Spiraea prunifolia var. simpliciflora Attenuates Oxidative Stress and Inflammatory Responses in a Murine Model of Lipopolysaccharide-Induced Acute Lung Injury and TNF-α-Stimulated NCI-H292 Cells

 ,
,
Abstract
1. Introduction
2. Materials and Methods
2.1. UPLC Q-TOF/MS Analysis
2.2. Animal Husbandry
2.3. LPS-induced ALI Model and Differential Cell Count in Bronchoalveolar Lavage Fluid (BALF) Collection
- Normal control (NC) group: treated with vehicle (2% DMSO) from day 0 to day 5 and given 50 μL PBS without LPS on day 3
- LPS group: treated with vehicle (2% DMSO) from day 0 to day 5 and given LPS 20 μg in 50 μL PBS on day 3
- DEX group: treated with DEX 3 mg/kg only from day 0 to day 5 and given LPS 20 μg in 50 μL PBS on day 3
- SP50 group: treated with SP 50 mg/kg only and given LPS 20 μg in 50 μL PBS on day 3.
- SP100 group: treated with SP 100 mg/kg only and given LPS 20 μg in 50 μL PBS on day 3.
2.4. Measurement of Cytokines and Protein Contents in BALF
2.5. Lung Tissue Histopathology
2.6. Immunoblotting
2.7. Oxidative Stress Markers Analysis
2.8. Effects of SP on ROS Production and 2,2-Diphenyl-1-Picryl Hydrazyl (DPPH) Radical Scavenging Activity
2.9. Cell Culture and Cell Viability
2.10. Effects of SP on Pro-Inflammatory Cytokine Levels and Oxidative Stress Markers in TNF-α-stimulated NCI-H292 Cells
2.11. Effects of SP on MAPKs and NF-κB in TNF-α-stimulated NCI-H292 Cells
2.12. Effects of SP on Nrf2 and HO-1 in TNF-α-Stimulated NCI-H292 Cells
2.13. Quantitative Real-Time Polymerase Chain Reaction (PCR)
2.14. Statistical Analysis
3. Results
3.1. Tentative Characterization of SP Extract
3.2. SP Treatment Decreases Inflammatory Cells and Pro-Inflammatory Cytokine Contents in BALF
3.3. SP Treatment Attenuates LPS-Induced Lung Histological Changes
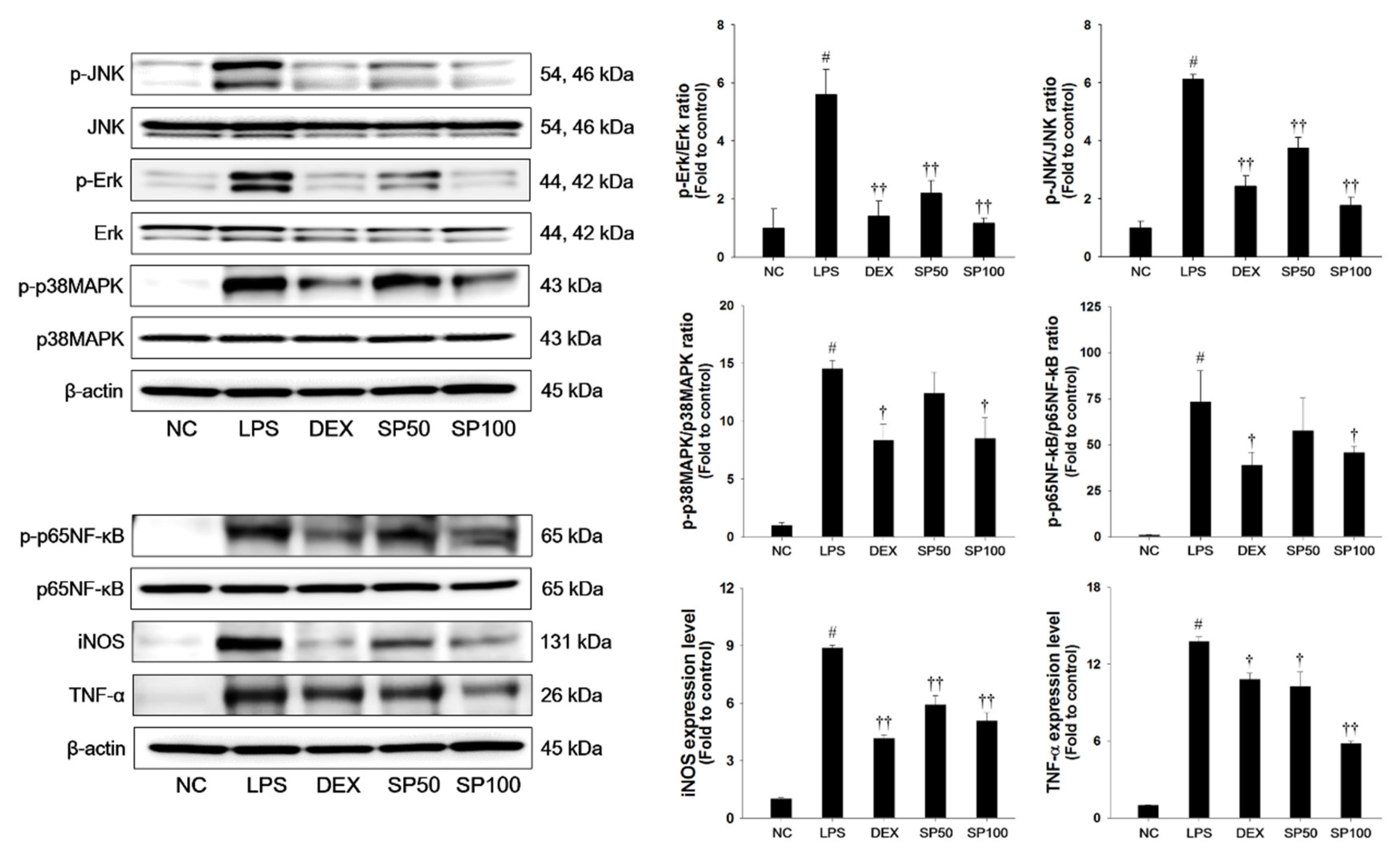
3.4. SP Treatment Suppresses LPS-Induced MAPKs, NF-κB Activation and Inflammatory Mediators in Lung Tissues
3.5. SP Treatment Decreases Lipid Peroxidation and Induces Nrf2 Pathway and Antioxidant Enzymes in Lung Tissues
3.6. SP Treatment Reduces Pro-Inflammatory Cytokine Production in TNF-α-stimulated NCI-H292 Cells
3.7. SP Treatment Decreases Phosphorylation of MAPKs and NF-κB Activation in TNF-α-Stimulated NCI-H292 Cells
3.8. SP Treatment Activates Nrf2 Pathways and Decreases Oxidative Stress, ROS Production and DPPH Radicals in TNF-α-stimulated NCI-H292 Cells
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| ALI | Acute lung injury |
| BALF | Bronchoalveolar lavage fluid |
| DEX | Dexamethasone |
| ELISA | Enzyme-linked immunosorbent assay |
| FBS | Fetal bovine serum |
| IL-1β | Interleukin-1β |
| IL-6 | Interleukin-6 |
| iNOS | Inducible nitric oxide synthase |
| LPS | Lipopolysaccharide |
| MAPKs | Mitogen-activated protein kinases |
| NF-κB | Nuclear factor-kappaB |
| NO | Nitric oxide |
| PCR | Polymerase chain reaction |
| SP | Spiraea prunifolia var. simpliciflora |
| TNF-α | Tumor necrosis factor-α |
| WST-1 | Water-soluble tetrazolium salt-1 |
| UPLC-QTOF/MS | Ultra-high performance liquid chromatography-quadrupole time-of-flight mass spectrometry |
| TLR | Toll-like receptor |
| Nrf2 | Nuclear factory erythroid-2 related factor 2 |
| HO-1 | Heme oxygenase-1 |
| NQO1 | NAD(P)H quinone dehydrogenase 1 |
| ROS | Reactive oxygen species |
| COPD | Chronic obstructive pulmonary disease |
| TBARS | Thiobarbituric acid-reactive substances |
| GSH | Glutathione |
| ARE | Antioxidant response element |
| Keap1 | Kelch-like ECH-associated protein 1 |
| ARDS | Acute respiratory distress syndrome |
References
- Liu, L.; Xiong, H.; Ping, J.; Ju, Y.; Zhang, X. Taraxacum officinale protects against lipopolysaccharide-induced acute lung injury in mice. J. Ethnopharmacol. 2010, 130, 392–397. [Google Scholar] [CrossRef]
- Xie, X.; Sun, S.; Zhong, W.; Soromou, L.W.; Wei, M.; Ren, Y.; Ding, Y. Zingerone attenuates lipopolysaccharide-induced acute lung injury in mice. Int. Immunopharmacol. 2014, 19, 103–109. [Google Scholar] [CrossRef]
- Jeyaseelan, S.; Chu, H.W.; Young, S.K.; Freeman, M.W.; Worthen, G.S. Distinct roles of pattern recognition receptors CD14 and toll-like receptor 4 in acute lung injury. Infect. Immun. 2005, 73, 1754–1763. [Google Scholar] [CrossRef] [PubMed]
- Piantadosi, C.A.; Schwartz, D.A. The acute respiratory distress syndrome. Ann. Intern. Med. 2004, 141, 460–470. [Google Scholar] [CrossRef] [PubMed]
- Eliana, C.; Eliana, Z.; Ewerton, P.; Ana Maria, G.; Carmen, B. Impact of distinct definitions of acute lung injury on its incidence and outcomes in Brazilian ICUs: Prospective evaluation of 7,133 patients. Crit. Care Med. 2014, 42, 574–582. [Google Scholar]
- Chen, J.; Wang, J.B.; Yu, C.H.; Chen, L.Q.; Xu, P.; Yu, W.Y. Total flavonoids of Mosla scabra leaves attenuates lipopolysaccharide-induced acute lung injury via down-regulation of inflammatory signaling in mice of inflammatory signaling in mice. J. Ethnopharmacol. 2013, 148, 835–841. [Google Scholar] [CrossRef]
- Shin, N.R.; Shin, I.S.; Song, H.H.; Hong, J.M.; Kwon, O.K.; Jeon, C.M.; Kim, J.H.; Lee, S.W.; Lee, J.K.; Jin, H.; et al. Callicarpa japonica Thunb. reduces inflammatory responses: A mouse model of lipopolysaccharide-induced acute lung injury. Int. Immunopharmacol. 2015, 26, 174–180. [Google Scholar] [CrossRef]
- Ho, Y.C.; Lee, S.S.; Yang, M.L.; Rosa, H.L.; Lee, C.Y.; Li, Y.C.; Kuan, Y.H. Zerumbone reduced the inflammatory response of acute lung injury in endotoxin treated mice via Akt-NF-κB pathway. Chem. Biol. Interact. 2017, 271, 9–14. [Google Scholar] [CrossRef]
- Tsai, C.L.; Lin, Y.C.; Wang, H.M.; Chou, T.C. Baicalein, an active component of Scutellaria baicalensis, protects against lipopolysaccharide-induced acute lung injury in rats. J. Ethnopharmacol. 2014, 153, 197–206. [Google Scholar] [CrossRef]
- Zhao, M.; Li, C.; Shen, F.; Wang, M.; Jia, N.; Wang, C. Naringenin ameliorates LPS-induced acute lung injury through its anti-oxidative and anti-inflammatory activity and by inhibition of the PI3K/AKT pathway. Exp. Ther. Med. 2017, 14, 2228–2234. [Google Scholar] [CrossRef]
- Fei, J.; Fu, L.; Hu, B.; Chen, Y.H.; Zhao, H.; Xu, D.X.; Li, J.B. Obeticholic acid alleviate lipopolysaccharide-induced acute lung injury via its anti-inflammatory effects in mice. Int. Immunopharmacol. 2019, 66, 177–184. [Google Scholar] [CrossRef] [PubMed]
- Chen, N.; Wu, Q.; Chi, G.; Soromou, L.W.; Hou, J.; Deng, Y.; Feng, H. Prime-O-glucosylcimifugin attenuates lipopolysaccharide-induced acute lung injury in mice. Int. Immunopharmacol. 2013, 16, 139–147. [Google Scholar] [CrossRef] [PubMed]
- Ding, H.; Ci, X.; Cheing, H.; Yu, Q.; Li, D. Chicoric acid alleviates lipopolysaccharide-induced acute lung injury in mice through anti-inflammatory and anti-oxidant activities. Int. Immunopharmacol. 2019, 66, 169–176. [Google Scholar] [CrossRef] [PubMed]
- Huang, X.T.; Liu, W.; Zhou, Y.; Sun, M.; Yang, H.H.; Zhang, C.Y. Galectin-1 ameliorates lipopolysaccharide-induced acute lung injury via AMPK-Nrf2 pathway in mice. Free Radic. Biol. Med. 2020, 146, 222–233. [Google Scholar] [CrossRef]
- Yang, H.; Lv, H.; Li, H.; Ci, X.; Peng, L. Oridonin protects LPS-induced acute lung injury by modulating Nrf2-mediated oxidative stress and Nrf2-independent NLRP3 and NF-κB pathways. Cell Commun. Signal. 2019, 17, 62. [Google Scholar] [CrossRef]
- Lu, Y.; Yu, T.; Liu, J.; Gu, L. Vitexin attenuates lipopolysaccharide-induced acute lung injury by controlling the Nrf2 pathway. PLoS ONE 2018, 13, e0196405. [Google Scholar] [CrossRef]
- Liu, Q.; Lv, H.; Wen, Z.; Ci, X.; Peng, L. Isoliquiritigenin activates nuclear factor erythroid-2 related factor 2 to suppress the NOD-like receptor protein 3 inflammasome and inhibits the NF-kappB pathway in macrophages and in acute lung injury. Front. Immunol. 2017, 8, 1518. [Google Scholar] [CrossRef]
- Lv, H.; Yu, Z.; Zheng, Y.; Wang, L.; Qin, X.; Cheng, G.; Ci, X. Isovitexin exerts anti-inflammatory and anti-oxidant activities on lipopolysaccharide-induced acute lung injury by inhibiting MAPK and NF-κB and activating HO-1/Nrf2 pathways. Int. J. Biol. Sci. 2016, 12, 72–86. [Google Scholar] [CrossRef]
- Wei, D.; Huang, Z. Anti-inflammatory effects of triptolide in LPS-induced acute lung injury in mice. Inflammation 2014, 37, 1307–1316. [Google Scholar] [CrossRef]
- Oh, H.; Oh, G.S.; Seo, W.G.; Pae, H.O.; Chai, K.Y.; Kwon, T.O.; Lee, Y.H.; Chung, H.T.; Lee, H.S. Prunioside A: A new terpene glycoside from Spiraea prunifolia. J. Nat. Prod. 2001, 64, 942–944. [Google Scholar] [CrossRef]
- Park, S.H.; Park, K.H.; Oh, M.H.; Kim, H.H.; Choe, K.I.; Kim, S.R.; Park, K.J.; Lee, M.W. Anti-oxidative and anti-inflammatory activities of caffeoyl hemiterpene glycosides from Spiraea prunifolia. Phytochemistry 2013, 96, 430–436. [Google Scholar] [CrossRef] [PubMed]
- So, H.S.; Park, R.; Oh, H.M.; Pae, H.O.; Lee, J.H.; Chai, K.Y.; Chung, S.Y.; Chung, H.T. The methanol extract of Spiraea prunifolia var. simpliciflora root inhibits the generation of nitric oxide and superoxide in RAW 264.7 cells. J. Ethnopharmacol. 1999, 68, 209–217. [Google Scholar] [CrossRef]
- Kim, C.S.; Oh, J.; Suh, W.S.; Jang, S.W.; Subedi, L.; Kim, S.Y.; Choi, S.U.; Lee, K.R. Investigation of chemical constituents from Spiraea prunifolia var. simpliciflora and their biological activities. Phytochem. Lett. 2017, 22, 255–260. [Google Scholar] [CrossRef]
- Card, J.W.; Carey, M.A.; Bradbury, J.A.; Degraff, L.M.; Morgan, D.L.; Moorman, M.P.; Flake, G.P.; Zeldin, D.C. Gender differences in murine airway responsiveness and lipopolysaccharide-induced inflammation. J. Immunol. 2006, 117, 621–630. [Google Scholar] [CrossRef] [PubMed]
- Ncube, E.N.; Mhlongo, M.I.; Piater, L.A.; Steenkamp, P.A.; Dubery, I.A.; Madala, N.E. Analyses of chlorogenic acids and related cinnamic acid derivatives from Nicotiana tabacum tissues with the aid of UPLC-QTOF-MS/MS based on the in-source collision-induced dissociation method. Chem. Cent. J. 2014, 8, 66. [Google Scholar] [CrossRef] [PubMed]
- Park, S.B.; Kang, J.Y.; Kim, J.M.; Park, S.K.; Park, S.H.; Kang, J.E.; Lee, C.J.; Kwon, B.S.; Yoo, S.K.; Lee, U.; et al. Aruncus dioicus var. kamtschaticus extract suppresses mitochondrial apoptosis induced-neurodegeneration in trimethyltin-injected ICR mice. J. Food Biochem. 2018, 42, e12667. [Google Scholar]
- Zimmermann, B.F.; Walch, S.G.; Tinzoh, L.N.; Stühlinger, W.; Lachenmeier, D.W. Rapid UHPLC determination of polyphenols in aqueous infusions of Salvia officinalis L. (sage tea). J. Chromatogr. B 2011, 879, 2459–2464. [Google Scholar] [CrossRef]
- Kajdžanoska, M.; Gjamovski, V.; Stefova, M. HPLC-DAD-ESI-MSn identification of phenolic compounds in cultivated strawberries from Macedonia. J. Chem. Chem. Eng. 2010, 29, 181–194. [Google Scholar] [CrossRef]
- Simirgiotis, M.J.; Benites, J.; Areche, C.; Sepúlveda, B. Antioxidant capacities and analysis of phenolic compounds in three endemic Nolana species by HPLC-PDA-ESI-MS. Molecules 2015, 20, 11490–11507. [Google Scholar] [CrossRef]
- Schieber, A.; Berardini, N.; Carle, R. Identification of flavonol and xanthone glycosides from mango (Mangifera indica L. Cv. "Tommy Atkins") peels by high-performance liquid chromatography-electrospray ionization mass spectrometry. J. Agric. Food Chem. 2003, 51, 5006–5011. [Google Scholar] [CrossRef]
- Jiménez-Aspee, F.; Theoduloz, C.; Soriano, M.D.P.C.; Ugalde-Arbizu, M.; Alberto, M.R.; Zampini, I.C.; Isla, M.I.; Simirigiotis, M.J.; Schmeda-Hirschmann, G. The native fruit Geoffroea decorticans from arid northern Chile: Phenolic composition, antioxidant activities and in vitro inhibition of pro-inflammatory and metabolic syndrome-associated enzymes. Molecules 2017, 22, 1565. [Google Scholar] [CrossRef] [PubMed]
- Jang, G.H.; Kim, H.W.; Lee, M.K.; Jeong, S.Y.; Bak, A.R.; Lee, D.J.; Kim, J.B. Characterization and quantification of flavonoid glycosides in the Prunus genus by UPLC-DAD-QTOF/MS. Saudi. J. Biol. Sci. 2018, 25, 1622–1631. [Google Scholar]
- Thabit, S.; Handoussa, H.; Roxo, M.; Cestari de Azevedo, B.; EL Sayed, N.S.; Wink, M. Evaluation of antioxidant and neuroprotective activities of Cassia fistula (L.) using the Caenorhabditis elegans model. Molecules 2019, 24, 2633. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.H.; Guo, H.; Xu, W.B.; Ge, J.; Li, X.; Alimu, M.; He, D.J. Rapid identification of flavonoid constituents directly from PTP1B inhibitive extract of raspberry (Rubus idaeus L.) leaves by HPLC-ESI-QTOF-MS-MS. J. Chromatogr. Sci. 2016, 54, 805–810. [Google Scholar] [CrossRef] [PubMed]
- Saluk-Juszczak, J.; Wachowicz, B. The proinflammatory activity of lipopolysaccharide. Postepy Biochem. 2005, 51, 280–287. [Google Scholar] [PubMed]
- Kitamura, Y.; Hashimoto, S.; Mizuta, N.; Kobayashi, A.; Kooguchi, K.; Fujiwara, I.; Nakajima, H. Fas/FasL-dependent apoptosis of alveolar cells after lipopolysaccharide-induced lung injury in mice. Am. J. Respir. Crit. Care Med. 2001, 163, 762–769. [Google Scholar] [CrossRef] [PubMed]
- Sung, J.; Sung, M.; Kim, Y.; Ham, H.; Jeong, H.S.; Lee, J. Anti-inflammatory effect of methanol extract from Erigeron canadensis L. may be involved with upregulation of heme oxygenase-1 expression and suppression of NFκB and MAPKs activation in macrophages. Nutr. Res. Pract. 2014, 8, 352–359. [Google Scholar] [CrossRef][Green Version]
- Chen, H.; Wu, S.; Lu, R.; Zhang, Y.G.; Zheng, Y.; Sun, J. Pulmonary permeability assessed by fluorescent-labeled dextran instilled intranasally into mice with LPS-induced acute lung injury. PLoS ONE 2014, 9, e101925. [Google Scholar] [CrossRef]
- Zhang, Y.; Du, Z.; Zhou, Q.; Wang, Y.; Li, J. Remifentanil attenuates lipopolysaccharide-induced acute lung injury by downregulating the NF-κB signaling pathway. Inflammation 2014, 37, 1654–1660. [Google Scholar] [CrossRef]
- Bhargava, R.; Altmann, C.J.; Andres-Hernando, A.; Webb, R.G.; Okamura, K.; Yang, Y.; Falk, S.; Schmidt, E.P.; Faubel, S. Acute lung injury and acute kidney injury are established by four hours in experimental sepsis and are improved with pre, but not post, sepsis administration of TNF-α antibodies. PLoS ONE 2013, 8, e79037. [Google Scholar] [CrossRef]
- Jnawali, H.N.; Lee, E.; Jeong, K.W.; Shin, A.; Heo, Y.S.; Kim, Y. Anti-inflammatory Activity of Rhamnetin and a model of its binding to c-Jun NH2-terminal kinase 1 and p38 MAPK. J. Nat. Prod. 2014, 77, 258–263. [Google Scholar] [CrossRef] [PubMed]
- Xia, M.Z.; Liang, Y.L.; Wang, H.; Chen, X.; Huang, Y.Y.; Zhang, Z.H.; Chen, Y.H.; Zhang, C.; Zhao, M.; Xu, D.X.; et al. Melatonin modulates TLR4-mediated inflammatory genes through MyD88-and TRIF-dependent signaling pathways in lipopolysaccharide stimulated RAW264.7 cells. J. Pineal Res. 2012, 53, 325–334. [Google Scholar] [CrossRef] [PubMed]
- Hong, J.M.; Kwon, O.K.; Shin, I.S.; Jeon, C.M.; Shin, N.R.; Lee, J.; Park, S.H.; Bach, T.T.; Hai, D.V.; Oh, S.R.; et al. Anti-inflammatory effects of methanol extract of Canarium lyi C.D. Dai & Yakovlev in RAW 264.7 macrophages and a murine model of lipopolysaccharide-induced lung injury. Int. J. Mol. Med. 2015, 35, 1403–1410. [Google Scholar] [PubMed]
- Gao, Y.; Jiang, W.; Dong, C.; Li, C.; Fu, X.; Min, L.; Tian, J.; Jin, H.; Shen, J. Anti-inflammatory effects of sophocarpine in LPS-induced RAW 264.7 cells via NF-κB and MAPKs signaling pathways. Toxicol. In Vitro. 2012, 26, 1–6. [Google Scholar] [CrossRef]
- Chen, X.; Yang, X.; Liu, T.; Guan, M.; Feng, X.; Dong, W.; Chu, X.; Liu, J.; Tian, X.; Ci, X.; et al. Kaempferol regulates MAPKs and NF-κB signaling pathways to attenuate LPS-induced acute lung injury in mice. Int. Immunopharmacol. 2012, 14, 209–216. [Google Scholar] [CrossRef]
- Chang, J.S.; Lin, H.J.; Deng, J.S.; Wu, W.T.; Huang, S.S.; Huang, G.J. Preventive effects of velvet antler (Cervus elaphus) against lipopolysaccharide-induced acute lung injury in mice by inhibiting MAPK/NF-κB activation and inducing AMPK/Nrf2 pathways. Evid. Based Complement. Alternat. Med. 2018, 2018, 2870503. [Google Scholar] [CrossRef]
- Tian, C.; Zhang, P.; Yang, J.; Zhang, Z.; Wang, H.; Guo, Y.; Liu, M. The protective effect of the flavonoid fraction of Abutilon theophrasti Medic. leaves on LPS-induced acute lung injury in mice via the NF-κB and MAPK signalling pathways. Biomed. Pharmacother. 2019, 109, 1024–1031. [Google Scholar] [CrossRef]
- Fengyang, L.; Yunhe, F.; Bo, L.; Zhicheng, L.; Depeng, L.; Dejie, L.; Wen, Z.; Yongguo, C.; Naisheng, Z.; Xichen, Z.; et al. Stevioside suppressed inflam-matory cytokine secretion by downregulation of NF-κB and MAPK signaling pathways in LPS-stimulated RAW264.7 cells. Inflammation 2012, 35, 1669–1675. [Google Scholar] [CrossRef]
- Tuntipopipat, S.; Muangnoi, C.; Chingsuwanrote, P.; Parengam, M.; Chantravisut, P.; Charoenkiatkul, S.; Svasti, S. Anti-inflammatory activities of red curry paste extract on lipo-polysaccharide-activated murine macrophage cell line. Nutrition 2011, 27, 479–487. [Google Scholar] [CrossRef]
- Li, W.; Li, W.; Zang, L.; Liu, F.; Yao, Q.; Zhao, J.; Zhi, W.; Niu, X. Fraxin ameliorates lipopolysaccharide-induced acute lung injury in mice by inhibiting the NF-κB and NLRP3 signalling pathways. Int. Immunopharmacol. 2019, 67, 1–12. [Google Scholar] [CrossRef]
- Jiang, K.; Guo, S.; Zhang, T.; Yang, Y.; Zhao, G.; Shaukat, A.; Wu, H.; Deng, G. Downregulation of TLR4 by miR-181a provides negative feedback regulation to lipopolysaccharide-induced inflammation. Front. Pharmacol. 2018, 9, 142. [Google Scholar] [CrossRef] [PubMed]
- Voss, O.H.; Murakami, Y.; Pena, M.Y.; Lee, H.N.; Tina, L.; Margulies, D.H.; Street, J.M.; Yuen, P.S.; Qi, C.F.; Krzewski, K.; et al. Lipopolysaccharide-induced CD300b receptor binding to Toll-like receptor 4 alters signaling to drive cytokine responses that enhance septic shock. Immunity 2016, 44, 1365–1378. [Google Scholar] [CrossRef] [PubMed]
- Hu, X.; Fu, Y.; Lu, X.; Zhang, Z.; Zhang, W.; Cao, Y.; Zhang, N. Protective effects of platycodin D on lipopolysaccharide-induced acute lung injury by activating LXRα-ABCA1 signaling pathway. Front. Immunol. 2017, 7, 644. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.M.; Johnson, J.A. An important role of Nrf2-ARE pathway in the cellular defense mechanism. J. Biochem. Mol. Biol. 2004, 37, 139–143. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.L.; Dodd, G.; Thomas, S.; Zhang, X.; Wasserman, M.A.; Rovin, B.H.; Kunsch, C. Activation of Nrf2/ARE pathway protects endothelial cells from oxidant injury and inhibits inflammatory gene expression. Am. J. Physiol. Heart Circ. Physiol. 2006, 290, H1862–H1870. [Google Scholar] [CrossRef] [PubMed]
- Ryter, S.W.; Alam, J.; Choi, A.M. Heme oxygenase-1/carbon monoxide: From basic science to therapeutic applications. Physiol. Rev. 2006, 86, 583–650. [Google Scholar] [CrossRef]
- Park, S.Y.; Kim, Y.H.; Kim, E.K.; Ryu, E.Y.; Lee, S.J. Heme oxygenase-1 signals are involved in preferential inhibition of pro-inflammatory cytokine release by surfactin in cells activated with porphyromanas gingivalis lipopolysaccharide. Chem. Biol. Interact. 2010, 188, 437–445. [Google Scholar] [CrossRef]
- Rushworth, S.A.; Macewan, D.J.; O’connell, M.A. Lipopolysaccharide-induced expression of NAD(P)H:quinone oxidoreductase 1 and heme oxygenase-1 protects against excessive inflammatory responses in human monocytes. J. Immunol. 2008, 181, 6730–6737. [Google Scholar] [CrossRef]
- Han, C.W.; Kwun, M.J.; Kim, K.H.; Choi, J.Y.; Oh, S.R.; Ahn, K.S.; Lee, J.H.; Joo, M. Ethanol extract of Alismatis Rhizoma reduces acute lung inflammation by suppressing NF-κB and activating Nrf2. J. Ethnopharmacol. 2013, 146, 402–410. [Google Scholar] [CrossRef]
- Park, S.R.; Kim, K.H.; Kwun, M.J.; Lee, J.Y.; Won, R.; Han, C.W.; Choi, J.Y.; Joo, M. Differential regulation of NF-κB and Nrf2 by Bojungikki-Tang is associated with suppressing lung inflammation. Evid. Based Complement. Alternat. Med. 2018, 2018, 5059469. [Google Scholar] [CrossRef]
- Xinmin, D.; Yunyou, D.; Chaosheng, P.; Huasong, F.; Pingkun, Z.; Jiguang, M.; Zhiigian, X.; Qinzhi, X. Dexamethasone treatment attenuates early seawater instillation-induced acute lung injury in rabbits. Pharmacol. Res. 2006, 53, 372–379. [Google Scholar] [CrossRef] [PubMed]
- Thompson, B.T. Glucocorticoids and acute lung injury. Crit. Care Med. 2003, 31, 253–257. [Google Scholar] [CrossRef] [PubMed]
- Hou, T.; Teng, Y.; Sun, Q.; Yu, Z.Y. A new fungitoxic metabolite from Spiraea alpina Pall. Fitoterapia 2009, 80, 237–240. [Google Scholar] [CrossRef] [PubMed]
- Choudhary, M.I.; Naheed, N.; Abbaskhan, A.; Ali, S.; Atta-ur-Rahman. Hemiterpene glucosides and other constituents from Spiraea canescens. Phytochemistry 2009, 70, 1467–1473. [Google Scholar] [CrossRef] [PubMed]
- Hung, T.M.; Na, M.; Thuong, P.T.; Su, N.D.; Sok, D.; Song, K.S.; Seong, Y.H.; Bae, K. Antioxidant activity of caffeoyl quinic acid derivatives from the roots of Dipsacus asper Wall. J. Ethnopharmacol. 2006, 108, 188–192. [Google Scholar] [CrossRef] [PubMed]
- Abdel Motaal, A.; Ezzat, S.M.; Tadros, M.S.; El-Askary, H.I. In vivo anti-inflammatory activity of caffeoylquinic acid derivatives from Solidago vigaurea in rats. Pharm. Biol. 2016, 54, 2864–2870. [Google Scholar] [CrossRef] [PubMed]
- Huang, R.; Zhong, T.; Wu, H. Quercetin protects against lipopolysaccharide-induced acute lung injury in rats through suppression of inflammation and oxidative stress. Arch. Med. Sci. 2015, 11, 427–432. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No. | tR (min) | Detected m/z [M − H]− | Exacted m/z [M − H]− | Error (ppm) | Formula | Fragments | Identification |
|---|---|---|---|---|---|---|---|
| 1 | 3.51 | 341.0879 | 341.0878 | 0.19 | C15H18O9 | 179 | caffeoyl glucoside |
| 2 | 4.49 | 353.0866 | 353.0878 | −0.81 | C16H18O9 | 191, 179, 135 | caffeoyl quinic acid |
| 3 | 4.69 | 325.0921 | 325.0929 | −0.65 | C15H18O8 | 163, 145, 135 | coumaroyl hexoside |
| 4 | 4.80 | 353.0866 | 353.0878 | −0.61 | C16H18O9 | 191, 179, 135 | peak 2 isomer |
| 5 | 7.44 | 439.1247 | 439.1246 | −0.76 | C20H24O11 | 341, 179, 161 | unknown |
| 6 | 7.69 | 463.0893 | 463.0882 | 0.12 | C21H20O12 | 301, 271, 255 | quercetin glucoside |
| 7 | 8.21 | 757.1626 | 757.1622 | 0.66 | C35H34O19 | 595, 301, 271 | quercetin hexosylpentoside-hexoside |
| 8 | 8.37 | 503.1180 | 503.1195 | −0.26 | C24H24O12 | 341, 179, 161 | dicaffeoyl glucoside |
| 9 | 8.74 | 741.1682 | 741.1672 | −0.17 | C35H34O18 | 579, 455, 285 | kaempferol hexosylpentoside-hexoside |
| 10 | 8.97 | 799.1735 | 799.1727 | −0.10 | C37H36O20 | 637, 595, 301 | quercetin acetylhexoside-pentoside hexoside |
| 11 | 9.14 | 637.1417 | 637.1410 | −0.06 | C28H30O17 | 595, 335, 301, | quercetin acetylhexoside-pentoside |
| 12 | 9.23 | 617.1504 | 617.1512 | 0.43 | C29H30O15 | 455, 395, 179, | unknown |
| 13 | 9.32 | 503.1208 | 503.1195 | −0.44 | C24H24O12 | 341, 179, 161 | peak 8 isomer |
| 14 | 9.41 | 487.1235 | 487.1246 | −0.36 | C24H24O11 | 323, 161 | unknown |
| 15 | 9.71 | 799.1735 | 799.1727 | −1.17 | C37H36O20 | 637, 301 | peak 10 isomer |
| 16 | 9.97 | 307.0825 | 307.0823 | 0.60 | C15H16O7 | 285, 161 | unknown |
| 17 | 10.11 | 799.1735 | 799.1727 | 1.18 | C37H36O20 | 637, 301 | peak 10 isomer |
| 18 | 10.46 | 601.1573 | 601.1563 | 0.23 | C29H30O14 | 439, 179, 135 | unknown |
| 19 | 10.71 | 783.1786 | 783.1778 | 0.01 | C37H36O19 | 621, 285 | kaempferol acetylhexoside-pentoside hexoside |
| 20 | 11.09 | 813.1893 | 813.1884 | 0.43 | C38H38O20 | 637, 595, 301 | quercetin acetylhexoside-pentoside glucuronide |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lee, B.-W.; Ha, J.-H.; Shin, H.-G.; Jeong, S.-H.; Jeon, D.-B.; Kim, J.-H.; Park, J.-Y.; Kwon, H.-J.; Jung, K.; Lee, W.-S.; et al. Spiraea prunifolia var. simpliciflora Attenuates Oxidative Stress and Inflammatory Responses in a Murine Model of Lipopolysaccharide-Induced Acute Lung Injury and TNF-α-Stimulated NCI-H292 Cells. Antioxidants 2020, 9, 198. https://doi.org/10.3390/antiox9030198
Lee B-W, Ha J-H, Shin H-G, Jeong S-H, Jeon D-B, Kim J-H, Park J-Y, Kwon H-J, Jung K, Lee W-S, et al. Spiraea prunifolia var. simpliciflora Attenuates Oxidative Stress and Inflammatory Responses in a Murine Model of Lipopolysaccharide-Induced Acute Lung Injury and TNF-α-Stimulated NCI-H292 Cells. Antioxidants. 2020; 9(3):198. https://doi.org/10.3390/antiox9030198
Chicago/Turabian StyleLee, Ba-Wool, Ji-Hye Ha, Han-Gyo Shin, Seong-Hun Jeong, Da-Bin Jeon, Ju-Hong Kim, Ji-Young Park, Hyung-Jun Kwon, Kyungsook Jung, Woo-Song Lee, and et al. 2020. "Spiraea prunifolia var. simpliciflora Attenuates Oxidative Stress and Inflammatory Responses in a Murine Model of Lipopolysaccharide-Induced Acute Lung Injury and TNF-α-Stimulated NCI-H292 Cells" Antioxidants 9, no. 3: 198. https://doi.org/10.3390/antiox9030198
APA StyleLee, B.-W., Ha, J.-H., Shin, H.-G., Jeong, S.-H., Jeon, D.-B., Kim, J.-H., Park, J.-Y., Kwon, H.-J., Jung, K., Lee, W.-S., Kim, H.-Y., Kim, S.-H., Jang, H.-J., Ryu, Y.-B., & Lee, I.-C. (2020). Spiraea prunifolia var. simpliciflora Attenuates Oxidative Stress and Inflammatory Responses in a Murine Model of Lipopolysaccharide-Induced Acute Lung Injury and TNF-α-Stimulated NCI-H292 Cells. Antioxidants, 9(3), 198. https://doi.org/10.3390/antiox9030198