immunogenicity study of an AS03-adjuvanted trivalent inactivated influenza vaccine in children aged 6 to 35 months. Hum. Vaccines Immunother. 2014, 10, 1959–1968. [Google Scholar] [CrossRef][Green Version]
- Garcon, N.; Chomez, P.; Van Mechelen, M. GlaxoSmithKline Adjuvant Systems in vaccines: Concepts, achievements and perspectives. Expert Rev. Vaccines 2007, 6, 723. [Google Scholar] [CrossRef]
- Garcon, N.; Van Mechelen, M. Recent clinical experience with vaccines using MPL and QS-21-containing adjuvant systems. Expert Rev. Vaccines 2011, 10, 471–486. [Google Scholar] [CrossRef] [PubMed]
- Grgacic, E.V.; Anderson, D.A. Virus-like particles: Passport to immune recognition. Methods 2006, 40, 60–65. [Google Scholar] [CrossRef] [PubMed]
- Kushnir, N.; Streatfield, S.J.; Yusibov, V. Virus-like particles as a highly efficient vaccine platform: Diversity of targets and production systems and advances in clinical development. Vaccine 2012, 31, 58–83. [Google Scholar] [CrossRef] [PubMed]
- Nooraei, S.; Bahrulolum, H.; Hoseini, Z.S.; Katalani, C.; Hajizade, A.; Easton, A.J.; Ahmadian, G. Virus-like particles: Preparation, immunogenicity and their roles as nanovaccines and drug nanocarriers. J. Nanobiotechnol. 2021, 19, 59. [Google Scholar] [CrossRef] [PubMed]
- Facciolà, A.; Visalli, G.; Laganà, P.; La Fauci, V.; Squeri, R.; Pellicanò, G.F.; Nunnari, G.; Trovato, M.; Di Pietro, A. The new era of vaccines: The “nanovaccinology”. Eur. Rev. Med. Pharmacol. Sci. 2019, 23, 7163–7182. [Google Scholar] [PubMed]
- Bachmann, M.F.; Jennings, G.T. Vaccine delivery: A matter of size, geometry, kinetics and molecular patterns. Immunology 2010, 10, 787–796. [Google Scholar] [CrossRef] [PubMed]
- Jeong, H.; Seong, B.L. Exploiting virus-like particles as innovative vaccines against emerging viral infections. J. Microbiol. 2017, 55, 220–230. [Google Scholar] [CrossRef]
- Pattenden, L.K.; Middelberg, A.P.J.; Niebert, M.; Lipin, D.I. Towards the preparative and large-scale precision manufacture of virus-like particles. Trends Biotechnol. 2005, 23, 523–529. [Google Scholar] [CrossRef]
- Zeltins, A. Construction and characterization of virus-like particles: A review. Mol. Biotechnol. 2013, 53, 92–107. [Google Scholar] [CrossRef]
- Sánchez-Rodríguez, S.P.; Münch-Anguiano, L.; Echeverría, O.; Vázquez-Nin, G.; Mora-Pale, M.; Dordick, J.S.; Bustos-Jaimes, I. Human parvovirus B19 virus-like particles: In vitro assembly and stability. Biochimie 2012, 94, 870–878. [Google Scholar] [CrossRef]
- Liew, M.W.O.; Chuan, Y.P.; Middelberg, A.P.J. High-yield and scalable cell-free assembly of virus-like particles by dilution. Biochem. Eng. J. 2012, 67, 88–96. [Google Scholar] [CrossRef]
- Lu, Y.; Welsh, J.P.; Chan, W.; Swartz, J.R. Escherichia coli-based cell free production of flagellin and ordered flagellin display on virus-like particles. Biotechnol. Bioeng. 2013, 110, 2073–2085. [Google Scholar] [CrossRef] [PubMed]
- Chuan, Y.P.; Lua, L.H.L.; Middelberg, A.P.J. Virus-Like Particle Bioprocessing; Biopharmaceutical Production Technology; Wiley-VCH Verlag GmbH & Co., KGaA: Weinheim, Germany, 2012; pp. 139–163. [Google Scholar]
- Lua, L.H.; Connors, N.K.; Sainsbury, F.; Chuan, Y.P.; Wibowo, N.; Middelberg, A.P. Bioengineering virus-like particles as vaccines. Biotechnol. Bioeng. 2014, 111, 425–440. [Google Scholar] [CrossRef] [PubMed]
- Stephenne, J. Production in yeast versus mammalian cells of the first efficacy, and economy: Hepatitis B vaccine. Adv. Biotechnol. Processes 1990, 14, 279–299. [Google Scholar]
- Keating, G.M.; Noble, S. Recombinant Hepatitis B Vaccine (Engerix-B®). Drugs 2003, 63, 1021–1051. [Google Scholar] [CrossRef]
- La Fauci, V.; Riso, R.; Facciolà, A.; Ceccio, C.; Lo Giudice, D.; Calimeri, S.; Squeri, R. Response to anti-HBV vaccine and 10-year follow-up of antibody levels in healthcare workers. Public Health 2016, 139, 198–202. [Google Scholar] [CrossRef] [PubMed]
- Szymonowicz, K.A.; Chen, J. Biological and clinical aspects of HPV-related cancers. Cancer Biol. Med. 2020, 17, 864–878. [Google Scholar] [CrossRef]
- Visalli, G.; Currò, M.; Facciolà, A.; Riso, R.; Mondello, P.; Laganà, P.; Di Pietro, A.; Picerno, I.; Spataro, P. Prevalence of human papillomavirus in saliva of women with HPV genital lesions. Infect. Agent Cancer 2016, 11, 48. [Google Scholar] [CrossRef]
- Zhai, L.; Tumban, E. Gardasil-9: A Global Survey of Projected Efficacy. Antiviral. Res. 2016, 130, 101–109. [Google Scholar] [CrossRef]
- European Medicines Agency (EMA). Gardasil 9. Summary of Product Characteristic. Available online: https://www.ema.europa.eu/en/documents/product-information/gardasil-9-epar-product-information_en.pdf (accessed on 18 February 2022).
- Cheng, L.; Wang, Y.; Du, J. Human Papillomavirus Vaccines: An Updated Review. Vaccines 2020, 8, 391. [Google Scholar] [CrossRef]
- Wilschut, J. Influenza vaccines: The virosome concept. Immunol. Lett. 2009, 122, 118–121. [Google Scholar] [CrossRef] [PubMed]
- Almeida, J.; Edwards, D.C.; Brand, C.; Heath, T. Formation of Virosomes from Influenza Subunits and Liposomes. Lancet 1975, 306, 899. [Google Scholar] [CrossRef]
- Bovier, P.A. Epaxal: A virosomal vaccine to prevent hepatitis A infection. Expert Rev. Vaccines 2008, 7, 1141–1150. [Google Scholar] [CrossRef] [PubMed]
- Herzog, C.; Hartmann, K.; Künzi, V.; Kürsteiner, O.; Mischler, R.; Lazar, H.; Glück, R. Eleven years of Inflexal V-a virosomal adjuvanted influenza vaccine. Vaccine 2009, 27, 4381–4387. [Google Scholar] [CrossRef]
- Soema, P.C.; Kompier, R.; Amorij, J.P.; Kersten, G.F. Current and Next Generation Influenza Vaccines: Formulation and Production Strategies. Eur. J. Pharm. Biopharm. 2015, 94, 251. [Google Scholar] [CrossRef]
- Huckriede, A.; Bungener, L.; Stegmann, T.; Daemen, T.; Medema, J.; Palache, A.M.; Wilschut, J. The Virosome Concept for Influenza Vaccines. Vaccine 2005, 23, S26–S38. [Google Scholar] [CrossRef]
- Dong, W.; Bhide, Y.; Marsman, S.; Holtrop, M.; Meijerhof, T.; de Vries–Idema, J.; de Haan, A.; Huckriede, A. Monophosphoryl Lipid a-Adjuvanted Virosomes with Ni-Chelating Lipids for Attachment of Conserved Viral Proteins as Cross-Protective Influenza Vaccine. Biotechnol. J. 2018, 13, e1700645. [Google Scholar] [CrossRef]
- Bron, R.; Ortiz, A.; Wilschut, J. Cellular cytoplasmic delivery of a polypeptide toxin by reconstituted influenza virus envelopes (virosomes). Biochemistry 1994, 9, 9110–9117. [Google Scholar] [CrossRef]
- Qian, C.; Liu, X.; Xu, Q.; Wang, Z.; Chen, J.; Li, T.; Zheng, Q.; Yu, H.; Gu, Y.; Li, S.; et al. Recent progress on the versatility of virus-like particles. Vaccines 2020, 8, 139. [Google Scholar] [CrossRef]
- Criscuolo, E.; Caputo, V.; Diotti, R.A.; Sautto, G.A.; Kirchenbaum, G.A.; Clementi, N. Alternative methods of vaccine delivery: An overview of edible and intradermal vaccines. J. Immunol. Res. 2019, 2019, 8303648. [Google Scholar] [CrossRef]
- Emadi, F.; Emadi, A.; Gholami, A. A comprehensive insight towards pharmaceutical aspects of graphene nanosheets. Curr. Pharm. Biotechnol. 2020, 21, 1016–1027. [Google Scholar] [CrossRef] [PubMed]
- Borzouyan Dastjerdi, M.; Amini, A.; Nazari, M.; Cheng, C.; Benson, V.; Gholami, A.; Ghasemi, Y. Novel versatile 3D bio-scaffold made of natural biocompatible hagfish exudate for tissue growth and organoid modeling. Int. J. Biol. Macromol. 2020, 158, 894–902. [Google Scholar] [CrossRef] [PubMed]
- Amacker, M.; Smardon, C.; Mason, L.; Sorrell, J.; Jeffery, K.; Adler, M.; Bhoelan, F.; Belova, O.; Spengler, M.; Punnamoottil, B.; et al. New GMP manufacturing processes to obtain thermostable HIV-1 gp41 virosomes under solid forms for various mucosal vaccination routes. Npj Vaccines 2020, 5, 41. [Google Scholar] [CrossRef] [PubMed]
- Bungener, L.; de Mare, A.; de Vries-Idema, J.; Sehr, P.; van der Zee, A.; Wilschut, J.; Daemen, T. A virosomal immunization strategy against cervical cancer and premalignant cervical disease. Antivir. Ther. 2006, 6, 717–727. [Google Scholar] [CrossRef]
- Lederhofer, J.; van Lent, J.; Bhoelan, F.; Karneva, Z.; de Haan, A.; Wilschut, J.C.; Stegmann, T. Development of a Virosomal RSV Vaccine Containing 3D-PHAD® Adjuvant: Formulation, Composition, and Long-Term Stability. Pharm. Res. 2018, 35, 172. [Google Scholar] [CrossRef]
- Pluschke, G.; Tamborrini, M. Development of a virosomal malaria vaccine candidate: From synthetic peptide design to clinical concept validation. Futur. Virol. 2012, 7, 779–790. [Google Scholar] [CrossRef]
- Ebensen, T.; Debarry, J.; Pedersen, G.K.; Blazejewska, P.; Weissmann, S.; Schulze, K.; McCullough, K.C.; Cox, R.J.; Guzmán, C.A. Mucosal Administration of Cycle-Di-Nucleotide-Adjuvanted Virosomes Efficiently Induces Protection against Influenza H5N1 in Mice. Front. Immunol. 2017, 8, 1223. [Google Scholar] [CrossRef]
- Duchemin, M.; Tudor, D.; Cottignies-Calamarte, A.; Bomsel, M. Antibody-dependent cellular phagocytosis of HIV-1-infected cells is efficiently triggered by IgA targeting HIV-1 envelope subunit gp41. Front. Immunol. 2020, 11, 1141. [Google Scholar] [CrossRef]
- Wong, Y.T.; Smardon, C.; Shirkhani, K.; Amacker, M.; Fleury, S.; Stegmann, A.J.H. Oral Dispersible Vaccine Comprising Virosomes. International Application No. PCT/EP2019/082940, 28 June 2019. Available online: https://patentscope.wipo.int/search/en/detail.jsf?docId=WO2020109485 (accessed on 18 March 2022).
- Lee, B.R.; Jeong, S.K.; Ahn, B.C.; Lee, B.-J.; Shin, S.J.; Yum, J.S.; Ha, S.J. Combination of TLR1/2 and TLR3 ligands enhances CD4+ T cell longevity and antibody responses by modulating type I IFN production. Sci. Rep. 2016, 6, 32526. [Google Scholar] [CrossRef]
- Jeong, S.K.; Heo, Y.K.; Jeong, J.H.; Ham, S.J.; Yum, J.S.; Ahn, B.C.; Song, C.S.; Chun, E.Y. COVID-19 Subunit Vaccine with a Combination of TLR1/2 and TLR3 Agonists Induces Robust and Protective Immunity. Vaccines 2021, 9, 957. [Google Scholar] [CrossRef]
- Hamley, I.W. Lipopeptides for Vaccine Development. Bioconjug Chem. 2021, 32, 1472–1490. [Google Scholar] [CrossRef] [PubMed]
- Takeuchi, O.; Kaufmann, A.; Grote, K.; Kawai, T.; Hoshino, K.; Morr, M.; Mühlradt, P.F.; Akira, S. Cutting Edge: Preferentially the RStereoisomer of the Mycoplasmal Lipopeptide Macrophage-Activating Lipopeptide-2 Activates Immune Cells through a Toll-like Receptor 2- and MyD88-Dependent Signaling Pathway. J. Immunol. 2000, 164, 554–557. [Google Scholar] [CrossRef] [PubMed]
- Halliday, A.; Turner, J.D.; Guimarães, A.; Bates, P.A.; Taylor, M.J. The TLR2/6 Ligand PAM2CSK4 Is a Th2 Polarizing Adjuvant in Leishmania Major and Brugia Malayi Murine Vaccine Models. Parasites Vectors 2016, 9, 96. [Google Scholar] [CrossRef] [PubMed]
- Powell, T.J. Microparticulate Vaccine against Malaria. International Application No. PCT/US2013/033070, 20 March 2013. [Google Scholar]
- Caproni, E.; Tritto, E.; Cortese, M.; Muzzi, A.; Mosca, F.; Monaci, E.; Baudner, B.; Seubert, A.; De Gregorio, E. MF59 and Pam3CSK4 Boost Adaptive Responses to Influenza Subunit Vaccine through an IFN Type I-Independent Mechanism of Action. J. Immunol. 2012, 188, 3088–3098. [Google Scholar] [CrossRef]
- Field, A.K.; Tytell, A.A.; Lampson, G.P.; Hilleman, M.R. Inducers of Interferon and Host Resistance. II. Multistranded Synthetic Polynucleotide Complexes. Proc. Natl. Acad. Sci. USA 1967, 58, 1004–1010. [Google Scholar] [CrossRef]
- Okahira, S.; Nishikawa, F.; Nishikawa, S.; Akazawa, T.; Seya, T.; Matsumoto, M. Interferon-Beta Induction Through Toll-Like Receptor 3 Depends on Double-Stranded RNA Structure. DNA Cell. Biol. 2005, 24, 614–623. [Google Scholar] [CrossRef]
- Kato, H.; Takeuchi, O.; Sato, S.; Yoneyama, M.; Yamamoto, M.; Matsui, K.; Uematsu, S.; Jung, A.; Kawai, T.; Ishii, K.J.; et al. Differential Roles of MDA5 and RIG-I Helicases in the Recognition of RNA Viruses. Nature 2006, 441, 101–105. [Google Scholar] [CrossRef]
- Longhi, M.P.; Trumpfheller, C.; Idoyaga, J.; Caskey, M.; Matos, I.; Kluger, C.; Salazar, A.M.; Colonna, M.; Steinman, R.M. Dendritic Cells Require a Systemic Type I Interferon Response to Mature and Induce CD4+ Th1 Immunity with Poly IC as Adjuvant. J. Exp. Med. 2009, 206, 1589–1602. [Google Scholar] [CrossRef]
- Schulz, O.; Diebold, S.S.; Chen, M.; Naslund, T.I.; Nolte, M.A.; Alexopoulou, L.; Azuma, Y.T.; Flavell, R.A.; Liljeström, P.; Reis e Sousa, C. Toll-Like Receptor 3 Promotes Cross-Priming to Virus-Infected Cells. Nature 2005, 433, 887–892. [Google Scholar] [CrossRef]
- Durand, V.; Wong, S.Y.; Tough, D.F.; Le Bon, A. IFN-Alpha/BetaDependent Cross-Priming Induced by Specific Toll-Like Receptor Agonists. Vaccine 2006, 24, S22–S23. [Google Scholar] [CrossRef]
- Kolumam, G.A.; Thomas, S.; Thompson, L.J.; Sprent, J.; Murali-Krishna, K. Type I Interferons Act Directly on CD8 T Cells to Allow Clonal Expansion and Memory Formation in Response to Viral Infection. J. Exp. Med. 2005, 202, 637–650. [Google Scholar] [CrossRef] [PubMed]
- Ngoi, S.M.; Tovey, M.G.; Vella, A.T. Targeting Poly(I:C) to the TLR3-Independent Pathway Boosts Effector CD8 T Cell Differentiation Through IFN-Alpha/Beta. J. Immunol. 2008, 181, 7670–7680. [Google Scholar] [CrossRef] [PubMed]
- Perret, R.; Sierro, S.R.; Botelho, N.K.; Corgnac, S.; Donda, A.; Romero, P. Adjuvants That Improve the Ratio of Antigen-Specific Effector to Regulatory T Cells Enhance Tumor Immunity. Cancer. Res. 2013, 73, 6597–6608. [Google Scholar] [CrossRef] [PubMed]
- Cornell, C.J., Jr.; Smith, K.A.; Cornwell, G.G., 3rd; Burke, G.P.; McIntyre, O.R. Systemic Effects of Intravenous Polyriboinosinic-Polyribocytidylic Acid in Man. J. Natl. Cancer Inst. 1976, 57, 1211–1216. [Google Scholar] [CrossRef] [PubMed]
- Robinson, R.A.; DeVita, V.T.; Levy, H.B.; Baron, S.; Hubbard, S.P.; Levine, A.S. A Phase I-II Trial of Multiple-Dose Polyriboinosic-Polyribocytidylic Acid in Patieonts with Leukemia or Solid Tumors. J. Natl. Cancer Inst. 1976, 57, 599–602. [Google Scholar] [CrossRef] [PubMed]
- Levy, H.B.; Baer, G.; Baron, S.; Buckler, C.E.; Gibbs, C.J.; Iadarola, M.J.; London, W.T.; Rice, J. A Modified Polyriboinosinic-Polyribocytidylic Acid Complex That Induces Interferon in Primates. J. Infect. Dis. 1975, 132, 434–439. [Google Scholar] [CrossRef] [PubMed]
- Caskey, M.; Lefebvre, F.; Filali-Mouhim, A.; Cameron, M.J.; Goulet, J.P.; Haddad, E.K.; Breton, G.; Trumpfheller, C.; Pollak, S.; Shimeliovich, I.; et al. Synthetic Double-Stranded RNA Induces Innate Immune Responses Similar to a Live Viral Vaccine in Humans. J. Exp. Med. 2011, 208, 2357–2366. [Google Scholar] [CrossRef]
- Kastenmuller, K.; Espinosa, D.A.; Trager, L.; Stoyanov, C.; Salazar, A.M.; Pokalwar, S.; Singh, S.; Dutta, S.; Ockenhouse, C.F.; Zavala, F.; et al. Full-Length Plasmodium Falciparum Circumsporozoite Protein Administered With Long-Chain Poly(I.C) or the Toll-Like Receptor 4 Agonist Glucopyranosyl Lipid Adjuvant-Stable Emulsion Elicits Potent Antibody and CD4+ T Cell Immunity and Protection in Mice. Infect. Immun. 2013, 81, 789–800. [Google Scholar] [CrossRef]
- Flynn, B.J.; Kastenmuller, K.; Wille-Reece, U.; Tomaras, G.D.; Alam, M.; Lindsay, R.W.; Salazar, A.M.; Perdiguero, B.; Gomez, C.E.; Wagner, R.; et al. Immunization with HIV Gag Targeted to Dendritic Cells Followed by Recombinant New York Vaccinia Virus Induces Robust T-Cell Immunity in Nonhuman Primates. Proc. Natl. Acad. Sci. USA 2011, 108, 7131–7136. [Google Scholar] [CrossRef]
- Ohlfest, J.R.; Andersen, B.M.; Litterman, A.J.; Xia, J.; Pennell, C.A.; Swier, L.E.; Salazar, M.A.; Olin, R. Vaccine Injection Site Matters: Qualitative and Quantitative Defects in CD8 T Cells Primed as a Function of Proximity to the Tumor in a Murine Glioma Model. J. Immunol. 2013, 190, 613–620. [Google Scholar] [CrossRef]
- Sabbatini, P.; Tsuji, T.; Ferran, L.; Ritter, E.; Sedrak, C.; Tuballes, K.; Jungbluth, A.A.; Ritter, G.; Aghajanian, C.; Bell-McGuinn, K.; et al. Phase I Trial of Overlapping Long Peptides from a Tumor Self-Antigen and Poly-ICLC Shows Rapid Induction of Integrated Immune Response in Ovarian Cancer Patients. Clin. Cancer Res. 2012, 18, 6497–6508. [Google Scholar] [CrossRef] [PubMed]
- Engel, A.L.; Holt, G.E.; Lu, H. The Pharmacokinetics of Toll-Like Receptor Agonists and the Impact on the Immune System. Expert. Rev. Clin. Pharmacol. 2011, 4, 275–289. [Google Scholar] [CrossRef] [PubMed]
- Martins, K.A.; Bavari, S.; Salazar, A.M. Vaccine Adjuvant Uses of Poly-IC and Derivatives. Expert Rev Vaccines 2015, 14, 447–459. [Google Scholar] [CrossRef] [PubMed]
- Gowen, B.B.; Wong, M.H.; Jung, K.H.; Sanders, A.B.; Mitchell, W.M.; Alexopoulou, L.; Flavell, R.A.; Sidwell, R.W. TLR3 is Essential for the Induction of Protective Immunity Against Punta Toro Virus Infection by the DoubleStranded RNA (dsRNA), Poly(I:C12U), But Not Poly(I:C): Differential Recognition of Synthetic dsRNA Molecules. J. Immunol. 2007, 178, 5200–5208. [Google Scholar] [CrossRef] [PubMed]
- Flamar, A.L.; Contreras, V.; Zurawski, S.; Montes, M.; Dereuddre-Bosquet, N.; Martinon, F.; Banchereau, J.; Le Grand, R.; Zurawski, G.; Levy, Y. Delivering HIV Gagp24 to DCIR Induces Strong Antibody Responses In Vivo. PLoS ONE 2015, 10, e0135513. [Google Scholar] [CrossRef] [PubMed]
- Overton, E.T.; Goepfert, P.A.; Cunningham, P.; Carter, W.A.; Horvath, J.; Young, D.; Strayer, D.R. Intranasal Seasonal Influenza Vaccine and a TLR-3 Agonist, Rintatolimod, Induced Cross-Reactive IgA Antibody Formation Against Avian H5N1 and H7N9 Influenza HA in Humans. Vaccine 2014, 32, 5490–5495. [Google Scholar] [CrossRef]
- Navabi, H.; Jasani, B.; Reece, A.; Clayton, A.; Tabi, Z.; Donninger, C.; Mason, M.; Adams, M. A Clinical Grade Poly I:C-Analogue (Ampligen) Promotes Optimal DC Maturation and Th1-Type T Cell Responses of Healthy Donors and Cancer Patients In Vitro. Vaccine 2009, 27, 107–115. [Google Scholar] [CrossRef]
- Seya, T.; Takeda, Y.; Matsumoto, M. Tumor Vaccines with dsRNA Adjuvant ARNAX Induces Antigen-Specific Tumor Shrinkage Without Cytokinemia. Oncoimmunology 2016, 5, e1043506. [Google Scholar] [CrossRef]
- Matsumoto, M.; Tatematsu, M.; Nishikawa, F.; Azuma, M.; Ishii, N.; Morii-Sakai, A.; Shime, H.; Seya, T. Defined TLR3-Specific Adjuvant That Induces NK and CTL Activation Without Significant Cytokine Production In Vivo. Nat. Commun. 2015, 6, 6280. [Google Scholar] [CrossRef]
- Takeda, Y.; Kataoka, K.; Yamagishi, J.; Ogawa, S.; Seya, T.; Matsumoto, M. A TLR3-Specific Adjuvant Relieves Innate Resistance to PD-L1 Blockade Without Cytokine Toxicity in Tumor Vaccine Immunotherapy. Cell. Rep. 2017, 19, 1874–1887. [Google Scholar] [CrossRef]
- Matsumoto, M.; Takeda, Y.; Seya, T. Targeting Toll-Like Receptor 3 in Dendritic Cells for Cancer Immunotherapy. Expert Opin. Biol. Ther. 2020, 20, 937–946. [Google Scholar] [CrossRef] [PubMed]
- Takeda, Y.; Takaki, H.; Fukui-Miyazaki, A.; Yoshida, S.; Matsumoto, M.; Seya, T. Vaccine Adjuvant ARNAX Promotes Mucosal IgA Production in Influenza HA Vaccination. Biochem. Biophys. Res. Commun. 2018, 506, 1019–1025. [Google Scholar] [CrossRef] [PubMed]
- White, M.T.; Verity, R.; Griffin, J.T.; Asante, K.P.; Owusu-Agyei, S.; Greenwood, B.; Drakeley, C.; Gesase, S.; Lusingu, J.; Ansong, D.; et al. Immunogenicity of the RTS,S/AS01 Malaria Vaccine and Implications for Duration of Vaccine Efficacy: Secondary Analysis of Data From a Phase 3 Randomised Controlled Trial. Lancet Infect. Dis. 2015, 15, 1450–1458. [Google Scholar] [CrossRef]
- Leroux-Roels, I.; Koutsoukos, M.; Clement, F.; Steyaert, S.; Janssens, M.; Bourguignon, P.; Cohen, K.; Altfeld, M.; Vandepapelière, P.; Pedneault, L.; et al. Strong and Persistent CD4+ T-Cell Response in Healthy Adults Immunized with a Candidate HIV-1 Vaccine Containing Gp120, Nef and Tat Antigens Formulated in Three Adjuvant Systems. Vaccine 2010, 28, 7016–7024. [Google Scholar] [CrossRef]
- Van Braeckel, E.; Bourguignon, P.; Koutsoukos, M.; Clement, F.; Janssens, M.; Carletti, I.; Collard, A.; Demoitié, M.A.; Voss, G.; Leroux-Roels, G.; et al. An Adjuvanted Polyprotein HIV-1 Vaccine Induces Polyfunctional Cross-Reactive CD4+ T Cell Responses in Seronegative Volunteers. Clin. Infect. Dis. 2011, 52, 522–531. [Google Scholar] [CrossRef]
- Leroux-Roels, I.; Forgus, S.; De Boever, F.; Clement, F.; Demoitié, M.A.; Mettens, P.; Moris, P.; Ledent, E.; Leroux-Roels, G.; Ofori-Anyinam, O.; et al. Improved CD4+ T Cell Responses to Mycobacterium Tuberculosis in PPD-Negative Adults by M72/AS01 as Compared to the M72/AS02 and Mtb72F/AS02 Tuberculosis Candidate Vaccine Formulations: A Randomized Trial. Vaccine 2013, 31, 2196–2206. [Google Scholar] [CrossRef]
- Kensil, C.R.; Patel, U.; Lennick, M.; Marciani, D. Separation and Characterization of Saponins with Adjuvant Activity from Quillaja Saponaria Molina Cortex. J. Immunol. 1991, 146, 431–437. [Google Scholar]
- Welsby, I.; Detienne, S.; N’Kuli, F.; Thomas, S.; Wouters, S.; Bechtold, V.; De Wit, D.; Gineste, R.; Reinheckel, T.; Elouahabi, A.; et al. Lysosome-Dependent Activation of Human Dendritic Cells by the Vaccine Adjuvant QS-21. Front. Immunol. 2017, 7, 663. [Google Scholar] [CrossRef]
- Watts, B.A.; George, T.; Sherwood, E.R.; Good, D.W. Monophosphoryl Lipid A Induces Protection Against LPS in Medullary Thick Ascending Limb Through a TLR4-TRIF-PI3K Signaling Pathway. Am. J. Physiol. Renal. Physiol. 2017, 313, F103–F115. [Google Scholar] [CrossRef]
- Beck, Z.; Matyas, G.R.; Alving, C.R. Detection of Liposomal Cholesterol and Monophosphoryl Lipid A by QS-21 Saponin and Limulus Polyphemus Amebocyte Lysate. Biochim Biophys Acta BBA Biomembr. 2015, 1848, 775–780. [Google Scholar] [CrossRef]
- Marty-Roix, R.; Vladimer, G.I.; Pouliot, K.; Weng, D.; Buglione-Corbett, R.; West, K.; MacMicking, J.D.; Chee, J.D.; Wang, S.; Lu, S.; et al. Identification of QS-21 as an Inflammasome-Activating Molecular Component of Saponin Adjuvants. J. Biol. Chem. 2016, 291, 1123–1136. [Google Scholar] [CrossRef] [PubMed]
- Detienne, S.; Welsby, I.; Collignon, C.; Wouters, S.; Coccia, M.; Delhaye, S.; Van Maele, L.; Thomas, S.; Swertvaegher, M.; Detavernier, A.; et al. Central Role of CD169+ Lymph Node Resident Macrophages in the Adjuvanticity of the QS-21 Component of AS01. Sci. Rep. 2016, 6, 39475. [Google Scholar] [CrossRef] [PubMed]
- Coccia, M.; Collignon, C.; Hervé, C.; Chalon, A.; Welsby, I.; Detienne, S.; van Helden, M.J.; Dutta, S.; Genito, C.J.; Waters, N.C.; et al. Cellular and Molecular Synergy in AS01-Adjuvanted Vaccines Results in an Early IFNγ Response Promoting Vaccine Immunogenicity. NPJ Vaccines 2017, 2, 25. [Google Scholar] [CrossRef] [PubMed]
- Hayashi, F.; Smith, K.D.; Ozinsky, A.; Hawn, T.R.; Yi, E.C.; Goodlett, D.R.; Eng, J.K.; Akira, S.; Underhill, D.M.; Aderem, A. The Innate Immune Response to Bacterial Flagellin is Mediated by Toll-Like Receptor 5. Nature 2001, 410, 1099–1103. [Google Scholar] [CrossRef]
- Huleatt, J.W.; Jacobs, A.R.; Tang, J.; Desai, P.; Kopp, E.B.; Huang, Y.; Song, L.; Nakaar, V.; Powell, T.J. Vaccination with Recombinant Fusion Proteins Incorporating Toll-Like Receptor Ligands Induces Rapid Cellular and Humoral Immunity. Vaccine 2007, 25, 763–775. [Google Scholar] [CrossRef]
- Franchi, L.; Amer, A.; Body-Malapel, M.; Kanneganti, T.D.; Ozoren, N.; Jagirdar, R.; Inohara, N.; Vandenabeele, P.; Bertin, J.; Coyle, A.; et al. Cytosolic Flagellin Requires Ipaf for Activation of Caspase-1 and Interleukin 1beta in Salmonella-Infected Macrophages. Nat. Immunol. 2006, 7, 576–582. [Google Scholar] [CrossRef]
- Zhao, Y.; Yang, J.; Shi, J.; Gong, Y.N.; Lu, Q.; Xu, H.; Liu, L.; Shao, F. The NLRC4 Inflammasome Receptors for Bacterial Flagellin and Type III Secretion Apparatus. Nature 2011, 477, 596–600. [Google Scholar] [CrossRef]
- Vijay-Kumar, M.; Carvalho, F.A.; Aitken, J.D.; Fifadara, N.H.; Gewirtz, A.T. TLR5 or NLRC4 is Necessary and Sufficient for Promotion of Humoral Immunity by Flagellin. Eur. J. Immunol. 2010, 40, 3528–3534. [Google Scholar] [CrossRef]
- Lopez-Yglesias, A.H.; Zhao, X.; Quarles, E.K.; Lai, M.A.; VandenBos, T.; Strong, R.K.; Smith, K.D. Flagellin Induces Antibody Responses Through a TLR5- and Inflammasome-Independent Pathway. J. Immunol. 2014, 192, 1587–1596. [Google Scholar] [CrossRef]
- Dos Reis, E.C.; Leal, V.N.C.; Soares, J.; Fernandes, F.P.; Souza de Lima, D.; de Alencar, B.C.; Pontillo, A. Flagellin/NLRC4 Pathway Rescues NLRP3-Inflammasome Defect in Dendritic Cells From HIV-Infected Patients: Perspective for New Adjuvant in Immunocompromised Individuals. Front. Immunol. 2019, 10, 1291. [Google Scholar] [CrossRef]
- Cui, B.; Liu, X.; Fang, Y.; Zhou, P.; Zhang, Y.; Wang, Y. Flagellin as a Vaccine Adjuvant. Expert Rev. Vaccines 2018, 17, 335–349. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.E.; Kim, S.Y.; Jeong, B.C.; Kim, Y.R.; Bae, S.J.; Ahn, O.S.; Lee, J.J.; Song, H.C.; Kim, J.M.; Choy, H.E.; et al. A Bacterial Flagellin, Vibrio Vulnificus FlaB, has a Strong Mucosal Adjuvant Activity to Induce Protective Immunity. Infect. Immun. 2006, 74, 694–702. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Liu, L.; Wen, K.; Huang, J.; Geng, S.; Shen, J.; Pan, Z.; Jiao, X. Chimeric flagellin expressed by Salmonella typhimurium induces an ESAT-6-specific Th1-type immune response and CTL effects following intranasal immunization. Cell Mol Immunol. 2011, 8, 496–501. [Google Scholar] [CrossRef] [PubMed]
- Skountzou, I.; Martin, M.P.; Wang, B.; Ling, Y.; Koutsonanos, D.; Weldon, W.; Jacob, J.; Compans, R.W. Salmonella flagellins are potent adjuvants for intranasally administered whole inactivated influenza vaccine. Vaccine 2010, 28, 4103–4112. [Google Scholar] [CrossRef]
- Chaung, H.C.; Cheng, L.T.; Hung, L.H.; Tsai, P.C.; Skountzou, I.; Wang, B.; Compans, R.W.; Lien, Y.Y. Salmonella flagellin enhances mucosal immunity of avian influenza vaccine in chickens. Vet. Microbiol. 2012, 157, 69–77. [Google Scholar] [CrossRef]
- Hong, S.H.; Byun, Y.H.; Nguyen, C.T.; Kim, S.Y.; Seong, B.L.; Park, S.; Woo, G.J.; Yoon, Y.; Koh, J.T.; Fujihashi, K.; et al. Intranasal Administration of a Flagellin-Adjuvanted Inactivated Influenza Vaccine Enhances Mucosal Immune Responses to Protect Mice Against Lethal Infection. Vaccine 2012, 30, 466–474. [Google Scholar] [CrossRef]
- Lai, C.H.; Tang, N.; Jan, J.T.; Huang, M.H.; Lu, C.Y.; Chiang, B.L.; Huang, L.M.; Wu, S.C. Use of recombinant flagellin in oil-inwater emulsions enhances hemagglutinin-specific mucosal IgA production and IL-17 secreting T cells against H5N1 avian influenza virus infection. Vaccine 2015, 33, 4321–4329. [Google Scholar] [CrossRef]
- Behrouz, B.; Mahdavi, M.; Amirmozafari, N.; Fatemi, M.J.; Irajian, G.; Bahroudi, M.; Hashemi, F.B. Immunogenicity of Pseudomonas aeruginosa recombinant b-type fagellin as a vaccine candidate: Protective efficacy in a murine burn wound sepsis model. Burns 2016, 2, S0305-417930028-6. [Google Scholar] [CrossRef]
- Chauhan, N.; Kumar, R.; Badhai, J.; Preet, A.; Yadava, P.K. Immunogenicity of cholera toxin B epitope inserted in Salmonella flagellin expressed on bacteria and administered as DNA vaccine. Mol. Cell Biochem. 2005, 276, 1–6. [Google Scholar] [CrossRef]
- Poirier, T.P.; Kehoe, M.A.; Beachey, E.H. Protective immunity evoked by oral administration of attenuated aroA Salmonella typhimurium expressing cloned streptococcal M protein. J Exp Med. 1988, 168, 25–32. [Google Scholar] [CrossRef]
- Verma, N.K.; Ziegler, H.K.; Wilson, M.; Khan, M.; Safley, S.; Stocker, B.A.; Schoolnik, G.K. Delivery of class I and class II MHC-restricted T-cell epitopes of listeriolysin of Listeria monocytogenes by attenuated Salmonella. Vaccine 1995, 13, 142–150. [Google Scholar] [CrossRef]
- Newton, S.M.; Joys, T.M.; Anderson, S.A.; Kennedy, R.C.; Hovi, M.E.; Stocker, B.A. Expression and immunogenicity of an 18-residue epitope of HIV1 gp41 inserted in the flagellar protein of a Salmonella live vaccine. Res. Microbiol. 1995, 146, 203–216. [Google Scholar] [CrossRef]
- Luna, M.G.; Martins, M.M.; Newton, S.M.; Costa, S.O.; Almeida, D.F.; Ferreira, L.C. Cloning and expression of colonization factor antigen I (CFA/I) epitopes of enterotoxigenic Escherichia coli (ETEC) in Salmonella flagellin. Res. Microbiol. 1997, 148, 217–228. [Google Scholar] [CrossRef]
- Talbot, R.J.; Newton, D.; Priest, N.D.; Austin, J.G.; Day, J.P. Inter-subject variability in the metabolism of aluminium following intravenous injection as citrate. Hum. Exp. Toxicol. 1995, 14, 595–599. [Google Scholar] [CrossRef]
- Treanor, J.J.; Taylor, D.N.; Tussey, L.; Hay, C.; Nolan, C.; Fitzgerald, T.; Liu, G.; Kavita, U.; Song, L.; Dark, I.; et al. Safety and Immunogenicity of a Recombinant Hemagglutinin Influenza-Flagellin Fusion Vaccine (VAX125) in Healthy Young Adults. Vaccine 2010, 28, 8268–8274. [Google Scholar] [CrossRef]
- Ghosh, T.K.; Mickelson, D.J.; Fink, J.; Solberg, J.C.; Inglefield, J.R.; Hook, D.; Gupta, S.K.; Gibson, S.; Alkan, S.S. Toll-Like Receptor (TLR) 2-9 Agonists-Induced Cytokines and Chemokines: I. Comparison with T Cell Receptor-Induced Responses. Cell. Immunol. 2006, 243, 48–57. [Google Scholar] [CrossRef]
- Kwissa, M.; Nakaya, H.I.; Oluoch, H.; Pulendran, B. Distinct TLR Adjuvants Differentially Stimulate Systemic and Local Innate Immune Responses in Nonhuman Primates. Blood 2012, 119, 2044–2055. [Google Scholar] [CrossRef]
- Kieffer, M.E.; Patel, A.M.; Hollingsworth, S.A.; Seganish, W.M. Small Molecule Agonists of Toll-Like Receptors 7 and 8: A Patent Review 2014–2020. Expert Opin. Ther. Pat. 2020, 30, 825–845. [Google Scholar] [CrossRef]
- Vasilakos, J.P.; Tomai, M.A. The Use of Toll-Like Receptor 7/8 Agonists as Vaccine Adjuvants. Expert Rev. Vaccines 2013, 12, 809–819. [Google Scholar] [CrossRef]
- Wu, T.Y.; Singh, M.; Miller, A.T.; De Gregorio, E.; Doro, F.; D’Oro, U.; Skibinski, D.A.; Mbow, M.L.; Bufali, S.; Herman, A.E.; et al. Rational design of small molecules as vaccine adjuvants. Sci. Transl. Med. 2014, 6, 263ra160. [Google Scholar] [CrossRef]
- Oh, J.Z.; Kedl, R.M. The Capacity to Induce Cross-Presentation Dictates the Success of a TLR7 Agonist-Conjugate Vaccine for Eliciting Cellular Immunity. J. Immunol. 2010, 185, 4602–4608. [Google Scholar] [CrossRef] [PubMed]
- Kastenmuller, K.; Wille-Reece, U.; Lindsay, R.W.; Trager, L.R.; Darrah, P.A.; Flynn, B.J.; Becker, M.R.; Udey, M.C.; Clausen, B.E.; Igyarto, B.Z.; et al. Protective T Cell Immunity in Mice Following Protein-TLR7/8 Agonist-Conjugate Immunization Requires Aggregationand Multiple DC Subsets. J. Clin. Investig. 2011, 121, 1782–1796. [Google Scholar] [CrossRef] [PubMed]
- Holbrook, B.C.; Aycock, S.T.; Machiele, E.; Clemens, E.; Gries, D.; Jorgensen, M.J.; Hadimani, M.B.; King, S.B.; Alexander-Miller, M.A. An R848 Adjuvanted Influenza Vaccine Promotes Early Activation of B Cells in the Draining Lymph Nodes of non-Human Primate Neonates. Immunology 2018, 153, 357–367. [Google Scholar] [CrossRef] [PubMed]
- Bhagchandani, S.; Johnson, J.; Irvine, D.J. Evolution of Toll-Like Receptor 7/8 Agonist Therapeutics and Their Delivery Approaches: From Antiviral Formulations to Vaccine Adjuvants. Adv. Drug Deliv. Rev. 2021, 175, 113803. [Google Scholar] [CrossRef] [PubMed]
- Fox, C.B.; Sivananthan, S.J.; Duthie, M.S.; Vergara, J.; Guderian, J.A.; Moon, E.; Coblentz, D.; Reed, S.G.; Carter, D. A Nanoliposome Delivery System to Synergistically Trigger TLR4 AND Tlr7. J. Nanobiotechnol. 2014, 12, 17. [Google Scholar] [CrossRef]
- Moody, M.A.; Santra, S.; Vandergrift, N.A.; Sutherland, L.L.; Gurley, T.C.; Drinker, M.S.; Allen, A.A.; Xia, S.M.; Meyerhoff, R.R.; Parks, R.; et al. Toll-Like Receptor 7/8 (TLR7/8) and TLR9 Agonists Cooperate to Enhance HIV-1 Envelope Antibody Responses in Rhesus Macaques. J. Virol. 2014, 88, 3329–3339. [Google Scholar] [CrossRef]
- Goff, P.H.; Hayashi, T.; Martinez-Gil, L.; Corr, M.; Crain, B.; Yao, S.; Cottam, H.B.; Chan, M.; Ramos, I.; Eggink, D.; et al. Synthetic Toll-Like Receptor 4 (TLR4) and TLR7 Ligands as Influenza Virus Vaccine Adjuvants Induce Rapid, Sustained, and Broadly Protective Responses. J. Virol. 2015, 89, 3221–3235. [Google Scholar] [CrossRef]
- Kawai, T.; Akira, S. The Role of Pattern-Recognition Receptors in Innate Immunity: Update on Toll-Like Receptors. Nat. Immunol. 2010, 11, 373. [Google Scholar] [CrossRef]
- Hanagata, N. Structure-Dependent Immunostimulatory Effect of CpG Oligodeoxynucleotides and Their Delivery System. Int. J. Nanomed. 2012, 7, 2181–2195. [Google Scholar] [CrossRef]
- Akira, S.; Uematsu, S.; Takeuchi, O. Pathogen Recognition and Innate Immunity. Cell 2006, 124, 783–801. [Google Scholar] [CrossRef]
- Vollmer, J.; Weeratna, R.; Payette, P.; Jurk, M.; Schetter, C.; Laucht, M.; Wader, T.; Tluk, S.; Liu, M.; Davis, H.L.; et al. Characterization of three CpG oligodeoxynucleotide classes with distinct immunostimulatory activities. Eur. J. Immunol. 2004, 34, 251–262. [Google Scholar] [CrossRef] [PubMed]
- Guiducci, C.; Ott, G.; Chan, J.H.; Damon, E.; Calacsan, C.; Matray, T.; Coffman, R.L.; Barrat, F.J. Properties Regulating the Nature of the Plasmacytoid Dendritic Cell Response to Toll-Like Receptor 9 Activation. J. Exp. Med. 2006, 203, 1999–2008. [Google Scholar] [CrossRef] [PubMed]
- Hartmann, G. Chapter Four—Nucleic Acid Immunity. In Advances in Immunology; Alt, F.W., Ed.; Elsevier: Amsterdam, The Netherlands, 2017; pp. 121–169. [Google Scholar]
- Barry, M.; Cooper, C. Review of Hepatitis B Surface Antigen-1018 ISS Adjuvant-Containing Vaccine Safety and Efficacy. Expert Opin. Biol. Ther. 2007, 7, 1731–1737. [Google Scholar] [CrossRef] [PubMed]
- Abramowicz, M.; Zuccotti, G.; Pflomm, J. A Two-Dose Hepatitis B Vaccine for Adults (Heplisav-B). JAMA 2018, 319, 822–823. [Google Scholar]
- Speiser, D.E.; Liénard, D.; Rufer, N.; Rubio-Godoy, V.; Rimoldi, D.; Lejeune, F.; Krieg, A.M.; Cerottini, J.C.; Romero, P. Rapid and Strong Human CD8+ T Cell Responses to Vaccination with Peptide, IFA, and CpG Oligodeoxynucleotide 7909. J. Clin. Investig. 2005, 115, 739–746. [Google Scholar] [CrossRef]
- Kuo, T.Y.; Lin, M.Y.; Coffman, R.L.; Campbell, J.D.; Traquina, P.; Lin, Y.-J.; Liu, L.T.C.; Cheng, J.; Wu, Y.C.; Wu, C.C.; et al. Development of CpG-Adjuvanted Stable Prefusion SARS-CoV-2 Spike Antigen as a Subunit Vaccine Against COVID-19. Sci. Rep. 2020, 10, 20085. [Google Scholar] [CrossRef]
- Ellis, R.D.; Wu, Y.; Martin, L.B.; Shaffer, D.; Miura, K.; Aebig, J.; Orcutt, A.; Rausch, K.; Zhu, D.; Mogensen, A.; et al. Phase 1 Study in Malaria Naive Adults of BSAM2/Alhydrogel(R)+CPG 7909, a Blood Stage Vaccine against P. Falciparum Malaria. PLoS ONE 2012, 7, e46094. [Google Scholar] [CrossRef]
- Kapp, K.; Kleuss, C.; Schroff, M.; Wittig, B. Genuine Immunomodulation with dSLIM. Mol. Ther. Nucleic Acids 2014, 3, e170. [Google Scholar] [CrossRef]
- Wittig, B.; Schmidt, M.; Scheithauer, W.; Schmoll, H.J. MGN1703, an Immunomodulator and Toll-Like Receptor 9 (TLR-9) Agonist: From Bench to Bedside. Crit. Rev. Oncol. Hematol. 2015, 94, 31–44. [Google Scholar] [CrossRef]
- Shoenfeld, Y.; Agmon-Levin, N. ‘ASIA’—Autoimmune/inflammatory syndrome induced by adjuvants. J. Autoimmun. 2011, 36, 4–8. [Google Scholar] [CrossRef]
- Watad, A.; Sharif, K.; Shoenfeld, Y. The ASIA syndrome: Basic concepts. Mediterr. J. Rheumatol. 2017, 28, 64–69. [Google Scholar] [CrossRef] [PubMed]
- Arango, M.T.; Perricone, C.; Kivity, S.; Cipriano, E.; Ceccarelli, F.; Valesini, G.; Shoenfeld, Y. HLA-DRB1 the notorious gene in the mosaic of autoimmunity. Immunol. Res. 2017, 65, 82–98. [Google Scholar] [CrossRef] [PubMed]
- Christen, U. Pathogen infection and autoimmune disease. Clin. Exp. Immunol. 2019, 195, 10–14. [Google Scholar] [CrossRef] [PubMed]
- Smatti, M.K.; Cyprian, F.S.; Nasrallah, G.K.; Al Thani, A.A.; Almishal, R.O.; Yassine, H.M. Viruses and Autoimmunity: A Review on the Potential Interaction and Molecular Mechanisms. Viruses 2019, 11, 762. [Google Scholar] [CrossRef]
- Caravantes-Cortes, M.I.; Roldan-Valadez, E.; Zwojewski-Martinez, R.D.; Salazar-Ruiz, S.Y.; Carballo-Zarate, A.A. Breast Prosthesis Syndrome: Pathophysiology and Management Algorithm. Aestheti. Plast. Surg. 2020, 44, 1423–1437. [Google Scholar] [CrossRef]
- Raslan, H.M.; Attia, H.R.; Salama, I.; Ibrahim, M.H.; Hassan, E.M.; El Hussieny, M.S.; El Menyawi, M.M.; Amr, K.S. Association of PTPN22 1858CT polymorphism, HLA-DRB1 shared epitope and autoantibodies with rheumatoid arthritis. Rheumatol. Int. 2016, 36, 1167–1175. [Google Scholar] [CrossRef]
- Steiner, S.; Becker, S.C.; Hartwig, J.; Sotzny, F.; Lorenz, S.; Bauer, S.; Löbel, M.; Stittrich, A.B.; Grabowski, P.; Scheibenbogen, C. Autoimmunity-Related Risk Variants in PTPN22 and CTLA4 Are Associated with ME/CFS with Infectious Onset. Front. Immunol. 2020, 11, 578. [Google Scholar] [CrossRef]
- Borba, V.; Malkova, A.; Basantsova, N.; Halpert, G.; Andreoli, L.; Tincani, A.; Amital, H.; Shoenfeld, Y. Classical Examples of the Concept of the ASIA Syndrome. Biomolecules 2020, 10, 1436. [Google Scholar]
- Vera-Lastra, O.; Medina, G.; Cruz-Domínguez, M.P.; Ramírez, G.M.; Blancas, R.B.P.; Amaro, A.L.P.; Martínez, A.V.; Delgado, J.S.; Jara, L.J. Autoimmune/inflammatory syndrome induced by mineral oil: A health problem. Clin. Rheumatol. 2018, 37, 1441–1448. [Google Scholar]
- Watad, A.; Rosenberg, V.; Tiosano, S.; Cohen Tervaert, J.W.; Yavne, Y.; Shoenfeld, Y.; Shalev, V.; Chodick, G.; Amital, H. Silicone breast implants and the risk of autoimmune/rheumatic disorders: A real-world analysis. Int. J. Epidemiol. 2018, 47, 1846–1854. [Google Scholar] [CrossRef]
- Tomljenovic, L.; Shaw, C.A. Mechanisms of aluminum adjuvant toxicity and autoimmunity in pediatric populations. Lupus 2012, 21, 223–230. [Google Scholar] [CrossRef] [PubMed]
- Geier, D.A.; Geier, M.R. A case-control study of quadrivalent human papillomavirus vaccine-associated autoimmune adverse events. Clin. Rheumatol. 2015, 34, 1225–1231. [Google Scholar] [CrossRef] [PubMed]
- Bragazzi, N.L.; Hejly, A.; Watad, A.; Adawi, M.; Amital, H.; Shoenfeld, Y. ASIA syndrome and endocrine autoimmune disorders. Best Pract. Res. Clin. Endocrinol. Metab. 2020, 34, 101412. [Google Scholar] [CrossRef] [PubMed]
- Linneberg, A.; Madsen, F.; Skaaby, T. Allergen-specific immunotherapy and risk of autoimmune disease. Curr. Opin. Allergy Clin. Immunol. 2012, 12, 635–639. [Google Scholar] [CrossRef] [PubMed]
- Cohen Tervaert, J.W. Autoinflammatory/autoimmunity syndrome induced by adjuvants (ASIA.; Shoenfeld’s syndrome): A new flame. Autoimmun. Rev. 2018, 17, 1259–1264. [Google Scholar] [CrossRef]
- Watad, A.; Bragazzi, N.L.; McGonagle, D.; Adawi, M.; Bridgewood, C.; Damiani, G.; Alijotas-Reig, J.; Esteve-Valverde, E.; Quaresma, M.; Amital, H.; et al. Autoimmune/inflammatory syndrome induced by adjuvants (ASIA) demonstrates distinct autoimmune and autoinflammatory disease associations according to the adjuvant subtype: Insights from an analysis of 500 cases. Clin. Immunol. 2019, 203, 1–8. [Google Scholar] [CrossRef]
- Ameratunga, R.; Gillis, D.; Gold, M.; Linneberg, A.; Elwood, J.M. Evidence Refuting the Existence of Autoimmune/Autoinflammatory Syndrome Induced by Adjuvants (ASIA). J. Allergy Clin. Immunol. Pract. 2017, 5, 1551–1555.e1. [Google Scholar] [CrossRef]
- Elwood, J.M.; Ameratunga, R. Autoimmune diseases after hepatitis B immunization in adults: Literature review and meta-analysis, with reference to ‘autoimmune/autoinflammatory syndrome induced by adjuvants’ (ASIA). Vaccine 2018, 36, 5796–5802. [Google Scholar] [CrossRef]
- Segal, Y.; Dahan, S.; Sharif, K.; Bragazzi, N.L.; Watad, A.; Amital, H. The value of Autoimmune Syndrome Induced by Adjuvant (ASIA)—Shedding light on orphan diseases in autoimmunity. Autoimmun. Rev. 2018, 17, 440–448. [Google Scholar] [CrossRef]


{kind=link}
| Adjuvant Groups | Types of Adjuvants |
|---|---|
| Delivery systems | |
| Mineral Salts | Aluminium salts |
| Emulsions | Freund’s adjuvants |
| MF59 | |
| AS03 | |
| Microparticles | Virus-like particles |
| Virosomes | |
| PLA/PLGA | |
| Immune Potentiators | |
| TLR1/2 agonists | L-pampo, MALP-2, Pam2CSK4 and Pam3CSK4 |
| TLR3 agonists | Poly(I:C) (polyinosinic:polycytidylic acid) |
| Poly-ICLC | |
| TLR4 agonists | Monophosphoryl lipid A (MPL) |
| TLR5 agonists | Flagellin |
| TLR7/8 agonists | Imiquimod (R837; 1-(2-methylpropyl)-1H-imidazo [4,5-c]quinolin-4-amine) and resiquimod (R848, 4-amino-2-(etoximetil)-a,a-dimethyl-1H-imidazo [4,5-c]quinoline-1-ethanol) |
| TLR9 agonists | CpG ODNs |
| Combined adjuvants | AS01 and AS02 |
| AS04 | |
| Mucosal adjuvants | Cholera toxin (CT) |
| Heat-labile enterotoxin (LTK3 and LTR72) | |
| Chitosan |
| Trade Name | Type | Adjuvant | Administration |
|---|---|---|---|
| ADACEL | Tetanus Toxoid, Reduced Diphtheria Toxoid and Acellular Pertussis Vaccine | Aluminum phosphate (0.3 mg Al3+) | Intramuscular |
| AVAXIM | Inactivated hepatitis A vaccine | Aluminium hydroxide (0.3 mg Al3+) | Intramuscular |
| BEXSERO | Group B meningococcal vaccine (recombinant) | Aluminium hydroxide (0.5 mg Al3+) | Intramuscular |
| BIOTHRAX | Anthrax Vaccine Adsorbed | Aluminium hydroxide (0.6 mg Al3+) | Intramuscular/ Subcutaneous |
| BOOSTRIX | Diphtheria, tetanus and pertussis (acellular component) vaccine (reduced antigen content) | Aluminium hydroxide, (0.3 mg Al3+) and Aluminum phosphate (0.2 mg Al3+) | Intramuscular |
| CERVARIX | Human Papillomavirus vaccine (types 16, 18) (recombinant) | AS04 containing 3-O-desacyl-4’-monophosphoryl lipid A (MPL) 50 μg adsorbed on Aluminum hydroxide (0.5 mg Al3+) | Intramuscular |
| DAPTACEL | Diphtheria & Tetanus Toxoids & Acellular Pertussis Vaccine Adsorbed | Aluminum phosphate (0.3 mg Al3+) | Intramuscular |
| DIFTETALL | Diphtheria and tetanus vaccine (reduced antigenic content) | Aluminium hydroxide (1.5 mg) | Intramuscular |
| ENGERIX B | Hepatitis B (recombinant) vaccine | Aluminium hydroxide, (0.25 mg Al3+) | Intramuscular |
| FENDRIX | Hepatitis B (recombinant) vaccine | AS04 containing 3-O-desacyl-4′-monophosphoryl lipid A (MPL) 50 μg adsorbed on Aluminum hydroxide (0.5 mg Al3+) | Intramuscular |
| FLUAD | Inactivated influenza vaccine, surface antigen | MF59 containing 9.75 mg of squalene; 1.175 mg of polysorbate 80; 1.175 mg of sorbitan trioleate; 0.66 mg of sodium citrate; 0.04 mg of citric acid. | Intramuscular |
| GARDASIL | Human Papillomavirus Quadrivalent (Types 6, 11, 16, 18) Vaccine (recombinant) | Amorphous aluminum hydroxyphosphate sulfate (0.225 mg Al3+) | Intramuscular |
| GARDASIL 9 | Human Papillomavirus 9-valent vaccine (recombinant) | Amorphous aluminum hydroxyphosphate sulfate (0.5 mg di Al3+). | Intramuscular |
| HAVRIX | Inactivated hepatitis A vaccine | Aluminium hydroxide (0.5 mg Al3+) | Intramuscular |
| HBVAXPRO | Hepatitis B vaccine, purified antigen (recombinant) | Amorphous aluminium hydroxyphosphate sulfate (0.25 mg Al3+) | intramuscular |
| HEPLISAV-B | Hepatitis B Vaccine (recombinant) | CpG 1018 | Intramuscular |
| HEXYON | Diphtheria, tetanus, pertussis (acellular component), hepatitis B (recombinant, poliomyelitis (inactivated) and Haemophilus influenzae type b conjugated vaccine | Aluminium hydroxide (0.6 mg Al3+) | Intramuscular |
| IMOVAX TETANO | Tetanus vaccine | Aluminium hydroxide (0.6 mg Al3+) | Intramuscular |
| INFANRIX HEXA | Diphtheria, tetanus, pertussis (acellular component), antihepatitis B (recombinant), polio (inactivated) and anti-Haemophilus influenzae type b conjugate vaccine. | Aluminium hydroxide (0.5 mg Al3+) | Intramuscular |
| IXIARO | Japanese Encephalitis Virus Vaccine, Inactivated, Adsorbed | Aluminum hydroxide (0.25 mg Al3+) | Intramuscular |
| KINRIX | Diphtheria and Tetanus Toxoids and Acellular Pertussis Adsorbed and Inactivated Poliovirus Vaccine | Aluminum hydroxide (0.6 mg Al3+) | Intramuscular |
| MENJUGATE | Group C Meningococcal vaccine Conjugated | Aluminium hydroxide (0.3–0.4 mg Al3+) | Intramuscular |
| NEISVAC-C | Conjugated polysaccharide vaccine | Aluminium hydroxide (0.5 mg Al3+) | Intramuscular |
| NUVAXOVID | COVID-19 Vaccine (recombinant, adjuvanted) | Matrix-M containing Fraction-A (42.5 μg) and Fraction-C (7.5 μg) of Quillaja saponaria Molina extract | Intramuscular |
| PEDIARIX | Diphtheria & Tetanus Toxoids & Acellular Pertussis Vaccine Adsorbed, Hepatitis B (recombinant) and Inactivated Poliovirus Vaccine Combined | Aluminum hydroxide and Aluminum phosphate (0.85 mg Al3+) | Intramuscular |
| PEDVAXHIB | Haemophilus influenzae b Conjugate Vaccine (Meningococcal Protein Conjugate) | Amorphous aluminum hydroxyphosphate sulfate (0.225 mg Al3+) | Intramuscular |
| PENTACEL | Diphtheria and Tetanus Toxoids and Acellular Pertussis Adsorbed, Inactivated Poliovirus and Haemophilus influenzae b Conjugate (Tetanus Toxoid Conjugate) Vaccine | Aluminum phosphate (0.3 mg Al3+) | Intramuscular |
| POLIOBOOSTRIX | Diphtheria, tetanus, pertussis (acellular component) and poliomyelitis (inactivated) vaccine (with reduced antigen content) | Aluminium hydroxide (0.3 mg Al3+) and Aluminum phosphate (0.2 mg Al3+) | Intramuscular |
| POLIOINFANRIX | Diphtheria, tetanus, pertussis (acellular component) and poliomyelitis (inactivated) vaccine | Aluminium hydroxide, (0.5 mg Al3+) | Intramuscular |
| PREVENAR 13 | Pneumococcal polysaccharide conjugate vaccine (13-valent) | Aluminium hydroxide (0.125 mg Al3+) | Intramuscular |
| QUADRACEL | Diphtheria and Tetanus Toxoids and Acellular Pertussis Adsorbed and Inactivated Poliovirus Vaccine | Aluminum phosphate (0.33 mg Al3+) | Intramuscular |
| RECOMBIVAX HB | Hepatitis B Vaccine (recombinant) | Amorphous aluminum hydroxyphosphate sulfate (0.25–0.5 mg Al3+) | Intramuscular |
| REVAXIS | Diphtheria, tetanus and poliomyelitis (inactivated) vaccine (reduced antigenic content) | Aluminium hydroxide (0.35 mg Al3+) | Intramuscular/ Subcutaneous |
| SHINGRIX | Herpes zoster vaccine (recombinant) | AS01B containing Quillaja saponaria Molina plant extract, fraction 21 (QS-21) 50 μg, 3-O-desacyl-4′-monophosphoryl lipid A (MPL) from Salmonella minnesota 50 μg | Intramuscular |
| SYNFLORIX | Pneumococcal polysaccharide conjugated vaccine | Aluminum phosphate (0.5 mg Al3+) | Intramuscular |
| TDVAX | Tetanus & Diphtheria Toxoids, Adsorbed | Aluminum phosphate (0.5 mg Al3+) | Intramuscular |
| TENIVAC | Tetanus & Diphtheria Toxoids Adsorbed for Adult Use | Aluminum phosphate (0.33 mg Al3+) | Intramuscular |
| TETRAVAC | Diphtheria, tetanus, pertussis (acellular component) and poliomyelitis (inactivated) vaccine | Aluminium hydroxide (0.3 mg Al3+) | Intramuscular |
| TICOVAC | Tick-borne encephalitis Vaccine (whole virus, inactivated) | Aluminium hydroxide (0.17 mg Al3+) | Intramuscular |
| TRIAXIS | Diphtheria, tetanus, pertussis (acellular components) vaccine (reduced antigenic content) | Aluminum phosphate (0.33 mg Al3+) | Intramuscular |
| TRIAXIS POLIO | Diphtheria, tetanus, pertussis (acellular components) and poliomyelitis (inactivated) vaccine (reduced antigenic content) | Aluminum phosphate (0.33 mg Al3+) | Intramuscular |
| TRIBACCINE | Diphtheria, tetanus and pertussis (acellular component) vaccine (reduced antigenic content) | Aluminium hydroxide (0.5 mg Al3+) | Intramuscular |
| TRUMENBA | Group B meningococcal vaccine (recombinant) | Aluminum phosphate (0.25 mg Al3+) | Intramuscular |
| TWINRIX | Hepatitis A (inactivated) and Hepatitis B (recombinant) vaccine | Aluminium hydroxide (0.05 mg Al3+) Aluminum phosphate (0.4 mg Al3+) | Intramuscular |
| TWINRIX ADULTS | Hepatitis A (inactivated) and Hepatitis B (recombinant) vaccine | Aluminium hydroxide (0.05 mg Al3+) | Intramuscular |
| VAQTA | Inactivated Hepatitis A vaccine | Amorphous aluminum hydroxyphosphate sulfate (0.45 mg di Al3+) | Intramuscular |
| VAXELIS | Diphtheria, tetanus, pertussis (acellular component), hepatitis B (recombinant), polyomyelitis (inactivated) and Haemophilus influenzae type b vaccine | Aluminum phosphate (0.17 mg Al3+) Amorphous aluminium hydroxyphosphate sulfate (0.15 mg Al3+) | Intramuscular |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Facciolà, A.; Visalli, G.; Laganà, A.; Di Pietro, A. An Overview of Vaccine Adjuvants: Current Evidence and Future Perspectives. Vaccines 2022, 10, 819. https://doi.org/10.3390/vaccines10050819
Facciolà A, Visalli G, Laganà A, Di Pietro A. An Overview of Vaccine Adjuvants: Current Evidence and Future Perspectives. Vaccines. 2022; 10(5):819. https://doi.org/10.3390/vaccines10050819
Chicago/Turabian StyleFacciolà, Alessio, Giuseppa Visalli, Antonio Laganà, and Angela Di Pietro. 2022. "An Overview of Vaccine Adjuvants: Current Evidence and Future Perspectives" Vaccines 10, no. 5: 819. https://doi.org/10.3390/vaccines10050819
APA StyleFacciolà, A., Visalli, G., Laganà, A., & Di Pietro, A. (2022). An Overview of Vaccine Adjuvants: Current Evidence and Future Perspectives. Vaccines, 10(5), 819. https://doi.org/10.3390/vaccines10050819