
Structural Identification and Molecular Interaction Modeling Analysis of Antioxidant Activity Selenium-Enriched Peptides from Selenium-Enriched Pleurotus eryngii

 , ,
, ,
Abstract
1. Introduction
2. Materials and Methods
2.1. Materials and Reagents
2.2. Preparation of SePEPs
2.3. Determination of Antioxidant Activity of SePEPs and PEPs In Vitro
2.4. Transmission Electron Microscopy (TEM)
2.5. UV Absorption Spectra
2.6. Fluorescence Spectra
2.7. Fourier Transform Infrared (FTIR) Spectra
2.8. Analysis of Amino Acid Composition
2.9. Liquid Chromatography–Mass Spectrometry/Mass Spectrometry (LC-MS/MS)
2.9.1. Sample Preparation
2.9.2. Liquid Chromatographic Conditions
2.9.3. Mass Spectrometry Conditions
2.10. Molecular Interaction Modeling Analysis
2.11. Peptide Synthesis
2.12. DPPH Radical Scavenging Rates of Selenium-Containing Peptide
2.13. Data Statistics and Analysis
3. Results
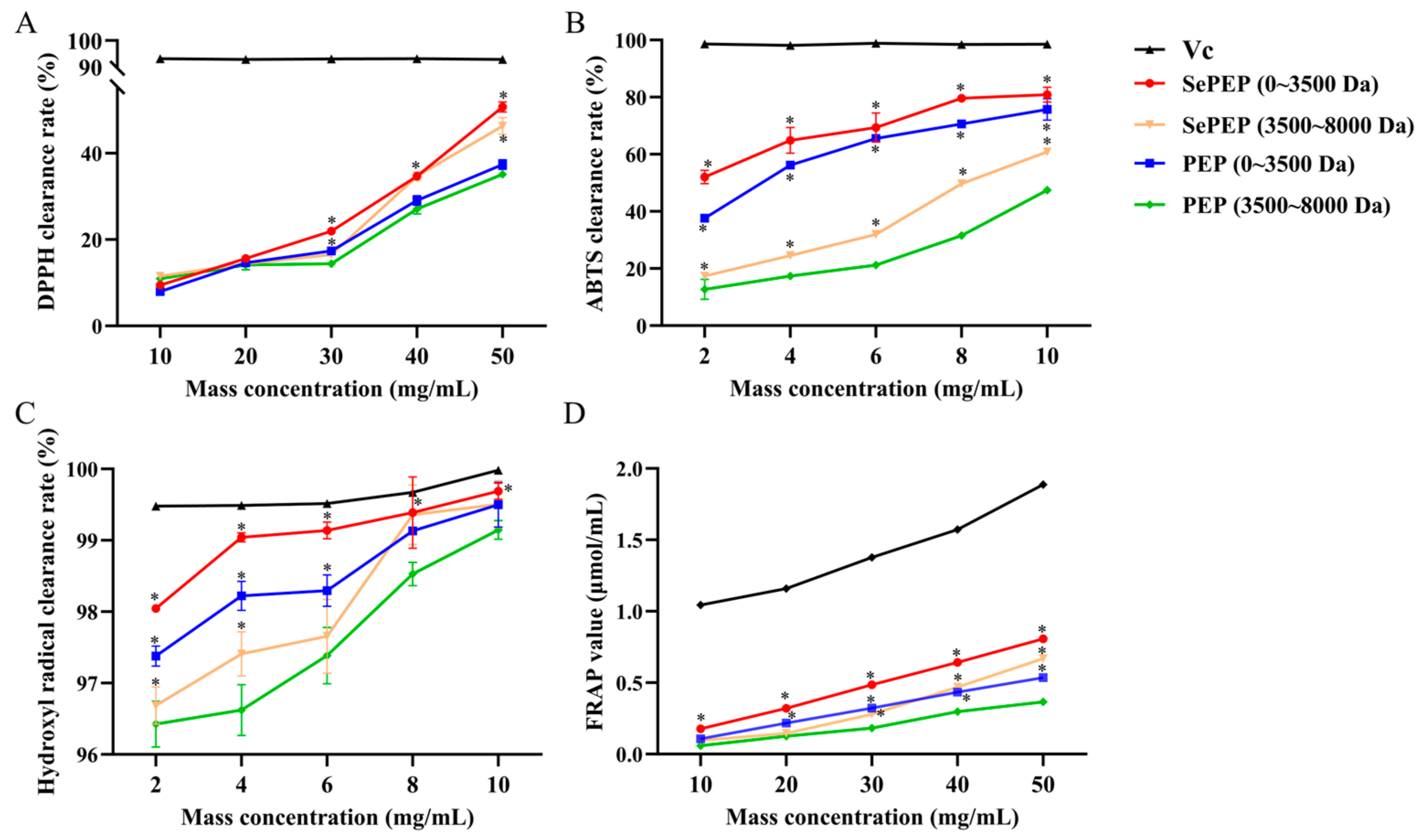
3.1. Comparison of In Vitro Antioxidant Activities of SePEPs and PEPs
3.2. TEM Analysis of SePEPs and PEPs
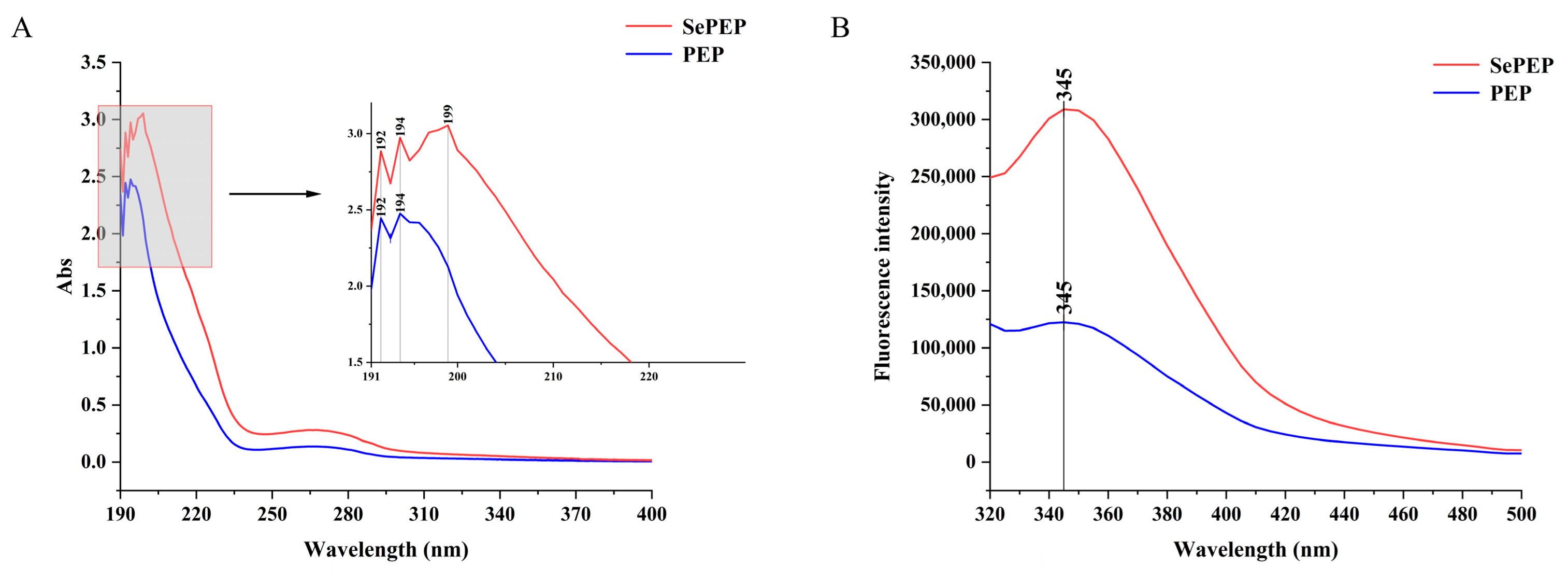
3.3. UV Absorption and Fluorescence Spectra
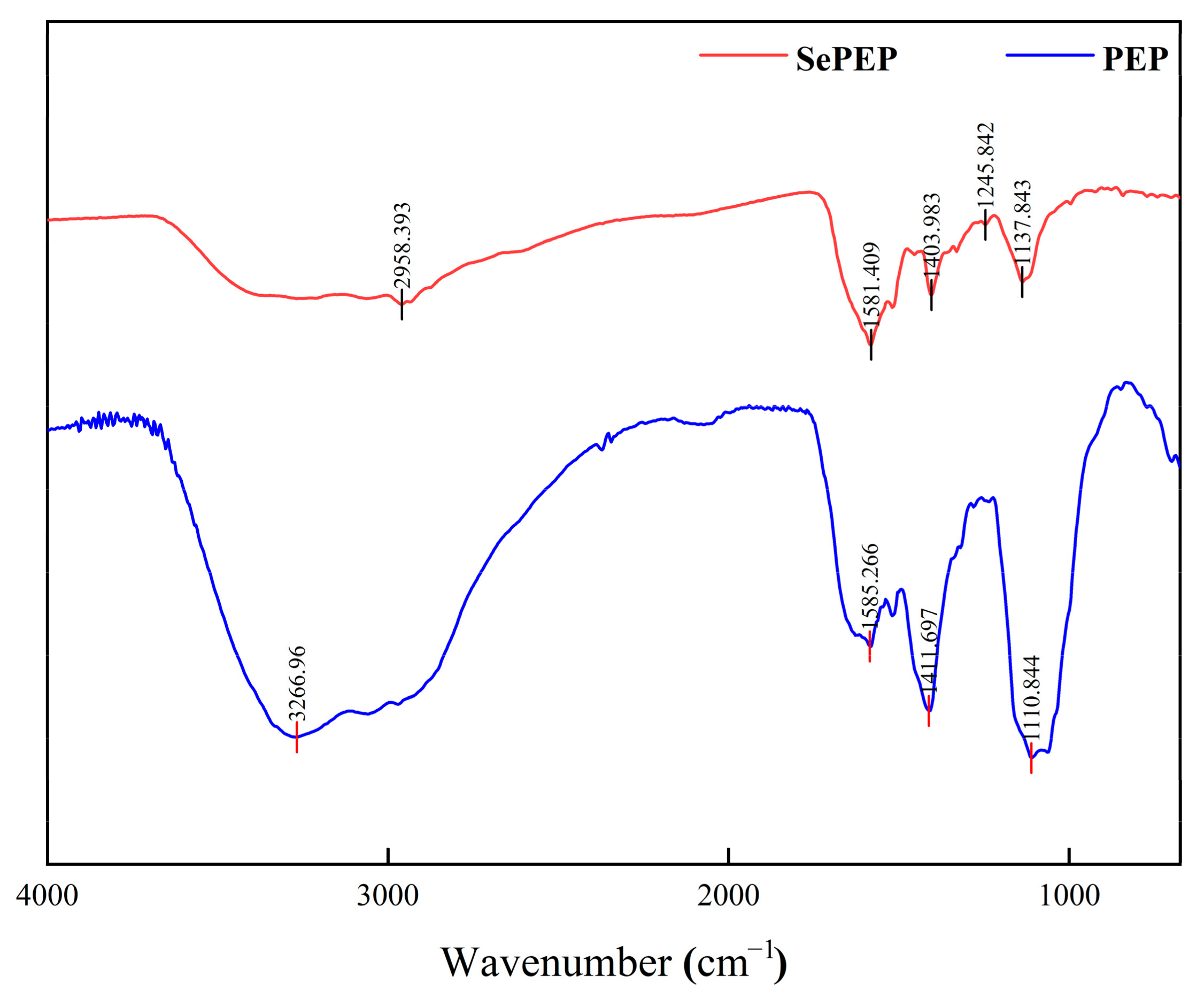
3.4. FTIR
3.5. Amino Acid Composition of SePEPs and PEPs
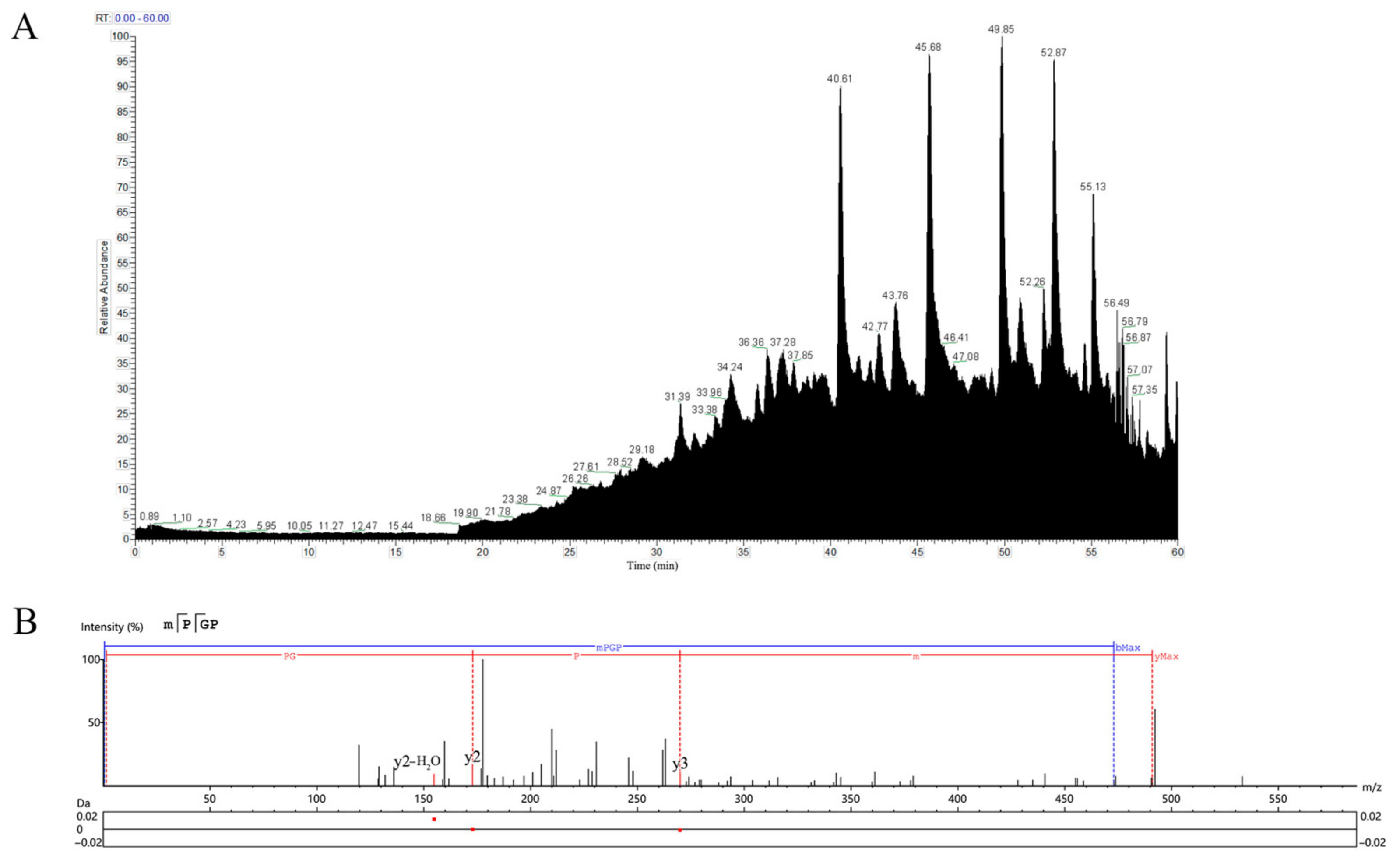
3.6. LC-MS/MS
3.7. Molecular Virtual Screening of Selenium-Containing Peptides with DPPH
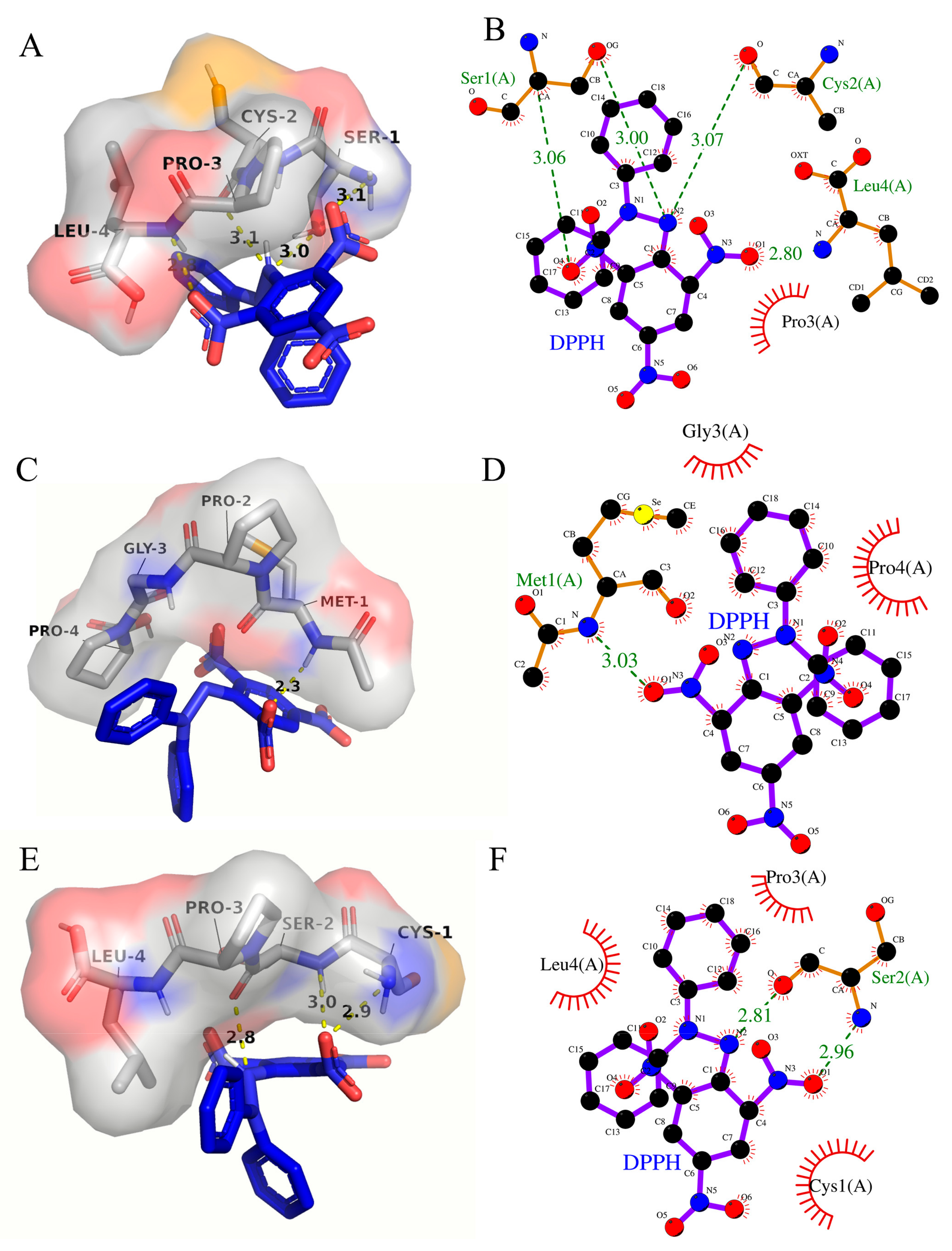
3.8. Molecular Interaction Modeling Visualization Analysis
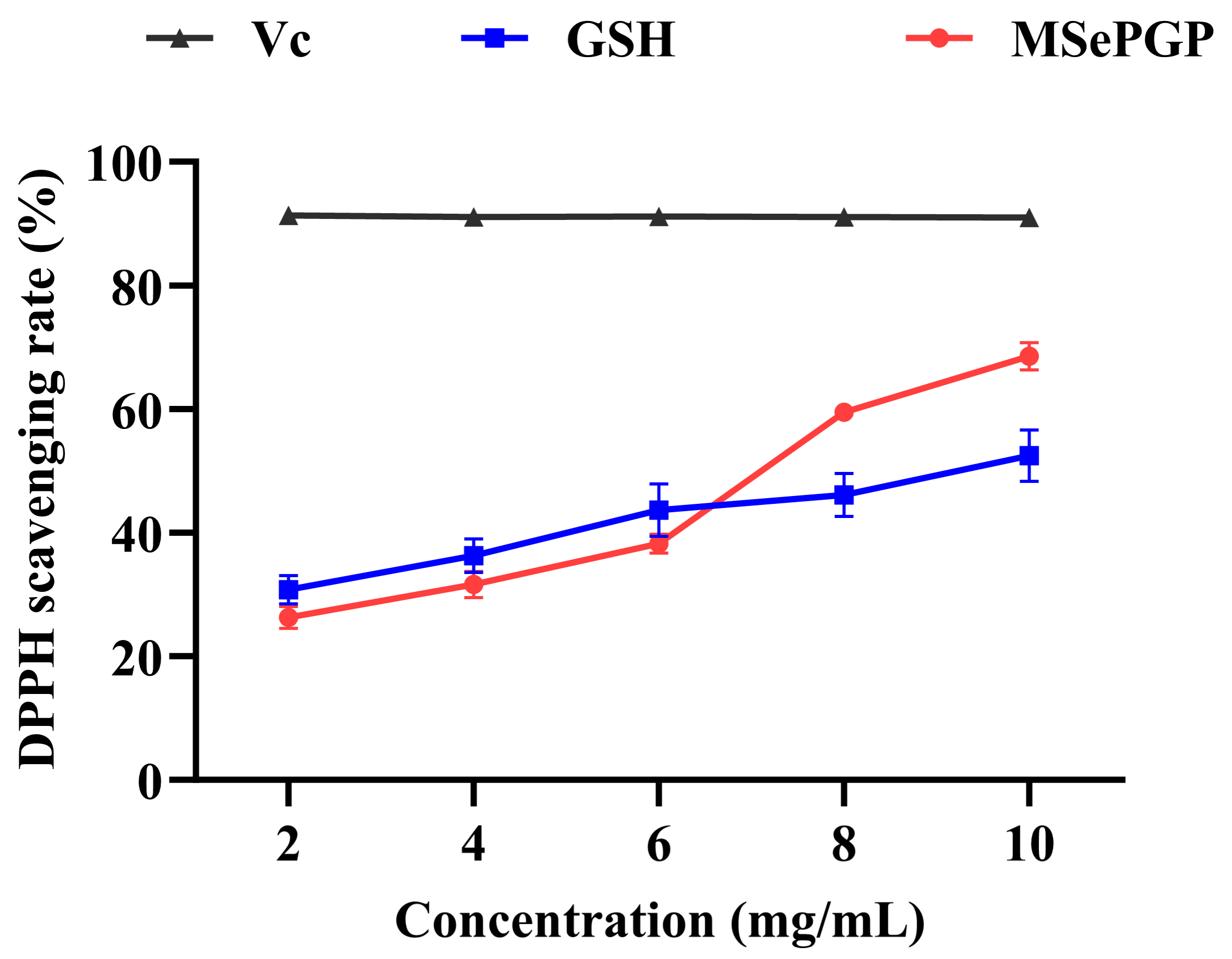
3.9. DPPH Radical Scavenging Rates of the Selenium-Containing Peptide
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Botha, L.K.; Namaumbo, S.; Kapito, N.J.; Ndovie, P.; Tsukuluza, D.C.; Jagot, F.; Mlangeni, A.T. Selenium health impacts and sub-saharan regional nutritional challenges: A review. Res. Chem. 2024, 12, 101920. [Google Scholar] [CrossRef]
- Rashid, M.T.; Liu, K.; Ning, M.; Ullah, K.; Wali, A.; Jatoi, M.A.; Muzaffar, N. Enhanced antioxidant activity of selenium-enriched brown rice protein against oxidative stress in mammalian erythrocytes under various cooking conditions. J. Agric. Food Res. 2024, 18, 101520. [Google Scholar] [CrossRef]
- Tutan, D.; Ulfberg, J.; Aydemir, N.; Eser, B.; Doğan, İ. Selenium, a notable micronutrient: A crucial element in the context of all-cause long-term mortality in renal failure. Biol. Trace Elem. Res. 2024. [Google Scholar] [CrossRef]
- Cai, J.; Su, W.; Chen, X.; Zheng, H. Advances in the study of selenium and human intestinal bacteria. Front. Nutr. 2022, 9, 1059358. [Google Scholar] [CrossRef]
- He, Z.; Gao, J.; Chen, X.; Ru, Y.; Zhang, D.; Pan, X. Efficient recovery of heavy metals and selenium from wastewater using granular sludge: The crucial role of glutathione (GSH). Water Res. 2025, 270, 122826. [Google Scholar] [CrossRef]
- De Souza, D.F.; Da Silva, M.D.S.; De Souza, T.C.; Rocha, G.C.; Kasuya, M.C.M.; Eller, M.R. Effect of selenium-enriched substrate on the chemical composition, mineral bioavailability, and yield of edible mushrooms. Biol. Trace Elem. Res. 2023, 201, 3077–3087. [Google Scholar] [CrossRef]
- Dong, Z.; Xiao, Y.; Wu, H. Selenium accumulation, speciation, and its effect on nutritive value of Flammulina velutipes (Golden needle mushroom). Food Chem. 2021, 350, 128667. [Google Scholar] [CrossRef]
- Yu, A.; Ji, Y.; Ma, G.; Xu, J.; Hu, Q. Identification and preparation of selenium-containing peptides from selenium-enriched Pleurotus eryngii and their protective effect on lead-induced oxidative damage in NCTC1469 hepatocytes. J. Sci. Food Agric. 2023, 103, 4522–4534. [Google Scholar] [CrossRef]
- Wang, Y.; Ji, Y.; Meng, K.; Zhang, J.; Zhong, L.; Zhan, Q.; Zhao, L. Effects of different selenium biofortification methods on Pleurotus eryngii polysaccharides: Structural characteristics, antioxidant activity, and binding capacity in vitro. Int. J. Biol. Macromol. 2024, 275, 133214. [Google Scholar] [CrossRef]
- Wang, B.; Zhao, N.; Li, J.; Xu, R.; Wang, T.; Guo, L.; Ma, M.; Fan, M.; Wei, X. Selenium-enriched Lactobacillus plantarum improves the antioxidant activity and flavor properties of fermented Pleurotus eryngii. Food Chem. 2021, 345, 128770. [Google Scholar] [CrossRef]
- Gąsecka, M.; Mleczek, M.; Siwulski, M.; Niedzielski, P. Phenolic composition and antioxidant properties of Pleurotus ostreatus and Pleurotus eryngii enriched with selenium and zinc. Eur. Food Res. Technol. 2016, 242, 723–773. [Google Scholar] [CrossRef]
- Wu, L.; Long, L.; Wen, X.; Qiu, D.; Yin, H.; Luo, K. Study on enzymatic preparation and antioxidant activity of selenium-rich peptides from kidney bean leaves. Cereals Oils 2023, 36, 83–89. [Google Scholar] [CrossRef]
- Li, L.; Jin, L.; Guo, P.; Liu, D.; Fu, J. Physicochemical properties, functional characteristics, and structure of selenium-enriched millet protein. Food Ferment. Ind. 2024, 50, 259–267. [Google Scholar] [CrossRef]
- Yu, T.; Guo, J.; Zhu, S.; Li, M.; Zhu, Z.; Cheng, S.; Wang, S.; Sun, Y.; Cong, X. Protective effects of selenium-enriched peptides from Cardaminutese violifolia against high-fat diet-induced obesity and its associated metabolic disorders in mice. Rsc Adv. 2020, 10, 31411–31424. [Google Scholar] [CrossRef]
- Wu, S.; Wu, Q.; Wang, J.; Li, Y.; Chen, B.; Zhu, Z.; Huang, R.; Chen, M.; Huang, A.; Xie, Y.; et al. Novel selenium peptides obtained from selenium-enriched Cordyceps militaris alleviate neuroinflammation and gut microbiota dysbacteriosis in LPS-injured mice. J. Agric. Food Chem. 2022, 70, 3194–3206. [Google Scholar] [CrossRef]
- Hallin, E.I.; Serkland, T.T.; Bjånes, T.K.; Skrede, S. High-throughput, low-cost quantification of 11 therapeutic antibodies using caprylic acid precipitation and LC-MS/MS. Anal. Chim. Acta 2024, 1313, 342789. [Google Scholar] [CrossRef]
- Olajide, O.E.; Zirpoli, M.; Kartowikromo, K.Y.; Zheng, J.; Hamid, A.M. Discriminutesation of common Escherichia coli strains in urine by liquid chromatography-ion mobility-tandem mass spectrometry and machine learning. J. Am. Soc. Mass Spectra. 2024, 35, 2706–2713. [Google Scholar] [CrossRef]
- Liu, X.; Liu, K. Optimization of preparation process and activity of selenium-enriched peanut peptides with cholesterol-lowering activity. China Oils Fats 2024, 49, 113–119. [Google Scholar] [CrossRef]
- Han, A.; Liu, H.; Dai, Y.; Sun, S.; Ma, H. Screening of umami peptides from fermented grains by machine learning, molecular docking, and molecular dynamics simulation. Food Biosci. 2024, 62, 105536. [Google Scholar] [CrossRef]
- Guo, Y.; Xu, M.; Hu, X.; Cen, L.; Pei, D.; Zhang, D.; Xu, J.; Shi, P.; Yang, L.; Cui, H. Extraction, purification, and mechanism of immunomodulatory peptides obtained from silkworm pupa protein hydrolysate. Int. J. Biol. Macromol. 2024, 283, 137863. [Google Scholar] [CrossRef]
- Chen, L.; Wu, H.; Xu, S.; Zhu, Z.; Xie, B.; Chen, B. Extraction process and antioxidant activities of selenopeptides from selenium-enriched Pleurotus eryngii. Mycosystema 2025, 44, 127–139. [Google Scholar] [CrossRef]
- Zhou, X.; Guo, T.; Lu, Y.; Hadiatullah, H.; Li, P.; Ding, K.; Zhao, G. Effects of amino acid composition of yeast extract on the microbiota and aroma quality of fermented soy sauce. Food Chem. 2022, 393, 133289. [Google Scholar] [CrossRef]
- Abid, K.; Rochat, B.; Lassahn, P.G.; Stöcklin, R.; Michalet, S.; Brakch, N.; Aubert, J.F.; Vatansever, B.; Tella, P.; De Meester, I.; et al. Kinetic study of neuropeptide Y (NPY) proteolysis in blood and identification of NPY3-35: A new peptide generated by plasma kallikrein. J. Biol. Chem. 2009, 284, 24715–24724. [Google Scholar] [CrossRef]
- Nema, R.; Vats, P.; Singh, J.; Srivastava, S.K.; Kumar, A. Competing endogenous TMPO-AS1-let-7c-5p- LDHA RNA network predicts the prognosis of lung adenocarcinoma patients. Asian Pac. J. Cancer Prev. 2024, 25, 3673–3689. [Google Scholar] [CrossRef]
- Wu, S.; Zhu, Z.; Chen, M.; Huang, A.; Xie, Y.; Hu, H.; Zhang, J.; Wu, Q.; Wang, J.; Ding, Y. Comparison of neuroprotection and regulating properties on gut microbiota between selenopeptide Val-Pro-Arg-Lys-Leu-SeMet and its native peptide Val-Pro-Arg-Lys-Leu-Met in vitro and in vivo. J. Agric. Food Chem. 2023, 71, 12203–12215. [Google Scholar] [CrossRef]
- Zhao, Q.; McClements, D.J.; Li, J.; Chang, C.; Su, Y.; Gu, L.; Yang, Y. Egg yolk selenopeptides: Preparation, characterization, and immunomodulatory activity. J. Agric. Food Chem. 2024, 72, 5237–5246. [Google Scholar] [CrossRef]
- Zhu, J.; Du, M.; Wu, M.; Yue, P.; Yang, X.; Wei, X.; Wang, Y. Preparation, physicochemical characterization, and identification of two novel mixed ACE-inhibiting peptides from two distinct tea alkali-soluble proteins. Eur. Food Res. Technol. 2020, 246, 1483–1494. [Google Scholar] [CrossRef]
- Xiao, Y.; Cai, W.; Zheng, Z.; Ma, H.; Huang, Q. Inhibition effect of Lignosus Rhinocerotis polysaccharides-selenium nanoparticles prepared by ultrasound treatment on non-enzymatic glycosylation. Food Sci. 2020, 41, 134–139. [Google Scholar] [CrossRef]
- Zhang, F.; Zhang, Y.; Luo, X.; Liu, P.; Lin, X.; Deng, Y.; Jiang, Y.; Chen, B. Purification and structural identification of polypeptides from fruiting bodies of Volvariella volvacea. Mycosystema 2020, 42, 1775–1784. [Google Scholar] [CrossRef]
- Wu, J.; Sun, N.; Lin, S.; Wu, W. Preparation and structural characterization of peptide-selenium complex from hoki (Macruronus novaezelandiae) skin gelatin. Food Sci. 2021, 42, 87–93. [Google Scholar] [CrossRef]
- Zhao, L.; Chen, Z.; Chen, H.; Hua, P.; Liu, B. Optimization of chelation of juncao Ganoderma lucidum peptides with selenium by response surface methodology. Food Sci. 2017, 38, 187–192. [Google Scholar] [CrossRef]
- Yang, Z.; Yin, H.; Wang, J. Structural characterization and antioxidant activity in vitro of selenized Bacillus subtilis peptidoglycan. J. Henan Univ. Technol. 2023, 44, 84–91. [Google Scholar] [CrossRef]
- Clarke, S.; Tamang, S.; Reiss, P.; Dahan, M. A simple and general route for monofunctionalization of fluorescent and magnetic nanoparticles using peptides. Nanotechnology 2011, 22, 175103. [Google Scholar] [CrossRef]
- Giordano, A.; Russo, C.; Raia, C.A.; Kuznetsova, I.M.; Stepanenko, O.V.; Turoverov, K.K. Highly UV-absorbing complex in selenomethionine-substituted alcohol dehydrogenase from Sulfolobus solfataricus. J. Proteome Res. 2004, 3, 613–620. [Google Scholar] [CrossRef]
- Borden, J.T.; Man, A.; Scott, D.A.; Liu, K.Z. Tobacco-induced alterations to the Fourier-transform infrared spectrum of serum. J. Mol. Med. 2003, 81, 788–794. [Google Scholar] [CrossRef]
- Jia, J.; Liu, Q.; Liu, H.; Yang, C.; Zhao, Q.; Xu, Y.; Wu, W. Structure characterization and antioxidant activity of abalone visceral peptides-selenium in vitro. Food Chem. 2024, 433, 137398. [Google Scholar] [CrossRef]
- Hu, Y.; Lian, W.; Xu, M.; Yuan, X. Preparation of selenium-polysaccharide from Huaishan yam and its inhibitory effect on α-glucosidase activity. J. Anhui Agric. Univ. 2023, 50, 911–916. [Google Scholar] [CrossRef]
- Chen, W.; Yue, L.; Jiang, Q.; Xia, W. Effect of chitosan with different molecular weights on the stability, antioxidant, and anticancer activities of well-dispersed selenium nanoparticles. IET Nanobiotechnol. 2019, 13, 30–35. [Google Scholar] [CrossRef]
- Wang, X.; Fu, J.; Bhullar, K.S.; Chen, B.; Liu, H.; Zhang, Y.; Wang, C.; Liu, C.; Su, D.; Ma, X.; et al. Identification, in silico selection, and mechanistic investigation of antioxidant peptides from corn gluten meal hydrolysate. Food Chem. 2024, 446, 138777. [Google Scholar] [CrossRef]
- Zhu, L.; Xie, C.; Su, Y.; Dong, Y.; Cheng, S.; He, J.; He, Y. Process optimization, structural characterization, and antioxidant activities of black pigment extracted from Enshi selenium-enriched Sesamum indicum L. LWT 2023, 173, 114223. [Google Scholar] [CrossRef]
- Terriente-Palacios, C.; Rubiño, S.; Hortós, M.; Peteiro, C.; Castellari, M. Taurine, homotaurine, GABA, and hydrophobic amino acids content influences “in vitro” antioxidant and SIRT1 modulation activities of enzymatic protein hydrolysates from algae. Sci. Rep. 2022, 12, 20832. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.; Yan, S.; Wang, P.; Fang, W.; Feng, G.; Bai, Y.; Xing, X.; Cai, H. Selenium-chelating peptides from the hydrolysates of degreased antarctic krill (Euphausia superba) powder: Preparation, structural characterization, and antioxidant properties. Food Biosci. 2024, 62, 105547. [Google Scholar] [CrossRef]
- Zhu, S.; Li, Y.; Chen, X.; Zhu, Z.; Li, S.; Song, J.; Zheng, Z.; Cong, X.; Cheng, S. Co-immobilization of Alcalase/Dispase for production of selenium-enriched peptide from Cardaminutese violifolia. Foods 2024, 13, 1753. [Google Scholar] [CrossRef]
- Pálla, T.; Mirzahosseini, A.; Noszál, B. Species-specific, pH-independent, standard redox potential of selenocysteine and selenocysteamine. Antioxidants 2020, 9, 465. [Google Scholar] [CrossRef] [PubMed]
- Ibrahim, M.; Muhammad, N.; Naeem, M.; Deobald, A.M.; Kamdem, J.P.; Rocha, J.B.T. In vitro evaluation of glutathione peroxidase (GPx)-like activity and antioxidant properties of an organoselenium compound. Toxicol. Vitr. 2015, 29, 947–952. [Google Scholar] [CrossRef]
- Khammuang, S.; Sarnthima, R.; Sanachai, K. Purification and identification of novel antioxidant peptides from silkworm pupae (Bombyx mori) protein hydrolysate and molecular docking study. Biocatal. Agric. Biotechnol. 2022, 42, 102367. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Amino Acid | Amount (mg/g) | |
|---|---|---|
| SePEPs | PEPs | |
| Glutamic acid (Glu) | 40.99 | 37.02 |
| Leucine (Leu) | 30.33 | 23.68 |
| Alanine (Ala) | 29.28 | 23.35 |
| Aspartic acid (Asp) | 25.92 | 21.18 |
| Valine (Val) | 23.64 | 17.78 |
| Phenylalanine (Phe) | 20.84 | 17.01 |
| Proline (Pro) | 15.85 | 8.68 |
| Isoleucine (Ile) | 15.51 | 13.07 |
| Lysine (Lys) | 14.89 | 13.44 |
| Glycine (Gly) | 13.98 | 14.91 |
| Serine (Ser) | 9.55 | 5.27 |
| Histidine (His) | 9.38 | 6.83 |
| Threonine (Thr) | 9.2 | 6.77 |
| Tryptophan (Tyr) | 8.44 | 7.26 |
| Arginine (Arg) | 8.15 | 5.10 |
| Methionine (Met) | 6.93 | 5.82 |
| Cysteine (Cys) | 0.9 | 1.59 |
| No. | Peptide | Mass (Da) | Length | ppm | m/z | z |
|---|---|---|---|---|---|---|
| 1 | CSeCL | 427.068 | 3 | −17.2 | 428.0679 | 1 |
| 2 | CSeGF | 415.065 | 3 | −3.6 | 416.0704 | 1 |
| 3 | FCSeH | 453.092 | 3 | 7.5 | 454.1022 | 1 |
| 4 | YGMSeT | 518.128 | 4 | 9.6 | 519.1402 | 1 |
| 5 | MSeCT | 443.063 | 3 | 13.9 | 440.0764 | 1 |
| 6 | CSeFA | 429.080 | 3 | −2.2 | 430.0866 | 1 |
| 7 | CSeACL | 498.105 | 4 | −14.4 | 499.1052 | 1 |
| 8 | SCSeSF | 490.097 | 4 | −4.9 | 491.1016 | 1 |
| 9 | CSeAF | 429.080 | 3 | −1.9 | 430.0867 | 1 |
| 10 | SCSePI | 466.133 | 4 | −13.9 | 467.1339 | 1 |
| 11 | SCSePL | 466.133 | 4 | −13.9 | 467.1339 | 1 |
| 12 | MSePGP | 490.133 | 4 | −10.5 | 491.1352 | 1 |
| 13 | CSeSPL | 466.133 | 4 | −13.9 | 467.1339 | 1 |
| 14 | CSePY | 429.080 | 3 | −4.2 | 430.0858 | 1 |
| 15 | CSeCPT | 512.084 | 4 | −13.8 | 513.0846 | 1 |
| 16 | FDGCSe | 488.081 | 4 | −3.2 | 489.0867 | 1 |
| No. | Peptide | ABTS | DPPH | FRAP | ORCA | Bio-Toxicity |
|---|---|---|---|---|---|---|
| 1 | CSeCL | Active | Non-active | Non-active | Non-active | Active |
| 2 | CSeGF | Non-active | Non-active | Non-active | Non-active | Active |
| 3 | FCSeH | Non-active | Non-active | Non-active | Non-active | Active |
| 4 | YGMSeT | Non-active | Non-active | Non-active | Non-active | Non-active |
| 5 | MSeCT | Non-active | Non-active | Non-active | Non-active | Active |
| 6 | CSeFA | Non-active | Non-active | Non-active | Non-active | Active |
| 7 | CSeACL | Non-active | Non-active | Active | Non-active | Active |
| 8 | SCSeSF | Active | Non-active | Non-active | Non-active | Active |
| 9 | CSeAF | Non-active | Non-active | Non-active | Non-active | Active |
| 10 | SCSePI | Active | Non-active | Non-active | Non-active | Active |
| 11 | SCSePL | Active | Active | Active | Active | Active |
| 12 | MSePGP | Active | Active | Active | Active | Non-active |
| 13 | CSeSPL | Active | Active | Active | Active | Active |
| No. | Name | Binding Energy |
|---|---|---|
| 1 | CCL | −3.0 |
| 2 | CGF | −3.8 |
| 3 | FCH | −3.9 |
| 4 | YGMT | −3.7 |
| 5 | MCT | −2.6 |
| 6 | CFA | −3.4 |
| 7 | CACL | −3.2 |
| 8 | SCSF | −3.8 |
| 9 | CAF | −4.0 |
| 10 | SCPI | −3.2 |
| 11 | SCPL | −3.1 |
| 12 | MPGP | −3.4 |
| 13 | CSPL | −2.8 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, L.; Nie, M.; Yang, J.; Zhang, W.; Hsiang, T.; Jiang, Y.; Xie, B.; Chen, B. Structural Identification and Molecular Interaction Modeling Analysis of Antioxidant Activity Selenium-Enriched Peptides from Selenium-Enriched Pleurotus eryngii. Antioxidants 2025, 14, 586. https://doi.org/10.3390/antiox14050586
Chen L, Nie M, Yang J, Zhang W, Hsiang T, Jiang Y, Xie B, Chen B. Structural Identification and Molecular Interaction Modeling Analysis of Antioxidant Activity Selenium-Enriched Peptides from Selenium-Enriched Pleurotus eryngii. Antioxidants. 2025; 14(5):586. https://doi.org/10.3390/antiox14050586
Chicago/Turabian StyleChen, Lili, Menghan Nie, Jing Yang, Weibin Zhang, Tom Hsiang, Yuji Jiang, Baogui Xie, and Bingzhi Chen. 2025. "Structural Identification and Molecular Interaction Modeling Analysis of Antioxidant Activity Selenium-Enriched Peptides from Selenium-Enriched Pleurotus eryngii" Antioxidants 14, no. 5: 586. https://doi.org/10.3390/antiox14050586
APA StyleChen, L., Nie, M., Yang, J., Zhang, W., Hsiang, T., Jiang, Y., Xie, B., & Chen, B. (2025). Structural Identification and Molecular Interaction Modeling Analysis of Antioxidant Activity Selenium-Enriched Peptides from Selenium-Enriched Pleurotus eryngii. Antioxidants, 14(5), 586. https://doi.org/10.3390/antiox14050586