Scrophularia koraiensis Nakai Attenuates Allergic Airway Inflammation via Suppression of NF-κB and Enhancement of Nrf2/HO-1 Signaling

 ,
,  ,
,  and
and 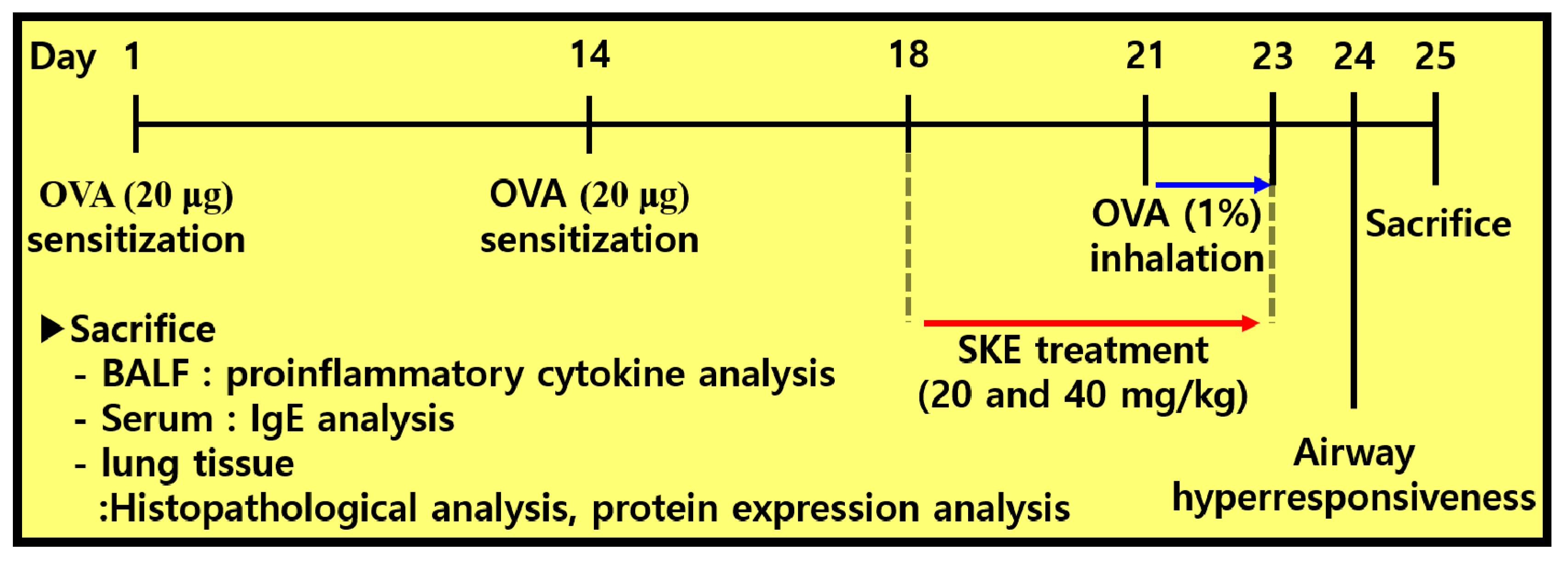{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Materials and Instruments
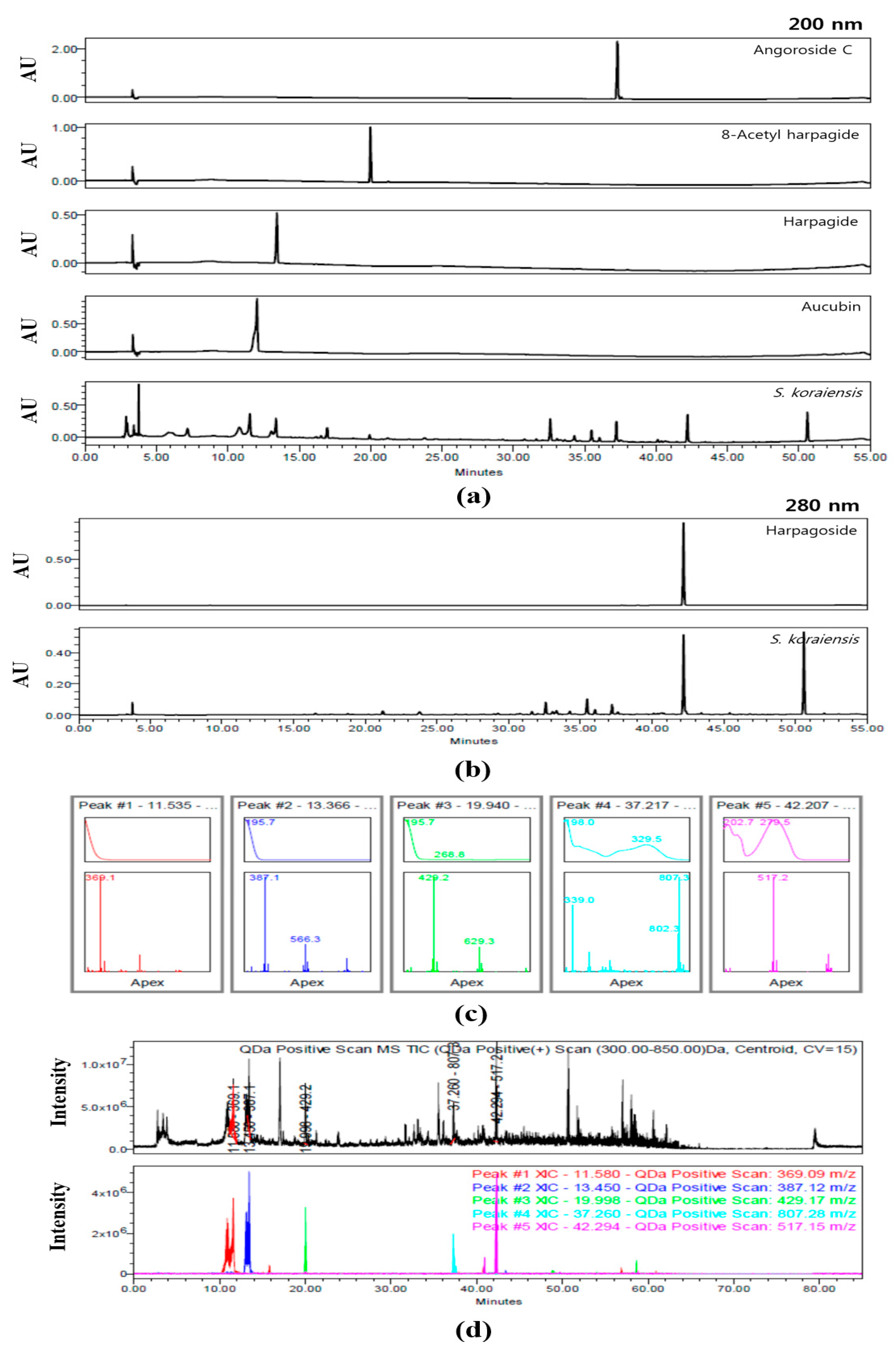
2.3. HPLC analysis
2.4. OVA-Induced Allergic Airway Inflammation Model
2.5. Measurement of Allergic Parameters in Bronchoalveolar Lavage Fluid (BALF) and Serum
2.6. Histopathology of Lung Tissue
2.7. Western Blotting
2.8. Statistical Analysis
3. Results
3.1. Identification of Compounds within SKE by HPLC Analysis
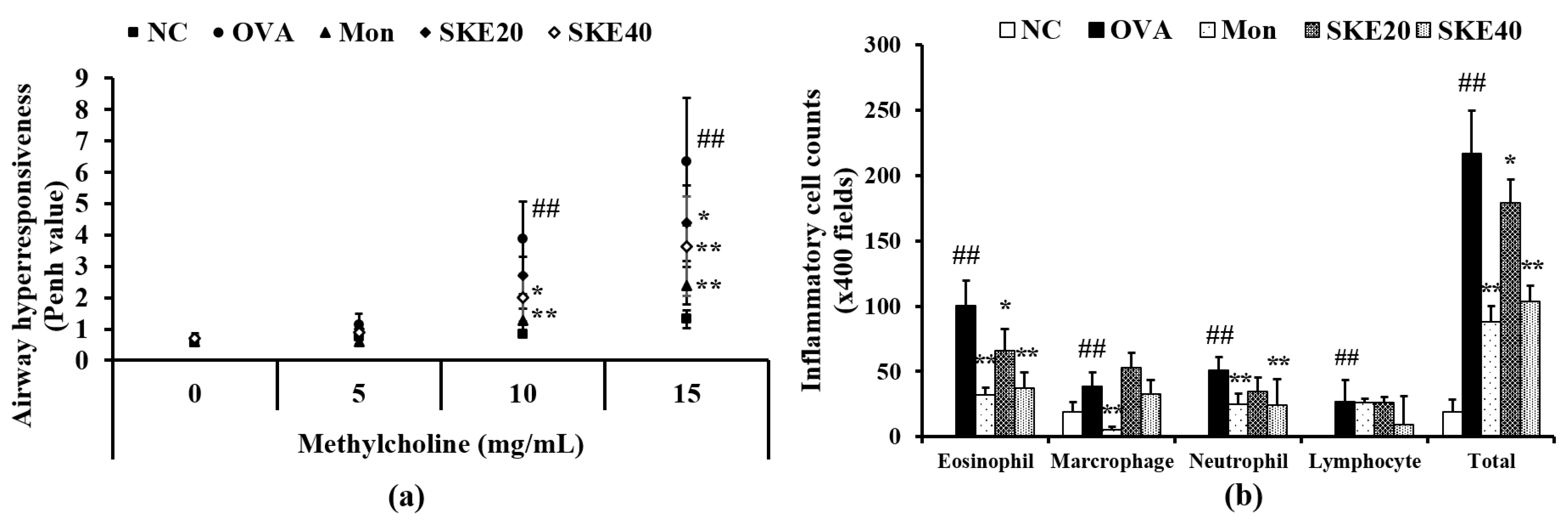
3.2. Effect of SKE on AHR and Inflammatory Cell Count During OVA-Induced Allergic Airway Inflammation
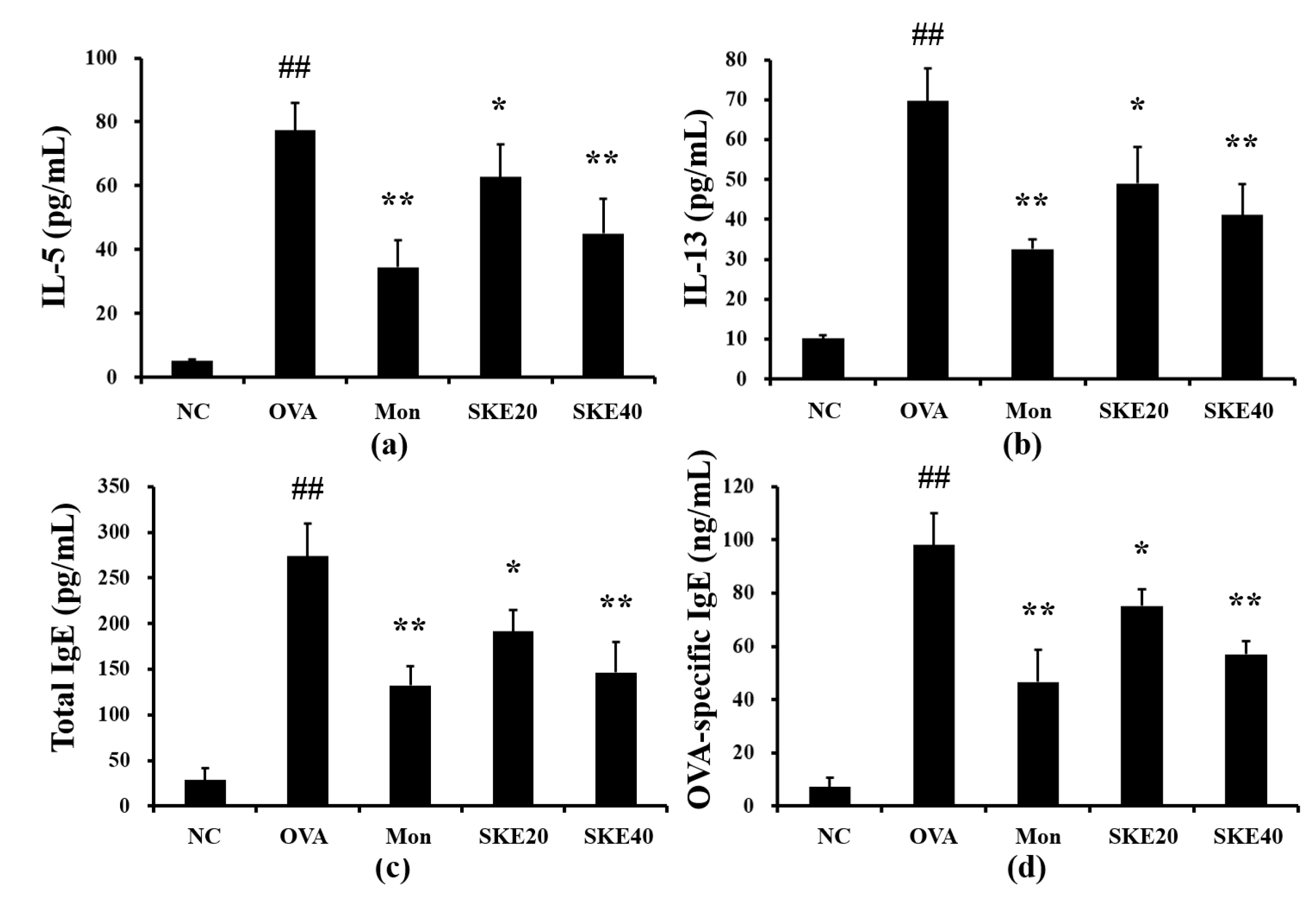
3.3. Effect of SKE on Pro-Inflammatory Cytokines and IgE Levels During OVA-Induced Allergic Airway Inflammation
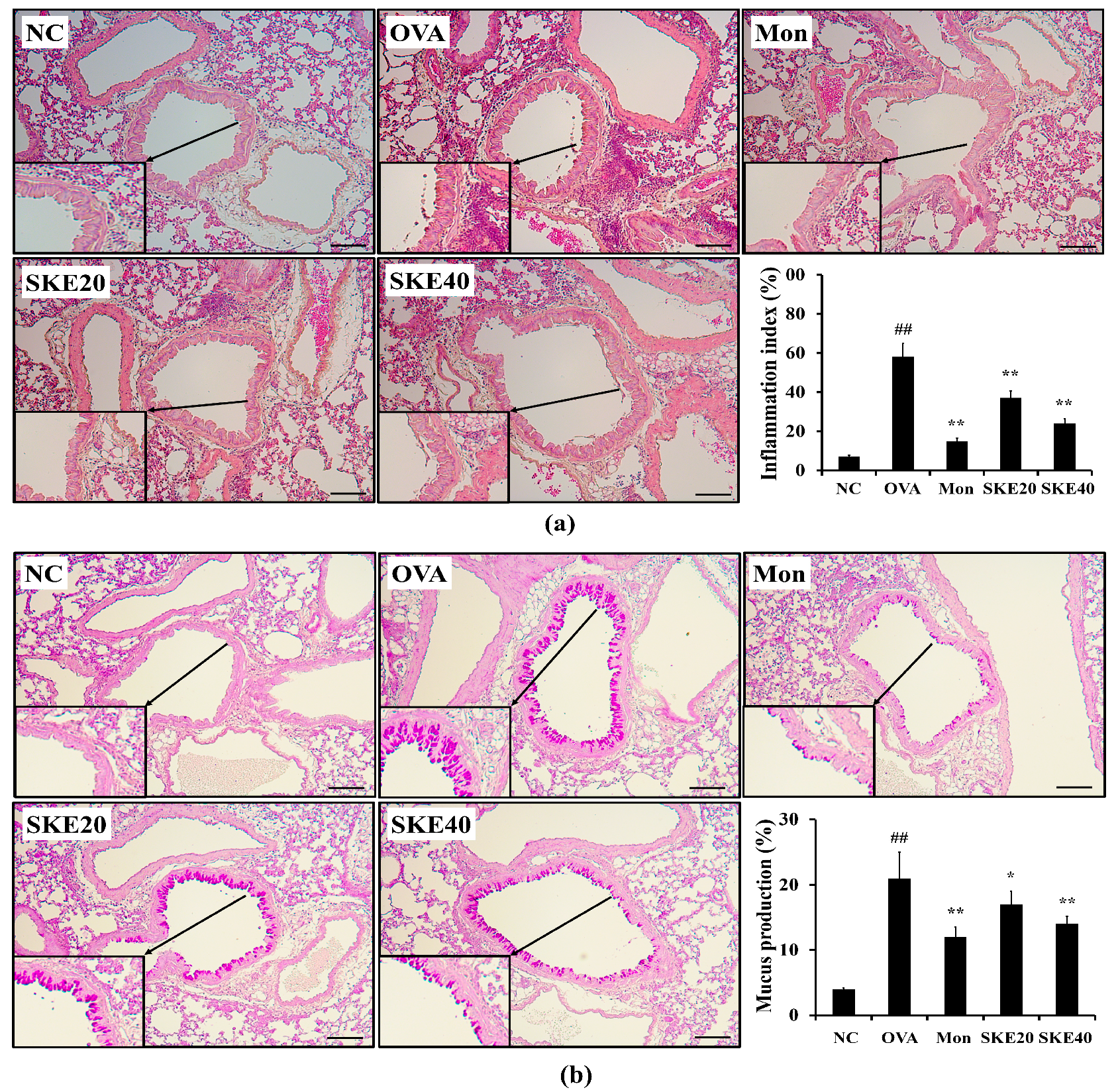
3.4. Effect of SKE on Airway Inflammation and Mucus Production During OVA-Induced Allergic Airway Inflammation
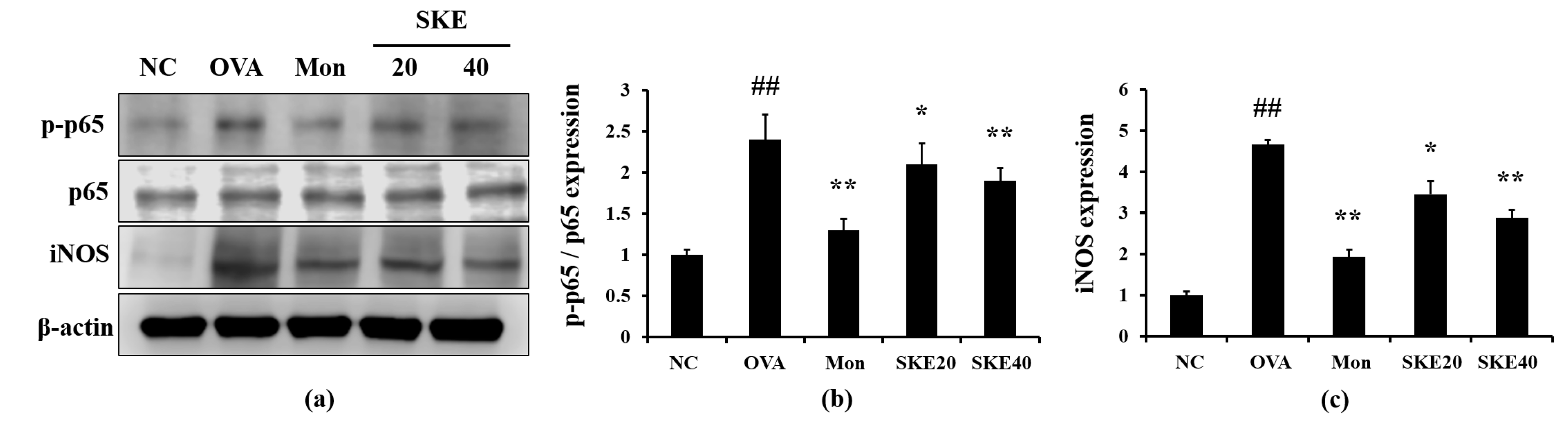
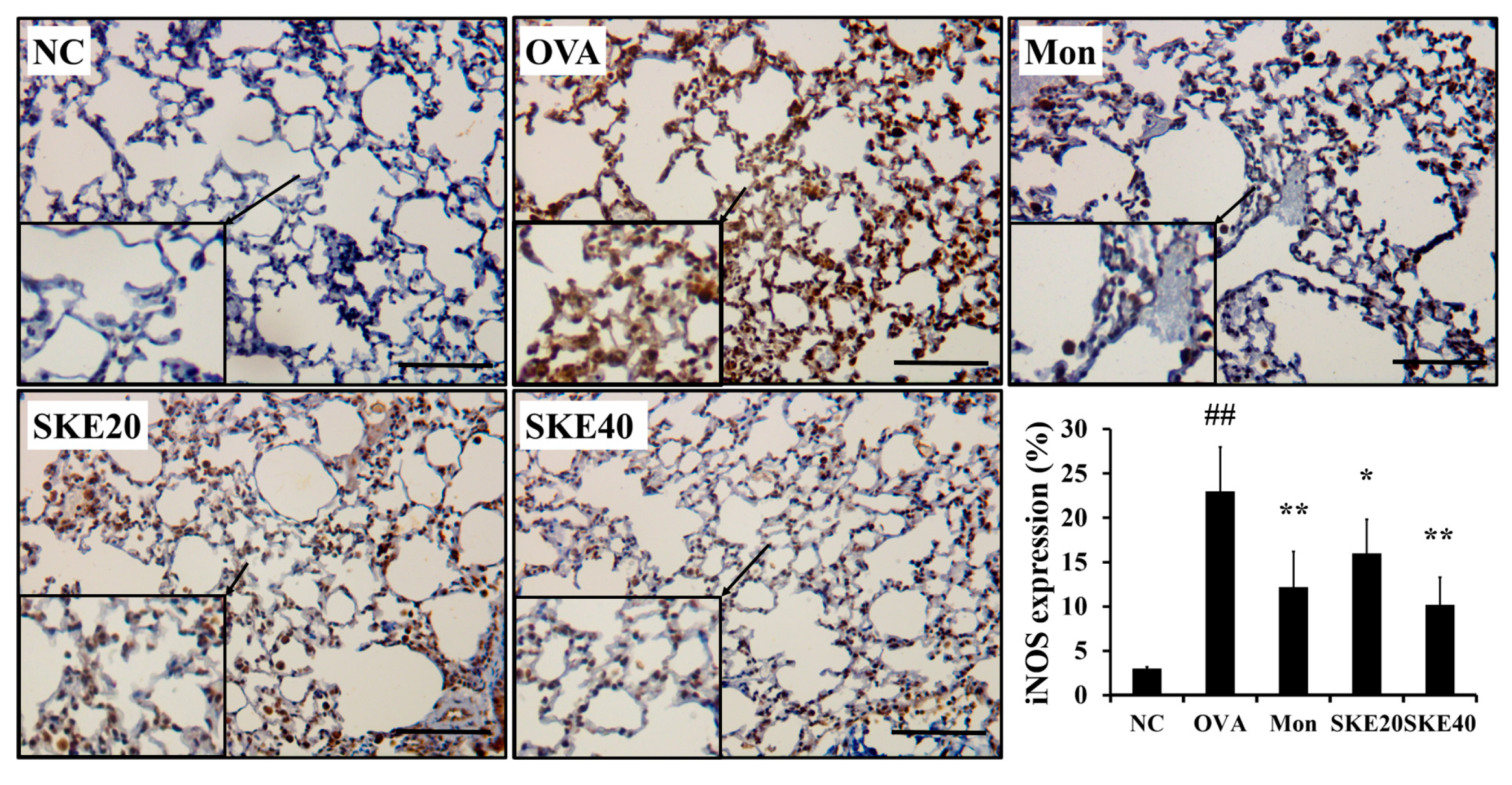
3.5. Effect of SKE on NF-κB Phosphorylation and iNOS Expression During OVA-Induced Allergic Airway Inflammation
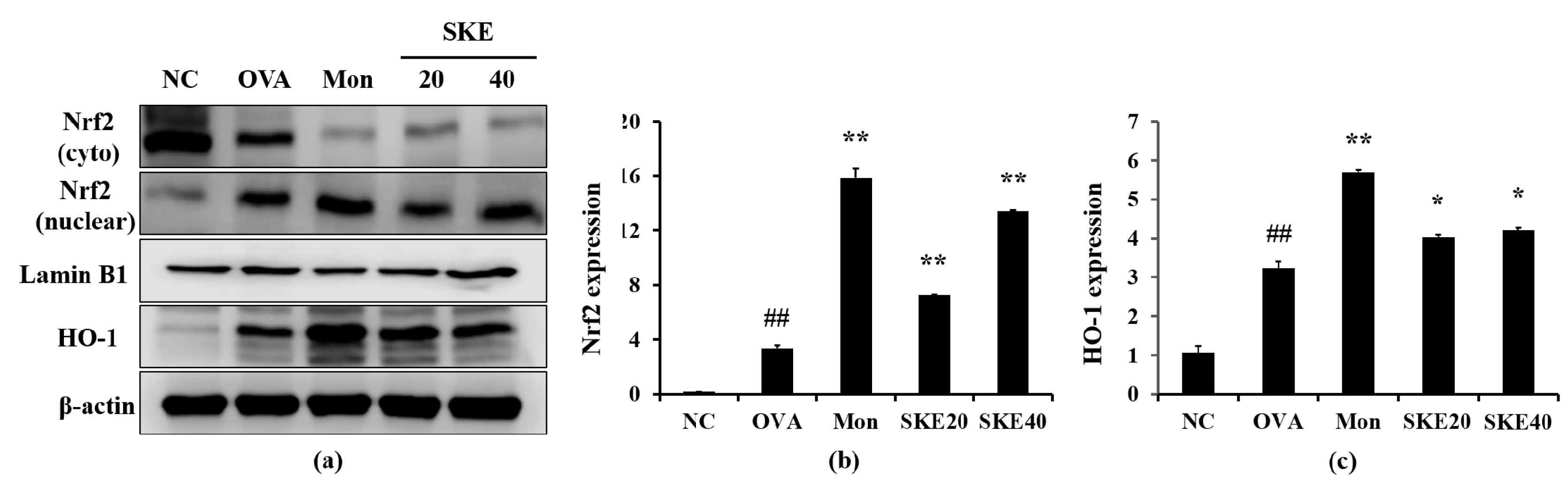
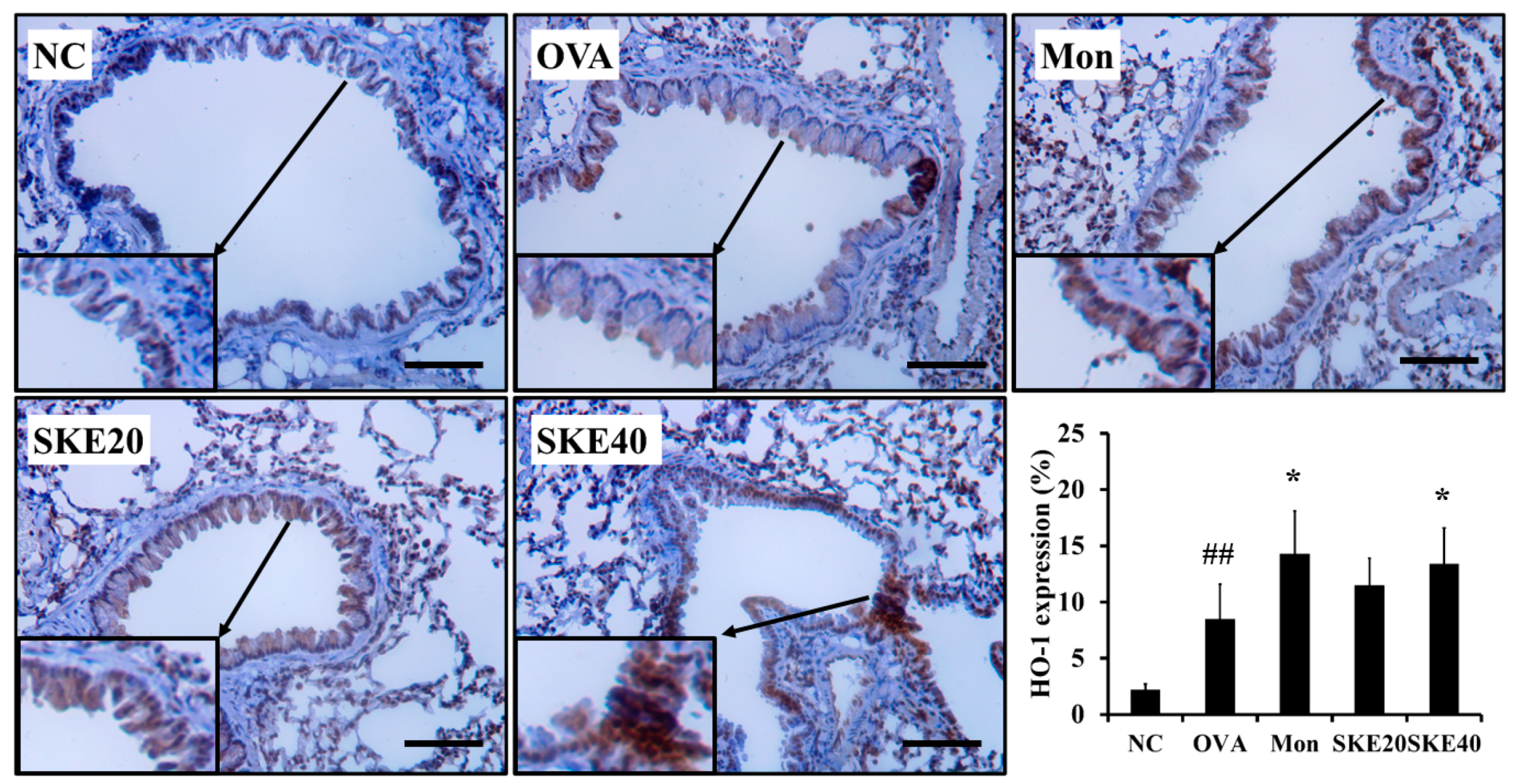
3.6. Effect of SKE on Nrf2/HO-1 Signaling During OVA-Induced Allergic Airway Inflammation
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Huang, C.C.; Chang, P.H.; Wu, P.W.; Wang, C.H.; Fu, C.H.; Huang, C.C.; Tseng, H.J.; Lee, T.J. Impact of nasal symptoms on the evaluation of asthma control. Medicine 2017, 96, e6147. [Google Scholar] [CrossRef] [PubMed]
- Shin, N.R.; Ryu, H.W.; Ko, J.W.; Park, S.H.; Yuk, H.J.; Kim, H.J.; Kim, J.C.; Jeong, S.H.; Shin, I.S. Artemisia argyi attenuates airway inflammation in ovalbumin-induced asthmatic animals. J. Ethnopharmacol. 2017, 209, 108–115. [Google Scholar] [CrossRef] [PubMed]
- Sahiner, U.M.; Birben, E.; Erzurum, S.; Sackesen, C.; Kalayci, Ö. Oxidative stress in asthma: Part of the puzzle. Pediatr. Allergy Immunol. 2018, 29, 789–800. [Google Scholar] [CrossRef] [PubMed]
- Sanchez-Solis, M. Early lung function and future asthma. Front. Pediatr. 2019, 7, 253. [Google Scholar] [CrossRef]
- Gauthier, M.; Ray, A.; Wenzel, S.E. Evolving concepts of asthma. Am. J. Respir. Crit. Care Med. 2015, 192, 660–668. [Google Scholar] [CrossRef]
- Akiki, Z.; Rava, M.; Diaz Gil, O.; Pin, I.; le Moual, N.; Siroux, V.; Guerra, S.; Chamat, S.; Matran, R.; Fitó, M.; et al. Serum cytokine profiles as predictors of asthma control in adults from the EGEA study. Respir. Med. 2017, 125, 57–64. [Google Scholar] [CrossRef]
- Fahy, J.V. Type 2 inflammation in asthma—Present in most, absent in many. Nat. Rev. Immunol. 2015, 15, 57–65. [Google Scholar] [CrossRef]
- Shergis, J.L.; Wu, L.; Zhang, A.L.; Guo, X.; Lu, C.; Xue, C.C. Herbal medicine for adults with asthma: A systematic review. J. Asthma 2016, 53, 650–659. [Google Scholar] [CrossRef]
- Hu, L.; Li, L.; Zhang, H.; Li, Q.; Jiang, S.; Qiu, J.; Sun, J.; Dong, J. Inhibition of airway remodeling and inflammatory response by Icariin in asthma. BMC Complement. Altern. Med. 2019, 19, 316. [Google Scholar] [CrossRef]
- Lee, J.W.; Min, J.H.; Kim, M.G.; Kim, S.M.; Kwon, O.K.; Oh, T.K.; Lee, J.K.; Kim, T.Y.; Lee, S.W.; Choi, S.; et al. Pistacia weinmannifolia root exerts a protective role in ovalbumin-induced lung inflammation in a mouse allergic asthma model. Int. J. Mol. Med. 2019, 44, 2171–2180. [Google Scholar] [CrossRef]
- Thannickal, V.J.; Fanburg, B.L. Reactive oxygen species in cell signaling. Am. J. Physiol. Lung Cell. Mol. Physiol. 2000, 279, L1005–L1028. [Google Scholar] [CrossRef] [PubMed]
- Bullone, M.; Lavoie, J.P. The Contribution of Oxidative Stress and Inflamm-Aging in Human and Equine Asthma. Int. J. Mol. Sci. 2017, 18, 2612. [Google Scholar] [CrossRef] [PubMed]
- Belenky, P.; Collins, J.J. Antioxidant strategies to tolerate antibiotics. Science 2011, 334, 915–916. [Google Scholar] [CrossRef]
- Rahman, I.; Biswas, S.K.; Kode, A. Oxidant and antioxidant balance in the airways and airway diseases. Eur. J. Pharmacol. 2006, 533, 222–239. [Google Scholar] [CrossRef] [PubMed]
- Ryter, S.W.; Otterbein, L.E.; Morse, D.; Choi, A.M. Heme oxygenase/carbon monoxide signaling pathways: Regulation and functional significance. Mol. Cell. Biochem. 2002, 234–235, 249–263. [Google Scholar] [CrossRef]
- Zhang, Z.H.; Zhu, W.; Ren, H.Z.; Zhao, X.; Wang, S.; Ma, H.C.; Shi, X.L. Mesenchymal stem cells increase expression of heme oxygenase-1 leading to anti-inflammatory activity in treatment of acute liver failure. Stem Cell Res. Ther. 2017, 8, 70. [Google Scholar] [CrossRef]
- Xiao, S.; Tolga, E.; Henry, J.G.; Thomas, E. Effects of Scrophularia ningpoensis Hemsl. on inhibition of proliferation, apoptosis induction and NF-κB signaling of immortalized and cancer cell lines. Pharmaceutical 2012, 5, 189–208. [Google Scholar] [CrossRef]
- Mitsuo, M.; Yoshiharu, O.; Sei-ichi, N.; Hiromu, K. Suppression of SOS-inducing activity of chemical mutagens by cinnamic acid derivatives from Scrophulia ningpoensis in the Salmonella typhimurium TA1535/pSK1002 umu test. J. Agric. Food Chem. 1998, 46, 904–910. [Google Scholar] [CrossRef]
- Han, K.S.; So, S.K.; Lee, C.H.; Kim, M.Y. Taxonomy of the genus Scrophularia (Scrophulariaceae) in Korea. Korean J. Plant Taxon. 2009, 39, 237–246. [Google Scholar] [CrossRef]
- Ko, J.W.; Shin, N.R.; Park, S.H.; Cho, Y.K.; Kim, J.C.; Seo, C.S.; Shin, I.S. Genipin inhibits allergic responses in ovalbumin-induced asthmatic mice. Int. Immunopharmacol. 2017, 53, 49–55. [Google Scholar] [CrossRef]
- Tran, T.N.; Zeiger, R.S.; Peters, S.P.; Colice, G.; Newbold, P.; Goldman, M.; Chipps, B.E. Overlap of atopic, eosinophilic, and Th2-high asthma phenotypes in a general population with current asthma. Ann. Allergy Asthma Immunol. 2016, 116, 37–42. [Google Scholar] [CrossRef]
- Shrestha Palikhe, N.; Bosonea, A.M.; Laratta, C.; Gandhi, V.D.; Nahirney, D.; Hillaby, A.; Bowen, M.; Bhutani, M.; Mayers, I.; Cameron, L.; et al. Stability of peripheral blood immune markers in patients with asthma. Allergy Asthma Clin. Immunol. 2019, 15, 30. [Google Scholar] [CrossRef]
- Narendra, D.K.; Hanania, N.A. Targeting IL-5 in COPD. Int. J. Chronic Obstr. Pulm. Dis. 2019, 14, 1045–1051. [Google Scholar] [CrossRef] [PubMed]
- Robinson, D.S.; Hamid, Q.; Ying, S.; Tsicopoulos, A.; Barkans, J.; Bentley, A.M.; Corrigan, C.; Durham, S.R.; Kay, A.B. Predominant Th2-like bronchoalveolar T-lymphocyte population in atopic asthma. N. Engl. J. Med. 1992, 326, 298–304. [Google Scholar] [CrossRef] [PubMed]
- Montero-Pérez, O.; Contreras-Rey, M.B.; Sánchez-Gómez, E. Effectiveness and safety of mepolizumab in severe refractory eosinophilic asthma; results in clinical practice. Drugs Context 2019, 8, 212584. [Google Scholar] [CrossRef] [PubMed]
- Chung, K.F.; Adcock, I.M. Multifaceted mechanisms in COPD: Inflammation, immunity, and tissue repair and destruction. Eur. Respir. J. 2008, 31, 1334–1356. [Google Scholar] [CrossRef] [PubMed]
- Kurakula, K.; Hamers, A.A.; van Loenen, P.; de Vries, C.J. 6-Mercaptopurine reduces cytokine and MUC5AC expression involving inhibition of NF-κB activation in airway epithelial cells. Respir. Res. 2015, 16, 73. [Google Scholar] [CrossRef]
- Ghosh, S.; Erzurum, S.C. Nitric oxide metabolism in asthma pathophysiology. Biochim. Biophys. Acta 2011, 1810, 1008–1016. [Google Scholar] [CrossRef]
- Lowry, J.L.; Brovkovych, V.; Zhang, Y.; Skidgel, R.A. Endothelial nitric-oxide synthase activation generates an inducible nitric-oxide synthase-like output of nitric oxide in inflamed enthothelium. J. Biol. Chem. 2013, 288, 4174–4193. [Google Scholar] [CrossRef]
- de Andres, M.C.; Takahashi, A.; Oreffo, R.O. Demethylation of an NF-κB enhancer element orchestrates iNOS induction in osteoarthritis and is associated with altered chondrocyte cell cycle. Osteoarthr. Cartil. 2016, 24, 1951–1960. [Google Scholar] [CrossRef]
- Blaise, G.A.; Gauvin, D.; Gangal, M.; Authier, S. Nitric oxide, cell signaling and cell death. Toxicology 2005, 208, 177–192. [Google Scholar] [CrossRef] [PubMed]
- Lim, J.C.; Goh, F.Y.; Sagineedu, S.R.; Yong, A.C.; Sidik, S.M.; Lajis, N.H.; Wong, W.S.; Stanslas, J. A semisynthetic diterpenoid lactone inhibits NF-κB signaling to ameliorate inflammation and airway hyperresponsiveness in a mouse asthma model. Toxicol. Appl. Pharmacol. 2016, 302, 10–22. [Google Scholar] [CrossRef] [PubMed]
- Shin, I.S.; Hong, J.M.; Jeon, C.M.; Shin, N.R.; Kwon, O.K.; Kim, H.S.; Kim, J.C.; Oh, S.R.; Ahn, K.S. Diallyl-disulfide, an organosulfur compound of garlic, attenuates airway inflammation via activation of the Nrf-2/HO-1 pathway and NF-kappaB suprresion. Food Chem. Toxicol. 2013, 62, 506–513. [Google Scholar] [CrossRef] [PubMed]
- Baird, L.; Dinkova-Kostova, A.T. The cytoprotective role of the Keap1-Nrf2 pathway. Arch. Toxicol. 2011, 85, 241–272. [Google Scholar] [CrossRef]
- Cheng, H.T.; Yen, C.J.; Chang, C.C.; Huang, K.T.; Chen, K.H.; Zhang, R.Y.; Lee, P.Y.; Miaw, S.C.; Huang, J.W.; Chiang, C.K.; et al. Ferritin heavy chain mediates the protective effect of heme oxygenase-1 against oxidative stress. Biochim. Biophys. Acta 2015, 1850, 2506–2517. [Google Scholar] [CrossRef]
- Han, C.W.; Kwun, M.J.; Kim, K.H.; Choi, J.Y.; Oh, S.R.; Ahn, K.S.; Lee, J.H.; Joo, M. Ethanol extract of Alismatis Rhizoma reduces acute lung inflammation by suppressing NF-κB and activating Nrf2. J. Ehtnopharmacol. 2013, 146, 402–410. [Google Scholar] [CrossRef]
- Nikkhah, E.; Heshmati Afshar, F.; Babaei, H.; Asgharian, P.; Delazar, A. Phytochemical analysis and in-vitro bioactivity of Scrophularia umbrosa Rhizome (Scrophulariaceae). Iran. J. Pharm. Res. 2018, 17, 685–694. [Google Scholar]
- Jeong, J.; Wahyudi, L.D.; Keum, Y.S.; Yang, H.; Kim, J.H. E-p-methoxycinnamoyl-α-l-rhamnopyranosyl ester, a phenylpropanoid isolated from Scrophularia buergeriana, increases nuclear factor erythroid-derived 2-related factor 2 stability by inhibiting ubiquitination in human keratinocytes. Molecules 2018, 23, 768. [Google Scholar] [CrossRef]
- Ding, B.; Lu, Y.; Li, Y.; Zhou, W.; Qin, F. Efficacy of treatment with montelukast, fluticasone propionate and budesonide liquid suspension for the prevention of recurrent asthma paroxysms in children with wheezing disorders. Exp. Ther. Med. 2019, 18, 3090–3094. [Google Scholar] [CrossRef]









© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jung, T.-Y.; Lee, A.Y.; Song, J.-H.; Lee, M.Y.; Lim, J.-O.; Lee, S.-J.; Ko, J.-W.; Shin, N.-R.; Kim, J.-C.; Shin, I.-S.; et al. Scrophularia koraiensis Nakai Attenuates Allergic Airway Inflammation via Suppression of NF-κB and Enhancement of Nrf2/HO-1 Signaling. Antioxidants 2020, 9, 99. https://doi.org/10.3390/antiox9020099
Jung T-Y, Lee AY, Song J-H, Lee MY, Lim J-O, Lee S-J, Ko J-W, Shin N-R, Kim J-C, Shin I-S, et al. Scrophularia koraiensis Nakai Attenuates Allergic Airway Inflammation via Suppression of NF-κB and Enhancement of Nrf2/HO-1 Signaling. Antioxidants. 2020; 9(2):99. https://doi.org/10.3390/antiox9020099
Chicago/Turabian StyleJung, Tae-Yang, A Yeong Lee, Jun-Ho Song, Min Young Lee, Je-Oh Lim, Se-Jin Lee, Je-Won Ko, Na-Rae Shin, Jong-Choon Kim, In-Sik Shin, and et al. 2020. "Scrophularia koraiensis Nakai Attenuates Allergic Airway Inflammation via Suppression of NF-κB and Enhancement of Nrf2/HO-1 Signaling" Antioxidants 9, no. 2: 99. https://doi.org/10.3390/antiox9020099
APA StyleJung, T.-Y., Lee, A. Y., Song, J.-H., Lee, M. Y., Lim, J.-O., Lee, S.-J., Ko, J.-W., Shin, N.-R., Kim, J.-C., Shin, I.-S., & Kim, J.-S. (2020). Scrophularia koraiensis Nakai Attenuates Allergic Airway Inflammation via Suppression of NF-κB and Enhancement of Nrf2/HO-1 Signaling. Antioxidants, 9(2), 99. https://doi.org/10.3390/antiox9020099