
Post-Immune Antibodies in HIV-1 Infection in the Context of Vaccine Development: A Variety of Biological Functions and Catalytic Activities




Abstract
1. Introduction
2. Neutralizing Antibodies in HIV-1 Infection
2.1. General Characteristics of nAbs
2.2. Antibodies against Env Epitopes
2.3. Hypervariable Domains gp120
3. Autoantibodies in HIV-1 Infection
4. Catalytic Antibodies in HIV-1
4.1. Catalytic Antibodies Cleave gp120
4.2. Antibodies Hydrolyze HIV-1 Integrase, Reverse Transcriptase, and Corresponding Oligopeptides
4.3. Antibodies Hydrolyze DNA, Histones, and Myelin Basic Protein
5. HIV-1 Vaccines: Production of Neutralizing Antibodies
5.1. HIV-1 Vaccine Development Strategies
5.2. Immunogen Structure Design
5.3. Vaccination Inducing Immune Response at Mucous Membranes
5.4. Covalent Epitope-Based Vaccines
5.5. HIV-1 mRNA Vaccines
6. Immune Exhaustion and Antibody Response
7. COVID-19 and HIV-1
7.1. Immunological Features of COVID-19 in HIV-1 Infected Patients
7.2. SARS-CoV-2 and HIV-1 Vaccines
8. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Barré-Sinoussi, F.; Ross, A.L.; Delfraissy, J.-F. Past, present and future: 30 years of HIV research. Nat. Rev. Microbiol. 2013, 11, 877–883. [Google Scholar] [CrossRef] [PubMed]
- Taylor, B.S.; Sobieszczyk, M.E.; McCutchan, F.E.; Hammer, S.M. The Challenge of HIV-1 Subtype Diversity. N. Engl. J. Med. 2008, 358, 1590–1602. [Google Scholar] [CrossRef]
- Hemelaar, J.; Gouws, E.; Ghys, P.D.; Osmanov, S. Global and regional distribution of HIV-1 genetic subtypes and recombinants in 2004. AIDS 2006, 20, W13–W23. [Google Scholar] [CrossRef] [PubMed]
- Korber, B.; Gaschen, B.; Yusim, K.; Thakallapally, R.; Kesmir, C.; Detours, V. Evolutionary and immunological implications of contemporary HIV-1 variation. Br. Med. Bull. 2001, 58, 19–42. [Google Scholar] [CrossRef] [PubMed]
- Wei, X.; Decker, J.M.; Wang, S.; Hui, H.; Kappes, J.C.; Wu, X.; Salazar-Gonzalez, J.F.; Salazar, M.G.; Kilby, J.M.; Saag, M.S.; et al. Antibody neutralization and escape by HIV-1. Nature 2003, 422, 307–312. [Google Scholar] [CrossRef] [PubMed]
- Richman, D.D.; Wrin, T.; Little, S.J.; Petropoulos, C.J. Rapid evolution of the neutralizing antibody response to HIV type 1 infection. Proc. Natl. Acad. Sci. USA 2003, 100, 4144–4149. [Google Scholar] [CrossRef]
- Liu, J.; Bartesaghi, A.; Borgnia, M.J.; Sapiro, G.; Subramaniam, S. Molecular architecture of native HIV-1 gp120 trimers. Nature 2008, 455, 109–113. [Google Scholar] [CrossRef]
- Kwong, P.D.; Wyatt, R.; Robinson, J.; Sweet, R.W.; Sodroski, J.; Hendrickson, W.A. Structure of an HIV gp120 envelope glycoprotein in complex with the CD4 receptor and a neutralizing human antibody. Nature 1998, 393, 648–659. [Google Scholar] [CrossRef]
- Chen, B.; Vogan, E.M.; Gong, H.; Skehel, J.J.; Wiley, D.C.; Harrison, S.C. Structure of an unliganded simian immunodeficiency virus gp120 core. Nature 2005, 433, 834–841. [Google Scholar] [CrossRef]
- Yu, W.-H.; Zhao, P.; Draghi, M.; Arevalo, C.; Karsten, C.B.; Suscovich, T.J.; Gunn, B.; Streeck, H.; Brass, A.L.; Tiemeyer, M.; et al. Exploiting glycan topography for computational design of Env glycoprotein antigenicity. PLoS Comput. Biol. 2018, 14, e1006093. [Google Scholar] [CrossRef]
- Barin, F.; McLane, M.F.; Allan, J.S.; Lee, T.H.; Groopman, J.E.; Essex, M. Virus Envelope Protein of HTLV-III Represents Major Target Antigen for Antibodies in AIDS Patients. Science 1985, 228, 1094–1096. [Google Scholar] [CrossRef] [PubMed]
- Allan, J.S.; Coligan, J.E.; Barin, F.; McLane, M.F.; Sodroski, J.G.; Rosen, C.A.; Haseltine, W.A.; Lee, T.H.; Essex, M. Major Glycoprotein Antigens That Induce Antibodies in AIDS Patients Are Encoded by HTLV-III. Science 1985, 228, 1091–1094. [Google Scholar] [CrossRef]
- Okabe, M.; Kubara, K.; Kawaguchi, H.; Kawano, T.; Nakashima, Y.; Fukuda, K.; Hiroki, T.; Arakawa, K.; Kikuchi, M. A case of myxedema with diffuse myocardial fibrosis proven by endomyocardial biopsy. Kokyu Junkan Respir. Circ. 1990, 38, 1159–1163. [Google Scholar]
- Moore, J.P.; Cao, Y.; Ho, D.D.; Koup, R.A. Development of the anti-gp120 antibody response during seroconversion to human immunodeficiency virus type 1. J. Virol. 1994, 68, 5142–5155. [Google Scholar] [CrossRef] [PubMed]
- Soudeyns, H.; Pantaleo, G. The moving target: Mechanisms of HIV persistence during primary infection. Immunol. Today 1999, 20, 446–450. [Google Scholar] [CrossRef]
- Cao, Y.; Qin, L.; Zhang, L.; Safrit, J.; Ho, D.D. Virologic and Immunologic Characterization of Long-Term Survivors of Human Immunodeficiency Virus Type 1 Infection. N. Engl. J. Med. 1995, 332, 201–208. [Google Scholar] [CrossRef]
- Walker, L.M.; Simek, M.D.; Priddy, F.; Gach, J.S.; Wagner, D.; Zwick, M.B.; Phogat, S.K.; Poignard, P.; Burton, D.R. A Limited Number of Antibody Specificities Mediate Broad and Potent Serum Neutralization in Selected HIV-1 Infected Individuals. PLoS Pathog. 2010, 6, e1001028. [Google Scholar] [CrossRef]
- Doria-Rose, N.A.; Klein, R.M.; Daniels, M.G.; O’Dell, S.; Nason, M.; Lapedes, A.; Bhattacharya, T.; Migueles, S.A.; Wyatt, R.T.; Korber, B.T.; et al. Breadth of Human Immunodeficiency Virus-Specific Neutralizing Activity in Sera: Clustering Analysis and Association with Clinical Variables. J. Virol. 2010, 84, 1631–1636. [Google Scholar] [CrossRef]
- Gray, E.S.; Madiga, M.C.; Hermanus, T.; Moore, P.L.; Wibmer, C.K.; Tumba, N.L.; Werner, L.; Mlisana, K.; Sibeko, S.; Williamson, C.; et al. The Neutralization Breadth of HIV-1 Develops Incrementally over Four Years and Is Associated with CD4+ T Cell Decline and High Viral Load during Acute Infection. J. Virol. 2011, 85, 4828–4840. [Google Scholar] [CrossRef]
- Burton, D.R.; Desrosiers, R.C.; Doms, R.W.; Koff, W.C.; Kwong, P.D.; Moore, J.P.; Nabel, G.J.; Sodroski, J.; Wilson, I.A.; Wyatt, R.T. HIV vaccine design and the neutralizing antibody problem. Nat. Immunol. 2004, 5, 233–236. [Google Scholar] [CrossRef]
- Kwong, P.D.; Doyle, M.L.; Casper, D.J.; Cicala, C.; Leavitt, S.A.; Majeed, S.; Steenbeke, T.D.; Venturi, M.; Chaiken, I.; Fung, M.; et al. HIV-1 evades antibody-mediated neutralization through conformational masking of receptor-binding sites. Nature 2002, 420, 678–682. [Google Scholar] [CrossRef] [PubMed]
- Mascola, J.R.; Montefiori, D.C. HIV-1: Nature’s master of disguise. Nat. Med. 2003, 9, 393–394. [Google Scholar] [CrossRef] [PubMed]
- Kessler, J.A.; McKenna, P.M.; Emini, E.A.; Chan, C.P.; Patel, M.D.; Gupta, S.K.; Mark, G.E.; Barbas, C.F.; Burton, D.R.; Conley, A.J. Recombinant Human Monoclonal Antibody IgG1b12 Neutralizes Diverse Human Immunodeficiency Virus Type 1 Primary Isolates. AIDS Res. Hum. Retroviruses 1997, 13, 575–582. [Google Scholar] [CrossRef] [PubMed]
- Parren, P.W.H.I.; Wang, M.; Trkola, A.; Binley, J.M.; Purtscher, M.; Katinger, H.; Moore, J.P.; Burton, D.R. Antibody Neutralization-Resistant Primary Isolates of Human Immunodeficiency Virus Type 1. J. Virol. 1998, 72, 10270–10274. [Google Scholar] [CrossRef]
- Trkola, A.; Purtscher, M.; Muster, T.; Ballaun, C.; Buchacher, A.; Sullivan, N.; Srinivasan, K.; Sodroski, J.; Moore, J.P.; Katinger, H. Human monoclonal antibody 2G12 defines a distinctive neutralization epitope on the gp120 glycoprotein of human immunodeficiency virus type 1. J. Virol. 1996, 70, 1100–1108. [Google Scholar] [CrossRef]
- Zwick, M.B.; Labrijn, A.F.; Wang, M.; Spenlehauer, C.; Saphire, E.O.; Binley, J.M.; Moore, J.P.; Stiegler, G.; Katinger, H.; Burton, D.R.; et al. Broadly Neutralizing Antibodies Targeted to the Membrane-Proximal External Region of Human Immunodeficiency Virus Type 1 Glycoprotein gp41. J. Virol. 2001, 75, 10892–10905. [Google Scholar] [CrossRef]
- Muster, T.; Steindl, F.; Purtscher, M.; Trkola, A.; Klima, A.; Himmler, G.; Rüker, F.; Katinger, H. A conserved neutralizing epitope on gp41 of human immunodeficiency virus type 1. J. Virol. 1993, 67, 6642–6647. [Google Scholar] [CrossRef]
- Wu, X.; Yang, Z.-Y.; Li, Y.; Hogerkorp, C.-M.; Schief, W.R.; Seaman, M.S.; Zhou, T.; Schmidt, S.D.; Wu, L.; Xu, L.; et al. Rational design of envelope identifies broadly neutralizing human monoclonal antibodies to HIV-1. Science 2010, 329, 856–861. [Google Scholar] [CrossRef]
- Conley, A.J.; Gorny, M.K.; Kessler, J.A.; Boots, L.J.; Ossorio-Castro, M.; Koenig, S.; Lineberger, D.W.; Emini, E.A.; Williams, C.; Zolla-Pazner, S. Neutralization of primary human immunodeficiency virus type 1 isolates by the broadly reactive anti-V3 monoclonal antibody, 447–452D. J. Virol. 1994, 68, 6994–7000. [Google Scholar] [CrossRef]
- Burton, D.R.; Pyati, J.; Koduri, R.; Sharp, S.J.; Thornton, G.B.; Parren, P.W.H.I.; Sawyer, L.S.W.; Hendry, R.M.; Dunlop, N.; Nara, P.L.; et al. Efficient Neutralization of Primary Isolates of HIV-1 by a Recombinant Human Monoclonal Antibody. Science 1994, 266, 1024–1027. [Google Scholar] [CrossRef]
- Binley, J.M.; Wrin, T.; Korber, B.; Zwick, M.B.; Wang, M.; Chappey, C.; Stiegler, G.; Kunert, R.; Zolla-Pazner, S.; Katinger, H.; et al. Comprehensive Cross-Clade Neutralization Analysis of a Panel of Anti-Human Immunodeficiency Virus Type 1 Monoclonal Antibodies. J. Virol. 2004, 78, 13232–13252. [Google Scholar] [CrossRef] [PubMed]
- Walker, L.M.; Phogat, S.K.; Chan-Hui, P.-Y.; Wagner, D.; Phung, P.; Goss, J.L.; Wrin, T.; Simek, M.D.; Fling, S.; Mitcham, J.L.; et al. Broad and Potent Neutralizing Antibodies from an African Donor Reveal a New HIV-1 Vaccine Target. Science 2009, 326, 285–289. [Google Scholar] [CrossRef] [PubMed]
- Corti, D.; Langedijk, J.P.M.; Hinz, A.; Seaman, M.S.; Vanzetta, F.; Fernandez-Rodriguez, B.M.; Silacci, C.; Pinna, D.; Jarrossay, D.; Balla-Jhagjhoorsingh, S.; et al. Analysis of Memory B Cell Responses and Isolation of Novel Monoclonal Antibodies with Neutralizing Breadth from HIV-1-Infected Individuals. PLoS ONE 2010, 5, e8805. [Google Scholar] [CrossRef]
- Zhou, T.; Xu, L.; Dey, B.; Hessell, A.J.; Van Ryk, D.; Xiang, S.-H.; Yang, X.; Zhang, M.-Y.; Zwick, M.B.; Arthos, J.; et al. Structural definition of a conserved neutralization epitope on HIV-1 gp120. Nature 2007, 445, 732–737. [Google Scholar] [CrossRef]
- Morrow, W.J.W.; Isenberg, D.A.; Sobol, R.E.; Stricker, R.B.; Kieber-Emmons, T. AIDS virus infection and autoimmunity: A perspective of the clinical, immunological, and molecular origins of the autoallergic pathologies associated with HIV disease. Clin. Immunol. Immunopathol. 1991, 58, 163–180. [Google Scholar] [CrossRef]
- González, C.M.; López-LONGO, F.J.; Samson, J.; Monteagudo, I.; Grau, R.; Rodríguez-Mahou, M.; ST-Cyr, C.; Lapointe, N.; Carreño, L. Antiribonucleoprotein Antibodies in Children with HIV Infection: A Comparative Study with Childhood-Onset Systemic Lupus Erythematosus. AIDS Patient Care STDS 1998, 12, 21–28. [Google Scholar] [CrossRef] [PubMed]
- Argov, S.; Schattner, A.; Burstein, R.; Handzel, Z.T.; Shoenfeld, Y.; Bentwich, Z. Autoantibodies in male homosexuals and HIV infection. Immunol. Lett. 1991, 30, 31–35. [Google Scholar] [CrossRef]
- Zandman-Goddard, G.; Shoenfeld, Y. HIV and autoimmunity. Autoimmun. Rev. 2002, 1, 329–337. [Google Scholar] [CrossRef]
- Sizyakina, L.P.; Buneva, V.N.; Nevinsky, G.A.; Odintsova, E.S.; Kharitonova, M.A.; Baranovskii, A.G.; Sizyakina, L.P.; Buneva, V.N.; Nevinsky, G.A. Proteolytic activity of IgG antibodies from blood of acquired immunodeficiency syndrome patients. Biochemistry 2006, 71, 251–261. [Google Scholar] [CrossRef]
- Baranova, S.V.; Buneva, V.N.; Nevinsky, G.A. Antibodies from the sera of HIV-infected patients efficiently hydrolyze all human histones. J. Mol. Recognit. 2016, 29, 346–362. [Google Scholar] [CrossRef]
- Baranova, S.V.; Dmitrienok, P.S.; Buneva, V.N.; Nevinsky, G.A. Autoantibodies in HIV-infected patients: Cross site-specific hydrolysis of H1 histone and myelin basic protein. BioFactors 2019, 45, 211–222. [Google Scholar] [CrossRef] [PubMed]
- Baranova, S.V.; Buneva, V.N.; Kharitonova, M.A.; Sizyakina, L.P.; Calmels, C.; Andreola, M.-L.; Parissi, V.; Nevinsky, G. HIV-1 integrase-hydrolyzing antibodies from sera of HIV-infected patients. Biochimie 2009, 91, 1081–1086. [Google Scholar] [CrossRef] [PubMed]
- Baranova, S.V.; Buneva, V.N.; Kharitonova, M.A.; Sizyakina, L.P.; Calmels, C.; Andreola, M.-L.M.-L.; Parissi, V.; Zakharova, O.D.; Nevinsky, G.A. HIV-1 integrase-hydrolyzing IgM antibodies from sera of HIV-infected patients. Int. Immunol. 2010, 22, 671–680. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Miedema, F. A brief history of HIV vaccine research: Stepping back to the drawing board? AIDS 2008, 22, 1699–1703. [Google Scholar] [CrossRef]
- Pitisuttithum, P.; Gilbert, P.; Gurwith, M.; Heyward, W.; Martin, M.; van Griensven, F.; Hu, D.; Tappero, J.W. Bangkok Vaccine Evaluation Group Randomized, Double-Blind, Placebo-Controlled Efficacy Trial of a Bivalent Recombinant Glycoprotein 120 HIV-1 Vaccine among Injection Drug Users in Bangkok, Thailand. J. Infect. Dis. 2006, 194, 1661–1671. [Google Scholar] [CrossRef]
- Sekaly, R.-P. The failed HIV Merck vaccine study: A step back or a launching point for future vaccine development? J. Exp. Med. 2008, 205, 7–12. [Google Scholar] [CrossRef]
- Rerks-Ngarm, S.; Pitisuttithum, P.; Nitayaphan, S.; Kaewkungwal, J.; Chiu, J.; Paris, R.; Premsri, N.; Namwat, C.; de Souza, M.; Adams, E.; et al. Vaccination with ALVAC and AIDSVAX to Prevent HIV-1 Infection in Thailand. N. Engl. J. Med. 2009, 361, 2209–2220. [Google Scholar] [CrossRef]
- Paul, S.; Planque, S.; Nishiyama, Y.; Escobar, M.; Hanson, C. Back to the future: Covalent epitope-based HIV vaccine development. Expert Rev. Vaccines 2010, 9, 1027–1043. [Google Scholar] [CrossRef]
- Spencer, D.A.; Shapiro, M.B.; Haigwood, N.L.; Hessell, A.J. Advancing HIV Broadly Neutralizing Antibodies: From Discovery to the Clinic. Front. Public Health 2021, 9, 610. [Google Scholar] [CrossRef]
- Williams, W.B.; Wiehe, K.; Saunders, K.O.; Haynes, B.F. Strategies for induction of HIV-1 envelope-reactive broadly neutralizing antibodies. J. Int. AIDS Soc. 2021, 24, e25831. [Google Scholar] [CrossRef]
- Verkoczy, L.; Alt, F.W.; Tian, M. Human Ig knockin mice to study the development and regulation of HIV-1 broadly neutralizing antibodies. Immunol. Rev. 2017, 275, 89–107. [Google Scholar] [CrossRef] [PubMed]
- Stamatatos, L.; Pancera, M.; McGuire, A.T. Germline-targeting immunogens. Immunol. Rev. 2017, 275, 203–216. [Google Scholar] [CrossRef] [PubMed]
- Huang, D.; Tran, J.T.; Olson, A.; Vollbrecht, T.; Tenuta, M.; Guryleva, M.V.; Fuller, R.P.; Schiffner, T.; Abadejos, J.R.; Couvrette, L.; et al. Vaccine elicitation of HIV broadly neutralizing antibodies from engineered B cells. Nat. Commun. 2020, 11, 5850. [Google Scholar] [CrossRef] [PubMed]
- Fauci, A.S.; Marston, H.D. Toward an HIV vaccine: A scientific journey. Science 2015, 349, 386–387. [Google Scholar] [CrossRef]
- Mascola, J.R.; Montefiori, D.C. The Role of Antibodies in HIV Vaccines. Annu. Rev. Immunol. 2010, 28, 413–444. [Google Scholar] [CrossRef]
- Pantophlet, R.; Burton, D.R. GP120: Target for Neutralizing HIV-1 Antibodies. Annu. Rev. Immunol. 2006, 24, 739–769. [Google Scholar] [CrossRef]
- Montefiori, D.C.; Morris, L.; Ferrari, G.; Mascola, J.R. Neutralizing and other antiviral antibodies in HIV-1 infection and vaccination. Curr. Opin. HIV AIDS 2007, 2, 169–176. [Google Scholar] [CrossRef]
- Cheng-Mayer, C.; Homsy, J.; Evans, L.A.; Levy, J.A. Identification of human immunodeficiency virus subtypes with distinct patterns of sensitivity to serum neutralization. Proc. Natl. Acad. Sci. USA 1988, 85, 2815–2819. [Google Scholar] [CrossRef]
- Brown, B.K.; Wieczorek, L.; Sanders-Buell, E.; Borges, A.R.; Robb, M.L.; Birx, D.L.; Michael, N.L.; McCutchan, F.E.; Polonis, V.R. Cross-clade neutralization patterns among HIV-1 strains from the six major clades of the pandemic evaluated and compared in two different models. Virology 2008, 375, 529–538. [Google Scholar] [CrossRef]
- Kostrikis, L.G.; Cao, Y.; Ngai, H.; Moore, J.P.; Ho, D.D. Quantitative analysis of serum neutralization of human immunodeficiency virus type 1 from subtypes A, B, C, D, E, F, and I: Lack of direct correlation between neutralization serotypes and genetic subtypes and evidence for prevalent serum-dependent infecti. J. Virol. 1996, 70, 445–458. [Google Scholar] [CrossRef]
- Haynes, B.F.; Burton, D.R.; Mascola, J.R. Multiple roles for HIV broadly neutralizing antibodies. Sci. Transl. Med. 2019, 11, eaaz2686. [Google Scholar] [CrossRef] [PubMed]
- Burton, D.R.; Hangartner, L. Broadly Neutralizing Antibodies to HIV and Their Role in Vaccine Design. Annu. Rev. Immunol. 2016, 34, 635–659. [Google Scholar] [CrossRef] [PubMed]
- Stamatatos, L.; Morris, L.; Burton, D.R.; Mascola, J.R. Neutralizing antibodies generated during natural HIV-1 infection: Good news for an HIV-1 vaccine? Nat. Med. 2009, 15, 866–870. [Google Scholar] [CrossRef] [PubMed]
- Sather, D.N.; Armann, J.; Ching, L.K.; Mavrantoni, A.; Sellhorn, G.; Caldwell, Z.; Yu, X.; Wood, B.; Self, S.; Kalams, S.; et al. Factors Associated with the Development of Cross-Reactive Neutralizing Antibodies during Human Immunodeficiency Virus Type 1 Infection. J. Virol. 2009, 83, 757–769. [Google Scholar] [CrossRef]
- Simek, M.D.; Rida, W.; Priddy, F.H.; Pung, P.; Carrow, E.; Laufer, D.S.; Lehrman, J.K.; Boaz, M.; Tarragona-Fiol, T.; Miiro, G.; et al. Human Immunodeficiency Virus Type 1 Elite Neutralizers: Individuals with Broad and Potent Neutralizing Activity Identified by Using a High-Throughput Neutralization Assay together with an Analytical Selection Algorithm. J. Virol. 2009, 83, 7337–7348. [Google Scholar] [CrossRef]
- Doria-Rose, N.A.; Klein, R.M.; Manion, M.M.; O’Dell, S.; Phogat, A.; Chakrabarti, B.; Hallahan, C.W.; Migueles, S.A.; Wrammert, J.; Ahmed, R.; et al. Frequency and Phenotype of Human Immunodeficiency Virus Envelope-Specific B Cells from Patients with Broadly Cross-Neutralizing Antibodies. J. Virol. 2009, 83, 188–199. [Google Scholar] [CrossRef]
- Montefiori, D.; Sattentau, Q.; Flores, J.; Esparza, J.; Mascola, J. Antibody-Based HIV-1 Vaccines: Recent Developments and Future Directions. PLoS Med. 2007, 4, e348. [Google Scholar] [CrossRef]
- Karsten, C.B.; Alter, G. The HIV-1 Glycan Shield: Strategically Placed Kinks in the Armor Improve Antigen Design. Cell Rep. 2017, 19, 669–670. [Google Scholar] [CrossRef][Green Version]
- Cohen, Y.Z.; Lavine, C.L.; Miller, C.A.; Garrity, J.; Carey, B.R.; Seaman, M.S. Glycan-Dependent Neutralizing Antibodies Are Frequently Elicited in Individuals Chronically Infected with HIV-1 Clade B or C. AIDS Res. Hum. Retroviruses 2015, 31, 1192–1201. [Google Scholar] [CrossRef]
- Gray, E.S.; Moore, P.L.; Choge, I.A.; Decker, J.M.; Bibollet-Ruche, F.; Li, H.; Leseka, N.; Treurnicht, F.; Mlisana, K.; Shaw, G.M.; et al. Neutralizing Antibody Responses in Acute Human Immunodeficiency Virus Type 1 Subtype C Infection. J. Virol. 2007, 81, 6187–6196. [Google Scholar] [CrossRef]
- Tomaras, G.D.; Yates, N.L.; Liu, P.; Qin, L.; Fouda, G.G.; Chavez, L.L.; Decamp, A.C.; Parks, R.J.; Ashley, V.C.; Lucas, J.T.; et al. Initial B-Cell Responses to Transmitted Human Immunodeficiency Virus Type 1: Virion-Binding Immunoglobulin M (IgM) and IgG Antibodies Followed by Plasma Anti-gp41 Antibodies with Ineffective Control of Initial Viremia. J. Virol. 2008, 82, 12449–12463. [Google Scholar] [CrossRef] [PubMed]
- Li, B.; Decker, J.M.; Johnson, R.W.; Bibollet-Ruche, F.; Wei, X.; Mulenga, J.; Allen, S.; Hunter, E.; Hahn, B.H.; Shaw, G.M.; et al. Evidence for Potent Autologous Neutralizing Antibody Titers and Compact Envelopes in Early Infection with Subtype C Human Immunodeficiency Virus Type 1. J. Virol. 2006, 80, 5211–5218. [Google Scholar] [CrossRef] [PubMed]
- Moore, P.L.; Gray, E.S.; Choge, I.A.; Ranchobe, N.; Mlisana, K.; Abdool Karim, S.S.; Williamson, C.; Morris, L. The C3-V4 Region Is a Major Target of Autologous Neutralizing Antibodies in Human Immunodeficiency Virus Type 1 Subtype C Infection. J. Virol. 2008, 82, 1860–1869. [Google Scholar] [CrossRef] [PubMed]
- Euler, Z.; van Gils, M.J.; Bunnik, E.M.; Phung, P.; Schweighardt, B.; Wrin, T.; Schuitemaker, H. Cross-Reactive Neutralizing Humoral Immunity Does Not Protect from HIV Type 1 Disease Progression. J. Infect. Dis. 2010, 201, 1045–1053. [Google Scholar] [CrossRef]
- Mikell, I.; Sather, D.N.; Kalams, S.A.; Altfeld, M.; Alter, G.; Stamatatos, L. Characteristics of the Earliest Cross-Neutralizing Antibody Response to HIV-1. PLoS Pathog. 2011, 7, e1001251. [Google Scholar] [CrossRef]
- Torán, J.L.; Kremer, L.; Sánchez-Pulido, L.; de Alborán, I.M.; del Real, G.; Llorente, M.; Valencia, A.; de Mon, M.A.; Martínez, A.C. Molecular analysis of HIV-1 gp120 antibody response using isotype IgM and IgG phage display libraries from a long-term non-progressor HIV-1-infected individual. Eur. J. Immunol. 1999, 29, 2666–2675. [Google Scholar] [CrossRef]
- Pancera, M.; McLellan, J.S.; Wu, X.; Zhu, J.; Changela, A.; Schmidt, S.D.; Yang, Y.; Zhou, T.; Phogat, S.; Mascola, J.R.; et al. Crystal Structure of PG16 and Chimeric Dissection with Somatically Related PG9: Structure-Function Analysis of Two Quaternary-Specific Antibodies That Effectively Neutralize HIV-1. J. Virol. 2010, 84, 8098–8110. [Google Scholar] [CrossRef]
- Zhou, T.; Georgiev, I.; Wu, X.; Yang, Z.-Y.; Dai, K.; Finzi, A.; Do Kwon, Y.; Scheid, J.F.; Shi, W.; Xu, L.; et al. Structural Basis for Broad and Potent Neutralization of HIV-1 by Antibody VRC01. Science 2010, 329, 811–817. [Google Scholar] [CrossRef]
- Moir, S.; Fauci, A.S. B cells in HIV infection and disease. Nat. Rev. Immunol. 2009, 9, 235–245. [Google Scholar] [CrossRef]
- van Gils, M.J.; Bunnik, E.M.; Burger, J.A.; Jacob, Y.; Schweighardt, B.; Wrin, T.; Schuitemaker, H. Rapid Escape from Preserved Cross-Reactive Neutralizing Humoral Immunity without Loss of Viral Fitness in HIV-1-Infected Progressors and Long-Term Nonprogressors. J. Virol. 2010, 84, 3576–3585. [Google Scholar] [CrossRef][Green Version]
- Haynes, B.F.; Montefiori, D.C. Aiming to induce broadly reactive neutralizing antibody responses with HIV-1 vaccine candidates. Expert Rev. Vaccines 2006, 5, 579–595. [Google Scholar] [CrossRef] [PubMed]
- Letvin, N.L. Progress and obstacles in the development of an AIDS vaccine. Nat. Rev. Immunol. 2006, 6, 930–939. [Google Scholar] [CrossRef] [PubMed]
- Barouch, D.H. Challenges in the development of an HIV-1 vaccine. Nature 2008, 455, 613–619. [Google Scholar] [CrossRef] [PubMed]
- Binley, J. Specificities of broadly neutralizing anti-HIV-1 sera. Curr. Opin. HIV AIDS 2009, 4, 364–372. [Google Scholar] [CrossRef]
- Gray, E.S.; Taylor, N.; Wycuff, D.; Moore, P.L.; Tomaras, G.D.; Wibmer, C.K.; Puren, A.; DeCamp, A.; Gilbert, P.B.; Wood, B.; et al. Antibody Specificities Associated with Neutralization Breadth in Plasma from Human Immunodeficiency Virus Type 1 Subtype C-Infected Blood Donors. J. Virol. 2009, 83, 8925–8937. [Google Scholar] [CrossRef]
- Li, Y.; Svehla, K.; Louder, M.K.; Wycuff, D.; Phogat, S.; Tang, M.; Migueles, S.A.; Wu, X.; Phogat, A.; Shaw, G.M.; et al. Analysis of Neutralization Specificities in Polyclonal Sera Derived from Human Immunodeficiency Virus Type 1-Infected Individuals. J. Virol. 2009, 83, 1045–1059. [Google Scholar] [CrossRef]
- Gray, E.S.; Madiga, M.C.; Moore, P.L.; Mlisana, K.; Abdool Karim, S.S.; Binley, J.M.; Shaw, G.M.; Mascola, J.R.; Morris, L. Broad Neutralization of Human Immunodeficiency Virus Type 1 Mediated by Plasma Antibodies against the gp41 Membrane Proximal External Region. J. Virol. 2009, 83, 11265–11274. [Google Scholar] [CrossRef]
- Sather, D.N.; Stamatatos, L. Epitope specificities of broadly neutralizing plasmas from HIV-1 infected subjects. Vaccine 2010, 28, B8–B12. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Moore, P.L.; Gray, E.S.; Sheward, D.; Madiga, M.; Ranchobe, N.; Lai, Z.; Honnen, W.J.; Nonyane, M.; Tumba, N.; Hermanus, T.; et al. Potent and Broad Neutralization of HIV-1 Subtype C by Plasma Antibodies Targeting a Quaternary Epitope Including Residues in the V2 Loop. J. Virol. 2011, 85, 3128–3141. [Google Scholar] [CrossRef]
- Wyatt, R.; Kwong, P.D.; Desjardins, E.; Sweet, R.W.; Robinson, J.; Hendrickson, W.A.; Sodroski, J.G. The antigenic structure of the HIV gp120 envelope glycoprotein. Nature 1998, 393, 705–711. [Google Scholar] [CrossRef]
- Binley, J.M.; Lybarger, E.A.; Crooks, E.T.; Seaman, M.S.; Gray, E.; Davis, K.L.; Decker, J.M.; Wycuff, D.; Harris, L.; Hawkins, N.; et al. Profiling the Specificity of Neutralizing Antibodies in a Large Panel of Plasmas from Patients Chronically Infected with Human Immunodeficiency Virus Type 1 Subtypes B and C. J. Virol. 2008, 82, 11651–11668. [Google Scholar] [CrossRef] [PubMed]
- Decker, J.M.; Bibollet-Ruche, F.; Wei, X.; Wang, S.; Levy, D.N.; Wang, W.; Delaporte, E.; Peeters, M.; Derdeyn, C.A.; Allen, S.; et al. Antigenic conservation and immunogenicity of the HIV coreceptor binding site. J. Exp. Med. 2005, 201, 1407–1419. [Google Scholar] [CrossRef] [PubMed]
- Labrijn, A.F.; Poignard, P.; Raja, A.; Zwick, M.B.; Delgado, K.; Franti, M.; Binley, J.; Vivona, V.; Grundner, C.; Huang, C.-C.; et al. Access of Antibody Molecules to the Conserved Coreceptor Binding Site on Glycoprotein gp120 Is Sterically Restricted on Primary Human Immunodeficiency Virus Type 1. J. Virol. 2003, 77, 10557–10565. [Google Scholar] [CrossRef] [PubMed]
- Haynes, B.F.; Fleming, J.; St Clair, E.W.; Katinger, H.; Stiegler, G.; Kunert, R.; Robinson, J.; Scearce, R.M.; Plonk, K.; Staats, H.F.; et al. Cardiolipin polyspecific autoreactivity in two broadly neutralizing HIV-1 antibodies. Science 2005, 308, 1906–1908. [Google Scholar] [CrossRef] [PubMed]
- Rong, R.; Bibollet-Ruche, F.; Mulenga, J.; Allen, S.; Blackwell, J.L.; Derdeyn, C.A. Role of V1V2 and Other Human Immunodeficiency Virus Type 1 Envelope Domains in Resistance to Autologous Neutralization during Clade C Infection. J. Virol. 2007, 81, 1350–1359. [Google Scholar] [CrossRef]
- Krachmarov, C.; Pinter, A.; Honnen, W.J.; Gorny, M.K.; Nyambi, P.N.; Zolla-Pazner, S.; Kayman, S.C. Antibodies That Are Cross-Reactive for Human Immunodeficiency Virus Type 1 Clade A and Clade B V3 Domains Are Common in Patient Sera from Cameroon, but Their Neutralization Activity Is Usually Restricted by Epitope Masking. J. Virol. 2005, 79, 780–790. [Google Scholar] [CrossRef]
- Krachmarov, C.P.; Honnen, W.J.; Kayman, S.C.; Gorny, M.K.; Zolla-Pazner, S.; Pinter, A. Factors Determining the Breadth and Potency of Neutralization by V3-Specific Human Monoclonal Antibodies Derived from Subjects Infected with Clade A or Clade B Strains of Human Immunodeficiency Virus Type 1. J. Virol. 2006, 80, 7127–7135. [Google Scholar] [CrossRef][Green Version]
- Gaschen, B.; Taylor, J.; Yusim, K.; Foley, B.; Gao, F.; Lang, D.; Novitsky, V.; Haynes, B.; Hahn, B.H.; Bhattacharya, T.; et al. Diversity Considerations in HIV-1 Vaccine Selection. Science 2002, 296, 2354–2360. [Google Scholar] [CrossRef]
- Li, M.; Gao, F.; Mascola, J.R.; Stamatatos, L.; Polonis, V.R.; Koutsoukos, M.; Voss, G.; Goepfert, P.; Gilbert, P.; Greene, K.M.; et al. Human Immunodeficiency Virus Type 1 env Clones from Acute and Early Subtype B Infections for Standardized Assessments of Vaccine-Elicited Neutralizing Antibodies. J. Virol. 2005, 79, 10108–10125. [Google Scholar] [CrossRef]
- Hartley, O.; Klasse, P.J.; Sattentau, Q.J.; Moore, J.P. V3: HIV’s Switch-Hitter. AIDS Res. Hum. Retroviruses 2005, 21, 171–189. [Google Scholar] [CrossRef]
- Pinter, A.; Honnen, W.J.; He, Y.; Gorny, M.K.; Zolla-Pazner, S.; Kayman, S.C. The V1/V2 Domain of gp120 Is a Global Regulator of the Sensitivity of Primary Human Immunodeficiency Virus Type 1 Isolates to Neutralization by Antibodies Commonly Induced upon Infection. J. Virol. 2004, 78, 5205–5215. [Google Scholar] [CrossRef] [PubMed]
- Kinsey, N.E.; Anderson, M.G.; Unangst, T.J.; Joag, S.V.; Narayan, O.; Zink, M.C.; Clements, J.E. Antigenic Variation of SIV: Mutations in V4 Alter the Neutralization Profile. Virology 1996, 221, 14–21. [Google Scholar] [CrossRef]
- Teeraputon, S.; Louisirirojchanakul, S.; Auewarakul, P. N-Linked Glycosylation in C2 Region of HIV-1 Envelope Reduces Sensitivity to Neutralizing Antibodies. Viral Immunol. 2005, 18, 343–353. [Google Scholar] [CrossRef]
- Fauci, A.S. Multifactorial Nature of Human Immunodeficiency Virus Disease: Implications for Therapy. Science 1993, 262, 1011–1018. [Google Scholar] [CrossRef]
- Tomar, R.H.; John, P.A.; Hennig, A.K.; Kloster, B. Cellular targets of antilymphocyte antibodies in AIDS and LAS. Clin. Immunol. Immunopathol. 1985, 37, 37–47. [Google Scholar] [CrossRef]
- Tausk, F.A.; McCutchan, A.; Spechko, P.; Schreiber, R.D.; Gigli, I. Altered erythrocyte C3b receptor expression, immune complexes, and complement activation in homosexual men in varying risk groups for acquired immune deficiency syndrome. J. Clin. Investig. 1986, 78, 977–982. [Google Scholar] [CrossRef]
- Ameglio, F.; Dolei, A.; Benedetto, A.; Sorrentino, R.; Tanigaki, N.; Tosi, R. Antibodies Reactive with Nonpolymorphic Epitopes on HLA Molecules Interfere in Screening Tests for the Human Immunodeficiency Virus. J. Infect. Dis. 1987, 156, 1034–1035. [Google Scholar] [CrossRef] [PubMed]
- Morris, L.; Distenfeld, A.; Amorosi, E.; Karpatkin, S. Autoimmune Thrombocytopenic Purpura in Homosexual Men. Ann. Intern. Med. 1982, 96, 714–717. [Google Scholar] [CrossRef] [PubMed]
- Stricker, R.B.; Abrams, D.I.; Corash, L.; Shuman, M.A. Target Platelet Antigen in Homosexual Men with Immune Thrombocytopenia. N. Engl. J. Med. 1985, 313, 1375–1380. [Google Scholar] [CrossRef]
- Kiprov, D.; Pfaeffl, W.; Parry, G.; Lippert, R.; Lang, W.; Miller, R. Antibody-mediated peripheral neuropathies associated with ARC and AIDS: Successful treatment with plasmapheresis. J. Clin. Apher. 1988, 4, 3–7. [Google Scholar] [CrossRef]
- de La Monte, S.M.; Gabuzda, D.H.; Ho, D.D.; Brown, R.H.; Hedley-Whyte, E.T.; Schooley, R.T.; Hirsch, M.S.; Bhan, A.K. Peripheral neuropathy in the acquired immunodeficiency syndrome. Ann. Neurol. 1988, 23, 485–492. [Google Scholar] [CrossRef] [PubMed]
- De Angelis, V.; Biasinutto, C.; Pradella, P.; Vaccher, E.; Spina, M.; Tirelli, U. Clinical significance of positive direct antiglobulin test in patients with HIV infection. Infection 1994, 22, 92–95. [Google Scholar] [CrossRef] [PubMed]
- Lai, M.; Visconti, E.; D’Onofrio, G.; Tamburrini, E.; Cauda, R.; Leone, G. Lower hemoglobin levels in human immunodeficiency virus-infected patients with a positive direct antiglobulin test (DAT): Relationship with DAT strength and clinical stages. Transfusion 2006, 46, 1237–1243. [Google Scholar] [CrossRef] [PubMed]
- Yen, Y.-F.; Lan, Y.-C.; Huang, C.-T.; Jen, I.-A.; Chen, M.; Lee, C.-Y.; Chuang, P.-H.; Lee, Y.; Morisky, D.E.; Chen, Y.-M.A. Human Immunodeficiency Virus Infection Increases the Risk of Incident Autoimmune Hemolytic Anemia: A Population-Based Cohort Study in Taiwan. J. Infect. Dis. 2017, 216, 1000–1007. [Google Scholar] [CrossRef] [PubMed]
- Yen, Y.-F.; Chuang, P.-H.; Jen, I.-A.; Chen, M.; Lan, Y.-C.; Liu, Y.-L.; Lee, Y.; Chen, Y.-H.; Chen, Y.-M.A. Incidence of autoimmune diseases in a nationwide HIV/AIDS patient cohort in Taiwan, 2000–2012. Ann. Rheum. Dis. 2017, 76, 661–665. [Google Scholar] [CrossRef] [PubMed]
- Schved, J.F.; Dupuy-Fons, C.; Biron, C.; Quéré, I.; Janbon, C. A Prospective Epidemiological Study on the Occurrence of Antiphospholipid Antibody: The Montpellier Antiphospholipid (MAP) Study. Pathophysiol. Haemost. Thromb. 1994, 24, 175–182. [Google Scholar] [CrossRef]
- Silvestris, F.; Frassanito, M.; Cafforio, P.; Potenza, D.; Di Loreto, M.; Tucci, M.; Grizzuti, M.; Nico, B.; Dammacco, F. Antiphosphatidylserine antibodies in human immunodeficiency virus-1 patients with evidence of T-cell apoptosis and mediate antibody- dependent cellular cytotoxicity [see comments]. Blood 1996, 87, 5185–5195. [Google Scholar] [CrossRef]
- Girón-González, J.A.; García del Río, E.; Rodríguez, C.; Rodríguez-Martorell, J.; Serrano, A. Antiphospholipid syndrome and asymptomatic carriers of antiphospholipid antibody: Prospective analysis of 404 individuals. J. Rheumatol. 2004, 31, 1560–1567. [Google Scholar]
- Calabrese, L.H.; Kirchner, E.; Shrestha, R. Rheumatic Complications of Human Immunodeficiency Virus Infection in the Era of Highly Active Antiretroviral Therapy: Emergence of a New Syndrome of Immune Reconstitution and Changing Patterns of Disease. Semin. Arthritis Rheum. 2005, 35, 166–174. [Google Scholar] [CrossRef]
- Reveille, J.D. The changing spectrum of rheumatic disease in human immunodeficiency virus infection. Semin. Arthritis Rheum. 2000, 30, 147–166. [Google Scholar] [CrossRef]
- Mouquet, H.; Scheid, J.F.; Zoller, M.J.; Krogsgaard, M.; Ott, R.G.; Shukair, S.; Artyomov, M.N.; Pietzsch, J.; Connors, M.; Pereyra, F.; et al. Polyreactivity increases the apparent affinity of anti-HIV antibodies by heteroligation. Nature 2010, 467, 591–595. [Google Scholar] [CrossRef] [PubMed]
- Liao, H.; Zhang, Z. Polyreactive antibodies in anti-HIV-1 responses. Curr. Mol. Med. 2018, 18, 126–133. [Google Scholar] [CrossRef] [PubMed]
- Kobie, J.J.; Alcena, D.C.; Zheng, B.; Bryk, P.; Mattiacio, J.L.; Brewer, M.; LaBranche, C.; Young, F.M.; Dewhurst, S.; Montefiori, D.C.; et al. 9G4 Autoreactivity Is Increased in HIV-Infected Patients and Correlates with HIV Broadly Neutralizing Serum Activity. PLoS ONE 2012, 7, e35356. [Google Scholar] [CrossRef] [PubMed]
- Jia, M.; Liberatore, R.A.; Guo, Y.; Chan, K.-W.; Pan, R.; Lu, H.; Waltari, E.; Mittler, E.; Chandran, K.; Finzi, A.; et al. VSV-Displayed HIV-1 Envelope Identifies Broadly Neutralizing Antibodies Class-Switched to IgG and IgA. Cell Host Microbe 2020, 27, 963–975.e5. [Google Scholar] [CrossRef]
- Cizmeci, D.; Lofano, G.; Rossignol, E.; Dugast, A.-S.; Kim, D.; Cavet, G.; Nguyen, N.; Tan, Y.C.; Seaman, M.S.; Alter, G.; et al. Distinct clonal evolution of B-cells in HIV controllers with neutralizing antibody breadth. Elife 2021, 10, e62648. [Google Scholar] [CrossRef]
- Ternynck, T.; Falanga, P.B.; Unterklrscher, C.; Gregolre, J.; da Silva, L.P.; Avrameas, S. Induction of high levels of IgG autoantibodies in mice infected with Plasmodium chabaudi. Int. Immunol. 1991, 3, 29–37. [Google Scholar] [CrossRef]
- Hentati, B.; Sato, M.N.; Payelle-Brogard, B.; Avrameas, S.; Ternynck, T. Beneficial effect of polyclonal immunoglobulins from malaria-infected BALB/c mice on the lupus-like syndrome of (NZB × NZW)F1 mice. Eur. J. Immunol. 1994, 24, 8–15. [Google Scholar] [CrossRef]
- Barzilai, O.; Ram, M.; Shoenfeld, Y. Viral infection can induce the production of autoantibodies. Curr. Opin. Rheumatol. 2007, 19, 636–643. [Google Scholar] [CrossRef]
- Odintsova, E.S.; Kharitonova, M.A.; Baranovskii, A.G.; Sizyakina, L.P.; Buneva, V.N.; Nevinsky, G.A. DNA-hydrolyzing IgG antibodies from the blood of patients with acquired immune deficiency syndrome. Mol. Biol. 2006, 40, 770–777. [Google Scholar] [CrossRef]
- Baranova, S.V.; Dmitrienok, P.S.; Ivanisenko, N.V.; Buneva, V.N.; Nevinsky, G.A. Antibodies to H1 histone from the sera of HIV-infected patients recognize and catalyze site-specific degradation of this histone. J. Mol. Recognit. 2017, 30, e2588. [Google Scholar] [CrossRef]
- Baranova, S.V.; Dmitrienok, P.S.; Ivanisenko, N.V.; Buneva, V.N.; Nevinsky, G.A. Antibodies to H2a and H2b histones from the sera of HIV-infected patients catalyze site-specific degradation of these histones. Mol. Biosyst. 2017, 13, 1090–1101. [Google Scholar] [CrossRef] [PubMed]
- Baranova, S.V.; Dmitrenok, P.S.; Zubkova, A.D.; Ivanisenko, N.V.; Odintsova, E.S.; Buneva, V.N.; Nevinsky, G.A. Antibodies against H3 and H4 histones from the sera of HIV-infected patients catalyze site-specific degradation of these histones. J. Mol. Recognit. 2018, 31, e2703. [Google Scholar] [CrossRef] [PubMed]
- Odintsova, E.S.; Baranova, S.V.; Dmitrenok, P.S.; Rasskazov, V.A.; Calmels, C.; Parissi, V.; Andreola, M.-L.M.-L.; Buneva, V.N.; Zakharova, O.D.; Nevinsky, G.A. Antibodies to HIV integrase catalyze site-specific degradation of their antigen. Int. Immunol. 2011, 23, 601–612. [Google Scholar] [CrossRef][Green Version]
- Odintsova, E.S.; Baranova, S.V.; Dmitrenok, P.S.; Calmels, C.; Parissi, V.; Andreola, M.-L.; Buneva, V.N.; Nevinsky, G.A. Anti-integrase abzymes from the sera of HIV-infected patients specifically hydrolyze integrase but nonspecifically cleave short oligopeptides. J. Mol. Recognit. 2012, 25, 193–207. [Google Scholar] [CrossRef] [PubMed]
- Townsley-Fuchs, J.; Kam, L.; Fairhurst, R.; Gange, S.J.; Goodglick, L.; Giorgi, J.V.; Sidell, N.; Detels, R.; Braun, J. Human immunodeficiency virus-1 (HIV-1) gp120 superantigen-binding serum antibodies. A host factor in homosexual HIV-1 transmission. J. Clin. Investig. 1996, 98, 1794–1801. [Google Scholar] [CrossRef] [PubMed]
- Planque, S.; Mitsuda, Y.; Taguchi, H.; Salas, M.; Morris, M.-K.; Nishiyama, Y.; Kyle, R.; Okhuysen, P.; Escobar, M.; Hunter, R.; et al. Characterization of gp120 Hydrolysis by IgA Antibodies from Humans without HIV Infection. AIDS Res. Hum. Retroviruses 2007, 23, 1541–1554. [Google Scholar] [CrossRef]
- Paul, S.; Karle, S.; Planque, S.; Taguchi, H.; Salas, M.; Nishiyama, Y.; Handy, B.; Hunter, R.; Edmundson, A.; Hanson, C. Naturally occurring proteolytic antibodies: Selective immunoglobulin M-catalyzed hydrolysis of HIV gp120. J. Biol. Chem. 2004, 279, 39611–39619. [Google Scholar] [CrossRef]
- Gao, Q.-S.; Sun, M.; Rees, A.R.; Paul, S. Site-directed Mutagenesis of Proteolytic Antibody Light Chain. J. Mol. Biol. 1995, 253, 658–664. [Google Scholar] [CrossRef]
- Chorny, A.; Puga, I.; Cerutti, A. Innate Signalling Networks in Mucosal IgA Class Switching. Adv Immunol 2010, 107, 31–69. [Google Scholar] [CrossRef]
- Olshevsky, U.; Helseth, E.; Furman, C.; Li, J.; Haseltine, W.; Sodroski, J. Identification of individual human immunodeficiency virus type 1 gp120 amino acids important for CD4 receptor binding. J. Virol. 1990, 64, 5701–5707. [Google Scholar] [CrossRef]
- Nishiyama, Y.; Karle, S.; Planque, S.; Taguchi, H.; Paul, S. Antibodies to the superantigenic site of HIV-1 gp120: Hydrolytic and binding activities of the light chain subunit. Mol. Immunol. 2007, 44, 2707–2718. [Google Scholar] [CrossRef] [PubMed]
- Belogurov, A.; Kozyr, A.; Ponomarenko, N.; Gabibov, A. Catalytic antibodies: Balancing between Dr. Jekyll and Mr. Hyde. Bioessays 2009, 31, 1161–1171. [Google Scholar] [CrossRef] [PubMed]
- Odintsova, E.S.; Dmitrenok, P.S.; Buneva, V.N.; Nevinsky, G.A. Specific anti-integrase abzymes from HIV-infected patients: A comparison of the cleavage sites of intact globular HIV integrase and two 20-mer oligopeptides corresponding to its antigenic determinants. J. Mol. Recognit. 2013, 26, 121–135. [Google Scholar] [CrossRef] [PubMed]
- Odintsova, E.S.; Dmitrenok, P.S.; Timofeeva, A.M.; Buneva, V.N.; Nevinsky, G.A. Why specific anti-integrase antibodies from HIV-infected patients can efficiently hydrolyze 21-mer oligopeptide corresponding to antigenic determinant of human myelin basic protein. J. Mol. Recognit. 2014, 27, 32–45. [Google Scholar] [CrossRef] [PubMed]
- Paul, S.; Li, L.; Kalaga, R.; Wilkins-Stevens, P.; Stevens, F.J.; Solomon, A. Natural Catalytic Antibodies: Peptide-hydrolyzing Activities of Bence Jones Proteins and VL Fragment. J. Biol. Chem. 1995, 270, 15257–15261. [Google Scholar] [CrossRef]
- Paul, S.; Li, L.; Kalaga, R.; O’Dell, J.; Dannenbring, R.E.; Swindells, S.; Hinrichs, S.; Caturegli, P.; Rose, N.R. Characterization of thyroglobulin-directed and polyreactive catalytic antibodies in autoimmune disease. J. Immunol. 1997, 159, 1530–1536. [Google Scholar]
- Ponomarenko, N.A.; Durova, O.M.; Vorobiev, I.I.; Belogurov, A.A.; Kurkova, I.N.; Petrenko, A.G.; Telegin, G.B.; Suchkov, S.V.; Kiselev, S.L.; Lagarkova, M.A.; et al. Autoantibodies to myelin basic protein catalyze site-specific degradation of their antigen. Proc. Natl. Acad. Sci. USA 2006, 103, 281–286. [Google Scholar] [CrossRef]
- Nevinsky, G.A. How Enzymes, Proteins, and Antibodies Recognize Extended DNAs; General Regularities. Int. J. Mol. Sci. 2021, 22, 1369. [Google Scholar] [CrossRef]
- Nevinsky, G.A.; Buneva, V.N. Natural catalytic antibodies in norm, autoimmune, viral, and bacterial diseases. Sci. World J. 2010, 10, 820417. [Google Scholar] [CrossRef]
- Odintsova, E.S.; Dmitrenok, P.S.; Baranova, S.V.; Timofeeva, A.M.; Buneva, V.N.; Nevinsky, G.A. Features of hydrolysis of specific and nonspecific globular proteins and oligopeptides by antibodies against viral integrase from blood of HIV-infected patients. Biochemistry 2015, 80, 180–201. [Google Scholar] [CrossRef]
- Baranova, S.V.; Dmitrenok, P.S.; Buneva, V.N.; Sedykh, S.E.; Nevinsky, G.A. HIV-Infected Patients: Cross Site-Specific Hydrolysis of H3 and H4 Histones and Myelin Basic Protein with Antibodies against These Three Proteins. Molecules 2021, 26, 316. [Google Scholar] [CrossRef] [PubMed]
- Gololobov, G.V.; Mikhalap, S.V.; Starov, A.V.; Kolesnikov, A.F.; Gabibov, A.G. DNA-protein complexes. Appl. Biochem. Biotechnol. 1994, 47, 305–315. [Google Scholar] [CrossRef] [PubMed]
- Legostaeva, G.A.; Polosukhina, D.I.; Bezuglova, A.M.; Doronin, B.M.; Buneva, V.N.; Nevinsky, G.A. Affinity and catalytic heterogeneity of polyclonal myelin basic protein-hydrolyzing IgGs from sera of patients with multiple sclerosis. J. Cell. Mol. Med. 2010, 14, 699–709. [Google Scholar] [CrossRef]
- Bezuglova, A.M.; Konenkova, L.P.; Buneva, V.N.; Nevinsky, G.A. IgGs containing light chains of the λ- and κ- type and of all subclasses (IgG1-IgG4) from the sera of patients with systemic lupus erythematosus hydrolyze myelin basic protein. Int. Immunol. 2012, 24, 759–770. [Google Scholar] [CrossRef]
- Bezuglova, A.M.; Konenkova, L.P.; Doronin, B.M.; Buneva, V.N.; Nevinsky, G.A. Affinity and catalytic heterogeneity and metal-dependence of polyclonal myelin basic protein-hydrolyzing IgGs from sera of patients with systemic lupus erythematosus. J. Mol. Recognit. 2011, 24, 960–974. [Google Scholar] [CrossRef] [PubMed]
- Gulick, R.M.; Flexner, C. Long-Acting HIV Drugs for Treatment and Prevention. Annu. Rev. Med. 2019, 70, 137–150. [Google Scholar] [CrossRef]
- Haynes, B.F.; Burton, D.R. Developing an HIV vaccine. Science 2017, 355, 1129–1130. [Google Scholar] [CrossRef] [PubMed]
- Seaman, M.S.; Janes, H.; Hawkins, N.; Grandpre, L.E.; Devoy, C.; Giri, A.; Coffey, R.T.; Harris, L.; Wood, B.; Daniels, M.G.; et al. Tiered Categorization of a Diverse Panel of HIV-1 Env Pseudoviruses for Assessment of Neutralizing Antibodies. J. Virol. 2010, 84, 1439–1452. [Google Scholar] [CrossRef]
- Sarzotti-Kelsoe, M.; Bailer, R.T.; Turk, E.; Lin, C.; Bilska, M.; Greene, K.M.; Gao, H.; Todd, C.A.; Ozaki, D.A.; Seaman, M.S.; et al. Optimization and validation of the TZM-bl assay for standardized assessments of neutralizing antibodies against HIV-1. J. Immunol. Methods 2014, 409, 131–146. [Google Scholar] [CrossRef]
- Montefiori, D.C.; Roederer, M.; Morris, L.; Seaman, M.S. Neutralization tiers of HIV-1. Curr. Opin. HIV AIDS 2018, 13, 128–136. [Google Scholar] [CrossRef]
- Früh, K.; Picker, L. CD8+ T cell programming by cytomegalovirus vectors: Applications in prophylactic and therapeutic vaccination. Curr. Opin. Immunol. 2017, 47, 52–56. [Google Scholar] [CrossRef] [PubMed]
- Mascola, J.R.; D’Souza, P.; Gilbert, P.; Hahn, B.H.; Haigwood, N.L.; Morris, L.; Petropoulos, C.J.; Polonis, V.R.; Sarzotti, M.; Montefiori, D.C. Recommendations for the Design and Use of Standard Virus Panels To Assess Neutralizing Antibody Responses Elicited by Candidate Human Immunodeficiency Virus Type 1 Vaccines. J. Virol. 2005, 79, 10103–10107. [Google Scholar] [CrossRef] [PubMed]
- Moldt, B.; Rakasz, E.G.; Schultz, N.; Chan-Hui, P.-Y.; Swiderek, K.; Weisgrau, K.L.; Piaskowski, S.M.; Bergman, Z.; Watkins, D.I.; Poignard, P.; et al. Highly potent HIV-specific antibody neutralization in vitro translates into effective protection against mucosal SHIV challenge in vivo. Proc. Natl. Acad. Sci. USA 2012, 109, 18921–18925. [Google Scholar] [CrossRef]
- Moog, C.; Dereuddre-Bosquet, N.; Teillaud, J.-L.; Biedma, M.E.; Holl, V.; Van Ham, G.; Heyndrickx, L.; Van Dorsselaer, A.; Katinger, D.; Vcelar, B.; et al. Protective effect of vaginal application of neutralizing and nonneutralizing inhibitory antibodies against vaginal SHIV challenge in macaques. Mucosal Immunol. 2014, 7, 46–56. [Google Scholar] [CrossRef]
- Kwong, P.D.; Mascola, J.R.; Nabel, G.J. Broadly neutralizing antibodies and the search for an HIV-1 vaccine: The end of the beginning. Nat. Rev. Immunol. 2013, 13, 693–701. [Google Scholar] [CrossRef] [PubMed]
- Haut, L.H.; Ertl, H.C.J. Obstacles to the successful development of an efficacious T cell-inducing HIV-1 vaccine. J. Leukoc. Biol. 2009, 86, 779–793. [Google Scholar] [CrossRef] [PubMed]
- Zolla-Pazner, S.; Cohen, S.; Pinter, A.; Krachmarov, C.; Wrin, T.; Wang, S.; Lu, S. Cross-clade neutralizing antibodies against HIV-1 induced in rabbits by focusing the immune response on a neutralizing epitope. Virology 2009, 392, 82–93. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Forthal, D.N.; Landucci, G.; Cole, K.S.; Marthas, M.; Becerra, J.C.; Van Rompay, K. Rhesus Macaque Polyclonal and Monoclonal Antibodies Inhibit Simian Immunodeficiency Virus in the Presence of Human or Autologous Rhesus Effector Cells. J. Virol. 2006, 80, 9217–9225. [Google Scholar] [CrossRef]
- Gómez-Román, V.R.; Patterson, L.J.; Venzon, D.; Liewehr, D.; Aldrich, K.; Florese, R.; Robert-Guroff, M. Vaccine-Elicited Antibodies Mediate Antibody-Dependent Cellular Cytotoxicity Correlated with Significantly Reduced Acute Viremia in Rhesus Macaques Challenged with SIV mac251. J. Immunol. 2005, 174, 2185–2189. [Google Scholar] [CrossRef] [PubMed]
- Kwong, P.D.; Mascola, J.R. Human Antibodies that Neutralize HIV-1: Identification, Structures, and B Cell Ontogenies. Immunity 2012, 37, 412–425. [Google Scholar] [CrossRef]
- Briney, B.S.; Willis, J.R.; Crowe, J.E. Human Peripheral Blood Antibodies with Long HCDR3s Are Established Primarily at Original Recombination Using a Limited Subset of Germline Genes. PLoS ONE 2012, 7, e36750. [Google Scholar] [CrossRef] [PubMed]
- Yu, L.; Guan, Y. Immunologic Basis for Long HCDR3s in Broadly Neutralizing Antibodies Against HIV-1. Front. Immunol. 2014, 5, 250. [Google Scholar] [CrossRef] [PubMed]
- Steichen, J.M.; Lin, Y.-C.; Havenar-Daughton, C.; Pecetta, S.; Ozorowski, G.; Willis, J.R.; Toy, L.; Sok, D.; Liguori, A.; Kratochvil, S.; et al. A generalized HIV vaccine design strategy for priming of broadly neutralizing antibody responses. Science 2019, 366, aax4380. [Google Scholar] [CrossRef] [PubMed]
- Flemming, A. bnAbs for HIV: Shepherding towards improbable mutations. Nat. Rev. Drug Discov. 2020, 19, 90. [Google Scholar] [CrossRef] [PubMed]
- Willis, J.R.; Finn, J.A.; Briney, B.; Sapparapu, G.; Singh, V.; King, H.; LaBranche, C.C.; Montefiori, D.C.; Meiler, J.; Crowe, J.E. Long antibody HCDR3s from HIV-naïve donors presented on a PG9 neutralizing antibody background mediate HIV neutralization. Proc. Natl. Acad. Sci. USA 2016, 113, 4446–4451. [Google Scholar] [CrossRef]
- Finney, J.; Kelsoe, G. Poly- and autoreactivity of HIV-1 bNAbs: Implications for vaccine design. Retrovirology 2018, 15, 53. [Google Scholar] [CrossRef]
- Deeks, S.G.; Schweighardt, B.; Wrin, T.; Galovich, J.; Hoh, R.; Sinclair, E.; Hunt, P.; McCune, J.M.; Martin, J.N.; Petropoulos, C.J.; et al. Neutralizing Antibody Responses against Autologous and Heterologous Viruses in Acute versus Chronic Human Immunodeficiency Virus (HIV) Infection: Evidence for a Constraint on the Ability of HIV To Completely Evade Neutralizing Antibody Responses. J. Virol. 2006, 80, 6155–6164. [Google Scholar] [CrossRef]
- Draenert, R.; Allen, T.M.; Liu, Y.; Wrin, T.; Chappey, C.; Verrill, C.L.; Sirera, G.; Eldridge, R.L.; Lahaie, M.P.; Ruiz, L.; et al. Constraints on HIV-1 evolution and immunodominance revealed in monozygotic adult twins infected with the same virus. J. Exp. Med. 2006, 203, 529–539. [Google Scholar] [CrossRef]
- Verkoczy, L.; Diaz, M. Autoreactivity in HIV-1 broadly neutralizing antibodies. Curr. Opin. HIV AIDS 2014, 9, 224–234. [Google Scholar] [CrossRef]
- Verkoczy, L.; Diaz, M.; Holl, T.M.; Ouyang, Y.-B.; Bouton-Verville, H.; Alam, S.M.; Liao, H.-X.; Kelsoe, G.; Haynes, B.F. Autoreactivity in an HIV-1 broadly reactive neutralizing antibody variable region heavy chain induces immunologic tolerance. Proc. Natl. Acad. Sci. USA 2010, 107, 181–186. [Google Scholar] [CrossRef]
- Pugh-Bernard, A.E.; Silverman, G.J.; Cappione, A.J.; Villano, M.E.; Ryan, D.H.; Insel, R.A.; Sanz, I. Regulation of inherently autoreactive VH4-34 B cells in the maintenance of human B cell tolerance. J. Clin. Investig. 2001, 108, 1061–1070. [Google Scholar] [CrossRef] [PubMed]
- Mehandru, S.; Poles, M.A.; Tenner-Racz, K.; Horowitz, A.; Hurley, A.; Hogan, C.; Boden, D.; Racz, P.; Markowitz, M. Primary HIV-1 Infection Is Associated with Preferential Depletion of CD4+ T Lymphocytes from Effector Sites in the Gastrointestinal Tract. J. Exp. Med. 2004, 200, 761–770. [Google Scholar] [CrossRef] [PubMed]
- Katlama, C.; Deeks, S.G.; Autran, B.; Martinez-Picado, J.; van Lunzen, J.; Rouzioux, C.; Miller, M.; Vella, S.; Schmitz, J.E.; Ahlers, J.; et al. Barriers to a cure for HIV: New ways to target and eradicate HIV-1 reservoirs. Lancet 2013, 381, 2109–2117. [Google Scholar] [CrossRef]
- Pavot, V.; Rochereau, N.; Lawrence, P.; Girard, M.P.; Genin, C.; Verrier, B.; Paul, S. Recent progress in HIV vaccines inducing mucosal immune responses. AIDS 2014, 28, 1701–1718. [Google Scholar] [CrossRef] [PubMed]
- Alexander, R.; Mestecky, J. Neutralizing Antibodies in Mucosal Secretions: IgG or IgA? Curr. HIV Res. 2007, 5, 588–593. [Google Scholar] [CrossRef] [PubMed]
- Watkins, J.D.; Sholukh, A.M.; Mukhtar, M.M.; Siddappa, N.B.; Lakhashe, S.K.; Kim, M.; Reinherz, E.L.; Gupta, S.; Forthal, D.N.; Sattentau, Q.J.; et al. Anti-HIV IgA isotypes. AIDS 2013, 27, F13–F20. [Google Scholar] [CrossRef] [PubMed]
- Shacklett, B.L. Mucosal immunity to HIV: A review of recent literature. Curr. Opin. HIV AIDS 2008, 3, 541–547. [Google Scholar] [CrossRef]
- Choi, R.Y.; Levinson, P.; Guthrie, B.L.; Lohman-Payne, B.; Bosire, R.; Liu, A.Y.; Hirbod, T.; Kiarie, J.; Overbaugh, J.; John-Stewart, G.; et al. Cervicovaginal HIV-1-neutralizing immunoglobulin A detected among HIV-1-exposed seronegative female partners in HIV-1-discordant couples. AIDS 2012, 26, 2155–2163. [Google Scholar] [CrossRef]
- Lin, S.W.; Cun, A.S.; Harris-McCoy, K.; Ertl, H.C. Intramuscular rather than oral administration of replication-defective adenoviral vaccine vector induces specific CD8+ T cell responses in the gut. Vaccine 2007, 25, 2187–2193. [Google Scholar] [CrossRef]
- Tatsis, N.; Lin, S.-W.; Harris-McCoy, K.; Garber, D.A.; Feinberg, M.B.; Ertl, H.C.J. Multiple immunizations with adenovirus and MVA vectors improve CD8+ T cell functionality and mucosal homing. Virology 2007, 367, 156–167. [Google Scholar] [CrossRef]
- Pavot, V.; Rochereau, N.; Genin, C.; Verrier, B.; Paul, S. New insights in mucosal vaccine development. Vaccine 2012, 30, 142–154. [Google Scholar] [CrossRef] [PubMed]
- Johansson, E.-L.; Bergquist, C.; Edebo, A.; Johansson, C.; Svennerholm, A.-M. Comparison of different routes of vaccination for eliciting antibody responses in the human stomach. Vaccine 2004, 22, 984–990. [Google Scholar] [CrossRef] [PubMed]
- Johansson, E.-L.; Wassén, L.; Holmgren, J.; Jertborn, M.; Rudin, A. Nasal and Vaginal Vaccinations Have Differential Effects on Antibody Responses in Vaginal and Cervical Secretions in Humans. Infect. Immun. 2001, 69, 7481–7486. [Google Scholar] [CrossRef] [PubMed]
- Jones, A.T.; Shen, X.; Walter, K.L.; LaBranche, C.C.; Wyatt, L.S.; Tomaras, G.D.; Montefiori, D.C.; Moss, B.; Barouch, D.H.; Clements, J.D.; et al. HIV-1 vaccination by needle-free oral injection induces strong mucosal immunity and protects against SHIV challenge. Nat. Commun. 2019, 10, 798. [Google Scholar] [CrossRef]
- Doan, L.X.; Li, M.; Chen, C.; Yao, Q. Virus-like particles as HIV-1 vaccines. Rev. Med. Virol. 2005, 15, 75–88. [Google Scholar] [CrossRef]
- Young, K.R.; McBurney, S.P.; Karkhanis, L.U.; Ross, T.M. Virus-like particles: Designing an effective AIDS vaccine. Methods 2006, 40, 98–117. [Google Scholar] [CrossRef]
- Stanley, M.; Pinto, L.A.; Trimble, C. Human Papillomavirus Vaccines—Immune Responses. Vaccine 2012, 30, F83–F87. [Google Scholar] [CrossRef]
- Landry, N.; Ward, B.J.; Trépanier, S.; Montomoli, E.; Dargis, M.; Lapini, G.; Vézina, L.-P. Preclinical and Clinical Development of Plant-Made Virus-Like Particle Vaccine against Avian H5N1 Influenza. PLoS ONE 2010, 5, e15559. [Google Scholar] [CrossRef]
- Mestecky, J.; Russell, M.W.; Elson, C.O. Perspectives on Mucosal Vaccines: Is Mucosal Tolerance a Barrier? J. Immunol. 2007, 179, 5633–5638. [Google Scholar] [CrossRef]
- Kang, S.; Hong, S.; Lee, Y.-K.; Cho, S. Oral Vaccine Delivery for Intestinal Immunity—Biological Basis, Barriers, Delivery System, and M Cell Targeting. Polymers 2018, 10, 948. [Google Scholar] [CrossRef]
- Ogra, P.L.; Faden, H.; Welliver, R.C. Vaccination Strategies for Mucosal Immune Responses. Clin. Microbiol. Rev. 2001, 14, 430–445. [Google Scholar] [CrossRef] [PubMed]
- Lavelle, E.C.; Ward, R.W. Mucosal vaccines—Fortifying the frontiers. Nat. Rev. Immunol. 2021. [Google Scholar] [CrossRef] [PubMed]
- Sanders, R.W.; Venturi, M.; Schiffner, L.; Kalyanaraman, R.; Katinger, H.; Lloyd, K.O.; Kwong, P.D.; Moore, J.P. The Mannose-Dependent Epitope for Neutralizing Antibody 2G12 on Human Immunodeficiency Virus Type 1 Glycoprotein gp120. J. Virol. 2002, 76, 7293–7305. [Google Scholar] [CrossRef] [PubMed]
- Karle, S.; Planque, S.; Nishiyama, Y.; Taguchi, H.; Zhou, Y.-X.; Salas, M.; Lake, D.; Thiagarajan, P.; Arnett, F.; Hanson, C.V.; et al. Cross-clade HIV-1 neutralization by an antibody fragment from a lupus phage display library. AIDS 2004, 18, 329–331. [Google Scholar] [CrossRef]
- Planque, S.; Salas, M.; Mitsuda, Y.; Sienczyk, M.; Escobar, M.A.; Mooney, J.P.; Morris, M.-K.; Nishiyama, Y.; Ghosh, D.; Kumar, A.; et al. Neutralization of genetically diverse HIV-1 strains by IgA antibodies to the gp120–CD4-binding site from long-term survivors of HIV infection. AIDS 2010, 24, 875–884. [Google Scholar] [CrossRef]
- Mu, Z.; Haynes, B.F.; Cain, D.W. HIV mRNA Vaccines—Progress and Future Paths. Vaccines 2021, 9, 134. [Google Scholar] [CrossRef]
- Pardi, N.; Hogan, M.J.; Naradikian, M.S.; Parkhouse, K.; Cain, D.W.; Jones, L.; Moody, M.A.; Verkerke, H.P.; Myles, A.; Willis, E.; et al. Nucleoside-modified mRNA vaccines induce potent T follicular helper and germinal center B cell responses. J. Exp. Med. 2018, 215, 1571–1588. [Google Scholar] [CrossRef]
- Cirelli, K.M.; Carnathan, D.G.; Nogal, B.; Martin, J.T.; Rodriguez, O.L.; Upadhyay, A.A.; Enemuo, C.A.; Gebru, E.H.; Choe, Y.; Viviano, F.; et al. Slow Delivery Immunization Enhances HIV Neutralizing Antibody and Germinal Center Responses via Modulation of Immunodominance. Cell 2020, 180, 206. [Google Scholar] [CrossRef]
- Moyer, T.J.; Zmolek, A.C.; Irvine, D.J. Beyond antigens and adjuvants: Formulating future vaccines. J. Clin. Investig. 2016, 126, 799–808. [Google Scholar] [CrossRef]
- Gause, K.T.; Wheatley, A.K.; Cui, J.; Yan, Y.; Kent, S.J.; Caruso, F. Immunological Principles Guiding the Rational Design of Particles for Vaccine Delivery. ACS Nano 2017, 11, 54–68. [Google Scholar] [CrossRef]
- López-Sagaseta, J.; Malito, E.; Rappuoli, R.; Bottomley, M.J. Self-assembling protein nanoparticles in the design of vaccines. Comput. Struct. Biotechnol. J. 2016, 14, 58–68. [Google Scholar] [CrossRef] [PubMed]
- Sliepen, K.; Ozorowski, G.; Burger, J.A.; van Montfort, T.; Stunnenberg, M.; LaBranche, C.; Montefiori, D.C.; Moore, J.P.; Ward, A.B.; Sanders, R.W. Presenting native-like HIV-1 envelope trimers on ferritin nanoparticles improves their immunogenicity. Retrovirology 2015, 12, 82. [Google Scholar] [CrossRef] [PubMed]
- He, L.; de Val, N.; Morris, C.D.; Vora, N.; Thinnes, T.C.; Kong, L.; Azadnia, P.; Sok, D.; Zhou, B.; Burton, D.R.; et al. Presenting native-like trimeric HIV-1 antigens with self-assembling nanoparticles. Nat. Commun. 2016, 7, 12041. [Google Scholar] [CrossRef] [PubMed]
- Pardi, N.; Hogan, M.J.; Weissman, D. Recent advances in mRNA vaccine technology. Curr. Opin. Immunol. 2020, 65, 14–20. [Google Scholar] [CrossRef]
- Zhang, P.; Gorman, J.; Geng, H.; Liu, Q.; Lin, Y.; Tsybovsky, Y.; Go, E.P.; Dey, B.; Andine, T.; Kwon, A.; et al. Interdomain Stabilization Impairs CD4 Binding and Improves Immunogenicity of the HIV-1 Envelope Trimer. Cell Host Microbe 2018, 23, 832–844.e6. [Google Scholar] [CrossRef]
- Henderson, R.; Lu, M.; Zhou, Y.; Mu, Z.; Parks, R.; Han, Q.; Hsu, A.L.; Carter, E.; Blanchard, S.C.; Edwards, R.J.; et al. Disruption of the HIV-1 Envelope allosteric network blocks CD4-induced rearrangements. Nat. Commun. 2020, 11, 520. [Google Scholar] [CrossRef]
- Do Kwon, Y.; Pancera, M.; Acharya, P.; Georgiev, I.S.; Crooks, E.T.; Gorman, J.; Joyce, M.G.; Guttman, M.; Ma, X.; Narpala, S.; et al. Crystal structure, conformational fixation and entry-related interactions of mature ligand-free HIV-1 Env. Nat. Struct. Mol. Biol. 2015, 22, 522–531. [Google Scholar] [CrossRef]
- Saunders, K.O.; Wiehe, K.; Tian, M.; Acharya, P.; Bradley, T.; Alam, S.M.; Go, E.P.; Scearce, R.; Sutherland, L.; Henderson, R.; et al. Targeted selection of HIV-specific antibody mutations by engineering B cell maturation. Science 2019, 366, eaay7199. [Google Scholar] [CrossRef] [PubMed]
- Khaitan, A.; Unutmaz, D. Revisiting Immune Exhaustion During HIV Infection. Curr. HIV/AIDS Rep. 2011, 8, 4–11. [Google Scholar] [CrossRef]
- Zajac, A.J.; Blattman, J.N.; Murali-Krishna, K.; Sourdive, D.J.D.; Suresh, M.; Altman, J.D.; Ahmed, R. Viral Immune Evasion Due to Persistence of Activated T Cells Without Effector Function. J. Exp. Med. 1998, 188, 2205–2213. [Google Scholar] [CrossRef]
- Urbani, S.; Amadei, B.; Tola, D.; Massari, M.; Schivazappa, S.; Missale, G.; Ferrari, C. PD-1 Expression in Acute Hepatitis C Virus (HCV) Infection Is Associated with HCV-Specific CD8 Exhaustion. J. Virol. 2006, 80, 11398–11403. [Google Scholar] [CrossRef] [PubMed]
- Radziewicz, H.; Ibegbu, C.C.; Fernandez, M.L.; Workowski, K.A.; Obideen, K.; Wehbi, M.; Hanson, H.L.; Steinberg, J.P.; Masopust, D.; Wherry, E.J.; et al. Liver-Infiltrating Lymphocytes in Chronic Human Hepatitis C Virus Infection Display an Exhausted Phenotype with High Levels of PD-1 and Low Levels of CD127 Expression. J. Virol. 2007, 81, 2545–2553. [Google Scholar] [CrossRef] [PubMed]
- Boni, C.; Fisicaro, P.; Valdatta, C.; Amadei, B.; Di Vincenzo, P.; Giuberti, T.; Laccabue, D.; Zerbini, A.; Cavalli, A.; Missale, G.; et al. Characterization of Hepatitis B Virus (HBV)-Specific T-Cell Dysfunction in Chronic HBV Infection. J. Virol. 2007, 81, 4215–4225. [Google Scholar] [CrossRef] [PubMed]
- Wherry, E.J.; Kurachi, M. Molecular and cellular insights into T cell exhaustion. Nat. Rev. Immunol. 2015, 15, 486–499. [Google Scholar] [CrossRef]
- Hashimoto, M.; Kamphorst, A.O.; Im, S.J.; Kissick, H.T.; Pillai, R.N.; Ramalingam, S.S.; Araki, K.; Ahmed, R. CD8 T Cell Exhaustion in Chronic Infection and Cancer: Opportunities for Interventions. Annu. Rev. Med. 2018, 69, 301–318. [Google Scholar] [CrossRef]
- Kahan, S.M.; Wherry, E.J.; Zajac, A.J. T cell exhaustion during persistent viral infections. Virology 2015, 479–480, 180–193. [Google Scholar] [CrossRef]
- Trautmann, L.; Janbazian, L.; Chomont, N.; Said, E.A.; Gimmig, S.; Bessette, B.; Boulassel, M.-R.; Delwart, E.; Sepulveda, H.; Balderas, R.S.; et al. Upregulation of PD-1 expression on HIV-specific CD8+ T cells leads to reversible immune dysfunction. Nat. Med. 2006, 12, 1198–1202. [Google Scholar] [CrossRef]
- Petrovas, C.; Casazza, J.P.; Brenchley, J.M.; Price, D.A.; Gostick, E.; Adams, W.C.; Precopio, M.L.; Schacker, T.; Roederer, M.; Douek, D.C.; et al. PD-1 is a regulator of virus-specific CD8+ T cell survival in HIV infection. J. Exp. Med. 2006, 203, 2281–2292. [Google Scholar] [CrossRef]
- Wherry, E.J.; Ha, S.-J.; Kaech, S.M.; Haining, W.N.; Sarkar, S.; Kalia, V.; Subramaniam, S.; Blattman, J.N.; Barber, D.L.; Ahmed, R. Molecular Signature of CD8+ T Cell Exhaustion during Chronic Viral Infection. Immunity 2007, 27, 670–684. [Google Scholar] [CrossRef]
- D’Souza, M.; Fontenot, A.P.; Mack, D.G.; Lozupone, C.; Dillon, S.; Meditz, A.; Wilson, C.C.; Connick, E.; Palmer, B.E. Programmed Death 1 Expression on HIV-Specific CD4 + T Cells Is Driven by Viral Replication and Associated with T Cell Dysfunction. J. Immunol. 2007, 179, 1979–1987. [Google Scholar] [CrossRef]
- Kaufmann, D.E.; Walker, B.D. PD-1 and CTLA-4 Inhibitory Cosignaling Pathways in HIV Infection and the Potential for Therapeutic Intervention. J. Immunol. 2009, 182, 5891–5897. [Google Scholar] [CrossRef] [PubMed]
- Chomont, N.; El-Far, M.; Ancuta, P.; Trautmann, L.; Procopio, F.A.; Yassine-Diab, B.; Boucher, G.; Boulassel, M.-R.; Ghattas, G.; Brenchley, J.M.; et al. HIV reservoir size and persistence are driven by T cell survival and homeostatic proliferation. Nat. Med. 2009, 15, 893–900. [Google Scholar] [CrossRef] [PubMed]
- Fromentin, R.; Bakeman, W.; Lawani, M.B.; Khoury, G.; Hartogensis, W.; DaFonseca, S.; Killian, M.; Epling, L.; Hoh, R.; Sinclair, E.; et al. CD4+ T Cells Expressing PD-1, TIGIT and LAG-3 Contribute to HIV Persistence during ART. PLoS Pathog. 2016, 12, e1005761. [Google Scholar] [CrossRef] [PubMed]
- Archin, N.M.; Sung, J.M.; Garrido, C.; Soriano-Sarabia, N.; Margolis, D.M. Eradicating HIV-1 infection: Seeking to clear a persistent pathogen. Nat. Rev. Microbiol. 2014, 12, 750–764. [Google Scholar] [CrossRef] [PubMed]
- Shan, L.; Deng, K.; Shroff, N.S.; Durand, C.M.; Rabi, S.A.; Yang, H.-C.; Zhang, H.; Margolick, J.B.; Blankson, J.N.; Siliciano, R.F. Stimulation of HIV-1-Specific Cytolytic T Lymphocytes Facilitates Elimination of Latent Viral Reservoir after Virus Reactivation. Immunity 2012, 36, 491–501. [Google Scholar] [CrossRef]
- Velu, V.; Titanji, K.; Zhu, B.; Husain, S.; Pladevega, A.; Lai, L.; Vanderford, T.H.; Chennareddi, L.; Silvestri, G.; Freeman, G.J.; et al. Enhancing SIV-specific immunity in vivo by PD-1 blockade. Nature 2009, 458, 206–210. [Google Scholar] [CrossRef]
- Cecchinato, V.; Tryniszewska, E.; Ma, Z.M.; Vaccari, M.; Boasso, A.; Tsai, W.-P.; Petrovas, C.; Fuchs, D.; Heraud, J.-M.; Venzon, D.; et al. Immune Activation Driven by CTLA-4 Blockade Augments Viral Replication at Mucosal Sites in Simian Immunodeficiency Virus Infection. J. Immunol. 2008, 180, 5439–5447. [Google Scholar] [CrossRef]
- Boyer, Z.; Palmer, S. Targeting Immune Checkpoint Molecules to Eliminate Latent HIV. Front. Immunol. 2018, 9. [Google Scholar] [CrossRef]
- Gay, C.L.; Bosch, R.J.; Ritz, J.; Hataye, J.M.; Aga, E.; Tressler, R.L.; Mason, S.W.; Hwang, C.K.; Grasela, D.M.; Ray, N.; et al. Clinical Trial of the Anti-PD-L1 Antibody BMS-936559 in HIV-1 Infected Participants on Suppressive Antiretroviral Therapy. J. Infect. Dis. 2017, 215, 1725–1733. [Google Scholar] [CrossRef]
- Wherry, E.J. T cell exhaustion. Nat. Immunol. 2011, 12, 492–499. [Google Scholar] [CrossRef]
- Dyavar Shetty, R.; Velu, V.; Titanji, K.; Bosinger, S.E.; Freeman, G.J.; Silvestri, G.; Amara, R.R. PD-1 blockade during chronic SIV infection reduces hyperimmune activation and microbial translocation in rhesus macaques. J. Clin. Investig. 2012, 122, 1712–1716. [Google Scholar] [CrossRef] [PubMed]
- Palmer, B.E.; Neff, C.P.; LeCureux, J.; Ehler, A.; DSouza, M.; Remling-Mulder, L.; Korman, A.J.; Fontenot, A.P.; Akkina, R. In Vivo Blockade of the PD-1 Receptor Suppresses HIV-1 Viral Loads and Improves CD4 + T Cell Levels in Humanized Mice. J. Immunol. 2013, 190, 211–219. [Google Scholar] [CrossRef] [PubMed]
- Kenmoe, S.; Bigna, J.J.; Fatawou Modiyingi, A.; Ndangang, M.S.; Ngoupo, P.A.; Simo, F.B.N.; Tchatchouang, S.; Temfack, E.; Njouom, R. Case fatality rate and viral aetiologies of acute respiratory tract infections in HIV positive and negative people in Africa: The VARIAFRICA-HIV systematic review and meta-analysis. J. Clin. Virol. 2019, 117, 96–102. [Google Scholar] [CrossRef] [PubMed]
- Neuzil, K.M.; Reed, G.W.; Mitchel, E.F., Jr.; Griffin, M.R. Influenza-Associated Morbidity and Mortality in Young and Middle-Aged Women. JAMA 1999, 281, 901–907. [Google Scholar] [CrossRef]
- Sheth, A.N.; Patel, P.; Peters, P.J. Influenza and HIV: Lessons from the 2009 H1N1 Influenza Pandemic. Curr. HIV/AIDS Rep. 2011, 8, 181–191. [Google Scholar] [CrossRef]
- Al-Omari, A.; Rabaan, A.A.; Salih, S.; Al-Tawfiq, J.A.; Memish, Z.A. MERS coronavirus outbreak: Implications for emerging viral infections. Diagn. Microbiol. Infect. Dis. 2019, 93, 265–285. [Google Scholar] [CrossRef]
- Moni, M.A.; Liò, P. Network-based analysis of comorbidities risk during an infection: SARS and HIV case studies. BMC Bioinform. 2014, 15, 333. [Google Scholar] [CrossRef]
- Edelman, E.J.; Aoun-Barakat, L.; Villanueva, M.; Friedland, G. Confronting Another Pandemic: Lessons from HIV can Inform Our COVID-19 Response. AIDS Behav. 2020, 24, 1977–1979. [Google Scholar] [CrossRef]
- Jiang, H.; Zhou, Y.; Tang, W. Maintaining HIV care during the COVID-19 pandemic. Lancet HIV 2020, 7, e308–e309. [Google Scholar] [CrossRef]
- Blanco, J.L.; Ambrosioni, J.; Garcia, F.; Martínez, E.; Soriano, A.; Mallolas, J.; Miro, J.M. COVID-19 in patients with HIV: Clinical case series. Lancet HIV 2020, 7, e314–e316. [Google Scholar] [CrossRef]
- Adepoju, P. Tuberculosis and HIV responses threatened by COVID-19. Lancet HIV 2020, 7, e319–e320. [Google Scholar] [CrossRef]
- Algarin, A.B.; Varas-Rodríguez, E.; Valdivia, C.; Fennie, K.P.; Larkey, L.; Hu, N.; Ibañez, G.E. Symptoms, Stress, and HIV-Related Care Among Older People Living with HIV During the COVID-19 Pandemic, Miami, Florida. AIDS Behav. 2020, 24, 2236–2238. [Google Scholar] [CrossRef]
- Harkness, A.; Behar-Zusman, V.; Safren, S.A. Understanding the Impact of COVID-19 on Latino Sexual Minority Men in a US HIV Hot Spot. AIDS Behav. 2020, 24, 2017–2023. [Google Scholar] [CrossRef] [PubMed]
- Amimo, F.; Lambert, B.; Magit, A. What does the COVID-19 pandemic mean for HIV, tuberculosis, and malaria control? Trop. Med. Health 2020, 48, 32. [Google Scholar] [CrossRef] [PubMed]
- Sun, S.; Hou, J.; Chen, Y.; Lu, Y.; Brown, L.; Operario, D. Challenges to HIV Care and Psychological Health During the COVID-19 Pandemic Among People Living with HIV in China. AIDS Behav. 2020, 24, 2764–2765. [Google Scholar] [CrossRef]
- Pinto, R.M.; Park, S. COVID-19 Pandemic Disrupts HIV Continuum of Care and Prevention: Implications for Research and Practice Concerning Community-Based Organizations and Frontline Providers. AIDS Behav. 2020, 24, 2486–2489. [Google Scholar] [CrossRef]
- Adadi, P.; Kanwugu, O.N. Living with HIV in the time of COVID-19: A glimpse of hope. J. Med. Virol. 2021, 93, 59–60. [Google Scholar] [CrossRef]
- Richardson, S.; Hirsch, J.S.; Narasimhan, M.; Crawford, J.M.; McGinn, T.; Davidson, K.W.; Barnaby, D.P.; Becker, L.B.; Chelico, J.D.; Cohen, S.L.; et al. Presenting Characteristics, Comorbidities, and Outcomes Among 5700 Patients Hospitalized With COVID-19 in the New York City Area. JAMA 2020, 323, 2052–2059. [Google Scholar] [CrossRef]
- Shalev, N.; Scherer, M.; LaSota, E.D.; Antoniou, P.; Yin, M.T.; Zucker, J.; Sobieszczyk, M.E. Clinical Characteristics and Outcomes in People Living With Human Immunodeficiency Virus Hospitalized for Coronavirus Disease 2019. Clin. Infect. Dis. 2020, 71, 2294–2297. [Google Scholar] [CrossRef]
- Okoh, A.K.; Bishburg, E.; Grinberg, S.; Nagarakanti, S. COVID-19 Pneumonia in Patients with HIV: A Case Series. JAIDS J. Acquir. Immune Defic. Syndr. 2020, 85, e4–e5. [Google Scholar] [CrossRef]
- Borobia, A.; Carcas, A.; Arnalich, F.; Álvarez-Sala, R.; Monserrat-Villatoro, J.; Quintana, M.; Figueira, J.; Torres Santos-Olmo, R.; García-Rodríguez, J.; Martín-Vega, A.; et al. A Cohort of Patients with COVID-19 in a Major Teaching Hospital in Europe. J. Clin. Med. 2020, 9, 1733. [Google Scholar] [CrossRef] [PubMed]
- Moher, D.; Liberati, A.; Tetzlaff, J.; Altman, D.G. PRISMA Group. Preferred Reporting Items for Systematic Reviews and Meta-Analyses: The PRISMA Statement. PLoS Med. 2009, 6, e1000097. [Google Scholar] [CrossRef] [PubMed]
- Cevik, M.; Kuppalli, K.; Kindrachuk, J.; Peiris, M. Virology, transmission, and pathogenesis of SARS-CoV-2. BMJ 2020, 371, m3862. [Google Scholar] [CrossRef]
- Sette, A.; Crotty, S. Adaptive immunity to SARS-CoV-2 and COVID-19. Cell 2021, 184, 861–880. [Google Scholar] [CrossRef]
- Choi, B.; Choudhary, M.C.; Regan, J.; Sparks, J.A.; Padera, R.F.; Qiu, X.; Solomon, I.H.; Kuo, H.-H.; Boucau, J.; Bowman, K.; et al. Persistence and Evolution of SARS-CoV-2 in an Immunocompromised Host. N. Engl. J. Med. 2020, 383, 2291–2293. [Google Scholar] [CrossRef]
- Kemp, S.A.; Collier, D.A.; Datir, R.P.; Ferreira, I.A.T.M.; Gayed, S.; Jahun, A.; Hosmillo, M.; Rees-Spear, C.; Mlcochova, P.; Lumb, I.U.; et al. SARS-CoV-2 evolution during treatment of chronic infection. Nature 2021, 592, 277–282. [Google Scholar] [CrossRef]
- Tegally, H.; Wilkinson, E.; Giovanetti, M.; Iranzadeh, A.; Fonseca, V.; Giandhari, J.; Doolabh, D.; Pillay, S.; San, E.J.; Msomi, N.; et al. Detection of a SARS-CoV-2 variant of concern in South Africa. Nature 2021, 592, 438–443. [Google Scholar] [CrossRef]
- Faria, N.R.; Mellan, T.A.; Whittaker, C.; Claro, I.M.; Candido, D.D.S.; Mishra, S.; Crispim, M.A.E.; Sales, F.C.S.; Hawryluk, I.; McCrone, J.T.; et al. Genomics and epidemiology of the P.1 SARS-CoV-2 lineage in Manaus, Brazil. Science 2021, 372, 815–821. [Google Scholar] [CrossRef]
- Cele, S.; Karim, F.; Lustig, G.; San, J.E.; Hermanus, T.; Tegally, H.; Snyman, J.; Moyo-Gwete, T.; Wilkinson, E.; Bernstein, M.; et al. SARS-CoV-2 prolonged infection during advanced HIV disease evolves extensive immune escape. Cell Host Microbe 2022, 30, 154–162.e5. [Google Scholar] [CrossRef]
- Tarhini, H.; Recoing, A.; Bridier-nahmias, A.; Rahi, M.; Lambert, C.; Martres, P.; Lucet, J.-C.; Rioux, C.; Bouzid, D.; Lebourgeois, S.; et al. Long-Term Severe Acute Respiratory Syndrome Coronavirus 2 (SARS-CoV-2) Infectiousness Among Three Immunocompromised Patients: From Prolonged Viral Shedding to SARS-CoV-2 Superinfection. J. Infect. Dis. 2021, 223, 1522–1527. [Google Scholar] [CrossRef]
- Sepulcri, C.; Dentone, C.; Mikulska, M.; Bruzzone, B.; Lai, A.; Fenoglio, D.; Bozzano, F.; Bergna, A.; Parodi, A.; Altosole, T.; et al. The Longest Persistence of Viable SARS-CoV-2 With Recurrence of Viremia and Relapsing Symptomatic COVID-19 in an Immunocompromised Patient—A Case Study. Open Forum Infect. Dis. 2021, 8, ofab217. [Google Scholar] [CrossRef] [PubMed]
- Riou, C.; du Bruyn, E.; Stek, C.; Daroowala, R.; Goliath, R.T.; Abrahams, F.; Said-Hartley, Q.; Allwood, B.W.; Hsiao, N.-Y.; Wilkinson, K.A.; et al. Relationship of SARS-CoV-2-specific CD4 response to COVID-19 severity and impact of HIV-1 and tuberculosis coinfection. J. Clin. Investig. 2021, 131, e149125. [Google Scholar] [CrossRef] [PubMed]
- Karim, F.; Gazy, I.; Cele, S.; Zungu, Y.; Krause, R.; Bernstein, M.; Khan, K.; Ganga, Y.; Rodel, H.; Mthabela, N.; et al. HIV status alters disease severity and immune cell responses in Beta variant SARS-CoV-2 infection wave. Elife 2021, 10. [Google Scholar] [CrossRef]
- Spinelli, M.A.; Lynch, K.L.; Yun, C.; Glidden, D.V.; Peluso, M.J.; Henrich, T.J.; Gandhi, M.; Brown, L.B. SARS-CoV-2 seroprevalence, and IgG concentration and pseudovirus neutralising antibody titres after infection, compared by HIV status: A matched case-control observational study. Lancet HIV 2021, 8, e334–e341. [Google Scholar] [CrossRef]
- Israelow, B.; Mao, T.; Klein, J.; Song, E.; Menasche, B.; Omer, S.B.; Iwasaki, A. Adaptive immune determinants of viral clearance and protection in mouse models of SARS-CoV-2. Sci. Immunol. 2021, 6, eabl4509. [Google Scholar] [CrossRef]
- Carmona, S.; Bor, J.; Nattey, C.; Maughan-Brown, B.; Maskew, M.; Fox, M.P.; Glencross, D.K.; Ford, N.; MacLeod, W.B. Persistent High Burden of Advanced HIV Disease Among Patients Seeking Care in South Africa’s National HIV Program: Data From a Nationwide Laboratory Cohort. Clin. Infect. Dis. 2018, 66, S111–S117. [Google Scholar] [CrossRef]
- Huerga, H.; Van Cutsem, G.; Ben Farhat, J.; Puren, A.; Bouhenia, M.; Wiesner, L.; Dlamini, L.; Maman, D.; Ellman, T.; Etard, J.-F. Progress towards the UNAIDS 90–90-90 goals by age and gender in a rural area of KwaZulu-Natal, South Africa: A household-based community cross-sectional survey. BMC Public Health 2018, 18, 303. [Google Scholar] [CrossRef]
- Madhi, S.; Koen, A.; Fairlie, L.; Cutland, C.; Baillie, V.; Padayachee, S.; Dheda, K.; Barnabas, S.; Bhorat, Q.E.; Briner, C.; et al. ChAdOx1 nCoV-19 (AZD1222) Vaccine in People Living with and without HIV; Research Square: Durham, NC, USA, 2021. [Google Scholar] [CrossRef]
- Jordan, R.E.; Adab, P.; Cheng, K.K. COVID-19: Risk factors for severe disease and death. BMJ 2020, 368, m1198. [Google Scholar] [CrossRef]
- Cooper, T.; Woodward, B.; Alom, S.; Harky, A. Coronavirus disease 2019 (COVID-19) outcomes in HIV/AIDS patients: A systematic review. HIV Med. 2020, 21, 567–577. [Google Scholar] [CrossRef]
- Huang, C.; Wang, Y.; Li, X.; Ren, L.; Zhao, J.; Hu, Y.; Zhang, L.; Fan, G.; Xu, J.; Gu, X.; et al. Clinical features of patients infected with 2019 novel coronavirus in Wuhan, China. Lancet 2020, 395, 497–506. [Google Scholar] [CrossRef]
- Xu, Z.; Shi, L.; Wang, Y.; Zhang, J.; Huang, L.; Zhang, C.; Liu, S.; Zhao, P.; Liu, H.; Zhu, L.; et al. Pathological findings of COVID-19 associated with acute respiratory distress syndrome. Lancet Respir. Med. 2020, 8, 420–422. [Google Scholar] [CrossRef]
- Qin, C.; Zhou, L.; Hu, Z.; Zhang, S.; Yang, S.; Tao, Y.; Xie, C.; Ma, K.; Shang, K.; Wang, W.; et al. Dysregulation of Immune Response in Patients With Coronavirus 2019 (COVID-19) in Wuhan, China. Clin. Infect. Dis. 2020, 71, 762–768. [Google Scholar] [CrossRef]
- Tan, M.; Liu, Y.; Zhou, R.; Deng, X.; Li, F.; Liang, K.; Shi, Y. Immunopathological characteristics of coronavirus disease 2019 cases in Guangzhou, China. Immunology 2020, 160, 261–268. [Google Scholar] [CrossRef] [PubMed]
- XU, B.; FAN, C.; WANG, A.; ZOU, Y.; YU, Y.; HE, C.; XIA, W.; ZHANG, J.; MIAO, Q. Suppressed T cell-mediated immunity in patients with COVID-19: A clinical retrospective study in Wuhan, China. J. Infect. 2020, 81, e51–e60. [Google Scholar] [CrossRef] [PubMed]
- Wan, S.; Yi, Q.; Fan, S.; Lv, J.; Zhang, X.; Guo, L.; Lang, C.; Xiao, Q.; Xiao, K.; Yi, Z.; et al. Relationships among lymphocyte subsets, cytokines, and the pulmonary inflammation index in coronavirus (COVID-19) infected patients. Br. J. Haematol. 2020, 189, 428–437. [Google Scholar] [CrossRef]
- Zhou, Y.; Zhang, Z.; Tian, J.; Xiong, S. Risk factors associated with disease progression in a cohort of patients infected with the 2019 novel coronavirus. Ann. Palliat. Med. 2020, 9, 428–436. [Google Scholar] [CrossRef]
- Chen, Y.; Klein, S.L.; Garibaldi, B.T.; Li, H.; Wu, C.; Osevala, N.M.; Li, T.; Margolick, J.B.; Pawelec, G.; Leng, S.X. Aging in COVID-19: Vulnerability, immunity and intervention. Ageing Res. Rev. 2021, 65, 101205. [Google Scholar] [CrossRef]
- Channappanavar, R.; Perlman, S. Pathogenic human coronavirus infections: Causes and consequences of cytokine storm and immunopathology. Semin. Immunopathol. 2017, 39, 529–539. [Google Scholar] [CrossRef]
- Lau, S.K.P.; Lau, C.C.Y.; Chan, K.-H.; Li, C.P.Y.; Chen, H.; Jin, D.-Y.; Chan, J.F.W.; Woo, P.C.Y.; Yuen, K.-Y. Delayed induction of proinflammatory cytokines and suppression of innate antiviral response by the novel Middle East respiratory syndrome coronavirus: Implications for pathogenesis and treatment. J. Gen. Virol. 2013, 94, 2679–2690. [Google Scholar] [CrossRef]
- Moore, J.B.; June, C.H. Cytokine release syndrome in severe COVID-19. Science 2020, 368, 473–474. [Google Scholar] [CrossRef]
- Guo, W.; Ming, F.; Dong, Y.; Zhang, Q.; Zhang, X.; Mo, P.; Feng, Y.; Liang, K. A Survey for COVID-19 Among HIV/AIDS Patients in Two Districts of Wuhan, China. SSRN Electron. J. 2020. [Google Scholar] [CrossRef]
- Minotti, C.; Tirelli, F.; Barbieri, E.; Giaquinto, C.; Donà, D. How is immunosuppressive status affecting children and adults in SARS-CoV-2 infection? A systematic review. J. Infect. 2020, 81, e61–e66. [Google Scholar] [CrossRef] [PubMed]
- SeyedAlinaghi, S.; Karimi, A.; MohsseniPour, M.; Barzegary, A.; Mirghaderi, S.P.; Fakhfouri, A.; Saeidi, S.; Razi, A.; Mojdeganlou, H.; Tantuoyir, M.M.; et al. The clinical outcomes of COVID-19 in HIV-positive patients: A systematic review of current evidence. Immunity, Inflamm. Dis. 2021, 9, 1160–1185. [Google Scholar] [CrossRef] [PubMed]
- Mascolo, S.; Romanelli, A.; Carleo, M.A.; Esposito, V. Could HIV infection alter the clinical course of SARS-CoV-2 infection? When less is better. J. Med. Virol. 2020, 92, 1777–1778. [Google Scholar] [CrossRef]
- Chen, X.P.; Cao, Y. Consideration of Highly Active Antiretroviral Therapy in the Prevention and Treatment of Severe Acute Respiratory Syndrome. Clin. Infect. Dis. 2004, 38, 1030–1032. [Google Scholar] [CrossRef]
- Choy, K.-T.; Wong, A.Y.-L.; Kaewpreedee, P.; Sia, S.F.; Chen, D.; Hui, K.P.Y.; Chu, D.K.W.; Chan, M.C.W.; Cheung, P.P.-H.; Huang, X.; et al. Remdesivir, lopinavir, emetine, and homoharringtonine inhibit SARS-CoV-2 replication in vitro. Antiviral Res. 2020, 178, 104786. [Google Scholar] [CrossRef]
- Elfiky, A.A. Ribavirin, Remdesivir, Sofosbuvir, Galidesivir, and Tenofovir against SARS-CoV-2 RNA dependent RNA polymerase (RdRp): A molecular docking study. Life Sci. 2020, 253, 117592. [Google Scholar] [CrossRef]
- Cao, B.; Wang, Y.; Wen, D.; Liu, W.; Wang, J.; Fan, G.; Ruan, L.; Song, B.; Cai, Y.; Wei, M.; et al. A Trial of Lopinavir–Ritonavir in Adults Hospitalized with Severe COVID-19. N. Engl. J. Med. 2020, 382, 1787–1799. [Google Scholar] [CrossRef] [PubMed]
- McMichael, A.J.; Borrow, P.; Tomaras, G.D.; Goonetilleke, N.; Haynes, B.F. The immune response during acute HIV-1 infection: Clues for vaccine development. Nat. Rev. Immunol. 2010, 10, 11–23. [Google Scholar] [CrossRef]
- Letko, M.; Marzi, A.; Munster, V. Functional assessment of cell entry and receptor usage for SARS-CoV-2 and other lineage B betacoronaviruses. Nat. Microbiol. 2020, 5, 562–569. [Google Scholar] [CrossRef]
- Wrapp, D.; Wang, N.; Corbett, K.S.; Goldsmith, J.A.; Hsieh, C.-L.; Abiona, O.; Graham, B.S.; McLellan, J.S. Cryo-EM structure of the 2019-nCoV spike in the prefusion conformation. Science 2020, 367, 1260–1263. [Google Scholar] [CrossRef] [PubMed]
- Walls, A.C.; Park, Y.-J.; Tortorici, M.A.; Wall, A.; McGuire, A.T.; Veesler, D. Structure, Function, and Antigenicity of the SARS-CoV-2 Spike Glycoprotein. Cell 2020, 181, 281–292.e6. [Google Scholar] [CrossRef] [PubMed]
- Guo, L.; Ren, L.; Yang, S.; Xiao, M.; Chang, D.; Yang, F.; Dela Cruz, C.S.; Wang, Y.; Wu, C.; Xiao, Y.; et al. Profiling Early Humoral Response to Diagnose Novel Coronavirus Disease (COVID-19). Clin. Infect. Dis. 2020, 71, 778–785. [Google Scholar] [CrossRef] [PubMed]
- Long, Q.-X.; Liu, B.-Z.; Deng, H.-J.; Wu, G.-C.; Deng, K.; Chen, Y.-K.; Liao, P.; Qiu, J.-F.; Lin, Y.; Cai, X.-F.; et al. Antibody responses to SARS-CoV-2 in patients with COVID-19. Nat. Med. 2020, 26, 845–848. [Google Scholar] [CrossRef]
- Perera, R.A.; Mok, C.K.; Tsang, O.T.; Lv, H.; Ko, R.L.; Wu, N.C.; Yuan, M.; Leung, W.S.; Chan, J.M.; Chik, T.S.; et al. Serological assays for severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2), March 2020. Eurosurveillance 2020, 25, 2000421. [Google Scholar] [CrossRef]




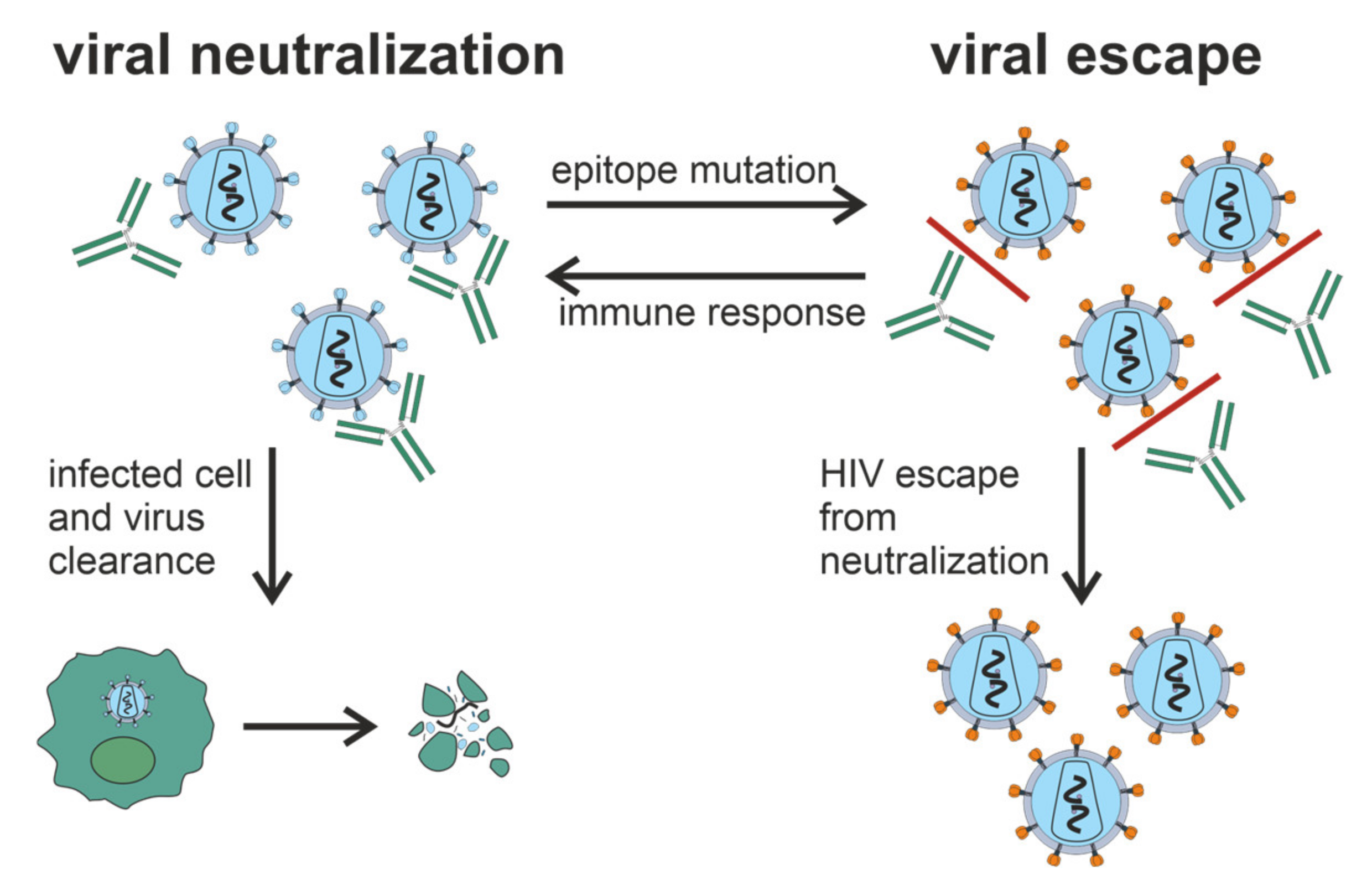{kind=link}
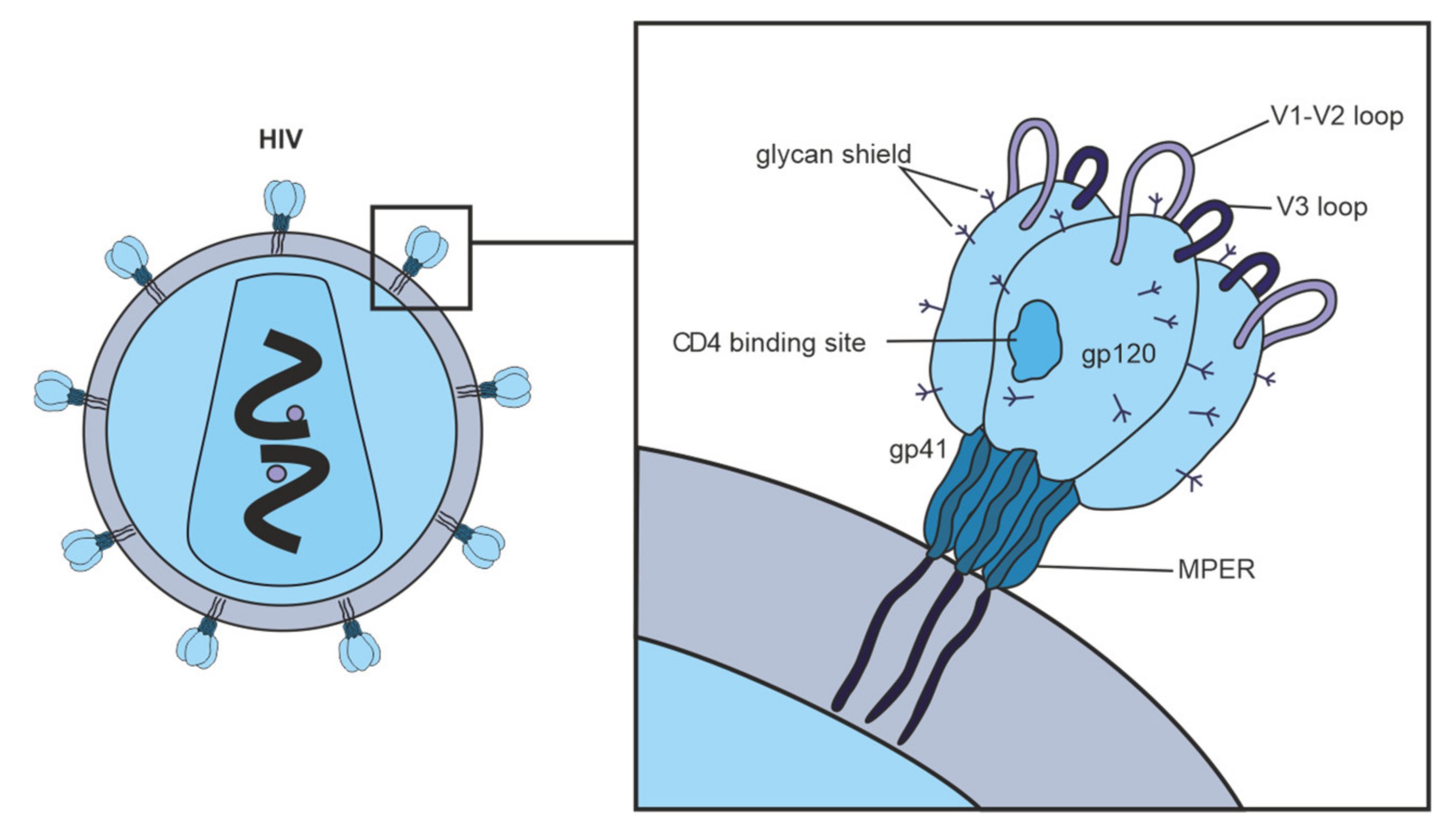{kind=link}
{kind=link}
| Specific Substrate | Type of Abs (Ab Fragment) | Ref |
|---|---|---|
| gp120 | L-chain, IgA | [136,137,138] |
| epitope 421–433 of gp120 | IgM, IgA, L-chain | [136,137,141] |
| HIV-1 integrase | IgG, IgM | [42,43] |
| HIV-1 integrase oligopeptides | IgG, IgM | [133,143] |
| HIV-1 reverse transcriptase | IgG, IgM | [39] |
| HIV-1 reverse transcriptase oligopeptides | IgG, IgM | [134] |
| β-casein | IgG | [39] |
| DNA | IgG, L-chain | [39,152] |
| histones | IgG | [40] |
| MBP | IgG | [41] |
| MBP oligopeptides | IgG | [144] |
| short non-specific oligopeptides | IgG | [133] |
| Title | Conditions | ID |
|---|---|---|
| HIV Testing & Womens Attitudes on HIV Vaccine Trials | HIV-1 | NCT00771537 |
| Efficacy and Safety of GSK Biologicals HIV Vaccine in Antiretroviral Therapy (ART)-naïve HIV Infected Persons | HIV-1 | NCT01218113 |
| Follow up of Thai Adult Volunteers With Breakthrough HIV Infection After Participation in a Preventive HIV Vaccine Trial | HIV-1 | NCT00337181 |
| Study to Optimize the Quality of Samples for Cell-mediated Immunity (CMI) in ART-naïve HIV-infected Subjects | HIV-1 | NCT01610427 |
| HIV Vaccine Trial in Thai Adults | HIV-1 | NCT00223080 |
| A Safety and Immune Response Study of 2 Experimental HIV Vaccines | HIV-1 | NCT02404311 |
| Safety and Immunogenicity of Clade C ALVAC and gp120 HIV Vaccine | HIV-1 | NCT03284710 |
| The Safety and Immunogenicity of a Potential HIV Vaccine | HIV-1 | NCT01966900 |
| Investigation of V520 in a HIV Vaccine Dose Refinement Study (V520-027) | HIV-1 | NCT00350623 |
| Investigation of V520 in an HIV Vaccine Proof-of-Concept Study (V520-023) | HIV-1 | NCT00095576 |
| Dose-ranging Study to Evaluate the Safety & Immunogenicity of a HIV Vaccine 732,461 in Healthy HIV Seronegative Volunteers | HIV-1 | NCT00434512 |
| Effectiveness of Two Hepatitis B Vaccines in HIV-negative Youths | Hepatitis B | NCT00107042 |
| A Pilot Study of a Dendritic Cell Vaccine in HIV-1 Infected Subjects | HIV-1 | NCT00833781 |
| HIV-1 Peptide Immunization of Individuals in West Africa to Prevent Disease | HIV-1 | NCT01141205 |
| Therapeutic Vaccine for HIV | HIV-1 | NCT01859325 |
| Safety and Immunogenicity of Anti-Pneumococcal Vaccines in HIV-Infected Pregnant Women | Pneumococcal | NCT02717494 |
| H1N1 Influenza Vaccine Immunogenicity in HIV-1 Infected Patients | HIV-1 | NCT01111162 |
| Impact of a Human Papilloma Virus (HPV) Vaccine in HIV-Infected Young Women | HIV-1 | NCT00710593 |
| Safety and Effectiveness of HIV-1 DNA Plasmid Vaccine and HIV-1 Recombinant Adenoviral Vector Vaccine in HIV-Uninfected, Circumcised Men and Male-to-Female (MTF) Transgender Persons Who Have Sex With Men | HIV-1 | NCT00865566 |
| A Study of Safety, Tolerability, and Immunogenicity of the MRKAd5 Gag/Pol/Nef Vaccine in Healthy Adults (V520-016) | HIV-1 | NCT00849680 |
| Improving Immunogenicity of Influenza Vaccine in HIV Infected Individuals | HIV-1 | NCT01262846 |
| Immunogenicity of Hepatitis B Vaccination in HIV-infected Adults | Hepatitis B | NCT03316807 |
| Immune Responses to Pneumococcal Vaccination Among HIV-infected Subjects | HIV-1 | NCT00706550 |
| Long-term Immunogenicity of the HIV gp120-NefTat/AS01B Vaccine (GSK SB732461) | HIV-1 | NCT03368053 |
| Early Versus Delayed BCG Vaccination of HIV-exposed Infants | HIV-1 | NCT00712530 |
| Combination Vaccination Before HIV Treatment Interruption | HIV-1 | NCT02062580 |
| Evaluating the Immunogenicity of the AIDSVAX B/E Vaccine and the MVA/HIV62B Vaccine in Healthy, HIV-1-Uninfected Adults Who Previously Received MVA/HIV62B in DNA/MVA or MVA/MVA Regimens in HVTN 205 | HIV-1 | NCT02852005 |
| Evaluation of Safety and Immunogenicity of a Human Papillomavirus (HPV) Vaccine in Human Immunodeficiency Virus (HIV) Infected Females | HPV-1 | NCT01031069 |
| Immunogenicity of Fluzone HD, A High Dose Influenza Vaccine, In Children With Cancer or HIV | HIV-1 | NCT01205581 |
| Shedding, Immunogenicity and Safety of Quadrivalent Live Intranasal Influenza Vaccine (QLAIV) in HIV-infected Children and Young Adults | HIV-1 | NCT02474901 |
| Live Zoster Vaccine in HIV-Infected Adults on Antiretroviral Therapy | Herpes Zoster | NCT00851786 |
| Safety of and Immune Response to an H1N1 Influenza Virus Vaccine in HIV Infected Children and Youth | H1N1 Influenza | NCT00992836 |
| Safety and Immune Response of a Rotavirus Vaccine in HIV-infected and Uninfected Children Born to HIV-infected Mothers | Rotavirus | NCT00880698 |
| Safety and Immunogenicity of 13-Valent Pneumococcal Conjugate Vaccine (13vPnC) in HIV-Infected Subjects 6 Years of Age or Older Who Are € to Pneumococcal Vaccine | Pneumococcal | NCT00962780 |
| Safety of and Immune Response to an H1N1 Influenza Vaccine in HIV Infected Pregnant Women | H1N1 Influenza | NCT00992017 |
| Immunization With HIV-1 Peptides in Adjuvant for Treatment of Patients With Chronic HIV-infection | HIV-1 | NCT01009762 |
| Safety and Immunogenicity of IMVAMUNE (MVA-BN) Smallpox Vaccine in HIV Infected Patients | HIV-1 | NCT00316589 |
| Autologous Dendritic Cell Vaccine in HIV1 Infection | HIV-1 | NCT00510497 |
| Quadrivalent HPV Vaccine to Prevent Anal HPV in HIV-infected Men and Women | HIV-1 | NCT01461096 |
| Safety of and Immune Response to the Human Papillomavirus (HPV) Vaccine in HIV-Infected Women | HPV-1 | NCT00604175 |
| Comparison of Three Hepatitis B Vaccination Regimens in HIV-Positive Youth | Hepatitis B | NCT00106964 |
| Safety of and Immune Response to a Novel Human Papillomavirus Vaccine in HIV Infected Children | HPV | NCT00339040 |
| Safety of and Immune Response to a Meningitis Vaccine in HIV Infected Children and Youth | Meningitis | NCT00459316 |
| Safety and Efficacy of Romidepsin and the Therapeutic Vaccine Vacc-4x for Reduction of the Latent HIV-1 Reservoir | HIV-I | NCT02092116 |
| Safety and Immunogenicity of GlaxoSmithKline Biologicals’ HPV Vaccine 580,299 (Cervarix) in HIV Infected Females | HPV | NCT00586339 |
| The Effect of HPV Vaccination on Recurrence Rates in HIV Patients With Condylomata | HPV | NCT00941889 |
| Immune Response After Booster Vaccination in HIV-Infected Patients Who Received Rabies Primary Vaccination | Rabies | NCT01286493 |
| Safety and Immunogenicity of a Candidate Tuberculosis (TB) Vaccine in HIV-positive Adults. | Tuberculosis | NCT00707967 |
| Safety & Immunogenicity of 13vPnC in HIV-Infected Subjects Aged 18 or Older Who Were Previously Immunized With 23vPS | Pneumococcal | NCT00963235 |
| Study to Evaluate GSK Biologicals’ Herpes Zoster Vaccine GSK1437173A in Human Immunodeficiency Virus HIV Infected Subjects | Herpes Zoster | NCT01165203 |
| Vaccine Therapy in Preventing HPV in HIV-Positive Women in India | HPV | NCT00667563 |
| Evaluation of the Immune Response of a HIV Candidate Vaccine After Administration of One Chloroquine Dose | HIV-1 | NCT00972725 |
| Evaluating the Safety and Immunogenicity of HIV Clade C DNA Vaccine and MF59- or AS01B-Adjuvanted Clade C Env Protein Vaccines in Various Combinations in Healthy, HIV-Uninfected Adults | HIV-1 | NCT02915016 |
| Safety and Immunogenicity of GSK Biologicals’ Investigational Malaria Vaccine in HIV Infected Infants and Children | Malaria | NCT01148459 |
| Magnitude of the Antibody Response to and Safety of a GBS Trivalent Vaccine in HIV Positive and HIV Negative Pregnant Women and Their Offsprings | Streptococcal | NCT01412801 |
| Vaccine Therapy in Preventing Human Papillomavirus Infection in Young HIV-Positive Male Patients Who Have Sex With Males | HPV | NCT01209325 |
| Primary and Booster Vaccination Study With a Pneumococcal Vaccine in HIV Infected, HIV Exposed Uninfected and HIV Uninfected Children 6 to 10 Weeks of Age. | Pneumococcal | NCT00829010 |
| Intradermal Versus Intramuscular Polio Vaccine Booster in HIV Infected Subjects | Polio | NCT01686503 |
| Human Papillomavirus Vaccine Therapy in Treating Men With HIV-1 Infection | HPV | NCT00513526 |
| MRKAd5 HIV-1 Gag Vaccine (V520) in Subjects With Chronic Hepatitis C (V520-022) | Hepatitis C | NCT00857311 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Timofeeva, A.; Sedykh, S.; Nevinsky, G. Post-Immune Antibodies in HIV-1 Infection in the Context of Vaccine Development: A Variety of Biological Functions and Catalytic Activities. Vaccines 2022, 10, 384. https://doi.org/10.3390/vaccines10030384
Timofeeva A, Sedykh S, Nevinsky G. Post-Immune Antibodies in HIV-1 Infection in the Context of Vaccine Development: A Variety of Biological Functions and Catalytic Activities. Vaccines. 2022; 10(3):384. https://doi.org/10.3390/vaccines10030384
Chicago/Turabian StyleTimofeeva, Anna, Sergey Sedykh, and Georgy Nevinsky. 2022. "Post-Immune Antibodies in HIV-1 Infection in the Context of Vaccine Development: A Variety of Biological Functions and Catalytic Activities" Vaccines 10, no. 3: 384. https://doi.org/10.3390/vaccines10030384
APA StyleTimofeeva, A., Sedykh, S., & Nevinsky, G. (2022). Post-Immune Antibodies in HIV-1 Infection in the Context of Vaccine Development: A Variety of Biological Functions and Catalytic Activities. Vaccines, 10(3), 384. https://doi.org/10.3390/vaccines10030384