Insight of Polyphenol Oxidase Enzyme Inhibition and Total Polyphenol Recovery from Cocoa Beans

 ,
, 
Abstract
1. Introduction
2. Materials and Methods
2.1. Reagents
2.2. Recovery of Cocoa Polyphenol Extract
2.3. Enzyme Inhibition
2.4. PPO Enzyme Extraction
2.5. PPO Enzyme Activity Measurement
2.6. Substrate Kinetic Constants of PPO
2.7. Recovery of Total Phenol Content
2.8. Determination of Total Polyphenol Content
2.9. Chromatographic Analysis by HPLC-DAD
2.10. Scanning Electron Microscopy and Image Analysis
2.11. Statistical Analysis
3. Results and Discussion
3.1. Preliminary Inhibition Assays
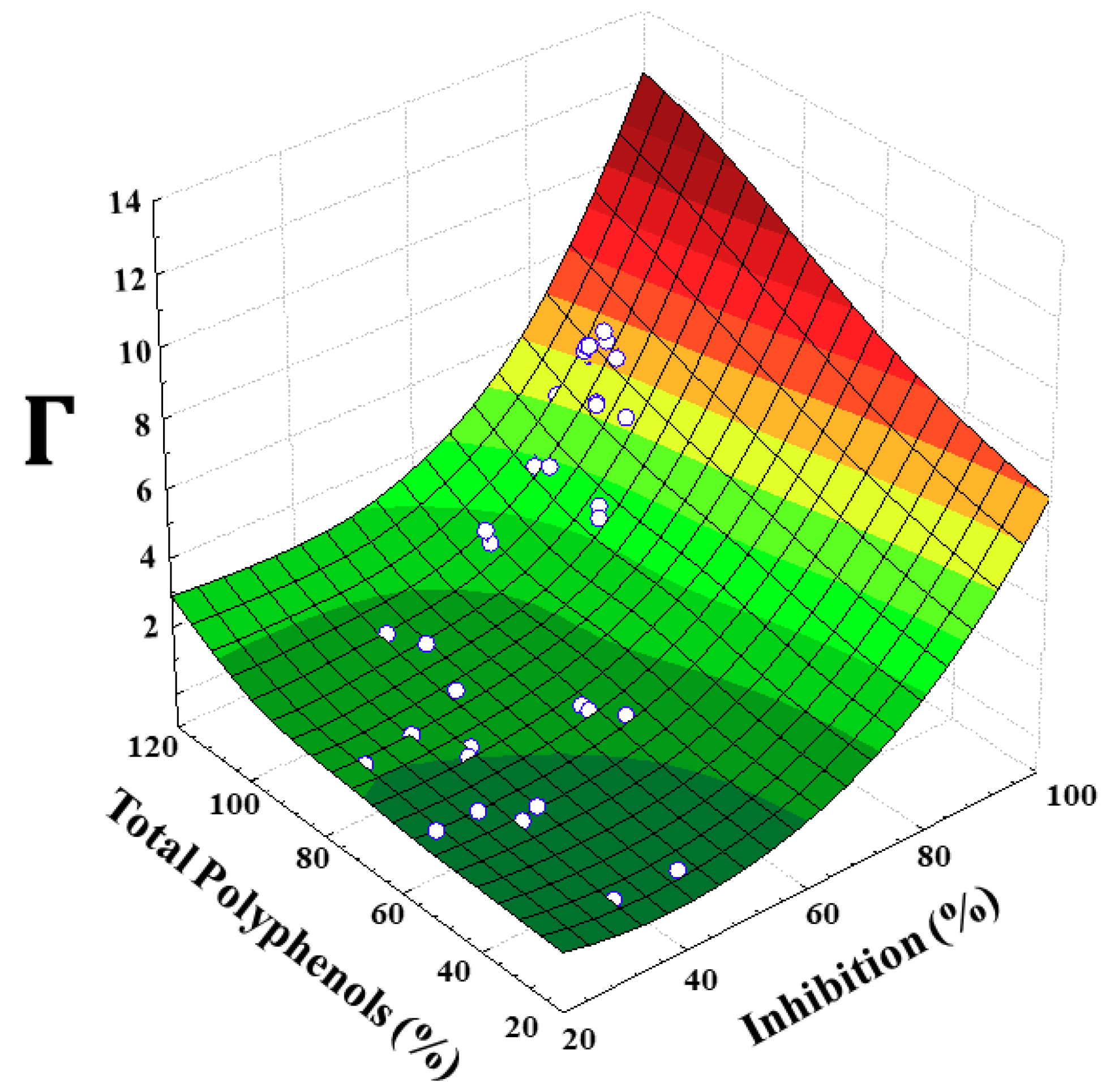
3.2. Influence of Inhibition Parameters on PPO Activity
3.3. Effects of Temperature, Time and Concentration of Inhibitor on PPO Activity
3.4. Kinetic Parameters of PPO Inhibition in Cocoa Beans
3.5. Microscopy Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Macedo, A.S.L.; de Rocha, F.S.; da Ribeiro, M.S.; Soares, S.E.; da Bispo, E.S. Characterization of polyphenol oxidase in two cocoa (Theobroma cacao L.) cultivars produced in the south of Bahia, Brazil. Food Sci. Technol. 2016, 36, 56–63. [Google Scholar] [CrossRef]
- Doǧru, Y.Z.; Erat, M. Investigation of some kinetic properties of polyphenol oxidase from parsley (Petroselinum crispum, Apiaceae). Food Res. Int. 2012, 49, 411–415. [Google Scholar] [CrossRef]
- De Oliveira Carvalho, J.; Orlanda, J.F.F. Heat stability and effect of pH on enzyme activity of polyphenol oxidase in buriti (Mauritia flexuosa Linnaeus f.) fruit extract. Food Chem. 2017, 233, 159–163. [Google Scholar] [CrossRef] [PubMed]
- Li, F.; Chen, G.; Zhang, B.; Fu, X. Current applications and new opportunities for the thermal and non-thermal processing technologies to generate berry product or extracts with high nutraceutical contents. Food Res. Int. 2017, 100, 19–30. [Google Scholar] [CrossRef] [PubMed]
- Osuga, D.; Van Der Schaaf, A.; Whitaker, J.R. Control of polyphenol oxidase activity using a catalytic mechanism. In Protein Structure-Function Relationships in Foods; Springer: Boston, MA, USA, 1994; pp. 62–88. ISBN 978-1-4615-2670-4. [Google Scholar]
- Araji, S.; Grammer, T.A.; Gertzen, R.; Anderson, S.D.; Mikulic-Petkovsek, M.; Veberic, R.; Phu, M.L.; Solar, A.; Leslie, C.A.; Dandekar, A.M.; et al. Novel roles for the polyphenol oxidase enzyme in secondary metabolism and the regulation of cell death in walnut. Plant Physiol. 2014, 164, 1191–1203. [Google Scholar] [CrossRef] [PubMed]
- Selamat, J.; Bakar, J.; Saari, N. Oxidation of polyphenols in unfermented and partly fermented cocoa beans by cocoa polyphenol oxidase and tyrosinase. J. Sci. Food Agric. 2002, 82, 559–566. [Google Scholar]
- Cheema, S.; Sommerhalter, M. Characterization of polyphenol oxidase activity in Ataulfo mango. Food Chem. 2015, 171, 382–387. [Google Scholar] [CrossRef]
- De Jesus, A.L.T.; Leite, T.S.; Cristianini, M. High isostatic pressure and thermal processing of açaí fruit (Euterpe oleracea Martius): Effect on pulp color and inactivation of peroxidase and polyphenol oxidase. Food Res. Int. 2018, 105, 853–862. [Google Scholar] [CrossRef]
- De la Rosa, L.A.; Alvarez-Parrilla, E.; González-Aguilar, G.A. Fruit and Vegetable Phytochemicals: Chemistry, Nutritional Value, and Stability, 1st ed.; De la Rosa, L.A., Ed.; Wiley-Blackwell: Ames, Iowa, 2009; ISBN 9780813803203. [Google Scholar]
- He, Q.; Luo, Y.; Chen, P. Elucidation of the mechanism of enzymatic browning inhibition by sodium chlorite. Food Chem. 2008, 110, 847–851. [Google Scholar] [CrossRef]
- Mishra, B.B.; Gautam, S.; Sharma, A. Free phenolics and polyphenol oxidase (PPO): The factors affecting post-cut browning in eggplant (Solanum melongena). Food Chem. 2013, 139, 105–114. [Google Scholar] [CrossRef]
- De Leonardis, A.; Angelico, R.; Macciola, V.; Ceglie, A. Effects of polyphenol enzymatic-oxidation on the oxidative stability of virgin olive oil. Food Res. Int. 2013, 54, 2001–2007. [Google Scholar] [CrossRef]
- De Brito, E.S.; García, N.H.P.; Amâncio, A.C. Effect of polyphenol oxidase (PPO) and air treatments on total phenol and tannin content of cocoa nibs. Food Sci. Technol. 2002, 22, 45–48. [Google Scholar] [CrossRef]
- Kruger, M.J.; Davies, N.; Myburgh, K.H.; Lecour, S. Proanthocyanidins, anthocyanins and cardiovascular diseases. Food Res. Int. 2014, 59, 41–52. [Google Scholar] [CrossRef]
- Schinella, G.; Mosca, S.; Cienfuegos-Jovellanos, E.; Pasamar, M.Á.; Muguerza, B.; Ramón, D.; Ríos, J.L. Antioxidant properties of polyphenol-rich cocoa products industrially processed. Food Res. Int. 2010, 43, 1614–1623. [Google Scholar] [CrossRef]
- Nakajima, V.M.; Moala, T.; Caria, C.R.e.P.; Moura, C.S.; Amaya-Farfan, J.; Gambero, A.; Macedo, G.A.; Macedo, J.A. Biotransformed citrus extract as a source of anti-inflammatory polyphenols: Effects in macrophages and adipocytes. Food Res. Int. 2017, 97, 37–44. [Google Scholar] [CrossRef]
- Marchese, A.; Coppo, E.; Sobolev, A.P.; Rossi, D.; Mannina, L.; Daglia, M. Influence of in vitro simulated gastroduodenal digestion on the antibacterial activity, metabolic profiling and polyphenols content of green tea (Camellia sinensis). Food Res. Int. 2014, 63, 182–191. [Google Scholar] [CrossRef]
- Karar, M.G.E.; Pletzer, D.; Jaiswal, R.; Weingart, H.; Kuhnert, N. Identification, characterization, isolation and activity against Escherichia coli of quince (Cydonia oblonga) fruit polyphenols. Food Res. Int. 2014, 65, 121–129. [Google Scholar] [CrossRef]
- Alañón, M.E.; Castle, S.M.; Siswanto, P.J.; Cifuentes-Gómez, T.; Spencer, J.P.E. Assessment of flavanol stereoisomers and caffeine and theobromine content in commercial chocolates. Food Chem. 2016, 208, 177–184. [Google Scholar] [CrossRef] [PubMed]
- Lamuela-Raventós, R.M.; Romero-Pérez, A.I.; Andrés-Lacueva, C.; Tornero, A. Review: Health effects of cocoa flavonoids. Food Sci. Technol. Int. 2005, 11, 159–176. [Google Scholar] [CrossRef]
- Pedan, V.; Fischer, N.; Bernath, K.; Hühn, T.; Rohn, S. Determination of oligomeric proanthocyanidins and their antioxidant capacity from different chocolate manufacturing stages using the NP-HPLC-online-DPPH methodology. Food Chem. 2017, 214, 523–532. [Google Scholar] [CrossRef]
- Guerrero-Beltrán, J.A.; Swanson, B.G.; Barbosa-Cánovas, G.V. Inhibition of polyphenoloxidase in mango puree with 4-hexylresorcinol, cysteine and ascorbic acid. LWT Food Sci. Technol. 2005, 38, 625–630. [Google Scholar] [CrossRef]
- Pizzocaro, F.; Torreggiani, D.; Gilardi, G. Inhibition of apple polyphenol oxidase (ppo) by ascorbic acid, citric acid and sodium chloride. J. Food Process. Preserv. 1993, 17, 21–30. [Google Scholar] [CrossRef]
- Menon, A.S.; Hii, C.L.; Law, C.L.; Suzannah, S.; Djaeni, M. Effects of water blanching on polyphenol reaction kinetics and quality of cocoa beans. In AIP Conference Proceedings; AIP Publishing LLC: Melville, NY, USA, 2015; Volume 1699, pp. 030006–1–030006–7. [Google Scholar]
- Ali, H.M.; El-Gizawy, A.M.; El-Bassiouny, R.E.I.; Saleh, M.A. Browning inhibition mechanisms by cysteine, ascorbic acid and citric acid, and identifying PPO-catechol-cysteine reaction products. J. Food Sci. Technol. 2015, 52, 3651–3659. [Google Scholar] [CrossRef] [PubMed]
- Lee, C.Y.; Whitaker, J.R. (Eds.) Enzymatic Browning and Its Prevention, 1st ed.; ACS Symposium Series; American Chemical Society: Washington, DC, USA, 1995; Volume 600, ISBN 0-8412-3249-0. [Google Scholar]
- Siddiq, M.; Dolan, K.D. Characterization and heat inactivation kinetics of polyphenol oxidase from blueberry (Vaccinium corymbosum L.). Food Chem. 2017, 218, 216–220. [Google Scholar] [CrossRef]
- Pons-Andreu, J.-V.; Cienfuegos-Jovellanos, E.; Ibarra, A. Process for Producing Cocoa Polyphenol Concentrate. U.S. Patent 0193629 A1, 2008. [Google Scholar]
- Manzocco, L.; Anese, M.; Nicoli, M.C. Radiofrequency inactivation of oxidative food enzymes in model systems and apple derivatives. Food Res. Int. 2008, 41, 1044–1049. [Google Scholar] [CrossRef]
- Yuan, G.; Lv, H.; Tang, W.; Zhang, X.; Sun, H. Effect of chitosan coating combined with pomegranate peel extract on the quality of Paci fi c white shrimp during iced storage. Food Control 2016, 59, 818–823. [Google Scholar] [CrossRef]
- Babu, B.R.; Rastogi, N.K.; Raghavarao, K.S.M.S. Liquid-liquid extraction of bromelain and polyphenol oxidase using aqueous two-phase system. Chem. Eng. Process. Process Intensif. 2008, 47, 83–89. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- AOCS. Official Methods of Analysis of AOAC International; AOCS: Washington, DC, USA, 1990; ISBN 0935584544. [Google Scholar]
- Singleton, V.L.; Orthofer, R.; Lamuela-Raventós, R.M. Analysis of total phenols and other oxidation substrates and antioxidants by means of folin-ciocalteu reagent. Methods Enzymol. 1998, 299, 152–178. [Google Scholar]
- Hernández-Carrión, M.; Hernando, I.; Sotelo-Díaz, I.; Quintanilla-Carvajal, M.X.; Quiles, A. Use of image analysis to evaluate the effect of high hydrostatic pressure and pasteurization as preservation treatments on the microstructure of red sweet pepper. Innov. Food Sci. Emerg. Technol. 2015, 27, 69–78. [Google Scholar] [CrossRef]
- Lee, P.M.; Lee, K.H.; Karim, M.I.A. Biochemical-Studies of Cocoa Bean Polyphenol Oxidase. J. Sci. Food Agric. 1991, 55, 251–260. [Google Scholar] [CrossRef]
- Dudley, E.D.; Hotchkiss, J.M. Cysteine as an inhibitor of polyphenoloxidase. J. Food Biochem. 1989, 13, 65–75. [Google Scholar] [CrossRef]
- Wuyts, N.; De Waele, D.; Swennen, R. Extraction and partial characterization of polyphenol oxidase from banana (Musa acuminata Grande naine) roots. Plant Physiol. Biochem. 2006, 44, 308–314. [Google Scholar] [CrossRef] [PubMed]
- Ho, K.K. Characterization of polyphenol oxidase from aerial roots of an orchid, Aranda “Christine 130”. Plant Physiol. Biochem. 1999, 37, 841–848. [Google Scholar] [CrossRef]
- Liu, L.; Cao, S.; Xie, B.; Sun, Z.; Li, X.; Miao, W. Characterization of polyphenol oxidase from litchi pericarp using (−)-epicatechin as substrate. J. Agric. Food Chem. 2007, 55, 7140–7143. [Google Scholar] [CrossRef]
- Martinez-Cayuela, M.; De Medina, L.S.; Faus, M.J.; GilL, A. Cherimoya (Annona cherimola Mill) polyphenoloxidase: Monophenolase and dihydroxyphenolase activities. J. Food Sci. 1988, 53, 1191–1194. [Google Scholar] [CrossRef]
- Dincer, B.; Colak, A.; Aydin, N.; Kadioglu, A.; Güner, S. Characterization of polyphenoloxidase from medlar fruits (Mespilus germanica L., Rosaceae). Food Chem. 2002, 77, 1–7. [Google Scholar] [CrossRef]
- Terefe, N.S.; Delon, A.; Buckow, R.; Versteeg, C. Blueberry polyphenol oxidase: Characterization and the kinetics of thermal and high pressure activation and inactivation. Food Chem. 2015, 188, 193–200. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Run | T (°C) | Time (min) | Inhibitor [mM] | Specific Activity [UPPO/mg] | Total Polyphenol [mg GAE/g] | Γ * |
|---|---|---|---|---|---|---|
| 1 | 90 | 1 | 50 | 8.97 ± 0.08 | 86.58 ± 2.53 | 3.74 |
| 2 | 70 | 5 | 0 | 18.48 ± 0.49 | 83.37 ± 1.45 | 1.68 |
| 3 | 63 | 3 | 25 | 8.40 ± 0.45 | 80.52 ± 3.21 | 3.45 |
| 4 | 90 | 1 | 0 | 20.42 ± 0.05 | 70.48 ± 0.90 | 1.05 |
| 5 | 90 | 5 | 0 | 12.30 ± 0.52 | 84.74 ± 4.65 | 2.61 |
| 6 | 80 | 3 | 67 | 6.86 ± 0.99 | 88.20 ± 3.43 | 5.07 |
| 7 | 80 | 3 | 25 | 15.05 ± 0.49 | 69.24 ± 1.22 | 1.36 |
| 8 | 80 | 3 | 25 | 18.42 ± 0.66 | 75.84 ± 1.34 | 1.38 |
| 9 | 80 | 1 | 25 | 20.08 ± 0.52 | 63.62 ± 4.50 | 0.81 |
| 10 | 70 | 1 | 0 | 21.11 ± 0.06 | 53.65 ± 2.32 | 0.41 |
| 11 | 70 | 1 | 50 | 21.15 ± 0.19 | 63.11 ± 1.01 | 0.75 |
| 12 | 80 | 6 | 25 | 5.60 ± 0.47 | 87.68 ± 3.21 | 6.14 |
| 13 | 70 | 5 | 50 | 5.17 ± 0.82 | 85.08 ± 2.21 | 6.27 |
| 14 | 80 | 3 | 25 | 14.03 ± 0.70 | 66.52 ±2.56 | 1.31 |
| 15 | 90 | 5 | 0 | 12.04 ± 0.41 | 85.58 ± 6.65 | 2.72 |
| 16 | 97 | 3 | 25 | 5.09 ± 0.36 | 86.32 ± 3.27 | 6.55 |
| 17 | 90 | 5 | 50 | 6.22 ± 0.90 | 85.21 ± 4.05 | 5.23 |
| 18 | 97 | 3 | 25 | 5.00 ± 0.31 | 86.73 ± 2.41 | 6.74 |
| 19 | 80 | 3 | 25 | 14.94 ± 0.07 | 68.57 ± 4.10 | 1.34 |
| 20 | 70 | 5 | 50 | 5.50 ± 1.14 | 87.67 ± 1.43 | 6.25 |
| 21 | 80 | 3 | 0 | 20.01 ± 0.23 | 71.01 ± 1.32 | 1.09 |
| 22 | 80 | 1 | 25 | 22.16 ± 1.14 | 65.75 ± 0.53 | 0.81 |
| 23 | 63 | 3 | 25 | 8.69 ± 0.60 | 79.99 ± 4.32 | 3.29 |
| 24 | 90 | 5 | 50 | 6.30 ± 0.41 | 85.15 ± 4.21 | 5.15 |
| 25 | 80 | 6 | 25 | 5.43 ± 0.16 | 87.50 ± 5.01 | 6.30 |
| 26 | 70 | 1 | 0 | 18.09 ± 0.04 | 53.24 ± 2.45 | 0.46 |
| 27 | 80 | 3 | 0 | 23.99 ± 0.79 | 74.21 ± 1.23 | 1.01 |
| 28 | 90 | 1 | 50 | 8.62 ± 0.86 | 85.60 ± 3.02 | 3.81 |
| 29 | 90 | 1 | 0 | 21.36 ± 0.43 | 74.44 ± 2.03 | 1.14 |
| 30 | 70 | 1 | 50 | 24.22 ± 0.26 | 65.94 ± 2.01 | 0.74 |
| 31 | 80 | 3 | 67 | 5.93 ± 0.33 | 82.54 ± 2.98 | 5.14 |
| 32 | 70 | 5 | 0 | 17.54 ± 0.78 | 80.84 ± 3.89 | 1.67 |
| Factor | SS | df | MS | p |
|---|---|---|---|---|
| T | 8.3298 | 1 | 8.32985 | 0.003001 |
| T2 | 19.4339 | 1 | 19.43392 | 0.000042 |
| t | 49.5296 | 1 | 49.52962 | 0.000000 |
| t2 | 2.2011 | 1 | 2.20111 | 0.100515 |
| Inh | 35.4793 | 1 | 35.47934 | 0.000001 |
| Inh2 | 0.0185 | 1 | 0.01846 | 0.876667 |
| T × t | 3.5258 | 1 | 3.52580 | 0.041099 |
| T × Inh | 0.0242 | 1 | 0.02417 | 0.859066 |
| t × Inh | 4.2416 | 1 | 4.24158 | 0.026414 |
| Error | 16.4760 | 22 | 0.74891 | |
| Total SS | 150.9190 | 31 |
| Substrate | Km [mM] | vmax (UPPO mL−1 min−1) | vmax/Km (UPPO/mM mL min) |
|---|---|---|---|
| Catechol | 0.61 ± 0.12 a | 2709 ± 21.89 a | 4440.98 |
| Catechol * | 8.36 ± 1.33 b | 106.7 ± 3.70 b | 12.76 |
| (−)-Epicatechin | 1.26 ± 0.37 a | 916.5 ± 20.59 c | 727.38 |
| (+)-Catechin | 1.45 ± 0.35 a | 924.8 ± 18.98 c | 637.79 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Toro-Uribe, S.; Godoy-Chivatá, J.; Villamizar-Jaimes, A.R.; Perea-Flores, M.d.J.; López-Giraldo, L.J. Insight of Polyphenol Oxidase Enzyme Inhibition and Total Polyphenol Recovery from Cocoa Beans. Antioxidants 2020, 9, 458. https://doi.org/10.3390/antiox9060458
Toro-Uribe S, Godoy-Chivatá J, Villamizar-Jaimes AR, Perea-Flores MdJ, López-Giraldo LJ. Insight of Polyphenol Oxidase Enzyme Inhibition and Total Polyphenol Recovery from Cocoa Beans. Antioxidants. 2020; 9(6):458. https://doi.org/10.3390/antiox9060458
Chicago/Turabian StyleToro-Uribe, Said, Jhair Godoy-Chivatá, Arley René Villamizar-Jaimes, María de Jesús Perea-Flores, and Luis J. López-Giraldo. 2020. "Insight of Polyphenol Oxidase Enzyme Inhibition and Total Polyphenol Recovery from Cocoa Beans" Antioxidants 9, no. 6: 458. https://doi.org/10.3390/antiox9060458
APA StyleToro-Uribe, S., Godoy-Chivatá, J., Villamizar-Jaimes, A. R., Perea-Flores, M. d. J., & López-Giraldo, L. J. (2020). Insight of Polyphenol Oxidase Enzyme Inhibition and Total Polyphenol Recovery from Cocoa Beans. Antioxidants, 9(6), 458. https://doi.org/10.3390/antiox9060458