
Foliar Spray of Cerium Oxide Nanoparticles (CeO2 NPs) Improves Lead (Pb) Resistance in Rice

 , ,
, ,
Abstract
1. Introduction
2. Materials and Methods
2.1. Synthesis of CeO2 NPs
2.2. Experimental Design
2.3. Morphology and Fresh Weight Determination
2.4. Photosynthesis-Related Index Detection
2.5. Antioxidant Enzyme Activity Detection
2.6. Transcriptome Detection
2.7. Metabolome Detection
2.8. Real-Time Quantitative PCR
2.9. Statistical Analysis
3. Results
3.1. Morphology and Fresh Weight
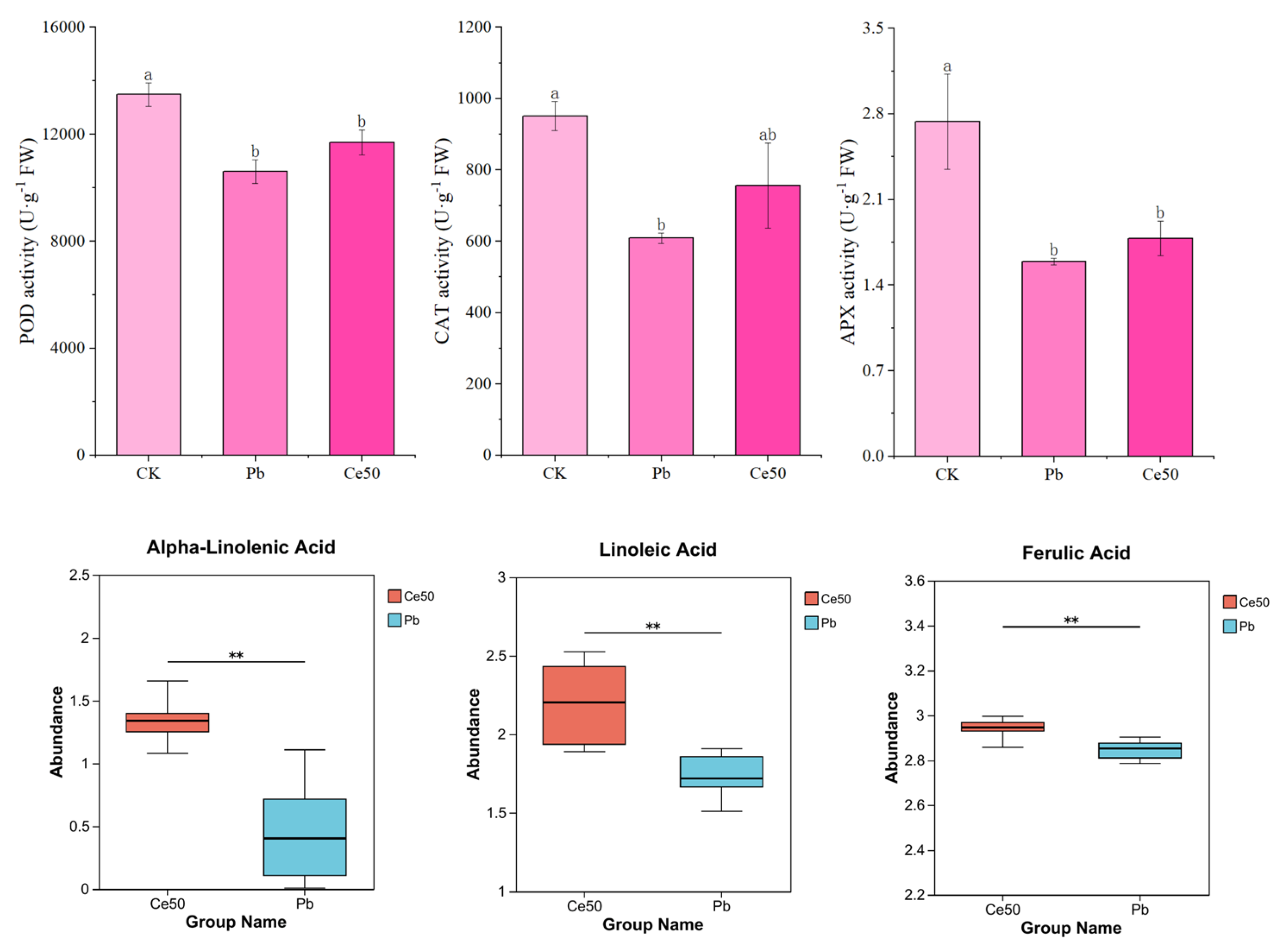
3.2. Antioxidant Enzyme Activity and Antioxidant Metabolite Abundance
3.3. Photosynthetic Pigment Content
3.4. Photosynthesis
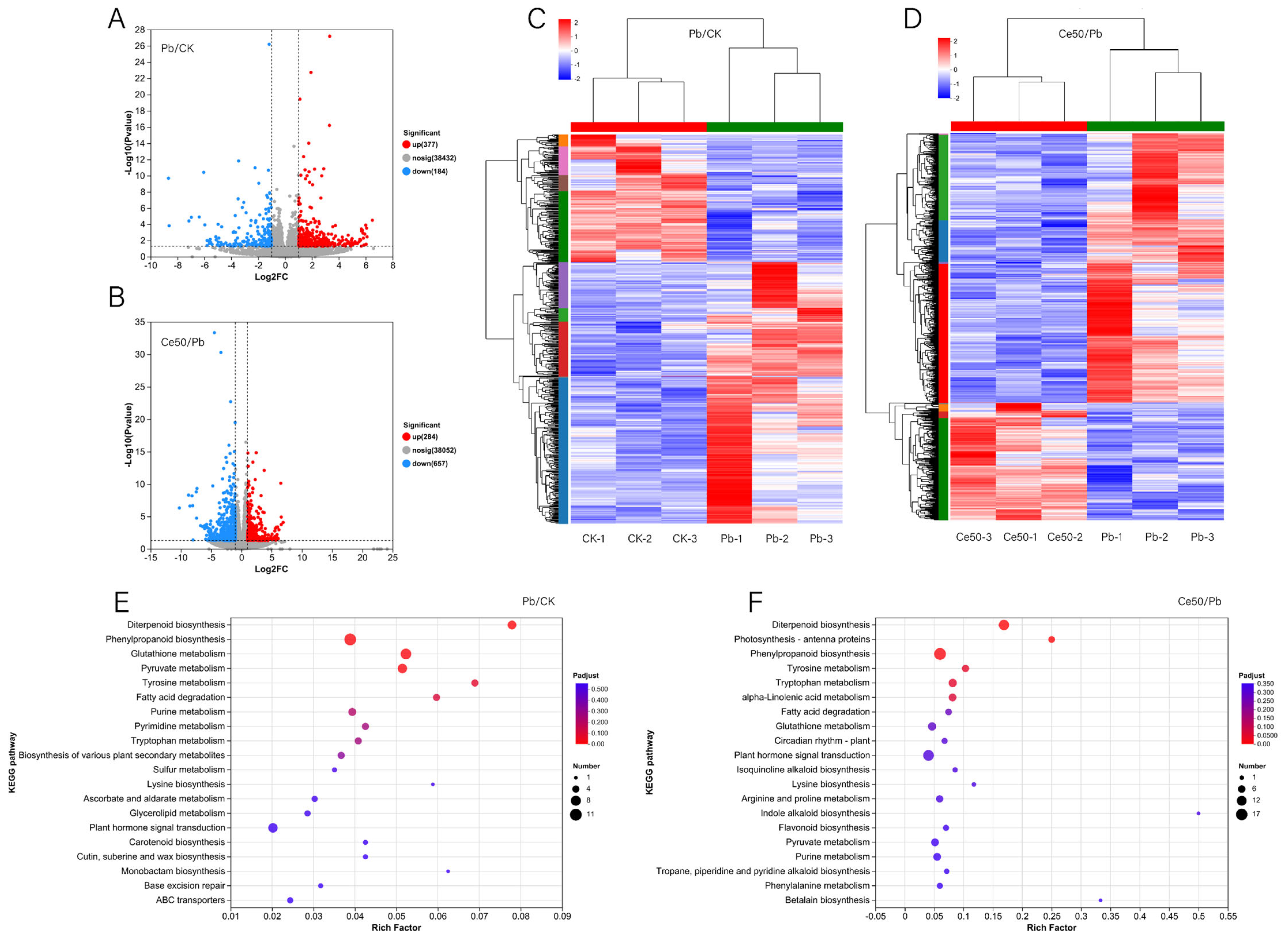
3.5. Transcriptome
3.5.1. Quality Control
3.5.2. Differentially Expressed Gene (DEG) Statistics
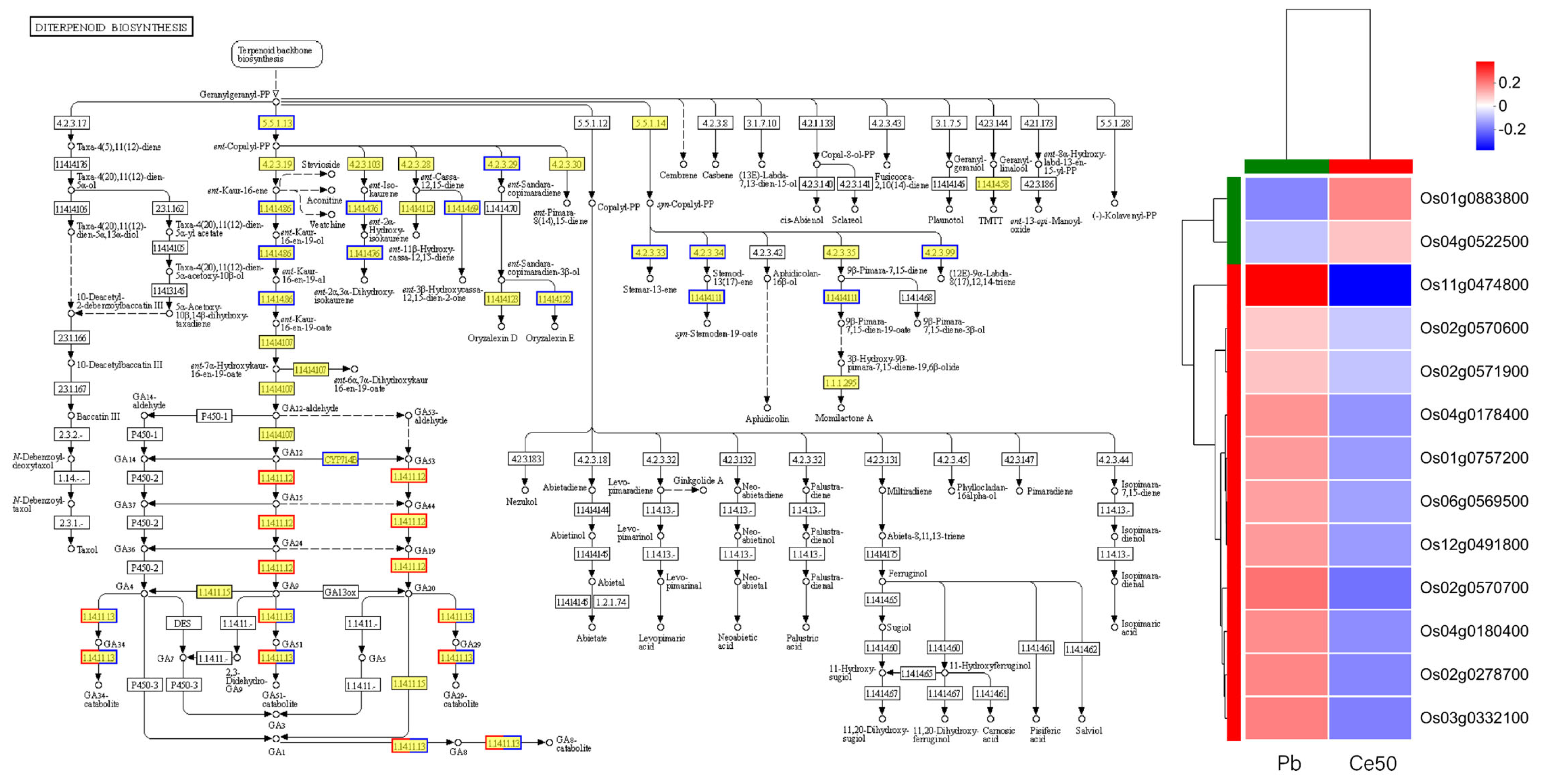
3.5.3. KEGG Annotation, KEGG Enrichment Analysis, and GO Enrichment Analysis
3.5.4. TF Gene Statistics
3.5.5. Expression of Stress Resistance-Related Genes
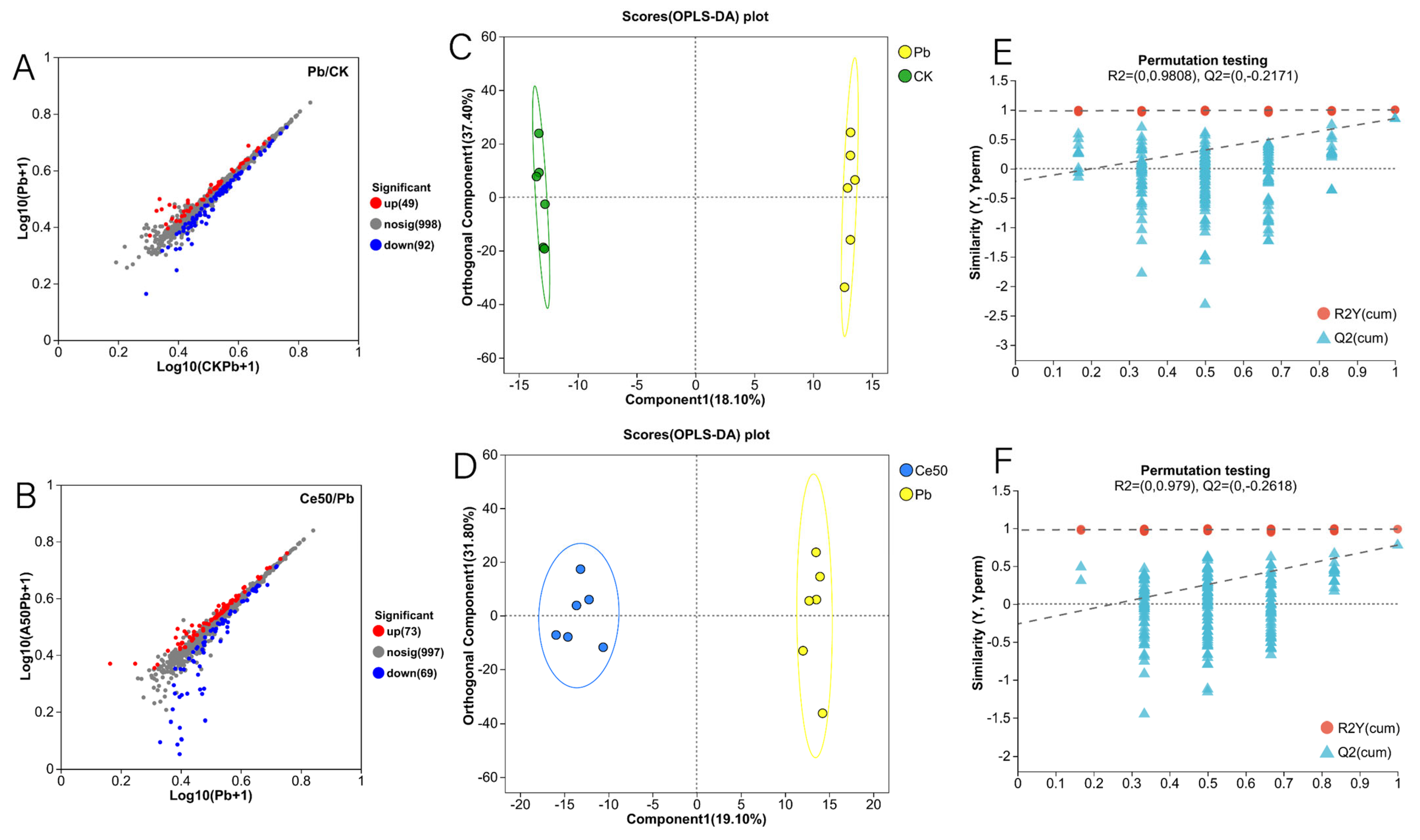
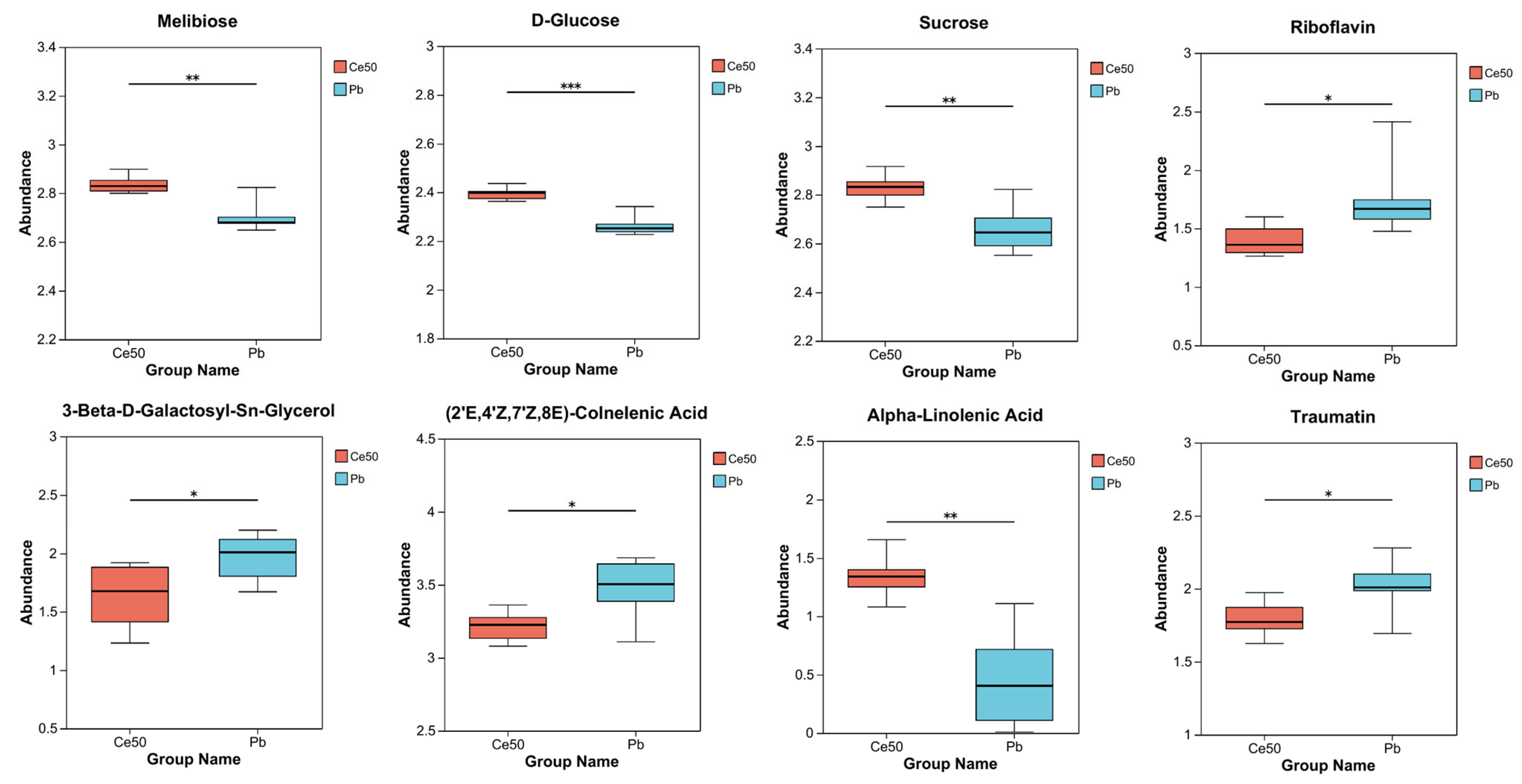
3.6. Metabolome
3.6.1. Differential Metabolite Statistics, OPLS-DA, and OPLS-DA Permutation Test
3.6.2. KEGG Enrichment Analysis
3.6.3. VIP Analysis
3.6.4. KEGG Compound Classification
4. Discussion
4.1. Physiological Responses of Rice to CeO2 NPs Under Pb Stress
4.2. Response of Rice Metabolism to CeO2 NPs Under Pb Stress
4.3. Response of Rice Gene Expression to CeO2 NPs Under Pb Stress
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wołejko, E.; Jabłońska-Trypuć, A.; Wydro, U.; Butarewicz, A.; Łozowicka, B. Soil biological activity as an indicator of soil pollution with pesticides—A review. Appl. Soil Ecol. 2020, 147, 103356. [Google Scholar] [CrossRef]
- Obaje, S.O.; Ogunyele, A.C.; Adeola, A.O.; Akingboye, A.S. Assessment of stream sediments pollution by potentially toxic elements in the active mining area of Okpella, Edo State, Nigeria. Rud.-Geološko-Naft. Zb. 2019, 34, 43–50. [Google Scholar] [CrossRef]
- Gupta, S.; Jena, V.; Matić, N.; Kapralova, V.; Solanki, J. Assessment of geo-accumulation indeks of heavy metal and source of contamination by multivariate factor analysis. Int. J. Hazard. Mater. 2014, 2, 18–22. [Google Scholar]
- Ale, T.O.; Ogunribido, T.H.; Ademila, O.; Akingboye, A.S. Soil pollution status due to potentially toxic elements in active open dumpsites: Insights from different Nigerian geological environments. Environ. Earth Sci. 2024, 83, 535. [Google Scholar] [CrossRef]
- Yang, H.; Wang, F.; Yu, J.; Huang, K.; Zhang, H.; Fu, Z. An improved weighted index for the assessment of heavy metal pollution in soils in Zhejiang, China. Environ. Res. 2021, 192, 110246. [Google Scholar] [CrossRef] [PubMed]
- Li, T.; Liu, Y.; Lin, S.; Liu, Y.; Xie, Y. Soil Pollution Management in China: A Brief Introduction. Sustainability 2019, 11, 556. [Google Scholar] [CrossRef]
- Yang, H.; Huang, X.; Thompson, J.R.; Flower, R.J. Soil pollution: Urban brownfields. Science 2014, 344, 691–692. [Google Scholar] [CrossRef]
- Chen, R.; De Sherbinin, A.; Ye, C.; Shi, G. China’s soil pollution: Farms on the frontline. Science 2014, 344, 691. [Google Scholar] [CrossRef]
- Yang, Q.; Li, Z.; Lu, X.; Duan, Q.; Huang, L.; Bi, J. A review of soil heavy metal pollution from industrial and agricultural regions in China: Pollution and risk assessment. Sci. Total Environ. 2018, 642, 690–700. [Google Scholar] [CrossRef]
- Peng, J.-Y.; Zhang, S.; Han, Y.; Bate, B.; Ke, H.; Chen, Y. Soil heavy metal pollution of industrial legacies in China and health risk assessment. Sci. Total Environ. 2022, 816, 151632. [Google Scholar] [CrossRef]
- Kumar, S.; Rahman, M.A.; Islam, M.R.; Hashem, M.A.; Rahman, M.M. Lead and other elements-based pollution in soil, crops and water near a lead-acid battery recycling factory in Bangladesh. Chemosphere 2022, 290, 133288. [Google Scholar] [CrossRef] [PubMed]
- Kushwaha, A.; Hans, N.; Kumar, S.; Rani, R. A critical review on speciation, mobilization and toxicity of lead in soil-microbe-plant system and bioremediation strategies. Ecotoxicol. Environ. Saf. 2018, 147, 1035–1045. [Google Scholar] [CrossRef] [PubMed]
- Singh, R.P.; Tripathi, R.D.; Sinha, S.; Maheshwari, R.; Srivastava, H. Response of higher plants to lead contaminated environment. Chemosphere 1997, 34, 2467–2493. [Google Scholar] [CrossRef]
- Qing, Z. Effect of La on Glycine max seedling under Pb stress. Chin. J. Appl. Environ. Biol. 1999, 5, 22–25. [Google Scholar]
- Pang, X.; Wang, D.; Xing, X.; Peng, A.; Zhang, F.; Li, C. Effect of La3+ on the activities of antioxidant enzymes in wheat seedlings under lead stress in solution culture. Chemosphere 2002, 47, 1033–1039. [Google Scholar] [CrossRef]
- Ahmad, M.S.A.; Ashraf, M.; Tabassam, Q.; Hussain, M.; Firdous, H. Lead (Pb)-induced regulation of growth, photosynthesis, and mineral nutrition in maize (Zea mays L.) plants at early growth stages. Biol. Trace Elem. Res. 2011, 144, 1229–1239. [Google Scholar] [CrossRef]
- Zhou, J.; Jiang, Z.; Ma, J.; Yang, L.; Wei, Y. The effects of lead stress on photosynthetic function and chloroplast ultrastructure of Robinia pseudoacacia seedlings. Environ. Sci. Pollut. Res. 2017, 24, 10718–10726. [Google Scholar] [CrossRef]
- Sharma, P.; Dubey, R.S. Lead toxicity in plants. Braz. J. Plant Physiol. 2005, 17, 35–52. [Google Scholar] [CrossRef]
- Yang, Y.; Han, X.; Liang, Y.; Ghosh, A.; Chen, J.; Tang, M. The Combined Effects of Arbuscular Mycorrhizal Fungi (AMF) and Lead (Pb) Stress on Pb Accumulation, Plant Growth Parameters, Photosynthesis, and Antioxidant Enzymes in Robinia pseudoacacia L. PLoS ONE 2015, 10, e0145726. [Google Scholar] [CrossRef]
- Rahman, S.U.; Qin, A.; Zain, M.; Mushtaq, Z.; Mehmood, F.; Riaz, L.; Naveed, S.; Ansari, M.J.; Saeed, M.; Ahmad, I. Pb uptake, accumulation, and translocation in plants: Plant physiological, biochemical, and molecular response: A review. Heliyon 2024, 10, e27724. [Google Scholar] [CrossRef]
- Hu, J.; Shi, G.; Xu, Q.; Wang, X.; Yuan, Q.; Du, K. Effects of Pb2+ on the active oxygen-scavenging enzyme activities and ultrastructure in Potamogeton crispus leaves. Russ. J. Plant Physiol. 2007, 54, 414–419. [Google Scholar] [CrossRef]
- Maleki, M.; Ghorbanpour, M.; Kariman, K. Physiological and antioxidative responses of medicinal plants exposed to heavy metals stress. Plant Gene 2017, 11, 247–254. [Google Scholar] [CrossRef]
- Sarraf, M.; Vishwakarma, K.; Kumar, V.; Arif, N.; Das, S.; Johnson, R.; Janeeshma, E.; Puthur, J.T.; Aliniaeifard, S.; Chauhan, D.K. Metal/metalloid-based nanomaterials for plant abiotic stress tolerance: An overview of the mechanisms. Plants 2022, 11, 316. [Google Scholar] [CrossRef] [PubMed]
- Preetha, J.S.Y.; Sriram, D.; Premasudha, P.; Pudake, R.N.; Arun, M. Cerium oxide as a nanozyme for plant abiotic stress tolerance: An overview of the mechanisms. Plant Nano Biol. 2023, 6, 100049. [Google Scholar] [CrossRef]
- Fallah Imani, A.; Gomarian, M.; Ghorbanpour, M.; Ramak, P.; Chavoshi, S. Foliar-applied nano-cerium dioxide differentially affect morpho-physiological traits and essential oil profile of Salvia mirzayanii Rech. f. & Esfand under drought stress and post-stress recovery conditions. Plant Physiol. Biochem. 2023, 203, 108046. [Google Scholar] [CrossRef]
- Ayub, M.A.; Ahmad, H.R.; Zia ur Rehman, M.; Waraich, E.A. Cerium oxide nanoparticles alleviates stress in wheat grown on Cd contaminated alkaline soil. Chemosphere 2023, 338, 139561. [Google Scholar] [CrossRef]
- An, J.; Hu, P.; Li, F.; Wu, H.; Shen, Y.; White, J.C.; Tian, X.; Li, Z.; Giraldo, J.P. Emerging investigator series: Molecular mechanisms of plant salinity stress tolerance improvement by seed priming with cerium oxide nanoparticles. Environ. Sci. Nano 2020, 7, 2214–2228. [Google Scholar] [CrossRef]
- Zhang, R.; Zheng, D.; Feng, N.; Qiu, Q.-S.; Zhou, H.; Liu, M.; Li, Y.; Meng, F.; Huang, X.; Huang, A. Prohexadione calcium enhances rice growth and tillering under NaCl stress. PeerJ 2023, 11, e14804. [Google Scholar] [CrossRef]
- Creelman, R.A.; Mullet, J.E. Biosynthesis and action of jasmonates in plants. Annu. Rev. Plant Biol. 1997, 48, 355–381. [Google Scholar] [CrossRef]
- Wasternack, C.; Feussner, I. The oxylipin pathways: Biochemistry and function. Annu. Rev. Plant Biol. 2018, 69, 363–386. [Google Scholar] [CrossRef]
- Zi, X.; Zhou, S.; Wu, B. Alpha-linolenic acid mediates diverse drought responses in maize (Zea mays L.) at seedling and flowering stages. Molecules 2022, 27, 771. [Google Scholar] [CrossRef] [PubMed]
- Ou, Y.; Teng, Z.; Shu, Y.; Wang, Y.; Wang, D.; Sun, C.; Lin, X. Linoleic acid alleviates aluminum toxicity by modulating fatty acid composition and redox homeostasis in wheat (Triticum aestivum) seedlings. J. Hazard. Mater. 2025, 487, 137156. [Google Scholar] [CrossRef] [PubMed]
- Quan, N.T.; Xuan, T.D. Foliar application of vanillic and p-hydroxybenzoic acids enhanced drought tolerance and formation of phytoalexin momilactones in rice. Arch. Agron. Soil Sci. 2018, 64, 1831–1846. [Google Scholar] [CrossRef]
- Khan, K.A.; Saleem, M.H.; Afzal, S.; Hussain, I.; Ameen, F.; Fahad, S. Ferulic acid: Therapeutic potential due to its antioxidant properties, role in plant growth, and stress tolerance. Plant Growth Regul. 2024, 104, 1329–1353. [Google Scholar] [CrossRef]
- Chen, H.-C.; Zhang, S.-L.; Wu, K.-J.; Li, R.; He, X.-R.; He, D.-N.; Huang, C.; Wei, H. The effects of exogenous organic acids on the growth, photosynthesis and cellular ultrastructure of Salix variegata Franch. Under Cd stress. Ecotoxicol. Environ. Saf. 2020, 187, 109790. [Google Scholar] [CrossRef]
- Si, P.; Shao, W.; Yu, H.; Xu, G.; Du, G. Differences in microbial communities stimulated by malic acid have the potential to improve nutrient absorption and fruit quality of grapes. Front. Microbiol. 2022, 13, 850807. [Google Scholar] [CrossRef]
- KiliÇ, T. Seed treatments with salicylic and succinic acid to mitigate drought stress in flowering kale cv. ‘Red Pigeon F1’. Sci. Hortic. 2023, 313, 111939. [Google Scholar] [CrossRef]
- Yoshikawa, M.; Hirai, N.; Wakabayashi, K.; Sugizaki, H.; Iwamura, H. Succinic and lactic acids as plant growth promoting compounds produced by rhizospheric Pseudomonas putida. Can. J. Microbiol. 1993, 39, 1150–1154. [Google Scholar] [CrossRef]
- Elzayat, H.E.; Taha, N.M.; Shakweer, N.H. Spraying biostimulants on Le Conte pear trees reduces fruit drop and enhances yield, improves fruit quality, and storability. Egypt. J. Agric. Res. 2024, 102, 251–261. [Google Scholar] [CrossRef]
- Zhang, Y.; Fernie, A.R. The role of TCA cycle enzymes in plants. Adv. Biol. 2023, 7, 2200238. [Google Scholar] [CrossRef]
- Lemoine, R. Sucrose transporters in plants: Update on function and structure. Biochim. Et Biophys. Acta (BBA)-Biomembr. 2000, 1465, 246–262. [Google Scholar] [CrossRef]
- Aluko, O.O.; Li, C.; Wang, Q.; Liu, H. Sucrose utilization for improved crop yields: A review article. Int. J. Mol. Sci. 2021, 22, 4704. [Google Scholar] [CrossRef] [PubMed]
- Thomas, A.; Beena, R. Sucrose Metabolism in Plants under Drought Stress Condition: A Review. Indian J. Agric. Res. 2024, 58, 943. [Google Scholar] [CrossRef]
- Siddiqui, H.; Sami, F.; Hayat, S. Glucose: Sweet or bitter effects in plants-a review on current and future perspective. Carbohydr. Res. 2020, 487, 107884. [Google Scholar] [CrossRef]
- De Luna, S.R.; Ramirez-Garza, R.; Saldivar, S.S. Review Article–Environmentally Friendly Methods for Flavonoid Extraction from Plant Material: Impact of Their Operating Conditions on Yield and Antioxidant Properties. Sci. World J. 2020, 2020, 6792069. [Google Scholar]
- Dias, M.C.; Pinto, D.C.; Silva, A.M. Plant flavonoids: Chemical characteristics and biological activity. Molecules 2021, 26, 5377. [Google Scholar] [CrossRef]
- Shomali, A.; Das, S.; Arif, N.; Sarraf, M.; Zahra, N.; Yadav, V.; Aliniaeifard, S.; Chauhan, D.K.; Hasanuzzaman, M. Diverse Physiological Roles of Flavonoids in Plant Environmental Stress Responses and Tolerance. Plants 2022, 11, 3158. [Google Scholar] [CrossRef]
- Ramzan, M.; Haider, S.T.A.; Hussain, M.B.; Ehsan, A.; Datta, R.; Alahmadi, T.A.; Ansari, M.J.; Alharbi, S.A. Potential of kaempferol and caffeic acid to mitigate salinity stress and improving potato growth. Sci. Rep. 2024, 14, 21657. [Google Scholar] [CrossRef]
- Conrath, U.; Beckers, G.J.; Flors, V.; García-Agustín, P.; Jakab, G.; Mauch, F.; Newman, M.-A.; Pieterse, C.M.; Poinssot, B.; Pozo, M.J. Priming: Getting ready for battle. Mol. Plant-Microbe Interact. 2006, 19, 1062–1071. [Google Scholar] [CrossRef]
- Frost, C.J.; Mescher, M.C.; Carlson, J.E.; De Moraes, C.M. Plant defense priming against herbivores: Getting ready for a different battle. Plant Physiol. 2008, 146, 818–824. [Google Scholar] [CrossRef]
- Krokene, P. Conifer defense and resistance to bark beetles. In Bark Beetles; Elsevier: Amsterdam, The Netherlands, 2015; pp. 177–207. [Google Scholar]
- Wu, L.; Wang, R.; Li, M.; Du, Z.; Jin, Y.; Shi, Y.; Jiang, W.; Chen, J.; Jiao, Y.; Hu, B. Functional analysis of a rice 12-oxo-phytodienoic acid reductase gene (OsOPR1) involved in Cd stress tolerance. Mol. Biol. Rep. 2024, 51, 198. [Google Scholar] [CrossRef] [PubMed]
- Agrawal, G.K.; Jwa, N.-S.; Shibato, J.; Han, O.; Iwahashi, H.; Rakwal, R. Diverse environmental cues transiently regulate OsOPR1 of the “octadecanoid pathway” revealing its importance in rice defense/stress and development. Biochem. Biophys. Res. Commun. 2003, 310, 1073–1082. [Google Scholar] [CrossRef] [PubMed]
- Huang, L.-F.; Lin, K.-H.; He, S.-L.; Chen, J.-L.; Jiang, J.-Z.; Chen, B.-H.; Hou, Y.-S.; Chen, R.-S.; Hong, C.-Y.; Ho, S.-L. Multiple patterns of regulation and overexpression of a ribonuclease-like pathogenesis-related protein gene, OsPR10a, conferring disease resistance in rice and Arabidopsis. PLoS ONE 2016, 11, e0156414. [Google Scholar] [CrossRef] [PubMed]
- McGee, J.D.; Hamer, J.E.; Hodges, T.K. Characterization of a PR-10 pathogenesis-related gene family induced in rice during infection with Magnaporthe grisea. Mol. Plant-Microbe Interact. 2001, 14, 877–886. [Google Scholar] [CrossRef]
- Zhou, J.; Deng, K.; Cheng, Y.; Zhong, Z.; Tian, L.; Tang, X.; Tang, A.; Zheng, X.; Zhang, T.; Qi, Y. CRISPR-Cas9 based genome editing reveals new insights into microRNA function and regulation in rice. Front. Plant Sci. 2017, 8, 1598. [Google Scholar] [CrossRef]
- Wang, R.; Jing, W.; Xiao, L.; Jin, Y.; Shen, L.; Zhang, W. The rice high-affinity potassium transporter1; 1 is involved in salt tolerance and regulated by an MYB-type transcription factor. Plant Physiol. 2015, 168, 1076–1090. [Google Scholar] [CrossRef]
- Zhou, H.; Wu, H.; Zhang, F.; Su, Y.; Guan, W.; Xie, Y.; Giraldo, J.P.; Shen, W. Molecular basis of cerium oxide nanoparticle enhancement of rice salt tolerance and yield. Environ. Sci. Nano 2021, 8, 3294–3311. [Google Scholar] [CrossRef]
- Li, L.; Li, N.; Song, S.; Li, Y.; Xia, X.; Fu, X.; Chen, G.; Deng, H. Cloning and characterization of the drought-resistance OsRCI2-5 gene in rice (Oryza sativa L.). Genet. Mol. Res. 2014, 13, 4022–4035. [Google Scholar] [CrossRef]
- Xiong, H.; Yu, J.; Miao, J.; Li, J.; Zhang, H.; Wang, X.; Liu, P.; Zhao, Y.; Jiang, C.; Yin, Z. Natural variation in OsLG3 increases drought tolerance in rice by inducing ROS scavenging. Plant Physiol. 2018, 178, 451–467. [Google Scholar] [CrossRef]
- Qian, Y.; Zhang, T.; Yu, Y.; Gou, L.; Yang, J.; Xu, J.; Pi, E. Regulatory mechanisms of bHLH transcription factors in plant adaptive responses to various abiotic stresses. Front. Plant Sci. 2021, 12, 677611. [Google Scholar] [CrossRef]
- Wagan, S.; Ali, M.; Khoso, M.A.; Alam, I.; Dinislam, K.; Hussain, A.; Brohi, N.A.; Manghwar, H.; Liu, F. Deciphering the role of WRKY transcription factors in plant resilience to alkaline salt stress. Plant Stress 2024, 13, 100526. [Google Scholar] [CrossRef]
- Mariyam, S.; Kumar, V.; Roychoudhury, A.; Ghodake, G.S.; Muneer, S.; Duhan, J.S.; Ahmad, F.; Sharma, R.K.; Singh, J.; Seth, C.S. Functional Diversification and Mechanistic Insights of MYB Transcription Factors in Mediating Plant Growth and Development, Secondary Metabolism, and Stress Responses. J. Plant Growth Regul. 2025, 44, 1465–1484. [Google Scholar] [CrossRef]
- Nakashima, K.; Takasaki, H.; Mizoi, J.; Shinozaki, K.; Yamaguchi-Shinozaki, K. NAC transcription factors in plant abiotic stress responses. Biochim. Et Biophys. Acta (BBA)-Gene Regul. Mech. 2012, 1819, 97–103. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment | First Sampling | Second Sampling | |
|---|---|---|---|
| CK | 39.142 ± 0.334 b | 42.667 ± 1.504 a | |
| Total chlorophyll content (mg·g−1) | Pb | 41.628 ± 0.395 a | 38.965 ± 0.494 b |
| Ce50 | 41.740 ± 0.598 a | 40.025 ± 0.413 ab | |
| CK | 27.897 ± 0.176 b | 29.086 ± 0.430 a | |
| Chlorophyll a content (mg·g−1) | Pb | 28.801 ± 0.176 a | 27.794 ± 0.233 b |
| Ce50 | 28.813 ± 0.181 a | 28.197 ± 0.139 ab | |
| CK | 11.245 ± 0.162 b | 13.581 ± 1.094 a | |
| Chlorophyll b content (mg·g−1) | Pb | 12.827 ± 0.226 a | 11.171 ± 0.270 b |
| Ce50 | 12.928 ± 0.417 a | 11.827 ± 0.277 ab | |
| CK | 5.756 ± 0.026 b | 5.975 ± 0.068 a | |
| Carotenoid content (mg·g−1) | Pb | 6.147 ± 0.058 a | 5.739 ± 0.039 b |
| Ce50 | 6.176 ± 0.022 a | 5.938 ± 0.018 a |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhou, H.; Liu, J.; Chen, Z.; An, J.; Huo, J.; Bu, Q.; Su, T.; Zhao, L.; Shen, X.; Xue, Y.; et al. Foliar Spray of Cerium Oxide Nanoparticles (CeO2 NPs) Improves Lead (Pb) Resistance in Rice. Antioxidants 2025, 14, 552. https://doi.org/10.3390/antiox14050552
Zhou H, Liu J, Chen Z, An J, Huo J, Bu Q, Su T, Zhao L, Shen X, Xue Y, et al. Foliar Spray of Cerium Oxide Nanoparticles (CeO2 NPs) Improves Lead (Pb) Resistance in Rice. Antioxidants. 2025; 14(5):552. https://doi.org/10.3390/antiox14050552
Chicago/Turabian StyleZhou, Hang, Junjie Liu, Ziyang Chen, Jing An, Jingxin Huo, Qing Bu, Tao Su, Liming Zhao, Xuefeng Shen, Yingbin Xue, and et al. 2025. "Foliar Spray of Cerium Oxide Nanoparticles (CeO2 NPs) Improves Lead (Pb) Resistance in Rice" Antioxidants 14, no. 5: 552. https://doi.org/10.3390/antiox14050552
APA StyleZhou, H., Liu, J., Chen, Z., An, J., Huo, J., Bu, Q., Su, T., Zhao, L., Shen, X., Xue, Y., Rao, G., Feng, N., Zheng, D., & Zhang, R. (2025). Foliar Spray of Cerium Oxide Nanoparticles (CeO2 NPs) Improves Lead (Pb) Resistance in Rice. Antioxidants, 14(5), 552. https://doi.org/10.3390/antiox14050552