
Nanoparticle-Based Delivery Systems for Vaccines

 ,
,  , , , and
, , , and 
Abstract
1. Introduction
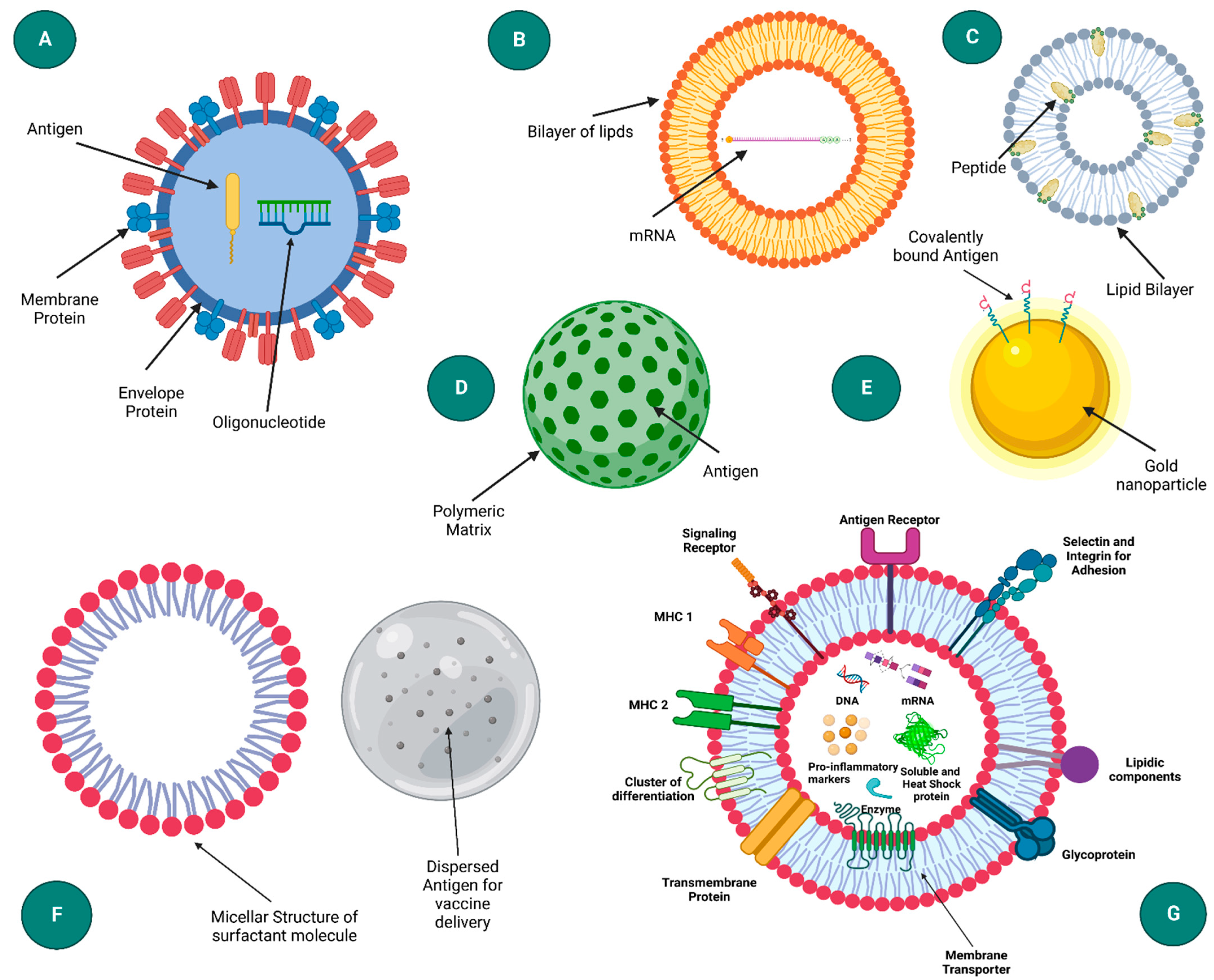
2. Nanoparticles and Nanovaccine Strategy
2.1. Virus-like Particles (VLPs) and Virosomes
2.2. Liposomes
2.3. Immunostimulatory Complexes (ISCOM)
2.4. Polymeric Nanoparticles
2.5. Inorganic Nanoparticles
2.6. Emulsion
2.7. Nanogels
2.8. Lipid Nanoparticles
3. Characterization of Nanoparticles
4. Exosome-Based Vaccine Delivery System
4.1. Exosome-Based Vaccine Delivery System for Viral Disease
4.2. Exosome-Based Treatment for Nonviral Disease
4.3. Exosome-Based Vaccines in Clinical Trials
4.4. Exosome-Based Vaccine against SARS-CoV-2
5. Nanoparticle-Based Nucleic Acid Vaccine
6. Nanoparticle Uptake and Immunity
7. Nanoparticle Vaccine Safety and Regulatory Concerns

{kind=link}
{kind=link}
| Biological | Condition | Phase | Sponsor | Reference |
|---|---|---|---|---|
| EBV Epstein-Barr Virus Infection Infectious Mononucleosis | Phase 1 | National Institute of Allergy and Infectious Diseases | [339] |
| Covid19 | Phase 3 | SK Bioscience Co., Ltd., Seongnam-si, Korea | [340] |
| COVID-19 (Healthy Volunteers) | Phase 1 Phase 2 | SK Bioscience Co., Ltd. | [341] |
| COVID-19 (Healthy Volunteers) | Phase 1 Phase 2 | SK Bioscience Co., Ltd. | [342] |
| Human Influenza | Phase 1 | Emergent BioSolutions | [343] |
| SARS-CoV-2 Infection | Phase 2 | Novavax | [344] |
| SARS-CoV-2 Infection | Phase 3 | Novavax | [345] |
| SARS-CoV Infection | Phase 3 | Novavax | [346] |
| SARS-CoV-2 Infection | Phase 1 | U.S. Army Medical Research and Development Command | [347] |
8. Cytotoxicity of Nanoparticles
9. Manufacturing Challenges
10. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Chavda, V.P.; Chen, Y.; Dave, J.; Chen, Z.-S.; Chauhan, S.C.; Yallapu, M.M.; Uversky, V.N.; Bezbaruah, R.; Patel, S.; Apostolopoulos, V. COVID-19 and Vaccination: Myths vs Science. Expert Rev. Vaccines 2022, 21, 1603–1620. [Google Scholar] [CrossRef] [PubMed]
- Fenner, F.; Henderson, D.A.; Arita, I.; Jezek, Z.; Ladnyi, I.D.; World Health Organization. Smallpox and Its Eradication; World Health Organization: Geneva, Switzerland, 1988. [Google Scholar]
- Hajj Hussein, I.; Chams, N.; Chams, S.; El Sayegh, S.; Badran, R.; Raad, M.; Gerges-Geagea, A.; Leone, A.; Jurjus, A. Vaccines Through Centuries: Major Cornerstones of Global Health. Front. Public Health 2015, 3, 269. [Google Scholar] [CrossRef] [PubMed]
- Pulendran, B.; Ahmed, R. Translating Innate Immunity into Immunological Memory: Implications for Vaccine Development. Cell 2006, 124, 849–863. [Google Scholar] [CrossRef] [PubMed]
- Biswas, P.; Wu, C.-Y. Nanoparticles and the Environment. J. Air Waste Manag. Assoc. 2005, 55, 708–746. [Google Scholar] [CrossRef] [PubMed]
- Chavda, V.P. Nanotherapeutics and Nanobiotechnology. In Applications of Targeted Nano Drugs and Delivery Systems; Elsevier: Amsterdam, The Netherlands, 2019; pp. 1–13. [Google Scholar]
- Chavda, V.P. Chapter 4—Nanobased Nano Drug Delivery: A Comprehensive Review. In Micro and Nano Technologies; Elsevier: Amsterdam, The Netherlands, 2019; pp. 69–92. [Google Scholar]
- Al-Halifa, S.; Gauthier, L.; Arpin, D.; Bourgault, S.; Archambault, D. Nanoparticle-Based Vaccines against Respiratory Viruses. Front. Immunol. 2019, 10, 22. [Google Scholar] [CrossRef]
- Angioletti-Uberti, S. Theory, Simulations and the Design of Functionalized Nanoparticles for Biomedical Applications: A Soft Matter Perspective. NPJ Comput. Mater. 2017, 3, 1–15. [Google Scholar] [CrossRef]
- Szeto, G.L.; Lavik, E.B. Materials Design at the Interface of Nanoparticles and Innate Immunity. J. Mater. Chem. B 2016, 4, 1610–1618. [Google Scholar] [CrossRef]
- Irvine, D.J.; Hanson, M.C.; Rakhra, K.; Tokatlian, T. Synthetic Nanoparticles for Vaccines and Immunotherapy. Chem. Rev. 2015, 115, 11109–11146. [Google Scholar] [CrossRef]
- Machhi, J.; Shahjin, F.; Das, S.; Patel, M.; Abdelmoaty, M.M.; Cohen, J.D.; Singh, P.A.; Baldi, A.; Bajwa, N.; Kumar, R.; et al. Nanocarrier Vaccines for SARS-CoV-2. Adv. Drug Deliv. Rev. 2021, 171, 215–239. [Google Scholar] [CrossRef]
- Chattopadhyay, S.; Chen, J.Y.; Chen, H.W.; Jack Hu, C.M. Nanoparticle Vaccines Adopting Virus-like Features for Enhanced Immune Potentiation. Nanotheranostics 2017, 1, 244–260. [Google Scholar] [CrossRef]
- Vora, L.K.; Moffatt, K.; Donnelly, R.F. 9—Long-Lasting Drug Delivery Systems Based on Microneedles. In Woodhead Publishing Series in Biomaterials; Woodhead Publishing: Amsterdam, The Netherlands, 2022; pp. 249–287. ISBN 978-0-12-821749-8. [Google Scholar]
- Gregory, A.E.; Titball, R.; Williamson, D. Vaccine Delivery Using Nanoparticles. Front. Cell. Infect. Microbiol. 2013, 3, 13. [Google Scholar] [CrossRef] [PubMed]
- Pachioni-Vasconcelos, J.D.A.; Lopes, A.M.; Apolinário, A.C.; Valenzuela-Oses, J.K.; Costa, J.S.R.; Nascimento, L.D.O.; Pessoa, A.; Barbosa, L.R.S.; Rangel-Yagui, C.D.O. Nanostructures for Protein Drug Delivery. Biomater. Sci. 2016, 4, 205–218. [Google Scholar] [CrossRef] [PubMed]
- Vora, L.K.; Moffatt, K.; Tekko, I.A.; Paredes, A.J.; Volpe-Zanutto, F.; Mishra, D.; Peng, K.; Raj Singh Thakur, R.; Donnelly, R.F. Microneedle Array Systems for Long-Acting Drug Delivery. Eur. J. Pharm. Biopharm. 2021, 159, 44–76. [Google Scholar] [CrossRef] [PubMed]
- Fredriksen, B.N.; Grip, J. PLGA/PLA Micro- and Nanoparticle Formulations Serve as Antigen Depots and Induce Elevated Humoral Responses after Immunization of Atlantic Salmon (Salmo Salar L.). Vaccine 2012, 30, 656–667. [Google Scholar] [CrossRef]
- Mamo, T.; Poland, G.A. Nanovaccinology: The next Generation of Vaccines Meets 21st Century Materials Science and Engineering. Vaccine 2012, 30, 6609–6611. [Google Scholar] [CrossRef]
- Zhu, M.; Wang, R.; Nie, G. Applications of Nanomaterials as Vaccine Adjuvants. Hum. Vaccines Immunother. 2014, 10, 2761–2774. [Google Scholar] [CrossRef]
- Ghiringhelli, F.; Apetoh, L.; Tesniere, A.; Aymeric, L.; Ma, Y.; Ortiz, C.; Vermaelen, K.; Panaretakis, T.; Mignot, G.; Ullrich, E.; et al. Activation of the NLRP3 Inflammasome in Dendritic Cells Induces IL-1β–Dependent Adaptive Immunity against Tumors. Nat. Med. 2009, 15, 1170–1178. [Google Scholar] [CrossRef]
- Tschopp, J.; Schroder, K. NLRP3 Inflammasome Activation: The Convergence of Multiple Signalling Pathways on ROS Production? Nat. Rev. Immunol. 2010, 10, 210–215. [Google Scholar] [CrossRef]
- Bruchard, M.; Mignot, G.; Derangère, V.; Chalmin, F.; Chevriaux, A.; Végran, F.; Boireau, W.; Simon, B.; Ryffel, B.; Connat, J.L.; et al. Chemotherapy-Triggered Cathepsin B Release in Myeloid-Derived Suppressor Cells Activates the Nlrp3 Inflammasome and Promotes Tumor Growth. Nat. Med. 2012, 19, 57–64. [Google Scholar] [CrossRef]
- Abderrazak, A.; Syrovets, T.; Couchie, D.; El Hadri, K.; Friguet, B.; Simmet, T.; Rouis, M. NLRP3 Inflammasome: From a Danger Signal Sensor to a Regulatory Node of Oxidative Stress and Inflammatory Diseases. Redox Biol. 2015, 4, 296–307. [Google Scholar] [CrossRef]
- He, Y.; Hara, H.; Núñez, G. Mechanism and Regulation of NLRP3 Inflammasome Activation. Trends Biochem. Sci. 2016, 41, 1012–1021. [Google Scholar] [CrossRef] [PubMed]
- Cassel, S.L.; Eisenbarth, S.C.; Iyer, S.S.; Sadler, J.J.; Colegio, O.R.; Tephly, L.A.; Carter, A.B.; Rothman, P.B.; Flavell, R.A.; Sutterwala, F.S. The Nalp3 Inflammasome Is Essential for the Development of Silicosis. Proc. Natl. Acad. Sci. USA 2008, 105, 9035–9040. [Google Scholar] [CrossRef] [PubMed]
- Scharf, B.; Clement, C.C.; Wu, X.X.; Morozova, K.; Zanolini, D.; Follenzi, A.; Larocca, J.N.; Levon, K.; Sutterwala, F.S.; Rand, J.; et al. Annexin A2 Binds to Endosomes Following Organelle Destabilization by Particulate Wear Debris. Nat. Commun. 2012, 3, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Niemi, K.; Teirilä, L.; Lappalainen, J.; Rajamäki, K.; Baumann, M.H.; Öörni, K.; Wolff, H.; Kovanen, P.T.; Matikainen, S.; Eklund, K.K. Serum Amyloid A Activates the NLRP3 Inflammasome via P2X7 Receptor and a Cathepsin B-Sensitive Pathway. J. Immunol. 2011, 186, 6119–6128. [Google Scholar] [CrossRef] [PubMed]
- Masters, S.L.; Dunne, A.; Subramanian, S.L.; Hull, R.L.; Tannahill, G.M.; Sharp, F.A.; Becker, C.; Franchi, L.; Yoshihara, E.; Chen, Z.; et al. Activation of the NLRP3 Inflammasome by Islet Amyloid Polypeptide Provides a Mechanism for Enhanced IL-1β in Type 2 Diabetes. Nat. Immunol. 2010, 11, 897–904. [Google Scholar] [CrossRef]
- Sharp, F.A.; Ruane, D.; Claass, B.; Creagh, E.; Harris, J.; Malyala, P.; Singh, M.; O’Hagan, D.T.; Pétrilli, V.; Tschopp, J.; et al. Uptake of Particulate Vaccine Adjuvants by Dendritic Cells Activates the NALP3 Inflammasome. Proc. Natl. Acad. Sci. USA 2009, 106, 870–875. [Google Scholar] [CrossRef]
- Halle, A.; Hornung, V.; Petzold, G.C.; Stewart, C.R.; Monks, B.G.; Reinheckel, T.; Fitzgerald, K.A.; Latz, E.; Moore, K.J.; Golenbock, D.T. The NALP3 Inflammasome Is Involved in the Innate Immune Response to Amyloid-β. Nat. Immunol. 2008, 9, 857–865. [Google Scholar] [CrossRef]
- Sheshala, R.; Madheswaran, T.; Panneerselvam, J.; Vora, L.; Thakur, R.R.S. Chapter 7—Stimuli-Responsive Nanomaterials in Infectious Diseases. In Developments in Microbiology; Academic Press: Cambridge, MA, USA, 2022; pp. 151–198. ISBN 978-0-323-91201-3. [Google Scholar]
- Laval, J.M.; Mazeran, P.E.; Thomas, D. Nanobiotechnology and Its Role in the Development of New Analytical Devices. Analyst 2000, 125, 29–33. [Google Scholar] [CrossRef]
- Schneider, C.S.; Xu, Q.; Boylan, N.J.; Chisholm, J.; Tang, B.C.; Schuster, B.S.; Henning, A.; Ensign, L.M.; Lee, E.; Adstamongkonkul, P.; et al. Nanoparticles That Do Not Adhere to Mucus Provide Uniform and Long-Lasting Drug Delivery to Airways Following Inhalation. Sci. Adv. 2017, 3, e1601556. [Google Scholar] [CrossRef]
- Parveen, K.; Banse, V.; Ledwani, L. Green Synthesis of Nanoparticles: Their Advantages and Disadvantages. In AIP Conference Proceedings; AIP Publishing LLC: Melville, NY, USA, 2016; Volume 1724, p. 020048. [Google Scholar] [CrossRef]
- Kheirollahpour, M.; Mehrabi, M.; Dounighi, N.M.; Mohammadi, M.; Masoudi, A. Nanoparticles and Vaccine Development. Pharm. Nanotechnol. 2019, 8, 6–21. [Google Scholar] [CrossRef]
- Chavda, V.P.; Yao, Q.; Vora, L.K.; Apostolopoulos, V.; Patel, C.A.; Bezbaruah, R.; Patel, A.B.; Chen, Z.-S. Fast-Track Development of Vaccines for SARS-CoV-2: The Shots That Saved the World. Front. Immunol. 2022, 13, 961198. [Google Scholar] [CrossRef] [PubMed]
- Gheibi Hayat, S.M.; Darroudi, M. Nanovaccine: A Novel Approach in Immunization. J. Cell. Physiol. 2019, 234, 12530–12536. [Google Scholar] [CrossRef] [PubMed]
- Cai, J.; Wang, H.; Wang, D.; Li, Y. Improving Cancer Vaccine Efficiency by Nanomedicine. Adv. Biosyst. 2019, 3, e1800287. [Google Scholar] [CrossRef] [PubMed]
- Mottram, P.L.; Leong, D.; Crimeen-Irwin, B.; Gloster, S.; Xiang, S.D.; Meanger, J.; Ghildyal, R.; Vardaxis, N.; Plebanski, M. Type 1 and 2 Immunity Following Vaccination Is Influenced by Nanoparticle Size: Formulation of a Model Vaccine for Respiratory Syncytial Virus. Mol. Pharm. 2006, 4, 73–84. [Google Scholar] [CrossRef] [PubMed]
- Karewicz, A. Polymeric and Liposomal Nanocarriers for Controlled Drug Delivery. In Biomaterials for Bone Regeneration; Elsevier: Amsterdam, The Netherlands, 2014; pp. 351–373. [Google Scholar] [CrossRef]
- Chavda, V.P.; Solanki, H.K.; Davidson, M.; Bojarska, J.; Apostolopoulos, V. Peptide-Drug Conjugates: A New Hope for Cancer Management. Molecules 2022, 27, 7232. [Google Scholar] [CrossRef]
- Nooraei, S.; Bahrulolum, H.; Hoseini, Z.S.; Katalani, C.; Hajizade, A.; Easton, A.J.; Ahmadian, G. Virus-like Particles: Preparation, Immunogenicity and Their Roles as Nanovaccines and Drug Nanocarriers. J. Nanobiotechnol. 2021, 19, 1–27. [Google Scholar] [CrossRef]
- Wang, N.; Chen, M.; Wang, T. Liposomes Used as a Vaccine Adjuvant-Delivery System: From Basics to Clinical Immunization. J. Control. Release 2019, 303, 130–150. [Google Scholar] [CrossRef]
- Sjölander, A.; Cox, J.C.; Barr, I.G. ISCOMs: An Adjuvant with Multiple Functions. J. Leukoc. Biol. 1998, 64, 713–723. [Google Scholar] [CrossRef]
- Guo, S.; Fu, D.; Utupova, A.; Sun, D.; Zhou, M.; Jin, Z.; Zhao, K. Applications of Polymer-Based Nanoparticles in Vaccine Field. Nanotechnol. Rev. 2019, 8, 143–155. [Google Scholar] [CrossRef]
- Zhang, Y.; Lin, S.; Wang, X.; Zhu, G. Nanovaccines for Cancer Immunotherapy. Wiley Interdiscip. Rev. Nanomed. Nanobiotechnol. 2019, 11, e1559. [Google Scholar] [CrossRef]
- Shah, R.R.; Brito, L.A.; O’Hagan, D.T.; Amiji, M.M. Emulsions as Vaccine Adjuvants BT—Subunit Vaccine Delivery; Springer: New York, NY, USA, 2015; pp. 59–76. ISBN 978-1-4939-1417-3. [Google Scholar]
- Hou, X.; Zaks, T.; Langer, R.; Dong, Y. Lipid Nanoparticles for MRNA Delivery. Nat. Rev. Mater. 2021, 6, 1078–1094. [Google Scholar] [CrossRef] [PubMed]
- Safety and Efficacy of Exosomes Overexpressing CD24 in Two Doses for Patients With Moderate or Severe COVID-19. Available online: https://www.clinicaltrials.gov/ct2/show/NCT04902183 (accessed on 25 September 2022).
- Tian, Y.; Wang, H.; Liu, Y.; Mao, L.; Chen, W.; Zhu, Z.; Liu, W.; Zheng, W.; Zhao, Y.; Kong, D.; et al. A Peptide-Based Nanofibrous Hydrogel as a Promising DNA Nanovector for Optimizing the Efficacy of HIV Vaccine. Nano Lett. 2014, 14, 1439–1445. [Google Scholar] [CrossRef]
- Touzé, A.; Gaitan, J.; Arnold, F.; Cazal, R.; Fleury, M.J.; Combelas, N.; Sizaret, P.Y.; Guyetant, S.; Maruani, A.; Baay, M.; et al. Generation of Merkel Cell Polyomavirus (MCV)-like Particles and Their Application to Detection of MCV Antibodies. J. Clin. Microbiol. 2010, 48, 1767–1770. [Google Scholar] [CrossRef] [PubMed]
- Kole, S.; Qadiri, S.S.N.; Shin, S.M.; Kim, W.S.; Lee, J.; Jung, S.J. PLGA Encapsulated Inactivated-Viral Vaccine: Formulation and Evaluation of Its Protective Efficacy against Viral Haemorrhagic Septicaemia Virus (VHSV) Infection in Olive Flounder (Paralichthys Olivaceus) Vaccinated by Mucosal Delivery Routes. Vaccine 2019, 37, 973–983. [Google Scholar] [CrossRef] [PubMed]
- Tao, W.; Ziemer, K.S.; Gill, H.S. Gold Nanoparticle–M2e Conjugate Coformulated with CpG Induces Protective Immunity against Influenza A Virus. Nanomedicine 2014, 9, 237–251. [Google Scholar] [CrossRef] [PubMed]
- Thomas, C.; Rawat, A.; Hope-Weeks, L.; Ahsan, F. Aerosolized PLA and PLGA Nanoparticles Enhance Humoral, Mucosal and Cytokine Responses to Hepatitis B Vaccine. Mol. Pharm. 2010, 8, 405–415. [Google Scholar] [CrossRef] [PubMed]
- Li, P.; Luo, Z.; Liu, P.; Gao, N.; Zhang, Y.; Pan, H.; Liu, L.; Wang, C.; Cai, L.; Ma, Y. Bioreducible Alginate-Poly (Ethylenimine) Nanogels as an Antigen-Delivery System Robustly Enhance Vaccine-Elicited Humoral and Cellular Immune Responses. J. Control. Release 2013, 168, 271–279. [Google Scholar] [CrossRef] [PubMed]
- Olczak, P.; Roden, R.B.S. Progress in L2-Based Prophylactic Vaccine Development for Protection against Diverse Human Papillomavirus Genotypes and Associated Diseases. Vaccines 2020, 8, 568. [Google Scholar] [CrossRef]
- Machhi, J.; Shahjin, F.; Das, S.; Patel, M.; Abdelmoaty, M.M.; Cohen, J.D.; Singh, P.A.; Baldi, A.; Bajwa, N.; Kumar, R.; et al. A Role for Extracellular Vesicles in SARS-CoV-2 Therapeutics and Prevention. J. Neuroimmune Pharmacol. 2021, 16, 270–288. [Google Scholar] [CrossRef]
- Donaldson, B.; Lateef, Z.; Walker, G.F.; Young, S.L.; Ward, V.K. Virus-like Particle Vaccines: Immunology and Formulation for Clinical Translation. Expert Rev. Vaccines 2018, 17, 833–849. [Google Scholar] [CrossRef]
- Chavda, V.P.; Patel, A.B.; Vora, L.K.; Apostolopoulos, V.; Uhal, B.D. Dendritic Cell-Based Vaccine: The State-of-the-Art Vaccine Platform for COVID-19 Management. Expert Rev. Vaccines 2022, 21, 1395–1403. [Google Scholar] [CrossRef] [PubMed]
- Zeltins, A. Construction and Characterization of Virus-like Particles: A Review. Mol. Biotechnol. 2013, 53, 92–107. [Google Scholar] [CrossRef] [PubMed]
- Young, K.R.; McBurney, S.P.; Karkhanis, L.U.; Ross, T.M. Virus-like Particles: Designing an Effective AIDS Vaccine. Methods 2006, 40, 98–117. [Google Scholar] [CrossRef] [PubMed]
- Patel, K.G.; Swartz, J.R. Surface Functionalization of Virus-like Particles by Direct Conjugation Using Azide-Alkyne Click Chemistry. Bioconjugate Chem. 2011, 22, 376–387. [Google Scholar] [CrossRef]
- Jarvis, D.L.; Howe, D.; Aumiller, J.J. Novel Baculovirus Expression Vectors That Provide Sialylation of Recombinant Glycoproteins in Lepidopteran Insect Cells. J. Virol. 2001, 75, 6223–6227. [Google Scholar] [CrossRef]
- Harrison, R.L.; Jarvis, D.L. Protein N-Glycosylation in the Baculovirus-Insect Cell Expression System and Engineering of Insect Cells to Produce “Mammalianized” Recombinant Glycoproteins. Adv. Virus Res. 2006, 68, 159–191. [Google Scholar]
- Vassileva, A.; Chugh, D.A.; Swaminathan, S.; Khanna, N. Effect of Copy Number on the Expression Levels of Hepatitis B Surface Antigen in the Methylotrophic Yeast Pichia Pastoris. Protein Expr. Purif. 2001, 21, 71–80. [Google Scholar] [CrossRef]
- Janowicz, Z.A.; Melber, K.; Merckelbach, A.; Jacobs, E.; Harford, N.; Comberbach, M.; Hollenberg, C.P. Simultaneous Expression of the S and L Surface Antigens of Hepatitis B, and Formation of Mixed Particles in the Methylotrophic Yeast, Hansenula Polymorpha. Yeast 1991, 7, 431–443. [Google Scholar] [CrossRef]
- Michel, M.-L.; Pontisso, P.; Sobczak, E.; Malpiece, Y.; Streeck, R.E.; Tiollais, P. Synthesis in Animal Cells of Hepatitis B Surface Antigen Particles Carrying a Receptor for Polymerized Human Serum Albumin. Proc. Natl. Acad. Sci. USA 1984, 81, 7708–7712. [Google Scholar] [CrossRef]
- Miyanohara, A.; Toh-e, A.; Nozaki, C.; Hamada, F.; Ohtomo, N.; Matsubara, K. Expression of Hepatitis B Surface Antigen Gene in Yeast. Proc. Natl. Acad. Sci. USA 1983, 80, 1–5. [Google Scholar] [CrossRef]
- Valenzuela, P.; Medina, A.; Rutter, W.J.; Ammerer, G.; Hall, B.D. Synthesis and Assembly of Hepatitis B Virus Surface Antigen Particles in Yeast. Nature 1982, 298, 347–350. [Google Scholar] [CrossRef] [PubMed]
- Michel, M.-L.; Tiollais, P. Hepatitis B Vaccines: Protective Efficacy and Therapeutic Potential. Pathol. Biol. 2010, 58, 288–295. [Google Scholar] [CrossRef] [PubMed]
- Bouvard, V.; Baan, R.; Straif, K.; Grosse, Y.; Secretan, B.; El Ghissassi, F.; Benbrahim-Tallaa, L.; Guha, N.; Freeman, C.; Galichet, L. A Review of Human Carcinogens—Part B: Biological Agents. Lancet Oncol. 2009, 10, 321–322. [Google Scholar] [CrossRef]
- Roldão, A.; Mellado, M.C.M.; Castilho, L.R.; Carrondo, M.J.; Alves, P.M. Virus-like Particles in Vaccine Development. Expert Rev. Vaccines 2010, 9, 1149–1176. [Google Scholar] [CrossRef]
- Kuate, S.; Cinatl, J.; Doerr, H.W.; Überla, K. Exosomal Vaccines Containing the S Protein of the SARS Coronavirus Induce High Levels of Neutralizing Antibodies. Virology 2007, 362, 26–37. [Google Scholar] [CrossRef]
- Chavda, V.P.; Vuppu, S.; Mishra, T.; Kamaraj, S.; Patel, A.B.; Sharma, N.; Chen, Z.S. Recent Review of COVID-19 Management: Diagnosis, Treatment and Vaccination. Pharmacol. Rep. 2022, 1–29. [Google Scholar] [CrossRef]
- Jeyanathan, M.; Afkhami, S.; Smaill, F.; Miller, M.S.; Lichty, B.D.; Xing, Z. Immunological Considerations for COVID-19 Vaccine Strategies. Nat. Rev. Immunol. 2020, 20, 615–632. [Google Scholar] [CrossRef]
- Aziz, S.A.; Kaneez, S.F. Analysis of SARS-CoV-2 and Factors Predicting next Spillover of Its More Contagious Variant. Int. J. Endorsing Health Sci. Res. 2021, 9, 405–419. [Google Scholar] [CrossRef]
- Khan, S.; Belgamwar, A.; Yeole, P. Nanovaccines against Viral Infections: Current Trends and Future Prospects. Nanotechnol. Appl. Virol. 2022, 315–328. [Google Scholar] [CrossRef]
- TRANSVAC. TRANSVAC-2: A European Network of Vaccine Research and Development-EATRIS; TRANSVAC: Alfreton, UK, 2020. [Google Scholar]
- Moser, C.; Müller, M.; Kaeser, M.D.; Weydemann, U.; Amacker, M. Influenza Virosomes as Vaccine Adjuvant and Carrier System. Expert Rev. Vaccines 2013, 12, 779–791. [Google Scholar] [CrossRef]
- Genito, C.J.; Batty, C.J.; Bachelder, E.M.; Ainslie, K.M. Considerations for Size, Surface Charge, Polymer Degradation, Co-Delivery, and Manufacturability in the Development of Polymeric Particle Vaccines for Infectious Diseases. Adv. NanoBiomed Res. 2020, 1, 2000041. [Google Scholar] [CrossRef] [PubMed]
- Das, A.; Ali, N. Nanovaccine: An Emerging Strategy. Expert Rev. Vaccines 2021, 20, 1273–1290. [Google Scholar] [CrossRef] [PubMed]
- Monpara, J.; Kanthou, C.; Tozer, G.M.; Vavia, P.R. Rational Design of Cholesterol Derivative for Improved Stability of Paclitaxel Cationic Liposomes. Pharm. Res. 2018, 35, 90. [Google Scholar] [CrossRef]
- Chavda, V.P.; Vihol, D.; Mehta, B.; Shah, D.; Patel, M.; Vora, L.K.; Pereira-Silva, M.; Paiva-Santos, A.C. Phytochemical-Loaded Liposomes for Anticancer Therapy: An Updated Review. Nanomedicine 2022, 17, 547–568. [Google Scholar] [CrossRef]
- Giddam, A.K.; Zaman, M.; Skwarczynski, M.; Toth, I. Liposome-Based Delivery System for Vaccine Candidates: Constructing an Effective Formulation. Nanomedicine 2012, 7, 1877–1893. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Ding, S.; Li, Y. Study on the Immunity Protection of 14-3-3–MPLA–Liposome Vaccine against Cystic Echinococcosis in Mice. Front. Lab. Med. 2018, 2, 146–151. [Google Scholar] [CrossRef]
- Nisini, R.; Poerio, N.; Mariotti, S.; De Santis, F.; Fraziano, M. The Multirole of Liposomes in Therapy and Prevention of Infectious Diseases. Release 2019, 303, 130–150. [Google Scholar] [CrossRef]
- Bhowmick, S.; Mazumdar, T.; Sinha, R.; Ali, N. Comparison of Liposome Based Antigen Delivery Systems for Protection against Leishmania Donovani. J. Control. Release 2010, 141, 199–207. [Google Scholar] [CrossRef]
- De Serrano, L.O.; Burkhart, D.J. Liposomal Vaccine Formulations as Prophylactic Agents: Design Considerations for Modern Vaccines. J. Nanobiotechnol. 2017, 15, 83. [Google Scholar] [CrossRef]
- Wui, S.R.; Ko, A.; Ryu, J.I.; Sim, E.; Lim, S.J.; Park, S.A.; Kim, K.S.; Kim, H.; Youn, H.; Lee, N.G. The Effect of a Tlr4 Agonist/Cationic Liposome Adjuvant on Varicella-zoster Virus Glycoprotein e Vaccine Efficacy: Antigen Presentation, Uptake and Delivery to Lymph Nodes. Pharmaceutics 2021, 13, 390. [Google Scholar] [CrossRef]
- Fu, Y.; Saraswat, A.; Vartak, R.; Patki, M.; Patel, K. Chapter 4—Liposomal Formulation: Opportunities, Challenges, and Industrial Applicability. In Micro and Nano Technologies; Elsevier: Amsterdam, The Netherlands, 2022; pp. 79–102. ISBN 978-0-323-85041-4. [Google Scholar]
- Bernasconi, V.; Norling, K.; Bally, M.; Höök, F.; Lycke, N.Y. Mucosal Vaccine Development Based on Liposome Technology. J. Immunol. Res. 2016, 2016, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Allison, A.C.; Gregoriadis, G. Liposomes as Immunological Adjuvants. Nature 1974, 252, 252. [Google Scholar] [CrossRef] [PubMed]
- Mai, Y.; Guo, J.; Zhao, Y.; Ma, S.; Hou, Y.; Yang, J. Intranasal Delivery of Cationic Liposome-Protamine Complex MRNA Vaccine Elicits Effective Anti-Tumor Immunity. Cell. Immunol. 2020, 354, 104143. [Google Scholar] [CrossRef]
- Wasan, E.K.; Syeda, J.; Strom, S.; Cawthray, J.; Hancock, R.E.; Wasan, K.M.; Gerdts, V. A Lipidic Delivery System of a Triple Vaccine Adjuvant Enhances Mucosal Immunity Following Nasal Administration in Mice. Vaccine 2019, 37, 1503–1515. [Google Scholar] [CrossRef] [PubMed]
- Espinosa, D.A.; Christensen, D.; Muñoz, C.; Singh, S.; Locke, E.; Andersen, P.; Zavala, F. Robust Antibody and CD8+ T-Cell Responses Induced by P. Falciparum CSP Adsorbed to Cationic Liposomal Adjuvant CAF09 Confer Sterilizing Immunity against Experimental Rodent Malaria Infection. NPJ Vaccines 2017, 2, 10. [Google Scholar] [CrossRef] [PubMed]
- Chavda, V.P.; Dawre, S.; Pandya, A.; Vora, L.K.; Modh, D.H.; Shah, V.; Dave, D.J.; Patravale, V. Lyotropic Liquid Crystals for Parenteral Drug Delivery. J. Control. Release 2022, 349, 533–549. [Google Scholar] [CrossRef]
- Chavda, V.P.; Vora, L.K.; Apostolopoulos, V. Inhalable Vaccines: Can They Help Control Pandemics? Vaccines 2022, 10, 1309. [Google Scholar] [CrossRef]
- Chavda, V.P.; Vora, L.K.; Pandya, A.K.; Patravale, V.B. Intranasal Vaccines for SARS-CoV-2: From Challenges to Potential in COVID-19 Management. Drug Discov. Today 2021, 26, 2619–2636. [Google Scholar] [CrossRef]
- Sun, H.; Xie, Y.; Ye, Y. ISCOMs and ISCOMATRIX. Vaccine 2009, 27, 4388–4401. [Google Scholar] [CrossRef]
- Junnuthula, V.; Kolimi, P.; Nyavanandi, D.; Sampathi, S.; Vora, L.K.; Dyawanapelly, S. Polymeric Micelles for Breast Cancer Therapy: Recent Updates, Clinical Translation and Regulatory Considerations. Pharmaceutics 2022, 14, 1860. [Google Scholar] [CrossRef]
- Morein, B.; Sundquist, B.; Höglund, S.; Dalsgaard, K.; Osterhaus, A. Iscom, a Novel Structure for Antigenic Presentation of Membrane Proteins from Enveloped Viruses. Nature 1984, 308, 457–460. [Google Scholar] [CrossRef] [PubMed]
- Alexyuk, P.G.; Bogoyavlenskiy, A.P.; Alexyuk, M.S.; Turmagambetova, A.S.; Zaitseva, I.A.; Omirtaeva, E.S.; Berezin, V. Adjuvant Activity of Multimolecular Complexes Based on Glycyrrhiza Glabra Saponins, Lipids, and Influenza Virus Glycoproteins. Arch. Virol. 2019, 164, 1793–1803. [Google Scholar] [CrossRef] [PubMed]
- Barr, I.G.; Sjölander, A.; Cox, J.C. ISCOMs and Other Saponin Based Adjuvants. Adv. Drug Deliv. Rev. 1998, 32, 247–271. [Google Scholar] [CrossRef]
- van Setten, D.C.; van de Werken, G. Molecular Structures of Saponins from Quillaja Saponaria Molina. Sapon. Used Tradit. Mod. Med. 1996, 185–193. [Google Scholar]
- Lovgren, K.; Morein, B. The Requirement of Lipids for the Formation of Immunostimulating Complexes (Iscoms). Biotechnol. Appl. Biochem. 1988, 10, 161–172. [Google Scholar] [CrossRef] [PubMed]
- Buglione-Corbett, R.; Pouliot, K.; Marty-Roix, R.; Li, W.; West, K.; Wang, S.; Morelli, A.B.; Lien, E.; Lu, S. Reduced MyD88 Dependency of ISCOMATRIXTM Adjuvant in a DNA Prime-Protein Boost HIV Vaccine. Hum. Vaccines Immunother. 2014, 10, 1078–1090. [Google Scholar] [CrossRef][Green Version]
- Peek, L.J.; Middaugh, C.R.; Berkland, C. Nanotechnology in Vaccine Delivery. Adv. Drug Deliv. Rev. 2008, 60, 915–928. [Google Scholar] [CrossRef]
- Pearse, M.J.; Drane, D. ISCOMATRIX® Adjuvant for Antigen Delivery. Adv. Drug Deliv. Rev. 2005, 57, 465–474. [Google Scholar] [CrossRef]
- Kersten, G.F.A.; Crommelin, D.J.A. Liposomes and ISCOMs. Vaccine 2003, 21, 915–920. [Google Scholar] [CrossRef]
- Cibulski, S.P.; Mourglia-Ettlin, G.; Teixeira, T.F.; Quirici, L.; Roehe, P.M.; Ferreira, F.; Silveira, F. Novel ISCOMs from Quillaja Brasiliensis Saponins Induce Mucosal and Systemic Antibody Production, T-Cell Responses and Improved Antigen Uptake. Vaccine 2016, 34, 1162–1171. [Google Scholar] [CrossRef]
- Shae, D.; Postma, A.; Wilson, J.T. Vaccine Delivery: Where Polymer Chemistry Meets Immunology. Ther. Deliv. 2016, 7, 193–196. [Google Scholar] [CrossRef] [PubMed]
- Chavda, V.P.; Jogi, G.; Shah, N.; Athalye, M.N.; Bamaniya, N.; Vora, L.K.; Cláudia Paiva-Santos, A. Advanced Particulate Carrier-Mediated Technologies for Nasal Drug Delivery. J. Drug Deliv. Sci. Technol. 2022, 74, 103569. [Google Scholar] [CrossRef]
- Chavda, V.P.; Patel, A.B.; Mistry, K.J.; Suthar, S.F.; Wu, Z.-X.; Chen, Z.-S.; Hou, K. Nano-Drug Delivery Systems Entrapping Natural Bioactive Compounds for Cancer: Recent Progress and Future Challenges. Front. Oncol. 2022, 12, 867655. [Google Scholar] [CrossRef]
- Akagi, T.; Baba, M.; Akashi, M. Biodegradable nanoparticles as vaccine adjuvants and delivery systems: Regulation of immune responses by nanoparticle-based vaccine. Adv. Polym. Sci. 2011, 247, 31–64. [Google Scholar] [CrossRef]
- Kim, H.; Griffith, T.S.; Panyam, J. Poly (d,l-Lactide-Co-Glycolide) Nanoparticles as Delivery Platforms for TLR7/8 Agonist-Based Cancer Vaccine. J. Pharmacol. Exp. Ther. 2019, 370, 715–724. [Google Scholar] [PubMed]
- Ekici, S.; Ilgin, P.; Butun, S.; Sahiner, N. Hyaluronic Acid Hydrogel Particles with Tunable Charges as Potential Drug Delivery Devices. Carbohydr. Polym. 2011, 84, 1306–1313. [Google Scholar] [CrossRef]
- Biswas, A.; Gomes, A.; Sengupta, J.; Datta, P.; Singha, S.; Dasgupta, A.K.; Gomes, A. Nanoparticle-Conjugated Animal Venom-Toxins and Their Possible Therapeutic Potential. J. Venom Res. 2012, 3, 15. [Google Scholar]
- Zolfagharian, H.; Damavandi, M.; Dounighi, N.M.; Moradi, S. Preparing and Characterizing Chitosan Nanoparticles Containing Hemiscorpius lepturus Scorpion Venom as an Antigen Delivery System. Arch. Razi Inst. 2012, 67, 145–153. [Google Scholar] [CrossRef]
- Mehrabi, M.; Dounighi, N.M.; Rezayat, S.M.; Doroud, D.; Amani, A.; Khoobi, M.; Ajdary, S. Novel Approach to Improve Vaccine Immunogenicity: Mannosylated Chitosan Nanoparticles Loaded with Recombinant Hepatitis B Antigen as a Targeted Vaccine Delivery System. J. Drug Deliv. Sci. Technol. 2018, 44, 19–26. [Google Scholar] [CrossRef]
- Feng, G.; Jiang, Q.; Xia, M.; Lu, Y.; Qiu, W.; Zhao, D.; Lu, L.; Peng, G.; Wang, Y. Enhanced Immune Response and Protective Effects of Nano-Chitosan-Based DNA Vaccine Encoding T Cell Epitopes of Esat-6 and FL against Mycobacterium Tuberculosis Infection. PLoS ONE 2013, 8, e61135. [Google Scholar] [CrossRef]
- Danhier, F.; Ansorena, E.; Silva, J.M.; Coco, R.; Le Breton, A.; Préat, V. PLGA-Based Nanoparticles: An Overview of Biomedical Applications. J. Control. Release 2012, 161, 505–522. [Google Scholar] [CrossRef] [PubMed]
- Panyam, J.; Labhasetwar, V. Biodegradable Nanoparticles for Drug and Gene Delivery to Cells and Tissue. Adv. Drug Deliv. Rev. 2003, 55, 329–347. [Google Scholar] [CrossRef]
- Lou, B.; De Beuckelaer, A.; Boonstra, E.; Li, D.; De Geest, B.G.; De Koker, S.; Mastrobattista, E.; Hennink, W.E. Modular Core-Shell Polymeric Nanoparticles Mimicking Viral Structures for Vaccination. J. Control. Release 2019, 293, 48–62. [Google Scholar] [CrossRef] [PubMed]
- Moon, J.J.; Suh, H.; Polhemus, M.E.; Ockenhouse, C.F.; Yadava, A.; Irvine, D.J. Antigen-Displaying Lipid-Enveloped PLGA Nanoparticles as Delivery Agents for a Plasmodium Vivax Malaria Vaccine. PLoS ONE 2012, 7, e31472. [Google Scholar] [CrossRef] [PubMed]
- Bilati, U.; Allémann, E.; Doelker, E. Development of a Nanoprecipitation Method Intended for the Entrapment of Hydrophilic Drugs into Nanoparticles. Eur. J. Pharm. Sci. 2005, 24, 67–75. [Google Scholar] [CrossRef]
- Boussif, O.; Lezoualc’h, F.; Zanta, M.A.; Mergny, M.D.; Scherman, D.; Demeneix, B.; Behr, J.-P. A Versatile Vector for Gene and Oligonucleotide Transfer into Cells in Culture and in Vivo: Polyethylenimine. Proc. Natl. Acad. Sci. USA 1995, 92, 7297–7301. [Google Scholar] [CrossRef]
- Shen, C.; Li, J.; Zhang, Y.; Li, Y.; Shen, G.; Zhu, J.; Tao, J. Polyethylenimine-Based Micro/Nanoparticles as Vaccine Adjuvants. Int. J. Nanomed. 2017, 12, 5443. [Google Scholar] [CrossRef]
- Bivas-Benita, M.; Lin, M.Y.; Bal, S.M.; van Meijgaarden, K.E.; Franken, K.L.M.C.; Friggen, A.H.; Junginger, H.E.; Borchard, G.; Klein, M.R.; Ottenhoff, T.H.M. Pulmonary Delivery of DNA Encoding Mycobacterium Tuberculosis Latency Antigen Rv1733c Associated to PLGA-PEI Nanoparticles Enhances T Cell Responses in a DNA Prime/Protein Boost Vaccination Regimen in Mice. Vaccine 2009, 27, 4010–4017. [Google Scholar] [CrossRef]
- Tanghe, A.; D’Souza, S.; Rosseels, V.; Denis, O.; Ottenhoff, T.H.M.; Dalemans, W.; Wheeler, C.; Huygen, K. Improved Immunogenicity and Protective Efficacy of a Tuberculosis DNA Vaccine Encoding Ag85 by Protein Boosting. Infect. Immun. 2001, 69, 3041–3047. [Google Scholar] [CrossRef]
- Niikura, K.; Matsunaga, T.; Suzuki, T.; Kobayashi, S.; Yamaguchi, H.; Orba, Y.; Kawaguchi, A.; Hasegawa, H.; Kajino, K.; Ninomiya, T. Gold Nanoparticles as a Vaccine Platform: Influence of Size and Shape on Immunological Responses in Vitro and in Vivo. ACS Nano 2013, 7, 3926–3938. [Google Scholar] [CrossRef]
- Yen, H.; Hsu, S.; Tsai, C. Cytotoxicity and Immunological Response of Gold and Silver Nanoparticles of Different Sizes. Small 2009, 5, 1553–1561. [Google Scholar] [CrossRef] [PubMed]
- Hutter, E.; Boridy, S.; Labrecque, S.; Lalancette-Hébert, M.; Kriz, J.; Winnik, F.M.; Maysinger, D. Microglial Response to Gold Nanoparticles. ACS Nano 2010, 4, 2595–2606. [Google Scholar] [CrossRef] [PubMed]
- Stone, J.W.; Thornburg, N.J.; Blum, D.L.; Kuhn, S.J.; Wright, D.W.; Crowe Jr, J.E. Gold Nanorod Vaccine for Respiratory Syncytial Virus. Nanotechnology 2013, 24, 295102. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.-S.; Hung, Y.-C.; Lin, W.-H.; Huang, G.S. Assessment of Gold Nanoparticles as a Size-Dependent Vaccine Carrier for Enhancing the Antibody Response against Synthetic Foot-and-Mouth Disease Virus Peptide. Nanotechnology 2010, 21, 195101. [Google Scholar] [CrossRef] [PubMed]
- Bianco, A.; Kostarelos, K.; Prato, M. Applications of Carbon Nanotubes in Drug Delivery. Curr. Opin. Chem. Biol. 2005, 9, 674–679. [Google Scholar] [CrossRef] [PubMed]
- Wang, T.; Zou, M.; Jiang, H.; Ji, Z.; Gao, P.; Cheng, G. Synthesis of a Novel Kind of Carbon Nanoparticle with Large Mesopores and Macropores and Its Application as an Oral Vaccine Adjuvant. Eur. J. Pharm. Sci. 2011, 44, 653–659. [Google Scholar] [CrossRef]
- Bachmann, M.F.; Jennings, G.T. Vaccine Delivery: A Matter of Size, Geometry, Kinetics and Molecular Patterns. Nat. Rev. Immunol. 2010, 10, 787–796. [Google Scholar] [CrossRef]
- Smart, S.K.; Cassady, A.I.; Lu, G.Q.; Martin, D.J. The Biocompatibility of Carbon Nanotubes. Carbon N. Y. 2006, 44, 1034–1047. [Google Scholar] [CrossRef]
- Ingle, S.G.; Pai, R.V.; Monpara, J.D.; Vavia, P.R. Liposils: An Effective Strategy for Stabilizing Paclitaxel Loaded Liposomes by Surface Coating with Silica. Eur. J. Pharm. Sci. 2018, 122, 51–63. [Google Scholar] [CrossRef]
- Ow, H.; Larson, D.R.; Srivastava, M.; Baird, B.A.; Webb, W.W.; Wiesner, U. Bright and Stable Core− Shell Fluorescent Silica Nanoparticles. Nano Lett. 2005, 5, 113–117. [Google Scholar] [CrossRef]
- Benezra, M.; Penate-Medina, O.; Zanzonico, P.B.; Schaer, D.; Ow, H.; Burns, A.; DeStanchina, E.; Longo, V.; Herz, E.; Iyer, S. Multimodal Silica Nanoparticles Are Effective Cancer-Targeted Probes in a Model of Human Melanoma. J. Clin. Investig. 2011, 121, 2768–2780. [Google Scholar] [CrossRef] [PubMed]
- Niu, Y.; Popat, A.; Yu, M.; Karmakar, S.; Gu, W.; Yu, C. Recent Advances in the Rational Design of Silica-Based Nanoparticles for Gene Therapy. Ther. Deliv. 2012, 3, 1217–1237. [Google Scholar] [CrossRef]
- He, Q.; Mitchell, A.R.; Johnson, S.L.; Wagner-Bartak, C.; Morcol, T.; Bell, S.J.D. Calcium Phosphate Nanoparticle Adjuvant. Clin. Diagn. Lab. Immunol. 2000, 7, 899–903. [Google Scholar] [CrossRef] [PubMed]
- He, Q.; Mitchell, A.; Morcol, T.; Bell, S.J.D. Calcium Phosphate Nanoparticles Induce Mucosal Immunity and Protection against Herpes Simplex Virus Type 2. Clin. Vaccine Immunol. 2002, 9, 1021–1024. [Google Scholar] [CrossRef]
- Joyappa, D.H.; Kumar, C.A.; Banumathi, N.; Reddy, G.R.; Suryanarayana, V.V.S. Calcium Phosphate Nanoparticle Prepared with Foot and Mouth Disease Virus P1-3CD Gene Construct Protects Mice and Guinea Pigs against the Challenge Virus. Vet. Microbiol. 2009, 139, 58–66. [Google Scholar] [CrossRef]
- Mody, K.T.; Popat, A.; Mahony, D.; Cavallaro, A.S.; Yu, C.; Mitter, N. Mesoporous Silica Nanoparticles as Antigen Carriers and Adjuvants for Vaccine Delivery. Nanoscale 2013, 5, 5167–5179. [Google Scholar] [CrossRef]
- Xu, L.; Liu, Y.; Chen, Z.; Li, W.; Liu, Y.; Wang, L.; Liu, Y.; Wu, X.; Ji, Y.; Zhao, Y. Surface-Engineered Gold Nanorods: Promising DNA Vaccine Adjuvant for HIV-1 Treatment. Nano Lett. 2012, 12, 2003–2012. [Google Scholar] [CrossRef]
- Wang, C.; Zhu, W.; Luo, Y.; Wang, B.-Z. Gold Nanoparticles Conjugating Recombinant Influenza Hemagglutinin Trimers and Flagellin Enhanced Mucosal Cellular Immunity. Nanomed. Nanotechnol. Biol. Med. 2018, 14, 1349–1360. [Google Scholar] [CrossRef]
- Dantas, G.P.F.; Ferraz, F.S.; Andrade, L.M.; Costa, G.M.J. Male Reproductive Toxicity of Inorganic Nanoparticles in Rodent Models: A Systematic Review. Chem. Biol. Interact. 2022, 363, 110023. [Google Scholar] [CrossRef]
- Chavda, V.P.; Shah, D. A Review on Novel Emulsification Technique: A Nanoemulsion. Res. Rev. J. Pharmacol. Toxicol. Stud. 2017, 5, 29–38. [Google Scholar]
- Chavda, V.P.; Shah, D. Self-Emulsifying Delivery Systems: One Step Ahead in Improving Solubility of Poorly Soluble Drugs. Nanostructures Cancer Ther. 2017, 653–718. [Google Scholar] [CrossRef]
- Chavda, V.P.; Shah, D. Chapter 25—Self-Emulsifying Delivery Systems: One Step Ahead in Improving Solubility of Poorly Soluble Drugs. In Micro and Nano Technologies; Elsevier: Amsterdam, The Netherlands, 2017; pp. 653–718. ISBN 978-0-323-46144-3. [Google Scholar]
- Chavda, V.P.; Shah, D.B.; Domadiya, K. Microemulsion: Novel Carrier for Drug Delivery. Trends Drug Deliv. 2016, 3, 1–18. [Google Scholar]
- O’Hagan, D.T. MF59 Is a Safe and Potent Vaccine Adjuvant That Enhances Protection against Influenza Virus Infection. Expert Rev. Vaccines 2007, 6, 699–710. [Google Scholar] [CrossRef]
- Nasiri, M.I.; Vora, L.K.; Juhaina, E.A.; Peng, K.; Tekko, I.A.; Donnelly, R.F. Nanoemulsion-Based Dissolving Microneedle Arrays for Enhanced Intradermal and Transdermal Delivery. Drug Deliv. Transl. Res. 2021, 12, 881–896. [Google Scholar] [CrossRef] [PubMed]
- Jain, J.; Fernandes, C.; Patravale, V. Formulation Development of Parenteral Phospholipid-Based Microemulsion of Etoposide. AAPS PharmSciTech 2010, 11, 826–831. [Google Scholar] [CrossRef] [PubMed]
- Chavda, V.; Soniwala, M.M.; Chavda, J.R. Lipid Based Delivery Systems: One Step Ahead in Improving Solubility Of Poorly Soluble Drugs. Inven. Rapid NDDS 2012. [Google Scholar]
- Lee, J.-J.; Shim, A.; Lee, S.Y.; Kwon, B.-E.; Kim, S.R.; Ko, H.-J.; Cho, H.-J. Ready-to-Use Colloidal Adjuvant Systems for Intranasal Immunization. J. Colloid Interface Sci. 2016, 467, 121–128. [Google Scholar] [CrossRef]
- Macedo, L.B.; Lobato, Z.I.P.; Fialho, S.L.; de Viott, A.M.; Guedes, R.M.C.; Silva-Cunha, A. Evaluation of Different Adjuvants Formulations for Bluetongue Vaccine. Braz. Arch. Biol. Technol. 2013, 56, 932–941. [Google Scholar] [CrossRef]
- Jiang, W.; Kim, B.Y.S.; Rutka, J.T.; Chan, W.C.W. Nanoparticle-Mediated Cellular Response Is Size-Dependent. Nat. Nanotechnol. 2008, 3, 145–150. [Google Scholar] [CrossRef]
- Reddy, S.T.; Rehor, A.; Schmoekel, H.G.; Hubbell, J.A.; Swartz, M.A. In Vivo Targeting of Dendritic Cells in Lymph Nodes with Poly(Propylene Sulfide) Nanoparticles. J. Control. Release 2006, 112, 26–34. [Google Scholar] [CrossRef]
- Vervarcke, S.; Ollevier, F.; Kinget, R.; Michoel, A. Oral Vaccination of African Catfish with Vibrio Anguillarum O2: Effect on Antigen Uptake and Immune Response by Absorption Enhancers in Lag Time Coated Pellets. Fish Shellfish Immunol. 2004, 16, 407–414. [Google Scholar] [CrossRef]
- Lamaisakul, S.; Tantituvanont, A.; Lipipun, V.; Ritthidej, G. Development of Novel Cationic Microemulsion as Parenteral Adjuvant for Influenza Vaccine. Asian J. Pharm. Sci. 2020, 15, 591–604. [Google Scholar] [CrossRef]
- Yang, A.; Yang, H.; Guo, S.; Xie, Y. MF59 Adjuvant Enhances the Immunogenicity and Protective Immunity of the OmpK/Omp22 Fusion Protein from Acineterbacter Baumannii through Intratracheal Inoculation in Mice. Scand. J. Immunol. 2019, 90, e12769. [Google Scholar] [CrossRef]
- Nakahashi-Ouchida, R.; Yuki, Y.; Kiyono, H. Development of a Nanogel-Based Nasal Vaccine as a Novel Antigen Delivery System. Expert Rev. Vaccines 2017, 16, 1231–1240. [Google Scholar] [CrossRef]
- Hubbell, J.A.; Thomas, S.N.; Swartz, M.A. Materials Engineering for Immunomodulation. Nature 2009, 462, 449–460. [Google Scholar] [CrossRef]
- Cone, R.A. Barrier Properties of Mucus. Adv. Drug Deliv. Rev. 2009, 61, 75–85. [Google Scholar] [CrossRef]
- Sharma, S.; Mukkur, T.K.S.; Benson, H.A.E.; Chen, Y. Pharmaceutical Aspects of Intranasal Delivery of Vaccines Using Particulate Systems. J. Pharm. Sci. 2009, 98, 812–843. [Google Scholar] [CrossRef]
- Owens, D., III; Peppas, N. Opsonization, Biodistribution, and Pharmacokinetics of Polymeric Nanoparticles. Int. J. Pharm. 2006, 307, 93–102. [Google Scholar] [CrossRef]
- Kabanov, A.V.; Vinogradov, S.V. Nanogels as Pharmaceutical Carriers: Finite Networks of Infinite Capabilities. Angew. Chem. Int. Ed. 2009, 48, 5418–5429. [Google Scholar] [CrossRef]
- Mishra, B.; Patel, B.B.; Tiwari, S. Colloidal Nanocarriers: A Review on Formulation Technology, Types and Applications toward Targeted Drug Delivery. Nanomed. Nanotechnol. Biol. Med. 2010, 6, 9–24. [Google Scholar] [CrossRef]
- Ferreira, S.A.; Gama, F.M.; Vilanova, M. Polymeric Nanogels as Vaccine Delivery Systems. Nanomed. Nanotechnol. Biol. Med. 2013, 9, 159–173. [Google Scholar] [CrossRef]
- Gao, C.; Lin, Z.; Jurado-Sánchez, B.; Lin, X.; Wu, Z.; He, Q. Stem Cell Membrane-coated Nanogels for Highly Efficient in Vivo Tumor Targeted Drug Delivery. Small 2016, 12, 4056–4062. [Google Scholar] [CrossRef]
- Harde, H.; Das, M.; Jain, S. Solid Lipid Nanoparticles: An Oral Bioavailability Enhancer Vehicle. Expert Opin. Drug Deliv. 2011, 8, 1407–1424. [Google Scholar] [CrossRef]
- Puri, A.; Loomis, K.; Smith, B.; Lee, J.-H.; Yavlovich, A.; Heldman, E.; Blumenthal, R. Lipid-Based Nanoparticles as Pharmaceutical Drug Carriers: From Concepts to Clinic. Crit. Rev. Ther. Drug Carr. Syst. 2009, 26. [Google Scholar] [CrossRef]
- Chavda, V.P.; Apostolopoulos, V. COVID-19 Vaccine Design and Vaccination Strategy for Emerging Variants. Expert Rev. Vaccines 2022, 21, 1359–1361. [Google Scholar] [CrossRef]
- Chavda, V.P.; Hossain, M.K.; Beladiya, J.; Apostolopoulos, V. Nucleic Acid Vaccines for COVID-19: A Paradigm Shift in the Vaccine Development Arena. Biologics 2021, 1, 337–356. [Google Scholar] [CrossRef]
- Das, S.; Chaudhury, A. Recent Advances in Lipid Nanoparticle Formulations with Solid Matrix for Oral Drug Delivery. Aaps Pharmscitech 2011, 12, 62–76. [Google Scholar] [CrossRef]
- O’Driscoll, C.M.; Griffin, B.T. Biopharmaceutical Challenges Associated with Drugs with Low Aqueous Solubility—the Potential Impact of Lipid-Based Formulations. Adv. Drug Deliv. Rev. 2008, 60, 617–624. [Google Scholar] [CrossRef]
- Porter, C.J.H.; Trevaskis, N.L.; Charman, W.N. Lipids and Lipid-Based Formulations: Optimizing the Oral Delivery of Lipophilic Drugs. Nat. Rev. Drug Discov. 2007, 6, 231–248. [Google Scholar] [CrossRef]
- Gursoy, R.N.; Benita, S. Self-Emulsifying Drug Delivery Systems (SEDDS) for Improved Oral Delivery of Lipophilic Drugs. Biomed. Pharmacother. 2004, 58, 173–182. [Google Scholar] [CrossRef]
- Patel, K.; Tyagi, M.; Monpara, J.; Vora, L.; Gupta, S.; Vavia, P. Arginoplexes: An Arginine-Anchored Nanoliposomal Carrier for Gene Delivery. J. Nanopart. Res. 2014, 16, 2345. [Google Scholar] [CrossRef]
- Ganesan, P.; Narayanasamy, D. Lipid Nanoparticles: Different Preparation Techniques, Characterization, Hurdles, and Strategies for the Production of Solid Lipid Nanoparticles and Nanostructured Lipid Carriers for Oral Drug Delivery. Sustain. Chem. Pharm. 2017, 6, 37–56. [Google Scholar] [CrossRef]
- Iqbal, M.A.; Md, S.; Sahni, J.K.; Baboota, S.; Dang, S.; Ali, J. Nanostructured Lipid Carriers System: Recent Advances in Drug Delivery. J. Drug Target. 2012, 20, 813–830. [Google Scholar] [CrossRef]
- Battaglia, L.; Gallarate, M. Lipid Nanoparticles: State of the Art, New Preparation Methods and Challenges in Drug Delivery. Expert Opin. Drug Deliv. 2012, 9, 497–508. [Google Scholar] [CrossRef]
- Kumar, S.; Dilbaghi, N.; Saharan, R.; Bhanjana, G. Nanotechnology as Emerging Tool for Enhancing Solubility of Poorly Water-Soluble Drugs. Bionanoscience 2012, 2, 227–250. [Google Scholar] [CrossRef]
- Pathak, K.; Raghuvanshi, S. Oral Bioavailability: Issues and Solutions via Nanoformulations. Clin. Pharmacokinet. 2015, 54, 325–357. [Google Scholar] [CrossRef]
- Schwarz, C.; Mehnert, W.; Lucks, J.S.; Müller, R.H. Solid Lipid Nanoparticles (SLN) for Controlled Drug Delivery. I. Production, Characterization and Sterilization. J. Control. Release 1994, 30, 83–96. [Google Scholar] [CrossRef]
- Westesen, K.; Bunjes, H.; Koch, M.H.J. Physicochemical Characterization of Lipid Nanoparticles and Evaluation of Their Drug Loading Capacity and Sustained Release Potential. J. Control. Release 1997, 48, 223–236. [Google Scholar] [CrossRef]
- Kaur, S.; Nautyal, U.; Singh, R.; Singh, S.; Devi, A. Nanostructure Lipid Carrier (NLC): The New Generation of Lipid Nanoparticles. Asian Pac. J. Health Sci. 2015, 2, 76–93. [Google Scholar] [CrossRef]
- Domingo, C.; Saurina, J. An Overview of the Analytical Characterization of Nanostructured Drug Delivery Systems: Towards Green and Sustainable Pharmaceuticals: A Review. Anal. Chim. Acta 2012, 744, 8–22. [Google Scholar] [CrossRef]
- Pardeshi, C.; Rajput, P.; Belgamwar, V.; Tekade, A.; Patil, G.; Chaudhary, K.; Sonje, A. Solid Lipid Based Nanocarriers: An Overview/Nanonosači Na Bazi Čvrstih Lipida: Pregled. Acta Pharm. 2012, 62, 433–472. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.-H.; Wu, C.-T. Optimization of Nanostructured Lipid Carriers for Lutein Delivery. Colloids Surf. A Physicochem. Eng. Asp. 2010, 353, 149–156. [Google Scholar] [CrossRef]
- Kisby, T.; Yilmazer, A.; Kostarelos, K. Reasons for Success and Lessons Learnt from Nanoscale Vaccines against COVID-19. Nat. Nanotechnol. 2021, 16, 843–850. [Google Scholar] [CrossRef] [PubMed]
- Gebre, M.S.; Brito, L.A.; Tostanoski, L.H.; Edwards, D.K.; Carfi, A.; Barouch, D.H. Novel Approaches for Vaccine Development. Cell 2021, 184, 1589–1603. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.; Eygeris, Y.; Gupta, M.; Sahay, G. Self-Assembled MRNA Vaccines. Adv. Drug Deliv. Rev. 2021, 170, 83–112. [Google Scholar] [CrossRef]
- Hassett, K.J.; Benenato, K.E.; Jacquinet, E.; Lee, A.; Woods, A.; Yuzhakov, O.; Himansu, S.; Deterling, J.; Geilich, B.M.; Ketova, T. Optimization of Lipid Nanoparticles for Intramuscular Administration of MRNA Vaccines. Mol. Ther. Acids 2019, 15, 1–11. [Google Scholar] [CrossRef]
- FDA. Drug Products, Including Biological Products, That Contain Nanomaterials-Guidance for Industry; U.S. Department of Health and Human Services: Washington, DC, USA, 2017.
- Kersten, G.F.; Teerlink, T.; Derks, H.J.; Verkleij, A.J.; Van Wezel, T.L.; Crommelin, D.J.; Beuvery, E.C. Incorporation of the Major Outer Membrane Protein of Neisseria Gonorrhoeae in Saponin-Lipid Complexes (Iscoms): Chemical Analysis, Some Structural Features, and Comparison of Their Immunogenicity with Three Other Antigen Delivery Systems. Infect. Immun. 1988, 56, 432–438. [Google Scholar] [CrossRef]
- Zhang, R.; Ulery, B.D. Synthetic Vaccine Characterization and Design. J. Bionanosci. 2018, 12, 1–11. [Google Scholar] [CrossRef]
- Carol, H.; Hernández, A.N.A.; Baz, A.; Nieto, A. Lack of Interspecies Barriers in Anti–Id Stimulated Antibody Production against Echinococcus Granulosus Antigens. Parasite Immunol. 1989, 11, 183–195. [Google Scholar] [CrossRef]
- Carol, H.; Nieto, A.; Villacres-Eriksson, M.; Morien, B. Intranasal Immunization of Mice with Echinococcus Granulosus Surface Antigens Iscoms Evokes a Strong Immune Response, Biased towards Glucidic Epitopes. Parasite Immunol. 1997, 19, 197–205. [Google Scholar]
- Erturk, M.; Jennings, R.; Phillpotts, R.J.; Potter, C.W. Biochemical Characterization of Herpes Simplex Virus Type-1-Immunostimulating Complexes (ISCOMs): A Multi-Glycoprotein Structure. Vaccine 1991, 9, 668–674. [Google Scholar] [CrossRef]
- Browning, M.; Reid, G.; Osborne, R.; Jarrett, O. Incorporation of Soluble Antigens into ISCOMs: HIV Gp120 ISCOMs Induce Virus Neutralizing Antibodies. Vaccine 1992, 10, 585–590. [Google Scholar] [CrossRef]
- Reid, G. Soluble Proteins Incorporate into ISCOMs after Covalent Attachment of Fatty Acid. Vaccine 1992, 10, 597–602. [Google Scholar] [CrossRef]
- Sundquist, B.G.; Czifra, G.; Stipkovits, L. Protective Immunity Induced in Chicken by a Single Immunization with Mycoplasma Gallisepticum Immunostimulating Complexes (ISCOMS). Vaccine 1996, 14, 892–897. [Google Scholar] [CrossRef]
- Hillyer, J.F.; Albrecht, R.M. Gastrointestinal Persorption and Tissue Distribution of Differently Sized Colloidal Gold Nanoparticles. J. Pharm. Sci. 2001, 90, 1927–1936. [Google Scholar] [CrossRef]
- Harkness, K.M.; Cliffel, D.E.; McLean, J.A. Characterization of Thiolate-Protected Gold Nanoparticles by Mass Spectrometry. Analyst 2010, 135, 868–874. [Google Scholar] [CrossRef]
- Athar, T.; Khan, M.K.; Pandey, A.; Hamurcu, M.; Saqib, Z.A.; Gezgin, S. Current Status and Future Directions for Examining Nanoparticles in Plants. Toxic. Nanopart. Plants 2022, 373–398. [Google Scholar] [CrossRef]
- Danino, D. Cryo-TEM of Soft Molecular Assemblies. Curr. Opin. Colloid Interface Sci. 2012, 17, 316–329. [Google Scholar] [CrossRef]
- Coleman, V.A.; Jämting, Å.K.; Catchpoole, H.J.; Roy, M.; Herrmann, J. Nanoparticles and Metrology: A Comparison of Methods for the Determination of Particle Size Distributions. SPIE 2011, 8105, 13–19. [Google Scholar] [CrossRef]
- Hernández-Garrido, J.C.; Moreno, M.S.; Ducati, C.; Pérez, L.A.; Midgley, P.A.; Coronado, E.A. Exploring the Benefits of Electron Tomography to Characterize the Precise Morphology of Core–Shell Au@Ag Nanoparticles and Its Implications on Their Plasmonic Properties. Nanoscale 2014, 6, 12696–12702. [Google Scholar] [CrossRef]
- Volkov, V.V.; Klechkovskaya, V.V.; Shtykova, E.V.; Dembo, K.A.; Arkharova, N.A.; Ivakin, G.I.; Smyslov, R.Y. Determination of the Size and Phase Composition of Silver Nanoparticles in a Gel Film of Bacterial Cellulose by Small-Angle X-Ray Scattering, Electron Diffraction, and Electron Microscopy. Crystallogr. Rep. 2009, 54, 169–173. [Google Scholar] [CrossRef]
- Edelman, I.S.; Petrakovskaja, E.A.; Petrov, D.A.; Zharkov, S.M.; Khaibullin, R.I.; Nuzhdin, V.I.; Stepanov, A.L. FMR and TEM Studies of Co and Ni Nanoparticles Implanted in the SiO2 Matrix. Appl. Magn. Reson. 2011, 40, 363–375. [Google Scholar] [CrossRef]
- Busó-Rogero, C.; Brimaud, S.; Solla-Gullon, J.; Vidal-Iglesias, F.J.; Herrero, E.; Behm, R.J.; Feliu, J.M. Ethanol Oxidation on Shape-Controlled Platinum Nanoparticles at Different PHs: A Combined in Situ IR Spectroscopy and Online Mass Spectrometry Study. J. Electroanal. Chem. 2016, 763, 116–124. [Google Scholar] [CrossRef]
- Williams, D.B.; Carter, C.B. The Transmission Electron Microscope. Transm. Electron Microsc. 1996, 3–17. [Google Scholar] [CrossRef]
- Allabashi, R.; Stach, W.; De La Escosura-Muñiz, A.; Liste-Calleja, L.; Merkoçi, A. ICP-MS: A Powerful Technique for Quantitative Determination of Gold Nanoparticles without Previous Dissolving. J. Nanopart. Res. 2009, 11, 2003–2011. [Google Scholar] [CrossRef]
- Liao, H.G.; Shao, Y.; Wang, C.; Lin, Y.; Jiang, Y.X.; Sun, S.G. TEM Study of Fivefold Twined Gold Nanocrystal Formation Mechanism. Mater. Lett. 2014, 116, 299–303. [Google Scholar] [CrossRef]
- Kauling, A.; Ebeling, G.; Morais, J.; Pádua, A.; Grehl, T.; Brongersma, H.H.; Dupont, J. Surface Composition/Organization of Ionic Liquids with Au Nanoparticles Revealed by High-Sensitivity Low-Energy Ion Scattering. Langmuir 2013, 29, 14301–14306. [Google Scholar] [CrossRef]
- Malik, R.; Annapoorni, S.; Lamba, S.; Raghavendra Reddy, V.; Gupta, A.; Sharma, P.; Inoue, A. Mössbauer and Magnetic Studies in Nickel Ferrite Nanoparticles: Effect of Size Distribution. J. Magn. Magn. Mater. 2010, 322, 3742–3747. [Google Scholar] [CrossRef]
- Filipe, V.; Hawe, A.; Jiskoot, W. Critical Evaluation of Nanoparticle Tracking Analysis (NTA) by NanoSight for the Measurement of Nanoparticles and Protein Aggregates. Pharm. Res. 2010, 27, 796–810. [Google Scholar] [CrossRef]
- Protesescu, L.; Yakunin, S.; Bodnarchuk, M.I.; Krieg, F.; Caputo, R.; Hendon, C.H.; Yang, R.X.; Walsh, A.; Kovalenko, M.V. Nanocrystals of Cesium Lead Halide Perovskites (CsPbX3, X = Cl, Br, and I): Novel Optoelectronic Materials Showing Bright Emission with Wide Color Gamut. Nano Lett. 2015, 15, 3692–3696. [Google Scholar] [CrossRef]
- Khatri, D.K.; Preeti, K.; Tonape, S.; Bhattacharjee, S.; Patel, M.; Shah, S.; Singh, P.K.; Srivastav, S.; Gugulothu, D.; Vora, L.; et al. Nanotechnological Advances for Nose to Brain Delivery of Therapeutics to Improve the Parkinson Therapy. Curr. Neuropharmacol. 2022, 20. [Google Scholar] [CrossRef] [PubMed]
- Beale, A.M.; Weckhuysen, B.M. EXAFS as a tool to interrogate the size and shape of mono and bimetallic catalyst nanoparticles. Phys. Chem. Chem. Phys. 2010, 12, 5562–5574. [Google Scholar] [CrossRef] [PubMed]
- De Haro, L.P.; Karaulanov, T.; Vreeland, E.C.; Anderson, B.; Hathaway, H.J.; Huber, D.L.; Matlashov, A.N.; Nettles, C.P.; Price, A.D.; Monson, T.C.; et al. Magnetic Relaxometry as Applied to Sensitive Cancer Detection and Localization. Biomed. Tech. 2015, 60, 445–455. [Google Scholar] [CrossRef] [PubMed]
- Sebby, K.B.; Mansfield, E. Determination of the Surface Density of Polyethylene Glycol on Gold Nanoparticles by Use of Microscale Thermogravimetric Analysis. Anal. Bioanal. Chem. 2015, 407, 2913–2922. [Google Scholar] [CrossRef]
- Upadhyay, S.; Parekh, K.; Pandey, B. Influence of Crystallite Size on the Magnetic Properties of Fe3O4 Nanoparticles. J. Alloys Compd. 2016, 678, 478–485. [Google Scholar] [CrossRef]
- Moderna, C.-V. European Medicines Agency: EMA/15689/2021 Corr.1* Assessment Report Covid-19 Vaccine Moderna. 2021, 11, p. 169. Available online: https://www.ema.europa.eu/en/documents/assessment-report/spikevax-previously-covid-19-vaccine-moderna-epar-public-assessment-report_en.pdf (accessed on 25 September 2022).
- Tran, J.C.; Zamdborg, L.; Ahlf, D.R.; Lee, J.E.; Catherman, A.D.; Durbin, K.R.; Tipton, J.D.; Vellaichamy, A.; Kellie, J.F.; Li, M. Mapping Intact Protein Isoforms in Discovery Mode Using Top-down Proteomics. Nature 2011, 480, 254–258. [Google Scholar] [CrossRef]
- Santos, P.; Almeida, F. Exosome-Based Vaccines: History, Current State, and Clinical Trials. Front. Immunol. 2021, 12, 1–15. [Google Scholar] [CrossRef]
- Zeng, F.; Morelli, A.E. Extracellular Vesicle-Mediated MHC Cross-Dressing in Immune Homeostasis, Transplantation, Infectious Diseases, and Cancer. Semin. Immunopathol. 2018, 40, 477–490. [Google Scholar] [CrossRef]
- Jiang, L.; Dong, H.; Cao, H.; Ji, X.; Luan, S.; Liu, J. Exosomes in Pathogenesis, Diagnosis, and Treatment of Alzheimer’s Disease. Med. Sci. Monit. 2019, 25, 3329–3335. [Google Scholar] [CrossRef]
- Console, L.; Scalise, M.; Indiveri, C. Exosomes in Inflammation and Role as Biomarkers. Clin. Chim. Acta 2019, 488, 165–171. [Google Scholar] [CrossRef] [PubMed]
- Yoo, K.H.O.; Thapa, N.; Kim, B.J.; Lee, J.O.K.; Jang, Y.N.A.; Chwae, Y.J.; Kim, J. Possibility of Exosome-Based Coronavirus Disease 2019 Vaccine (Review). Mol. Med. Rep. 2022, 25, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Sun, D.; Zhuang, X.; Xiang, X.; Liu, Y.; Zhang, S.; Liu, C.; Barnes, S.; Grizzle, W.; Miller, D.; Zhang, H.G. A Novel Nanoparticle Drug Delivery System: The Anti-Inflammatory Activity of Curcumin Is Enhanced When Encapsulated in Exosomes. Mol. Ther. 2010, 18, 1606–1614. [Google Scholar] [CrossRef] [PubMed]
- Saleh, A.F.; Lázaro-Ibáñez, E.; Forsgard, M.A.-M.; Shatnyeva, O.; Osteikoetxea, X.; Karlsson, F.; Heath, N.; Ingelsten, M.; Rose, J.; Harris, J.; et al. Extracellular Vesicles Induce Minimal Hepatotoxicity and Immunogenicity. Nanoscale 2019, 11, 6990–7001. [Google Scholar] [CrossRef]
- Krishnan, A.; Gangadaran, P.; Chavda, V.P.; Jogalekar, M.P.; Muthusamy, R.; Valu, D.; Vadivalagan, C.; Ramani, P.; Laishevtcev, A.; Katari, N.K.; et al. Convalescent Serum-Derived Exosomes: Attractive Niche as COVID-19 Diagnostic Tool and Vehicle for MRNA Delivery. Exp. Biol. Med. 2022, 15353702221092984. [Google Scholar] [CrossRef]
- Anand, K.; Ramesh, M.; Singh, T.; Balakumar, C.; Chithravel, V.; Prasher, P.; Katari, N.K.; Gupta, G.; Singh, S.K.; Chellappan, D.K.; et al. One-Step Synthesis of Picolinohydrazides from Fusaric Acid: DFT, Structural Characterization and Molecular Inhibitory Studies on Metastatic Tumor-Derived Exosomal and Non-Exosomal Proteins. J. Mol. Struct. 2022, 1255, 132442. [Google Scholar] [CrossRef]
- Thakur, A.; Parra, D.C.; Motallebnejad, P.; Brocchi, M.; Chen, H.J. Exosomes: Small Vesicles with Big Roles in Cancer, Vaccine Development, and Therapeutics. Bioact. Mater. 2022, 10, 281–294. [Google Scholar] [CrossRef]
- Balachandran, B.; Yuana, Y. Extracellular Vesicles-Based Drug Delivery System for Cancer Treatment. Cogent Med. 2019, 6, 1635806. [Google Scholar] [CrossRef]
- Hazrati, A.; Soudi, S.; Malekpour, K.; Mahmoudi, M.; Rahimi, A.; Hashemi, S.M.; Varma, R.S. Immune Cells-Derived Exosomes Function as a Double-Edged Sword: Role in Disease Progression and Their Therapeutic Applications. Biomark. Res. 2022, 10, 130. [Google Scholar] [CrossRef]
- Chiozzini, C.; Manfredi, F.; Arenaccio, C.; Ferrantelli, F.; Leone, P.; Federico, M. N-Terminal Fatty Acids of NEFMUT Are Required for the CD8+ T-Cell Immunogenicity of In Vivo Engineered Extracellular Vesicles. Vaccines 2020, 8, 243. [Google Scholar] [CrossRef]
- Shi, X.; Sun, J.; Li, H.; Lin, H.; Xie, W.; Li, J.; Tan, W. Antitumor Efficacy of Interferon-γ-modified Exosomal Vaccine in Prostate Cancer. Prostate 2020, 80, 811–823. [Google Scholar] [CrossRef] [PubMed]
- Maemura, T.; Fukuyama, S.; Kawaoka, Y. High Levels of MiR-483-3p Are Present in Serum Exosomes Upon Infection of Mice With Highly Pathogenic Avian Influenza Virus. Front. Microbiol. 2020, 11. [Google Scholar] [CrossRef] [PubMed]
- Sotillo, J.; Pearson, M.; Potriquet, J.; Becker, L.; Pickering, D.; Mulvenna, J.; Loukas, A. Extracellular Vesicles Secreted by Schistosoma Mansoni Contain Protein Vaccine Candidates. Int. J. Parasitol. 2016, 46, 1–5. [Google Scholar] [CrossRef] [PubMed]
- Samoil, V.; Dagenais, M.; Ganapathy, V.; Aldridge, J.; Glebov, A.; Jardim, A.; Ribeiro, P. Vesicle-Based Secretion in Schistosomes: Analysis of Protein and MicroRNA (MiRNA) Content of Exosome-like Vesicles Derived from Schistosoma Mansoni. Sci. Rep. 2018, 8, 3286. [Google Scholar] [CrossRef]
- Morse, M.A.; Garst, J.; Osada, T.; Khan, S.; Hobeika, A.; Clay, T.M.; Valente, N.; Shreeniwas, R.; Sutton, M.A.; Delcayre, A.; et al. A Phase I Study of Dexosome Immunotherapy in Patients with Advanced Non-Small Cell Lung Cancer. J. Transl. Med. 2005, 3. [Google Scholar] [CrossRef]
- Escudier, B.; Dorval, T.; Chaput, N.; André, F.; Caby, M.P.; Novault, S.; Flament, C.; Leboulaire, C.; Borg, C.; Amigorena, S.; et al. Vaccination of Metastatic Melanoma Patients with Autologous Dendritic Cell (DC) Derived-Exosomes: Results of the First Phase 1 Clinical Trial. J. Transl. Med. 2005, 3. [Google Scholar] [CrossRef]
- Besse, B.; Charrier, M.; Lapierre, V.; Dansin, E.; Lantz, O.; Planchard, D.; Le Chevalier, T.; Livartoski, A.; Barlesi, F.; Laplanche, A.; et al. Dendritic Cell-Derived Exosomes as Maintenance Immunotherapy after First Line Chemotherapy in NSCLC. Oncoimmunology 2016, 5, e1071008. [Google Scholar] [CrossRef]
- Nassar, W.; El-Ansary, M.; Sabry, D.; Mostafa, M.A.; Fayad, T.; Kotb, E.; Temraz, M.; Saad, A.-N.; Essa, W.; Adel, H. Umbilical Cord Mesenchymal Stem Cells Derived Extracellular Vesicles Can Safely Ameliorate the Progression of Chronic Kidney Diseases. Biomater. Res. 2016, 20, 21. [Google Scholar] [CrossRef]
- Dai, S.; Wei, D.; Wu, Z.; Zhou, X.; Wei, X.; Huang, H.; Li, G. Phase I Clinical Trial of Autologous Ascites-Derived Exosomes Combined with GM-CSF for Colorectal Cancer. Mol. Ther. 2008, 16, 782–790. [Google Scholar] [CrossRef]
- Pocsfalvi, G.; Mammadova, R.; Ramos Juarez, A.P.; Bokka, R.; Trepiccione, F.; Capasso, G. COVID-19 and Extracellular Vesicles: An Intriguing Interplay. Kidney Blood Press Res. 2020, 45, 661–670. [Google Scholar] [CrossRef]
- Wang, Z.; Popowski, K.D.; Zhu, D.; de Juan Abad, B.L.; Wang, X.; Liu, M.; Lutz, H.; De Naeyer, N.; DeMarco, C.T.; Denny, T.N.; et al. Exosomes Decorated with a Recombinant SARS-CoV-2 Receptor-Binding Domain as an Inhalable COVID-19 Vaccine. Nat. Biomed. Eng. 2022, 6, 791–805. [Google Scholar] [CrossRef] [PubMed]
- Monpara, J.D.; Sodha, S.J.; Gupta, P.K. COVID-19 Associated Complications and Potential Therapeutic Targets. Eur. J. Pharmacol. 2020, 886, 173548. [Google Scholar] [CrossRef] [PubMed]
- Tsai, S.-J.; Guo, C.; Atai, N.A.; Gould, S.J. Exosome-Mediated MRNA Delivery For SARS-CoV-2 Vaccination. bioRxiv 2020, 11, 371419. [Google Scholar] [CrossRef]
- Geall, A.J.; Mandl, C.W.; Ulmer, J.B. RNA: The New Revolution in Nucleic Acid Vaccines. Semin. Immunol. 2013, 25, 152–159. [Google Scholar] [CrossRef] [PubMed]
- Ferraro, B.; Morrow, M.P.; Hutnick, N.A.; Shin, T.H.; Lucke, C.E.; Weiner, D.B. Clinical Applications of DNA Vaccines: Current Progress. Clin. Infect. Dis. 2011, 53, 296–302. [Google Scholar] [CrossRef] [PubMed]
- Han, S. Clinical Vaccine Development. Clin. Exp. Vaccine Res. 2015, 4, 46. [Google Scholar] [CrossRef]
- Thanh Le, T.; Andreadakis, Z.; Kumar, A.; Gómez Román, R.; Tollefsen, S.; Saville, M.; Mayhew, S. The COVID-19 Vaccine Development Landscape. Nat. Rev. Drug Discov. 2020, 19, 305–306. [Google Scholar] [CrossRef]
- Hudu, S.A.; Shinkafi, S.H.; Umar, S. An Overview of Recombinant Vaccine Technology, Adjuvants and Vaccine Delivery Methods. Int. J. Pharm. Pharm. Sci. 2016, 8, 19–24. [Google Scholar] [CrossRef]
- Ho, W.; Gao, M.; Li, F.; Li, Z.; Zhang, X.Q.; Xu, X. Next-Generation Vaccines: Nanoparticle-Mediated DNA and MRNA Delivery. Adv. Healthc. Mater. 2021, 10, 2001812. [Google Scholar] [CrossRef]
- Coban, C.; Koyama, S.; Takeshita, F.; Akira, S.; Ishii, K.J. Molecular and Cellular Mechanisms of DNA Vaccines. Hum. Vaccines 2008, 4, 453–457. [Google Scholar] [CrossRef]
- Zhang, C.; Maruggi, G.; Shan, H.; Li, J. Advances in MRNA Vaccines for Infectious Diseases. Front. Immunol. 2019, 10, 594. [Google Scholar] [CrossRef] [PubMed]
- Bedford, H.; Elliman, D. Concerns about Immunisation. BMJ 2000, 320, 240–243. [Google Scholar] [CrossRef] [PubMed]
- Chrastina, A.; Massey, K.A.; Schnitzer, J.E. Overcoming in Vivo Barriers to Targeted Nanodelivery. Wiley Interdiscip. Rev. Nanomed. Nanobiotechnol. 2011, 3, 421–437. [Google Scholar] [CrossRef] [PubMed]
- Davis, M.E.; Chen, Z.; Shin, D.M. Nanoparticle Therapeutics: An Emerging Treatment Modality for Cancer. Nat. Rev. Drug Discov. 2008, 7, 771–782. [Google Scholar] [CrossRef]
- Chow, E.K.H.; Ho, D. Cancer Nanomedicine: From Drug Delivery to Imaging. Sci. Transl. Med. 2013, 5, 216rv4. [Google Scholar] [CrossRef]
- Fries, C.N.; Curvino, E.J.; Chen, J.L.; Permar, S.R.; Fouda, G.G.; Collier, J.H. Advances in Nanomaterial Vaccine Strategies to Address Infectious Diseases Impacting Global Health. Nat. Nanotechnol. 2020, 16, 1–14. [Google Scholar] [CrossRef]
- Reddy, S.T.; Van Der Vlies, A.J.; Simeoni, E.; Angeli, V.; Randolph, G.J.; O’Neil, C.P.; Lee, L.K.; Swartz, M.A.; Hubbell, J.A. Exploiting Lymphatic Transport and Complement Activation in Nanoparticle Vaccines. Nat. Biotechnol. 2007, 25, 1159–1164. [Google Scholar] [CrossRef]
- Xu, X.; Ho, W.; Zhang, X.; Bertrand, N.; Farokhzad, O. Cancer Nanomedicine: From Targeted Delivery to Combination Therapy. Trends Mol. Med. 2015, 21, 223–232. [Google Scholar] [CrossRef]
- Suschak, J.J.; Williams, J.A.; Schmaljohn, C.S. Advancements in DNA Vaccine Vectors, Non-Mechanical Delivery Methods, and Molecular Adjuvants to Increase Immunogenicity. Hum. Vaccines Immunother. 2017, 13, 2837–2848. [Google Scholar] [CrossRef]
- Sullivan, S.M.; Doukas, J.; Hartikka, J.; Smith, L.; Rolland, A. Vaxfectin: A Versatile Adjuvant for Plasmid DNA- and Protein-Based Vaccines. Expert Opin. Drug Deliv. 2010, 7, 1433–1446. [Google Scholar] [CrossRef]
- Smith, L.R.; Wloch, M.K.; Ye, M.; Reyes, L.R.; Boutsaboualoy, S.; Dunne, C.E.; Chaplin, J.A.; Rusalov, D.; Rolland, A.P.; Fisher, C.L.; et al. Phase 1 Clinical Trials of the Safety and Immunogenicity of Adjuvanted Plasmid DNA Vaccines Encoding Influenza A Virus H5 Hemagglutinin. Vaccine 2010, 28, 2565–2572. [Google Scholar] [CrossRef] [PubMed]
- Danko, J.R.; Kochel, T.; Teneza-Mora, N.; Luke, T.C.; Raviprakash, K.; Sun, P.; Simmons, M.; Moon, J.E.; De La Barrera, R.; Martinez, L.J.; et al. Safety and Immunogenicity of a Tetravalent Dengue DNA Vaccine Administered with a Cationic Lipid-Based Adjuvant in a Phase 1 Clinical Trial. Am. J. Trop. Med. Hyg. 2018, 98, 849–856. [Google Scholar] [CrossRef] [PubMed]
- Shiao, S.L.; Ganesan, A.P.; Rugo, H.S.; Coussens, L.M. Immune Microenvironments in Solid Tumors: New Targets for Therapy. Genes Dev. 2011, 25, 2559–2572. [Google Scholar] [CrossRef] [PubMed]
- Gnjatic, S.; Altorki, N.K.; Tang, D.N.; Tu, S.-M.; Kundra, V.; Ritter, G.; Old, L.J.; Logothetis, C.J.; Sharma, P. NY-ESO-1 DNA Vaccine Induces T-Cell Responses That Are Suppressed by Regulatory T Cells. Clin. Cancer Res. 2009, 15, 2130–2139. [Google Scholar] [CrossRef] [PubMed]
- Anselmo, A.C.; Mitragotri, S. Nanoparticles in the Clinic. Bioeng. Transl. Med. 2016, 1, 10–29. [Google Scholar] [CrossRef] [PubMed]
- Lim, M.; Badruddoza, A.Z.M.; Firdous, J.; Azad, M.; Mannan, A.; Al-Hilal, T.A.; Cho, C.S.; Islam, M.A. Engineered Nanodelivery Systems to Improve Dna Vaccine Technologies. Pharmaceutics 2020, 12, 30. [Google Scholar] [CrossRef]
- Boraschi, D.; Italiani, P.; Palomba, R.; Decuzzi, P.; Duschl, A.; Fadeel, B.; Moghimi, S.M. Nanoparticles and Innate Immunity: New Perspectives on Host Defence. Semin. Immunol. 2017, 34, 33–51. [Google Scholar] [CrossRef]
- Behzadi, S.; Serpooshan, V.; Tao, W.; Hamaly, M.A.; Alkawareek, M.Y.; Dreaden, E.C.; Brown, D.; Alkilany, A.M.; Farokhzad, O.C.; Mahmoudi, M. Cellular Uptake of Nanoparticles: Journey inside the Cell. Chem. Soc. Rev. 2017, 46, 4218–4244. [Google Scholar] [CrossRef]
- Grego, E.A.; Siddoway, A.C.; Uz, M.; Liu, L.; Christiansen, J.C.; Ross, K.A.; Kelly, S.M.; Mallapragada, S.K.; Wannemuehler, M.J.; Narasimhan, B. Polymeric Nanoparticle-Based Vaccine Adjuvants and Delivery Vehicles. Curr. Top. Microbiol. Immunol. 2021, 433, 29–76. [Google Scholar] [CrossRef]
- Uto, T.; Wang, X.; Sato, K.; Haraguchi, M.; Akagi, T.; Akashi, M.; Baba, M. Targeting of Antigen to Dendritic Cells with Poly(Gamma-Glutamic Acid) Nanoparticles Induces Antigen-Specific Humoral and Cellular Immunity. J. Immunol. 2007, 178, 2979–2986. [Google Scholar] [CrossRef]
- Alaraby, M.; Abass, D.; Villacorta, A.; Hernández, A.; Marcos, R. Antagonistic in Vivo Interaction of Polystyrene Nanoplastics and Silver Compounds. A Study Using Drosophila. Sci. Total Environ. 2022, 842, 156923. [Google Scholar] [CrossRef] [PubMed]
- Choo, P.; Liu, T.; Odom, T.W. Nanoparticle Shape Determines Dynamics of Targeting Nanoconstructs on Cell Membranes. J. Am. Chem. Soc. 2021, 143, 4550. [Google Scholar] [CrossRef]
- Jiang, Q.; Peng, M.; Yin, M.; Shen, J.; Yan, S. Nanocarrier-Loaded Imidaclothiz Promotes Plant Uptake and Decreases Pesticide Residue. Int. J. Mol. Sci. 2022, 23, 6651. [Google Scholar] [CrossRef] [PubMed]
- Diaz-Diestra, D.M.; Palacios-Hernandez, T.; Liu, Y.; Smith, D.E.; Nguyen, A.K.; Todorov, T.; Gray, P.J.; Zheng, J.; Skoog, S.A.; Goering, P.L. Impact of Surface Chemistry of Ultrasmall Superparamagnetic Iron Oxide Nanoparticles on Protein Corona Formation and Endothelial Cell Uptake, Toxicity, and Barrier Function. Toxicol. Sci. 2022, 188, 261–275. [Google Scholar] [CrossRef]
- Leopold, L.F.; Ruginǎ, D.; Oprea, I.; Diaconeasa, Z.; Leopold, N.; Suciu, M.; Coman, V.; Vodnar, D.C.; Pintea, A.; Coman, C. Warfarin-Capped Gold Nanoparticles: Synthesis, Cytotoxicity, and Cellular Uptake. Molecules 2019, 24, 4145. [Google Scholar] [CrossRef]
- de Boer, I.; Richards, C.J.; Åberg, C. Simultaneous Exposure of Different Nanoparticles Influences Cell Uptake. Pharmaceutics 2022, 14, 136. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Xi, W.S.; Su, Q.; Li, Y.; Yan, G.H.; Liu, Y.; Wang, H.; Cao, A. Unexpected Size Effect: The Interplay between Different-Sized Nanoparticles in Their Cellular Uptake. Small 2019, 15, 1901687. [Google Scholar] [CrossRef]
- Sun, W.; Gao, J.; Fan, R.; Zhang, T.; Tian, Y.; Wang, Z.; Zhang, H.; Zheng, A. The Effect of Particle Size on the Absorption of Cyclosporin A Nanosuspensions. Int. J. Nanomed. 2022, 17, 1741. [Google Scholar] [CrossRef]
- Win, K.Y.; Feng, S.S. Effects of particle size and surface coating on cellular uptake of polymeric nanoparticles for oral delivery of anticancer drugs. Biomaterials 2005, 26, 2713–2722. [Google Scholar] [CrossRef]
- Borzęcka, W.; Pereira, P.M.R.; Fernandes, R.; Trindade, T.; Torres, T.; Tomé, J.P.C. Spherical and Rod Shaped Mesoporous Silica Nanoparticles for Cancer-Targeted and Photosensitizer Delivery in Photodynamic Therapy. J. Mater. Chem. B 2022, 10, 3248–3259. [Google Scholar] [CrossRef]
- Enea, M.; de Almeida, M.P.; Eaton, P.; da Silva, D.D.; Pereira, E.; Soares, M.E.; Bastos, M.D.L.; Carmo, H. A multiparametric study of gold nanoparticles cytotoxicity, internalization and permeability using an in vitro model of blood–brain barrier. Influence of size, shape and capping agent. Nanotoxicology 2019, 13, 990–1004. [Google Scholar] [CrossRef] [PubMed]
- Bastings, M.M.C.; Anastassacos, F.M.; Ponnuswamy, N.; Leifer, F.G.; Cuneo, G.; Lin, C.; Ingber, D.E.; Ryu, J.H.; Shih, W.M. Modulation of the Cellular Uptake of DNA Origami through Control over Mass and Shape. Nano Lett. 2018, 18, 3557–3564. [Google Scholar] [CrossRef] [PubMed]
- Xie, X.; Liao, J.; Shao, X.; Li, Q.; Lin, Y. The Effect of Shape on Cellular Uptake of Gold Nanoparticles in the Forms of Stars, Rods, and Triangles. Sci. Rep. 2017, 7, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Bewersdorff, T.; Gruber, A.; Eravci, M.; Dumbani, M.; Klinger, D.; Haase, A. Amphiphilic Nanogels: Influence of Surface Hydrophobicity on Protein Corona, Biocompatibility and Cellular Uptake. Int. J. Nanomed. 2019, 14, 7861. [Google Scholar] [CrossRef]
- Dai, Z.; Song, J.; Chen, Y.; Feng, L.; Xu, Y.; Li, D.; Wu, C.; Zhang, Z.; Liu, J. Study on the Bioavailability of Stevioside-Encapsulized Lutein and Its Mechanism. Food Chem. 2021, 354. [Google Scholar] [CrossRef]
- Tomeh, M.A.; Hadianamrei, R.; Sun, W.; Xu, D.; Brown, S.; Zhao, X. Stiffness-Tuneable Nanocarriers for Controlled Delivery of ASC-J9 into Colorectal Cancer Cells. J. Colloid Interface Sci. 2021, 594, 513–521. [Google Scholar] [CrossRef]
- Ho, H.M.K.; Craig, D.Q.M.; Day, R.M. Design of Experiment Approach to Modeling the Effects of Formulation and Drug Loading on the Structure and Properties of Therapeutic Nanogels. Mol. Pharm. 2022, 19, 602–615. [Google Scholar] [CrossRef]
- Olivieri, P.H.; Jesus, M.B.; Nader, H.B.; Justo, G.Z.; Sousa, A.A. Cell-Surface Glycosaminoglycans Regulate the Cellular Uptake of Charged Polystyrene Nanoparticles. Nanoscale 2022, 14, 7350–7363. [Google Scholar] [CrossRef]
- Hanot, C.C.; Choi, Y.S.; Anani, T.B.; Soundarrajan, D.; David, A.E. Effects of Iron-Oxide Nanoparticle Surface Chemistry on Uptake Kinetics and Cytotoxicity in CHO-K1 Cells. Int. J. Mol. Sci. 2015, 17, 54. [Google Scholar] [CrossRef]
- Chen, L.; Zhao, T.; Zhao, M.; Wang, W.; Sun, C.; Liu, L.; Li, Q.; Zhang, F.; Zhao, D.; Li, X. Size and Charge Dual-Transformable Mesoporous Nanoassemblies for Enhanced Drug Delivery and Tumor Penetration. Chem. Sci. 2020, 11, 2819. [Google Scholar] [CrossRef]
- Petrini, M.; Lokerse, W.J.M.; Mach, A.; Hossann, M.; Merkel, O.M.; Lindner, L.H. Effects of Surface Charge, PEGylation and Functionalization with Dipalmitoylphosphatidyldiglycerol on Liposome–Cell Interactions and Local Drug Delivery to Solid Tumors via Thermosensitive Liposomes. Int. J. Nanomed. 2021, 16, 4045. [Google Scholar] [CrossRef] [PubMed]
- David, S.; Patel, D.Y.; Cardona, S.M.; Kirby, N.; Mayer, K.M. Cellular Uptake and Cytotoxicity of PEGylated Gold Nanoparticles in C33A Cervical Cancer Cells. Nano Express 2022, 3, 025006. [Google Scholar] [CrossRef]
- Arathi, A.; Joseph, X.; Akhil, V.; Mohanan, P. L-cysteine capped Zinc Oxide nanoparticles induced cellular response on adenocarcinomic human alveolar basal epithelial cells using a conventional and organ-on-a-chip approach. Colloids Surf. B Biointerfaces 2021, 211, 112300. [Google Scholar] [CrossRef]
- Bolaños, K.; Kogan, M.J.; Araya, E. Capping gold nanoparticles with albumin to improve their biomedical properties. Int. J. Nanomed. 2019, 14, 6387–6406. [Google Scholar] [CrossRef] [PubMed]
- Foroozandeh, P.; Aziz, A.A. Insight into Cellular Uptake and Intracellular Trafficking of Nanoparticles. Nanoscale Res. Lett. 2018, 13, 339. [Google Scholar] [CrossRef]
- Panariti, A.; Miserocchi, G.; Rivolta, I. The Effect of Nanoparticle Uptake on Cellular Behavior: Disrupting or Enabling Functions? Nanotechnol. Sci. Appl. 2012, 5, 87. [Google Scholar] [CrossRef]
- Shang, L.; Nienhaus, K.; Nienhaus, G.U. Engineered Nanoparticles Interacting with Cells: Size Matters. J. Nanobiotechnol. 2014, 12, 1–11. [Google Scholar] [CrossRef]
- Kumari, S.; Mg, S.; Mayor, S. Endocytosis Unplugged: Multiple Ways to Enter the Cell. Cell Res. 2010, 20, 256–275. [Google Scholar] [CrossRef]
- Wei, X.; She, G.; Wu, T.; Xue, C.; Cao, Y. PEDV Enters Cells through Clathrin-, Caveolae-, and Lipid Raft-Mediated Endocytosis and Traffics via the Endo-/Lysosome Pathway. Vet. Res. 2020, 51, 1–18. [Google Scholar] [CrossRef]
- Damm, E.M.; Pelkmans, L.; Kartenbeck, J.; Mezzacasa, A.; Kurzchalia, T.; Helenius, A. Clathrin- and Caveolin-1–Independent Endocytosis: Entry of Simian Virus 40 into Cells Devoid of Caveolae. J. Cell Biol. 2005, 168, 477. [Google Scholar] [CrossRef]
- Du, X.; Wang, J.; Zhou, Q.; Zhang, L.; Wang, S.; Zhang, Z.; Yao, C. Advanced Physical Techniques for Gene Delivery Based on Membrane Perforation. Drug Deliv. 2018, 25, 1516. [Google Scholar] [CrossRef] [PubMed]
- Vora, L.; Tyagi, M.; Patel, K.; Gupta, S.; Vavia, P. Self-Assembled Nanocomplexes of Anionic Pullulan and Polyallylamine for DNA and PH-Sensitive Intracellular Drug Delivery. J. Nanopart. Res. 2014, 16, 2781. [Google Scholar] [CrossRef]
- Malachowski, T.; Hassel, A. Engineering Nanoparticles to Overcome Immunological Barriers for Enhanced Drug Delivery. Eng. Regen. 2020, 1, 35–50. [Google Scholar] [CrossRef]
- Sabourian, P.; Yazdani, G.; Ashraf, S.S.; Frounchi, M.; Mashayekhan, S.; Kiani, S.; Kakkar, A. Effect of Physico-Chemical Properties of Nanoparticles on Their Intracellular Uptake. Int. J. Mol. Sci. 2020, 21, 8019. [Google Scholar] [CrossRef] [PubMed]
- van Leent, M.M.T.; Priem, B.; Schrijver, D.P.; de Dreu, A.; Hofstraat, S.R.J.; Zwolsman, R.; Beldman, T.J.; Netea, M.G.; Mulder, W.J.M. Regulating Trained Immunity with Nanomedicine. Nat. Rev. Mater. 2022, 7, 465–481. [Google Scholar] [CrossRef]
- Pavlin, M.; Lojk, J.; Strojan, K.; Hafner-Bratkovič, I.; Jerala, R.; Leonardi, A.; Križaj, I.; Drnovšek, N.; Novak, S.; Veranič, P.; et al. The Relevance of Physico-Chemical Properties and Protein Corona for Evaluation of Nanoparticles Immunotoxicity—In Vitro Correlation Analysis on THP-1 Macrophages. Int. J. Mol. Sci. 2022, 23, 6197. [Google Scholar] [CrossRef]
- Toraskar, S.; Madhukar Chaudhary, P.; Kikkeri, R. The Shape of Nanostructures Encodes Immunomodulation of Carbohydrate Antigen and Vaccine Development. ACS Chem. Biol. 2021, 17, 1122–1130. [Google Scholar] [CrossRef]
- Vandebriel, R.J.; Remy, S.; Vermeulen, J.P.; Hurkmans, E.G.E.; Kevenaar, K.; Bastús, N.G.; Pelaz, B.; Soliman, M.G.; Puntes, V.F.; Parak, W.J.; et al. Pathways Related to NLRP3 Inflammasome Activation Induced by Gold Nanorods. Int. J. Mol. Sci. 2022, 23, 5763. [Google Scholar] [CrossRef]
- Weh, E.; Mei, L.; Walsh, L.; Yi, M.; Lin, C.-M. Delivery of a Hydrophobic Small Molecule LXR Agonist via SHDL Nanoparticles in a Preclinical Model of Dry AMD | IOVS | ARVO Journals. Investig. Ophthalmol. Vis. Sci. 2022, 63, 290–293. [Google Scholar]
- Farfán-Castro, S.; García-Soto, M.J.; Comas-García, M.; Arévalo-Villalobos, J.I.; Palestino, G.; González-Ortega, O.; Rosales-Mendoza, S. Synthesis and Immunogenicity Assessment of a Gold Nanoparticle Conjugate for the Delivery of a Peptide from SARS-CoV-2. Nanomedicine 2021, 34, 102372. [Google Scholar] [CrossRef]
- Mekonnen, T.W.; Darge, H.F.; Tsai, H.C.; Birhan, Y.S.; Hanurry, E.Y.; Gebrie, H.T.; Chou, H.Y.; Lai, J.Y.; Lin, S.Z.; Harn, H.J.; et al. Combination of Ovalbumin-Coated Iron Oxide Nanoparticles and Poly(Amidoamine) Dendrimer-Cisplatin Nanocomplex for Enhanced Anticancer Efficacy. Colloids Surf. B Biointerfaces 2022, 213, 112391. [Google Scholar] [CrossRef] [PubMed]
- McCright, J.; Skeen, C.; Yarmovsky, J.; Maisel, K. Nanoparticles with Dense Poly(Ethylene Glycol) Coatings with near Neutral Charge Are Maximally Transported across Lymphatics and to the Lymph Nodes. Acta Biomater. 2022, 145, 146–158. [Google Scholar] [CrossRef] [PubMed]
- Mitarotonda, R.; Saraceno, M.; Todone, M.; Giorgi, E.; Malchiodi, E.L.; Desimone, M.F.; De Marzi, M.C. Surface Chemistry Modification of Silica Nanoparticles Alters the Activation of Monocytes. Ther. Deliv. 2021, 12, 443–459. [Google Scholar] [CrossRef] [PubMed]
- González-García, L.E.; Macgregor, M.N.; Visalakshan, R.M.; Lazarian, A.; Cavallaro, A.A.; Morsbach, S.; Mierczynska-Vasilev, A.; Mailänder, V.; Landfester, K.; Vasilev, K. Nanoparticles Surface Chemistry Influence on Protein Corona Composition and Inflammatory Responses. Nanomaterials 2022, 12, 682. [Google Scholar] [CrossRef] [PubMed]
- Jia, J.; Zhang, Y.; Xin, Y.; Jiang, C.; Yan, B.; Zhai, S. Interactions Between Nanoparticles and Dendritic Cells: From the Perspective of Cancer Immunotherapy. Front. Oncol. 2018, 8, 1–11. [Google Scholar] [CrossRef]
- O’Hagan, D.T.; Fox, C.B. New Generation Adjuvants—From Empiricism to Rational Design. Vaccine 2015, 33, B14–B20. [Google Scholar] [CrossRef]
- Mignani, S.; El Kazzouli, S.; Bousmina, M.; Majoral, J.P. Expand Classical Drug Administration Ways by Emerging Routes Using Dendrimer Drug Delivery Systems: A Concise Overview. Adv. Drug Deliv. Rev. 2013, 65, 1316–1330. [Google Scholar] [CrossRef]
- Tacken, P.J.; Figdor, C.G. Targeted Antigen Delivery and Activation of Dendritic Cells in Vivo: Steps towards Cost Effective Vaccines. Semin. Immunol. 2011, 23, 12–20. [Google Scholar] [CrossRef]
- Yi, X.; Shi, X.; Gao, H. Cellular Uptake of Elastic Nanoparticles. Phys. Rev. Lett. 2011, 107, 098101. [Google Scholar] [CrossRef]
- Tzlil, S.; Deserno, M.; Gelbart, W.M.; Ben-Shaul, A. A Statistical-Thermodynamic Model of Viral Budding. Biophys. J. 2004, 86, 2037–2048. [Google Scholar] [CrossRef]
- Florindo, H.F.; Lopes, J.; Silva, L.C.; Corvo, M.L.; Martins, M.B.; Gaspar, R. Regulatory Development of Nanotechnology-Based Vaccines. Micro-Nanotechnol. Vaccine Dev. 2017, 393–410. [Google Scholar] [CrossRef]
- Cruz, L.J.; Tacken, P.J.; Zeelenberg, I.S.; Srinivas, M.; Bonetto, F.; Weigelin, B.; Eich, C.; De Vries, I.J.; Figdor, C.G. Tracking Targeted Bimodal Nanovaccines: Immune Responses and Routing in Cells, Tissue, and Whole Organism. Mol. Pharm. 2014, 11, 4299–4313. [Google Scholar] [CrossRef] [PubMed]
- Nandedkar, T.D. Nanotechnology a Path to Nanovaccine. Int. J. Pharma Bio Sci. 2012, 3, 290–292. [Google Scholar]
- Guy, B. Adjuvants for Protein-and Carbohydrate-Based Vaccines. Carbohydr.-Based Vaccines Immunother. 2009, 8, 89. [Google Scholar] [CrossRef]
- Gonzalez-Aramundiz, J.V.; Cordeiro, A.S.; Csaba, N.; De La Fuente, M.; Alonso, M.J. Nanovaccines: Nanocarriers for Antigen Delivery. Biol. Aujourdhui. 2012, 206, 249–261. [Google Scholar] [CrossRef]
- Safety and Immunogenicity of an Epstein-Barr Virus (EBV) Gp350-Ferritin Nanoparticle Vaccine in Healthy Adults with or Without EBV Infection. Available online: https://clinicaltrials.gov/ct2/show/NCT04645147 (accessed on 25 September 2022).
- Immunogenicity and Safety Study of SK SARS-CoV-2 Recombinant Nanoparticle Vaccine (GBP510) Adjuvanted With AS03 (COVID-19). Available online: https://clinicaltrials.gov/ct2/show/NCT05007951 (accessed on 25 September 2022).
- Safety and Immunogenicity Study of SARS-CoV-2 Nanoparticle Vaccine (GBP510) Adjuvanted with or without AS03 (COVID-19). Available online: https://clinicaltrials.gov/ct2/show/NCT04750343 (accessed on 25 September 2022).
- Safety and Immunogenicity Study of SARS-CoV-2 Nanoparticle Vaccine (GBP510) Adjuvanted with Aluminum Hydroxide (COVID-19). Available online: https://clinicaltrials.gov/ct2/show/NCT04742738 (accessed on 25 September 2022).
- Universal Influenza A Vaccine in Healthy Adults. Available online: https://clinicaltrials.gov/ct2/show/NCT05155319 (accessed on 25 September 2022).
- A Study to Evaluate Safety and Immunogenicity of a COVID-19 Vaccine in People Living with HIV at Risk for SARS-CoV-2 (COVID-19). Available online: https://clinicaltrials.gov/ct2/show/NCT05112848 (accessed on 25 September 2022).
- A Study Looking at the Effectiveness, Immune Response, and Safety of a COVID-19 Vaccine in Adults in the United Kingdom. Available online: https://clinicaltrials.gov/ct2/show/NCT04583995 (accessed on 25 September 2022).
- Dunkle, L.M.; Kotloff, K.L.; Gay, C.L.; Áñez, G.; Adelglass, J.M.; Barrat Hernández, A.Q.; Harper, W.L.; Duncanson, D.M.; McArthur, M.A.; Florescu, D.F.; et al. Efficacy and Safety of NVX-CoV2373 in Adults in the United States and Mexico. N. Engl. J. Med. 2022, 386, 531–543. [Google Scholar] [CrossRef]
- SARS-COV-2-Spike-Ferritin-Nanoparticle (SpFN) Vaccine with ALFQ Adjuvant for Prevention of COVID-19 in Healthy Adults. Available online: https://clinicaltrials.gov/ct2/show/NCT04784767 (accessed on 25 September 2022).
- Pourmand, A.; Abdollahi, M. Current Opinion on Nanotoxicology. DARU J. Pharm. Sci. 2012, 20, 95. [Google Scholar] [CrossRef]
- Petrarca, C.; Clemente, E.; Amato, V.; Pedata, P.; Sabbioni, E.; Bernardini, G.; Iavicoli, I.; Cortese, S.; Niu, Q.; Otsuki, T.; et al. Engineered Metal Based Nanoparticles and Innate Immunity. Clin. Mol. Allergy 2015, 13, 13. [Google Scholar] [CrossRef]
- Diaz-Arévalo, D.; Zeng, M. Nanoparticle-Based Vaccines. In Nanopharmaceuticals; Elsevier: Amsterdam, The Netherlands, 2020; pp. 135–150. ISBN 9780128177792. [Google Scholar]
- ISO, IEC, NIST and OECD International Workshop on Documentary Standards for Measurement and Characterization for Nanotechnologies. 2008. Available online: https://www.google.com.hk/url?sa=t&rct=j&q=&esrc=s&source=web&cd=&ved=2ahUKEwjcooKDxqD7AhW24zgGHRXuBAYQFnoECA8QAQ&url=https%3A%2F%2Fwww.pnnl.gov%2Fnano%2Fpdf%2FISO_IEC_NIST_OECDwkshp.pdf&usg=AOvVaw1yDOj-qFCVOISAAKJRrTci (accessed on 25 September 2022).
- Nel, A.; Xia, T.; Mädler, L.; Li, N. Toxic Potential of Materials at the Nanolevel. Science 2006, 311, 622–627. [Google Scholar] [CrossRef]
- Lewinski, N.; Colvin, V.; Drezek, R. Cytotoxicity of Nanopartides. Small 2008, 4, 26–49. [Google Scholar] [CrossRef]
- Jia, G.; Wang, H.; Yan, L.; Wang, X.; Pei, R.; Yan, T.; Zhao, Y.; Guo, X. Cytotoxicity of Carbon Nanomaterials: Single-Wall Nanotube, Multi-Wall Nanotube, and Fullerene. Environ. Sci. Technol. 2005, 39, 1378–1383. [Google Scholar] [CrossRef] [PubMed]
- Uo, M.; Akasaka, T.; Watari, F.; Sato, Y.; Tohji, K. Toxicity Evaluations of Various Carbon Nanomaterials. Dent. Mater. J. 2011, 30, 245–263. [Google Scholar] [CrossRef] [PubMed]
- Wick, P.; Manser, P.; Limbach, L.K.; Dettlaff-Weglikowska, U.; Krumeich, F.; Roth, S.; Stark, W.J.; Bruinink, A. The degree and kind of agglomeration affect carbon nanotube cytotoxicity. Toxicol. Lett. 2007, 168, 121–131. [Google Scholar] [CrossRef] [PubMed]
- Sato, Y.; Yokoyama, A.; Shibata, K.I.; Akimoto, Y.; Ogino, S.I.; Nodasaka, Y.; Kohgo, T.; Tamura, K.; Akasaka, T.; Uo, M.; et al. Influence of Length on Cytotoxicity of Multi-Walled Carbon Nanotubes against Human Acute Monocytic Leukemia Cell Line THP-1 in Vitro and Subcutaneous Tissue of Rats in Vivo. Mol. Biosyst. 2005, 1, 176–182. [Google Scholar] [CrossRef]
- Takahashi, H.; Niidome, Y.; Niidome, T.; Kaneko, K.; Kawasaki, H.; Yamada, S. Modification of Gold Nanorods Using Phosphatidylcholine to Reduce Cytotoxicity. Langmuir 2006, 22, 2–5. [Google Scholar] [CrossRef] [PubMed]
- Goodman, C.M.; McCusker, C.D.; Yilmaz, T.; Rotello, V.M. Toxicity of Gold Nanoparticles Functionalized with Cationic and Anionic Side Chains. Bioconjug. Chem. 2004, 15, 897–900. [Google Scholar] [CrossRef] [PubMed]
- Xu, M.; Fujita, D.; Kajiwara, S.; Minowa, T.; Li, X.; Takemura, T.; Iwai, H.; Hanagata, N. Contribution of Physicochemical Characteristics of Nano-Oxides to Cytotoxicity. Biomaterials 2010, 31, 8022–8031. [Google Scholar] [CrossRef]
- Sun, H.; Jiang, C.; Wu, L.; Bai, X.; Zhai, S. Cytotoxicity-Related Bioeffects Induced by Nanoparticles: The Role of Surface Chemistry. Front. Bioeng. Biotechnol. 2019, 7, 414. [Google Scholar] [CrossRef]
- Semisch, A.; Ohle, J.; Witt, B.; Hartwig, A. Cytotoxicity and Genotoxicity of Nano - and Microparticulate Copper Oxide: Role of Solubility and Intracellular Bioavailability. Part. Fibre Toxicol. 2014, 11, 1–16. [Google Scholar] [CrossRef]
- Sahu, D.; Kannan, G.M.; Tailang, M.; Vijayaraghavan, R. In Vitro Cytotoxicity of Nanoparticles: A Comparison between Particle Size and Cell Type. J. Nanosci. 2016, 2016, 1–9. [Google Scholar] [CrossRef]
- Singh, S. Zinc Oxide Nanoparticles Impacts: Cytotoxicity, Genotoxicity, Developmental Toxicity, and Neurotoxicity. Toxicol. Mech. Methods 2019, 29, 300–311. [Google Scholar] [CrossRef] [PubMed]
- Bhamidipati, M.; Fabris, L. Multiparametric Assessment of Gold Nanoparticle Cytotoxicity in Cancerous and Healthy Cells: The Role of Size, Shape, and Surface Chemistry. Bioconjug. Chem. 2017, 28, 449–460. [Google Scholar] [CrossRef] [PubMed]
- Desai, N. Challenges in Development of Nanoparticle-Based Therapeutics. AAPS J. 2012, 14, 282–295. [Google Scholar] [CrossRef] [PubMed]
- Moghimi, S. Modulation of Lymphatic Distribution of Subcutaneously Injected Poloxamer 407-Coated Nanospheres: The Effect of the Ethylene Oxide Chain Configuration. FEBS Lett. 2003, 540, 241–244. [Google Scholar] [CrossRef]
- Moghimi, S.M.; Hunter, A.C.; Murray, J.C. Long-circulating and target-specific nanoparticles: Theory to practice. Pharmacol. Rev. 2001, 53, 283–318. [Google Scholar]
- Moghimi, S.M.; Szebeni, J. Stealth Liposomes and Long Circulating Nanoparticles: Critical Issues in Pharmacokinetics, Opsonization and Protein-Binding Properties. Prog. Lipid Res. 2003, 42, 463–478. [Google Scholar] [CrossRef]
- O’Brien, M.E.R.; Wigler, N.; Inbar, M.; Rosso, R.; Grischke, E.; Santoro, A.; Catane, R.; Kieback, D.G.; Tomczak, P.; Ackland, S.P.; et al. Reduced Cardiotoxicity and Comparable Efficacy in a Phase III Trial of Pegylated Liposomal Doxorubicin HCl (CAELYX/Doxil) versus Conventional Doxorubicin for First-Line Treatment of Metastatic Breast Cancer. Ann. Oncol. Off. J. Eur. Soc. Med. Oncol. 2004, 15, 440–449. [Google Scholar] [CrossRef]
- Gaumet, M.; Vargas, A.; Gurny, R.; Delie, F. Nanoparticles for Drug Delivery: The Need for Precision in Reporting Particle Size Parameters. Eur. J. Pharm. Biopharm. 2008, 69, 1–9. [Google Scholar] [CrossRef]
- Feng, S.-S.; Mu, L.; Win, K.; Huang, G. Nanoparticles of Biodegradable Polymers for Clinical Administration of Paclitaxel. Curr. Med. Chem. 2005, 11, 413–424. [Google Scholar] [CrossRef]
- Kempner, E.S. Effects of High-Energy Electrons and Gamma Rays Directly on Protein Molecules. J. Pharm. Sci. 2001, 90, 1637–1646. [Google Scholar] [CrossRef]
- Özcan, I.; Bouchemal, K.; Segura-Sánchez, F.; Abaci, Ö.; Özer, Ö.; Güneri, T.; Ponchel, G. Effects of Sterilization Techniques on the PEGylated Poly (γ-Benzyl-L-Glutamate) (PBLG) Nanoparticles. Acta Pharm. Sci. 2009, 51, 211–218. [Google Scholar]
- FDA Guidance for Industry: Liposome Drug Products-Chemistry, Manufacturing, and Controls; Human Pharmacokinetics and Bioavailability; and Labeling Documentation-ECA Academy; FDA: Silver Spring, MD, USA, 2018.
- Davis, M.E. The First Targeted Delivery of SiRNA in Humans via a Self-Assembling, Cyclodextrin Polymer-Based Nanoparticle: From Concept to Clinic. Mol. Pharm. 2009, 6, 659–668. [Google Scholar] [CrossRef] [PubMed]
- Larrañeta, E.; Vora, L. Delivery of Nanomedicines Using Microneedles. Microneedles Drug Vaccine Deliv. Patient Monit. 2018, 177–205. [Google Scholar] [CrossRef]
- Trynda-Lemiesz, L. Paclitaxel–HSA Interaction. Binding Sites on HSA Molecule. Bioorg. Med. Chem. 2004, 12, 3269–3275. [Google Scholar] [CrossRef] [PubMed]


| Types of Nanoparticle Systems | Advantages | Disadvantages | Example | Ref. |
|---|---|---|---|---|
| Virus-like particles (VLP) | Free of infectious material | Polydispersed particle size | MalariVax against malaria. (contains core proteins of HBV + epitopes of circumsporozoite proteins of Plasmodium falciparum) | [43] |
| Shielding of immune- modulators prevent off-target effects | Limited encapsidation | |||
| Self-adjuvant | Lack of reproducibility | |||
| Liposomes | Phospholipids possess intrinsic adjuvant property When modified, is stable in GI fluids | Less stable than polymer particles Limited antigen loading esp. hydrophilic antigens | Mosqurix® against both Plasmodium falciparum and HBV | [44] |
| Can take in both hydrophilic and hydrophobic antigens | Poor stability of GI naked liposomes | |||
| Immunostimulatory complexes (ISCOMs) | Lipophilic antigens is easy encapsulate | Antigens that are hydrophilic are difficult to incorporate | Quilvax-L™, a vaccine for dogs for prevention of canine lyme disease | [45] |
| Natural adjuvant | Exert no depot release profile | |||
| It is biodegradable | ||||
| Built- in adjuvant property of Quil A | ||||
| Polymeric | It is biodegradable | Insufficient protection against antigens | Chitosan-DNA nanoparticles displayed effectiveness against infections caused by T.pyogenes | [46] |
| Surface properties can be easily modified to improve immunogenicity | Antigens are released prematurely | |||
| Antigens can be released in targeted sites | Low antigen protection | |||
| Inorganic nanoparticles | Better protection of adsorbed antigens | Nonbiodegradable | Synthetic SNAs (spherical nucleic acids) based nanovaccines for cancer immunotherapy | [47] |
| Chances of premature release is less | Poor aqueous solubility | |||
| Easy surface modification | ||||
| Emulsion | Can take in both hydrophilic and hydrophobic antigens | Poor GI stability | Influenza vaccine Prepandrix® and Pandemrix® | [48] |
| Self-adjuvant | Premature release of antigens | |||
| Lipid nanoparticle | Safe and versatile vehicles for drugs Biodegradable and biocompatible Improve oral drug bioavailability | Low loading efficiency Drug expulsion during storage | Moderna COVID-19 (mRNA-1273) vaccine and BNT162b from Pfizer-BioNTech against SARS-CoV-2 virus. | [49] |
| Exosomes | Can deliver vaccine to targeted site | Methods for quantification are not sensitive enough. | A phase II NCT04902183 against moderate or severe COVID-19 | [50] |
| Modification of the cell is possible by transferring active molecules between cells | High cost |
| Virus | Nanocarrier Platforms | Constituents | Route of Administration | Experimental Outcomes | Advantage | Phase | Ref. |
|---|---|---|---|---|---|---|---|
| HIV | Peptide-based nanofibrous hydrogel | Methylamino group, DNA, Naphthalene acetic acid-Glycine-Phenylalanine-Tyrosine | Intramuscular, intradermal, and subcutaneous | The level of IFN-γ and IL-4 cytokine was markedly increased | Biocompatible and biosafety | Preclinical | [51] |
| Merkel Cell Polyomavirus (MCV) | MCV-like particles | Viral proteins 1 (VP1), Polyomavirus capsid proteins | Intramuscular | Anti-MCV antibody seroprevalence was found to be significantly high in mice inoculated with MCV VLPs. Low antibody seroprevalence of cross-reactive antibodies was reported against LPV VLPs and BKV VLPs, nevertheless, providing for 4.4% and 2.6% of the reactivity against MCV VLPs, respectively. | Nontoxic, biodegradable, and biocompatible | Phase I | [52] |
| Hemorrhagic septicemia virus (HSV) | Polymeric nanoparticles | Poly (lactic-co-glycolic acid) | Mucosal | In exposure to the attenuated viral antigen, the inoculated fish produce anti-VHSV immunoglobulin (Ig), triggering the elements of the humoral immunological response. The anti-VHSV inhibition percentage was significantly increased in immunized groups as compared to a nonvaccinated challenged group | Fabrication is easy, biodegradable, nonimmunogenic, and low toxicity | Preclinical | [53] |
| Influenza A virus | Gold NPs (AuNPs) | Cytosine-guanine-rich oligonucleotide, AuNPs | Intranasal | An enhancement in anti-M2e serum Ig concentrations was observed when M2e was coupled to AuNPs. | Improved bioavailability and half-life, low toxic, and biocompatible | Preclinical | [54] |
| Hepatitis B | Polymeric nanoparticles | Poly (lactic-co-glycolic acid), Poly-lactic acid | Intramuscular and pulmonary | Anti-HBsAg antibody concentration of PLA and PLGA was substantially elevated as compared to plain HBsAg | Site-specific targeted drug delivery, biodegradable, nonimmunogenic, and low toxicity | Phase II | [55] |
| Viral infections | Nanogel | Cationic alginate-poly ethylenimine | Intraperitoneal | Nanogels dramatically augmented anti-OVA IgG1 synthesis but had minimal influence on IgG2a and IgG2b expression. However, IgG isotypes and anti-OVA IgG were enhanced by nanogels in a dose-dependent manner and improved OVA-specific IFN-γ by 60 times. | Nonimmunogenic, highly biocompatible, controlled as well as sustained drug delivery is achieved | Preclinical | [56] |
| Human papillomavirus (HPV) | Virus-like particles (VLPs) | VLPs, L1, and L2 proteins | --- | --- | Targeted site specific, biodegradable, and biocompatible. | Marketed | [57] |
| Methods | Outcomes | Reference |
|---|---|---|
| Atomic force microscopy (AFM) | Rapid interpretation: chemical component, nanoparticle dimensions, and design in 3D modality | [210] |
| Cryo-electron microscopy (Cryo-TEM) | Explore intricate growth pathways, agglomeration mechanism | [211] |
| Differential centrifugal sedimentation (DCS) | Size variation and nanoparticle dimensions | [212] |
| Dynamic light scattering (DLS) | Detecting coalescence, hydrodynamic dimension | [101] |
| Electron tomography | Analytical data at the molecular level, representation of accurate 3D nanoparticles, images, and video | [213] |
| Electron diffraction | Investigate order-disorder transition, lattice variables, and crystal component | [214] |
| Ferromagnetic resonance (FMR) | Nanoparticle dimension, size variable, M values, magnetostatic domain, surface features, shape, and magnetic anisotropic constant | [215] |
| Fourier transform infrared spectroscopy (FTIR) | Ligand interaction, surface features | [216] |
| High-resolution transmission electron microscopy (HRTEM) | Differentiate between polycrystalline, amorphous, and monocrystalline nanoparticles. | [217] |
| Inductively coupled plasma-mass spectrometry (ICP-MS) | Nanoparticle dimension and concentration, size variation, and characterization of the elements | [218] |
| Liquid TEM | Explore intricate growth pathways, fabrication of crystal structures, and real-time mapping of nanoparticle development | [219] |
| Low energy ion scattering (LEIS) | Nanoparticle chemical constitution and thickness | [220] |
| Mossbauer | Thermal unblocking, differentiate between iron oxides, symmetry, surface spins, magnetic anisotropy energy, and oxidation state | [221] |
| Nanoparticle tracking analysis (NTA) | Size variation and nanoparticle dimensions | [222] |
| Photoluminescence spectroscopy | Correlation of optical features and structural properties including flaws, composition, and dimensions | [223] |
| Scanning transmission electron microscopy (STEM) | Investigate the atomic architecture of heterointerfaces, and crystal geometry | [224] |
| Small-angle X-ray scattering (SAXS) | Size variation, growth kinetics, and size of the particle | [225] |
| Superparamagnetic relaxometry | Identify and pinpoint NPs with superparamagnetism, and hydrodynamic size variation | [226] |
| Thermal gravimetric analysis (TGA) | Stabilizers component and weight | [227] |
| X-ray diffraction (XRD) | Dimension of crystal granules, crystal geometry, and composition | [228] |
| Liquid Chromatography (LC) | Quantify and identify the lipid nanocarriers (NCs) of LNC-mRNA approved vaccines | [229] |
| Nuclear magnetic resonance (NMR) spectroscopy | To quantify and learn more about the new lipid excipients’ molecular structure | [229] |
| Capillary gel electrophoresis | To investigate the glycosylation and disulfide bonds in proteins, molecular weight, nucleic acid, concentration and protein integrity | [230] |
| Physicochemical Properties | Nanoparticles | Uptake Mechanism | Remarks | Reference |
|---|---|---|---|---|
| Size | Silver and polystyrene NPs | Polystyrene NPs size 50 nm (PS-50) via clathrin and caveolae-mediated transport; PS-500 via macropinocytosis | Increased uptake in PS-50 due to smaller particle size | [284] |
| Carboxylated polystyrene NPs | Endocytosis | Obstruction of 100 nm particles by particles of size 40 nm in a polydisperse sample, increased uptake of smaller sized particles | [289] | |
| Silica NPs (SNP) | Clathrin-dependent endocytosis | Inhibition of SNP100 uptake in presence of SNP50 | [290] | |
| Cyclosporin A NPs (CsA-NPs) | Endocytosis (<500 nm); lymphatic uptake (>500 nm) | Increased uptake and intestinal absorption of 280 nm sized CsA-NPs than 522 nm and 2967 nm | [291] | |
| Coumarin-6-polylactic-co-glycolic acid NPs | Receptor-mediated endocytosis and phagocytosis | Increased uptake of NPs in the size range of 100–200 nm (optimum size) as compared to 50 nm, 500 nm and 1000 nm | [292] | |
| Shape | Mesoporous silica NPs | Endocytosis | Increased uptake and phototoxicity in cancer cells of spherical shaped particles than the rod-shaped particles | [293] |
| DNA aptamer AS1411 gold NPs | Passive diffusion | Increased uptake in nanostars as compared to nanospheres in cancer cells | [285] | |
| Gold NPs | Clathrin-mediated/caveolae-mediated endocytosis | Increased uptake of Au nanospheres than the corresponding Au nanostars | [294] | |
| DNA-origami shaped designer NPs | Endocytosis | Larger particles with moderate aspect ratio (width: height) showed increased uptake than elongated particles with high aspect ratio | [295] | |
| Methyl-polyethylene glycol-gold NPs | Clathrin/caveolae-mediated uptake and lipid raft-mediated endocytosis | Increased uptake in stars, intermediate in rods, and least in triangles | [296] | |
| Hydrophobicity | Amphiphilic nanogels | Passive transport | Hydrophobic NPs displayed increased uptake and protein binding in THP-1 cells | [297] |
| Lutein stevioside NP | Passive diffusion/clathrin-mediated endocytosis/caveolae/lipid raft dependent endocytosis | Entrapment of lutein in the hydrophobic core of stevioside caused increased uptake and bioavailability of lutein | [298] | |
| Star shaped polymer-imidaclothiz NP | Clathrin-mediated endocytosis | Entrapment of imidaclothiz in the lipophilic core of star polymer, increased plant uptake of imidaclothiz | [286] | |
| Liposomal silk fibroin and sodium alginate (https://www.sciencedirect.com/topics/chemical-engineering/sodium-alginate)-dimethylcurcumin NP | Endocytosis | Combination of silk fibroin and sodium alginate (https://www.sciencedirect.com/topics/chemical-engineering/sodium-alginate) provide a platform for loading poorly water-soluble dimethylcurcumin (anticancer activity) through hydrophobic interactions | [299] | |
| Propranolol-chitosan nanogels | Endocytic uptake | Hydrophobic drug reduced size of nanogel with increased uptake | [300] | |
| Surface chemistry | Anionic polystyrene NPs | Clathrin-mediated medocytosis and micropinocytosis (cations); clathrin-mediated and lipid raft-dependent internalization (anions) | Increased uptake of anionic particles due to cleavage of heparan sulfate from cell surface | [301] |
| Ultrasmall superparamagnetic iron oxide NPs (USPION) | Endocytosis | Increased uptake of amine-functionalized USPION as compared to carboxyl-functionalized USPION | [287] | |
| Superparamagnetic iron-oxide NPs (SPIONs) | lysosomal pathway | Increased uptake of aminated SPIONs and as compared to polyethylene glycolated SPIONS | [302] | |
| Mesoporous silica NPs and up/downconverting NPs | Endocytosis | Increased uptake of negatively charged particles in acidic pH and conversion to positive charge | [303] | |
| Thermosensitive liposomal NPs with surface polyethylene glycol/phospholipid 1,2-dipalmitoyl-sn-glycero-3-phosphodiglycerol (DPPG2) | Endocytosis | DPPG2 functionalized anionic NPs exhibited increased uptake of doxorubicin in cancer cells | [304] | |
| Capping | Gold NPs | Clathrin-mediated/caveolae-mediated endocytosis | 11-mercaptoundecanoic acid (MUA)-coated particles were more greatly internalized relative to citrate-coated particles | [294] |
| Gold NPs | Clathrin-mediated/caveolae-mediated endocytosis | Carboxyl-polyethylene glycol-thiol (PEG) capped particles showed increased uptake in cancer cells | [305] | |
| Gold NPs | Endocytosis | Increased uptake of Au released from sodium warfarin capped particles | [288] | |
| Zinc oxide NPs | Endocytosis | L-cysteine capped particles increased uptake in cancer cells | [306] | |
| Gold NPs | Transcytosis | Increased uptake caused by albumin-capped particles | [307] |
| Physicochemical Properties | Nanoparticles | Response of Immune System | Reference |
|---|---|---|---|
| Size | NPs | Regulation of innate immunity (activation of Kupffer cells and B cells) increased in large (100–250 nm) and decreased in medium (10–100 nm) sized NPs | [318] |
| Silver, silica, titanium, and magnetic NPs | Increased IL-8 and IL-1β cytokine secretion by all NPs except silver NPs | [319] | |
| Shape | Gold NPs | Increased immune response and IgG production in spherical and star shaped | [320] |
| Polyethylene glycol gold NPs | Nanorods increased inflammasome activation | [321] | |
| Hydrophobicity | Synthetic-HDL NPs | Increased immune response of hydrophobic liver X receptor agonist | [322] |
| SARS-CoV-2 spike protein B-cell epitome (S461-493)-gold NPs | Increased IgG responses when compared to sole S461-493 with increased released of cytokines | [323] | |
| Coating | Cisplatin -ovalbumin-coated iron oxide NPs | Increased macrophage polarization and proinflammatory cytokines | [324] |
| Polyethylene glycol coated NPs | Increased lymphatic transport and modulate immune response | [325] | |
| Surface charge | Silica NPs | Amine functionalized particles decreased monocyte/macrophages activation (nitrite secretion, CD40/CD80, and pro-inflammatory cytokines) | [326] |
| Silica NPs | Hydrocarbon groups at the surface formed albumin rich corona, increased pro-inflammatory cytokines (IL6 and TNFα) | [327] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bezbaruah, R.; Chavda, V.P.; Nongrang, L.; Alom, S.; Deka, K.; Kalita, T.; Ali, F.; Bhattacharjee, B.; Vora, L. Nanoparticle-Based Delivery Systems for Vaccines. Vaccines 2022, 10, 1946. https://doi.org/10.3390/vaccines10111946
Bezbaruah R, Chavda VP, Nongrang L, Alom S, Deka K, Kalita T, Ali F, Bhattacharjee B, Vora L. Nanoparticle-Based Delivery Systems for Vaccines. Vaccines. 2022; 10(11):1946. https://doi.org/10.3390/vaccines10111946
Chicago/Turabian StyleBezbaruah, Rajashri, Vivek P. Chavda, Lawandashisha Nongrang, Shahnaz Alom, Kangkan Deka, Tutumoni Kalita, Farak Ali, Bedanta Bhattacharjee, and Lalitkumar Vora. 2022. "Nanoparticle-Based Delivery Systems for Vaccines" Vaccines 10, no. 11: 1946. https://doi.org/10.3390/vaccines10111946
APA StyleBezbaruah, R., Chavda, V. P., Nongrang, L., Alom, S., Deka, K., Kalita, T., Ali, F., Bhattacharjee, B., & Vora, L. (2022). Nanoparticle-Based Delivery Systems for Vaccines. Vaccines, 10(11), 1946. https://doi.org/10.3390/vaccines10111946