
HSA Adductomics in the Shanghai Women’s Health Study Links Lung Cancer in Never-Smokers with Air Pollution, Redox Biology, and One-Carbon Metabolism

 , , , , ,
, , , , ,
Abstract

1. Introduction
2. Materials and Methods
2.1. Study Population
2.2. Lung Cancer Ascertainment
2.3. Mass Spectrometry and Data Acquisition
2.4. Statistical Analysis
3. Results
4. Discussion
4.1. Air Pollution and Lung Cancer in Never-Smokers
4.2. ROS Production and Removal
4.3. The Role of OCM in Lung Cancer
4.4. A Connection to Microbial Translocation?
4.5. Influence of Covariates
4.6. Strengths and Weaknesses
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
References
- LoPiccolo, J.; Gusev, A.; Christiani, D.C.; Janne, P.A. Lung cancer in patients who have never smoked—An emerging disease. Nat. Rev. Clin. Oncol. 2024, 21, 121–146. [Google Scholar] [CrossRef]
- Epplein, M.; Schwartz, S.M.; Potter, J.D.; Weiss, N.S. Smoking-adjusted lung cancer incidence among Asian-Americans (United States). Cancer Causes Control CCC 2005, 16, 1085–1090. [Google Scholar] [CrossRef]
- Zhong, L.; Goldberg, M.S.; Gao, Y.T.; Jin, F. A case-control study of lung cancer and environmental tobacco smoke among nonsmoking women living in Shanghai, China. Cancer Causes Control CCC 1999, 10, 607–616. [Google Scholar] [CrossRef] [PubMed]
- Hosgood, H.D., 3rd; Pao, W.; Rothman, N.; Hu, W.; Pan, Y.H.; Kuchinsky, K.; Jones, K.D.; Xu, J.; Vermeulen, R.; Simko, J.; et al. Driver mutations among never smoking female lung cancer tissues in China identify unique EGFR and KRAS mutation pattern associated with household coal burning. Respir. Med. 2013, 107, 1755–1762. [Google Scholar] [CrossRef] [PubMed]
- Yu, I.T.; Chiu, Y.L.; Au, J.S.; Wong, T.W.; Tang, J.L. Dose-response relationship between cooking fumes exposures and lung cancer among Chinese nonsmoking women. Cancer Res. 2006, 66, 4961–4967. [Google Scholar] [CrossRef] [PubMed]
- Yang, G.; Shu, X.O.; Chow, W.H.; Zhang, X.; Li, H.L.; Ji, B.T.; Cai, H.; Wu, S.; Gao, Y.T.; Zheng, W. Soy food intake and risk of lung cancer: Evidence from the Shanghai Women’s Health Study and a meta-analysis. Am. J. Epidemiol. 2012, 176, 846–855. [Google Scholar] [CrossRef]
- Takata, Y.; Shu, X.O.; Yang, G.; Li, H.; Dai, Q.; Gao, J.; Cai, Q.; Gao, Y.T.; Zheng, W. Calcium intake and lung cancer risk among female nonsmokers: A report from the Shanghai Women’s Health Study. Cancer Epidemiol. Biomark. Prev. 2013, 22, 50–57. [Google Scholar] [CrossRef]
- Loomis, D.; Grosse, Y.; Lauby-Secretan, B.; El Ghissassi, F.; Bouvard, V.; Benbrahim-Tallaa, L.; Guha, N.; Baan, R.; Mattock, H.; Straif, K.; et al. The carcinogenicity of outdoor air pollution. Lancet Oncol. 2013, 14, 1262–1263. [Google Scholar] [CrossRef]
- Hill, W.; Lim, E.L.; Weeden, C.E.; Lee, C.; Augustine, M.; Chen, K.; Kuan, F.C.; Marongiu, F.; Evans, E.J., Jr.; Moore, D.A.; et al. Lung adenocarcinoma promotion by air pollutants. Nature 2023, 616, 159–167. [Google Scholar] [CrossRef]
- Hogg, J.C.; van Eeden, S. Pulmonary and systemic response to atmospheric pollution. Respirology 2009, 14, 336–346. [Google Scholar] [CrossRef]
- Valavanidis, A.; Vlachogianni, T.; Fiotakis, K.; Loridas, S. Pulmonary oxidative stress, inflammation and cancer: Respirable particulate matter, fibrous dusts and ozone as major causes of lung carcinogenesis through reactive oxygen species mechanisms. Int. J. Environ. Res. Public Health 2013, 10, 3886–3907. [Google Scholar] [CrossRef]
- Straif, K.; Baan, R.; Grosse, Y.; Secretan, B.; El Ghissassi, F.; Cogliano, V.; WHO International Agency for Research on Cancer Monograph Working Group. Carcinogenicity of polycyclic aromatic hydrocarbons. Lancet Oncol. 2005, 6, 931–932. [Google Scholar] [CrossRef] [PubMed]
- Chiang, T.A.; Wu, P.F.; Ko, Y.C. Identification of carcinogens in cooking oil fumes. Environ. Res. 1999, 81, 18–22. [Google Scholar] [CrossRef]
- Moorthy, B.; Chu, C.; Carlin, D.J. Polycyclic aromatic hydrocarbons: From metabolism to lung cancer. Toxicol. Sci. 2015, 145, 5–15. [Google Scholar] [CrossRef] [PubMed]
- Palackal, N.T.; Lee, S.H.; Harvey, R.G.; Blair, I.A.; Penning, T.M. Activation of polycyclic aromatic hydrocarbon trans-dihydrodiol proximate carcinogens by human aldo-keto reductase (AKR1C) enzymes and their functional overexpression in human lung carcinoma (A549) cells. J. Biol. Chem. 2002, 277, 24799–24808. [Google Scholar] [CrossRef] [PubMed]
- IARC. Diesel and Gasoline Engine Exhausts and Some Nitroarenes. Iarc Monographs on the Evaluation of Carcinogenic Risks to Humans. IARC Monogr. Eval. Carcinog. Risks Hum. 2014, 105, 9–699. [Google Scholar]
- Lui, K.H.; Bandowe, B.A.; Tian, L.; Chan, C.S.; Cao, J.J.; Ning, Z.; Lee, S.C.; Ho, K.F. Cancer risk from polycyclic aromatic compounds in fine particulate matter generated from household coal combustion in Xuanwei, China. Chemosphere 2017, 169, 660–668. [Google Scholar] [CrossRef]
- Barone-Adesi, F.; Chapman, R.S.; Silverman, D.T.; He, X.; Hu, W.; Vermeulen, R.; Ning, B.; Fraumeni, J.F., Jr.; Rothman, N.; Lan, Q. Risk of lung cancer associated with domestic use of coal in Xuanwei, China: Retrospective cohort study. BMJ 2012, 345, e5414. [Google Scholar] [CrossRef]
- Lee, K.B.; Ang, L.; Yau, W.P.; Seow, W.J. Association between Metabolites and the Risk of Lung Cancer: A Systematic Literature Review and Meta-Analysis of Observational Studies. Metabolites 2020, 10, 362. [Google Scholar] [CrossRef]
- Zhang, T.; Hoang, P.H.; Wong, J.Y.; Yang, K.; Chen, K.; Wong, M.P.; Vermeulen, R.C.; Xuanwei Study Team; Huang, Y.; Chanock, S.J.; et al. Distinct Genomic Landscape of Lung Adenocarcinoma from Household Use of Smoky Coal. Am. J. Respir. Crit. Care Med. 2023, 208, 733–736. [Google Scholar] [CrossRef]
- Ho, K.F.; Chang, C.C.; Tian, L.; Chan, C.S.; Musa Bandowe, B.A.; Lui, K.H.; Lee, K.Y.; Chuang, K.J.; Liu, C.Y.; Ning, Z.; et al. Effects of polycyclic aromatic compounds in fine particulate matter generated from household coal combustion on response to EGFR mutations in vitro. Environ. Pollut. 2016, 218, 1262–1269. [Google Scholar] [CrossRef] [PubMed]
- Lan, Q.; Mumford, J.L.; Shen, M.; Demarini, D.M.; Bonner, M.R.; He, X.; Yeager, M.; Welch, R.; Chanock, S.; Tian, L.; et al. Oxidative damage-related genes AKR1C3 and OGG1 modulate risks for lung cancer due to exposure to PAH-rich coal combustion emissions. Carcinogenesis 2004, 25, 2177–2181. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.J.; Ha, S.; Lee, H.Y.; Lee, K.J. ROSics: Chemistry and proteomics of cysteine modifications in redox biology. Mass Spectrom. Rev. 2015, 34, 184–208. [Google Scholar] [CrossRef]
- Key, T.J.; Allen, N.E.; Spencer, E.A.; Travis, R.C. The effect of diet on risk of cancer. Lancet 2002, 360, 861–868. [Google Scholar] [CrossRef] [PubMed]
- Wei, X.; Zhu, C.; Ji, M.; Fan, J.; Xie, J.; Huang, Y.; Jiang, X.; Xu, J.; Yin, R.; Du, L.; et al. Diet and Risk of Incident Lung Cancer: A Large Prospective Cohort Study in UK Biobank. Am. J. Clin. Nutr. 2021, 114, 2043–2051. [Google Scholar] [CrossRef]
- Yu, J.; Bi, X.; Yu, B.; Chen, D. Isoflavones: Anti-Inflammatory Benefit and Possible Caveats. Nutrients 2016, 8, 361. [Google Scholar] [CrossRef]
- Seow, W.J.; Shu, X.O.; Nicholson, J.K.; Holmes, E.; Walker, D.I.; Hu, W.; Cai, Q.; Gao, Y.T.; Xiang, Y.B.; Moore, S.C.; et al. Association of Untargeted Urinary Metabolomics and Lung Cancer Risk Among Never-Smoking Women in China. JAMA Netw. Open 2019, 2, e1911970. [Google Scholar] [CrossRef]
- Ducker, G.S.; Rabinowitz, J.D. One-Carbon Metabolism in Health and Disease. Cell Metab. 2017, 25, 27–42. [Google Scholar] [CrossRef]
- Baltar, V.T.; Xun, W.W.; Johansson, M.; Ferrari, P.; Chuang, S.C.; Relton, C.; Ueland, P.M.; Midttun, O.; Slimani, N.; Jenab, M.; et al. A structural equation modelling approach to explore the role of B vitamins and immune markers in lung cancer risk. Eur. J. Epidemiol. 2013, 28, 677–688. [Google Scholar] [CrossRef]
- Yang, J.; Li, H.; Deng, H.; Wang, Z. Association of One-Carbon Metabolism-Related Vitamins (Folate, B6, B12), Homocysteine and Methionine With the Risk of Lung Cancer: Systematic Review and Meta-Analysis. Front. Oncol. 2018, 8, 493. [Google Scholar] [CrossRef]
- Aldini, G.; Vistoli, G.; Regazzoni, L.; Gamberoni, L.; Facino, R.M.; Yamaguchi, S.; Uchida, K.; Carini, M. Albumin is the main nucleophilic target of human plasma: A protective role against pro-atherogenic electrophilic reactive carbonyl species? Chem. Res. Toxicol. 2008, 21, 824–835. [Google Scholar] [CrossRef] [PubMed]
- Dalle-Donne, I.; Rossi, R.; Colombo, R.; Giustarini, D.; Milzani, A. Biomarkers of oxidative damage in human disease. Clin. Chem. 2006, 52, 601–623. [Google Scholar] [CrossRef]
- Watanabe, H.; Imafuku, T.; Otagiri, M.; Maruyama, T. Clinical Implications Associated with the Posttranslational Modification-Induced Functional Impairment of Albumin in Oxidative Stress-Related Diseases. J. Pharm. Sci. 2017, 106, 2195–2203. [Google Scholar] [CrossRef]
- Grigoryan, H.; Imani, P.; Dudoit, S.; Rappaport, S.M. Extending the HSA-Cys34-Adductomics Pipeline to Modifications at Lys525. Chem. Res. Toxicol. 2021, 34, 2549–2557. [Google Scholar] [CrossRef] [PubMed]
- Zheng, W.; Chow, W.H.; Yang, G.; Jin, F.; Rothman, N.; Blair, A.; Li, H.L.; Wen, W.; Ji, B.T.; Li, Q.; et al. The Shanghai Women’s Health Study: Rationale, study design, and baseline characteristics. Am. J. Epidemiol. 2005, 162, 1123–1131. [Google Scholar] [CrossRef] [PubMed]
- Yang, Z.; Lin, Y.; Wang, S.; Liu, X.; Cullinan, P.; Chung, K.F.; Zhang, J. Urinary Amino-Polycyclic Aromatic Hydrocarbons in Urban Residents: Finding a Biomarker for Residential Exposure to Diesel Traffic. Environ. Sci. Technol. 2021, 55, 10569–10577. [Google Scholar] [CrossRef]
- Grigoryan, H.; Imani, P.; Sacerdote, C.; Masala, G.; Grioni, S.; Tumino, R.; Chiodini, P.; Dudoit, S.; Vineis, P.; Rappaport, S.M. HSA Adductomics Reveals Sex Differences in NHL Incidence and Possible Involvement of Microbial Translocation. Cancer Epidemiol. Biomark. Prev. 2023, 32, 1217–1226. [Google Scholar] [CrossRef]
- Zhu, H.; Zhang, S. Body mass index and lung cancer risk in never smokers: A meta-analysis. BMC Cancer 2018, 18, 635. [Google Scholar] [CrossRef]
- Grigoryan, H.; Edmands, W.; Lu, S.S.; Yano, Y.; Regazzoni, L.; Iavarone, A.T.; Williams, E.R.; Rappaport, S.M. Adductomics Pipeline for Untargeted Analysis of Modifications to Cys34 of Human Serum Albumin. Anal. Chem. 2016, 88, 10504–10512. [Google Scholar] [CrossRef]
- Lu, S.S.; Grigoryan, H.; Edmands, W.M.; Hu, W.; Iavarone, A.T.; Hubbard, A.; Rothman, N.; Vermeulen, R.; Lan, Q.; Rappaport, S.M. Profiling the Serum Albumin Cys34 Adductome of Solid Fuel Users in Xuanwei and Fuyuan, China. Environ. Sci. Technol. 2017, 51, 46–57. [Google Scholar] [CrossRef]
- Grigoryan, H.; Edmands, W.M.B.; Lan, Q.; Carlsson, H.; Vermeulen, R.; Zhang, L.; Yin, S.N.; Li, G.L.; Smith, M.T.; Rothman, N.; et al. Adductomic signatures of benzene exposure provide insights into cancer induction. Carcinogenesis 2018, 39, 661–668. [Google Scholar] [CrossRef]
- Liu, S.; Grigoryan, H.; Edmands, W.M.B.; Dagnino, S.; Sinharay, R.; Cullinan, P.; Collins, P.; Chung, K.F.; Barratt, B.; Kelly, F.J.; et al. Cys34 Adductomes Differ between Patients with Chronic Lung or Heart Disease and Healthy Controls in Central London. Environ. Sci. Technol. 2018, 52, 2307–2313. [Google Scholar] [CrossRef] [PubMed]
- Dagnino, S.; Bodinier, B.; Grigoryan, H.; Rappaport, S.M.; Karimi, M.; Guida, F.; Polidoro, S.; Edmands, W.B.; Naccarati, A.; Fiorito, G.; et al. Agnostic Cys34-albumin adductomics and DNA methylation: Implication of N-acetylcysteine in lung carcinogenesis years before diagnosis. Int. J. Cancer 2019, 146, 3294–3303. [Google Scholar] [CrossRef] [PubMed]
- Grigoryan, H.; Schiffman, C.; Gunter, M.J.; Naccarati, A.; Polidoro, S.; Dagnino, S.; Dudoit, S.; Vineis, P.; Rappaport, S.M. Cys34 Adductomics Links Colorectal Cancer with the Gut Microbiota and Redox Biology. Cancer Res. 2019, 79, 6024–6031. [Google Scholar] [CrossRef]
- Wong, J.Y.Y.; Imani, P.; Grigoryan, H.; Bassig, B.A.; Dai, Y.; Hu, W.; Blechter, B.; Rahman, M.L.; Ji, B.T.; Duan, H.; et al. Exposure to diesel engine exhaust and alterations to the Cys34/Lys525 adductome of human serum albumin. Environ. Toxicol. Pharmacol. 2022, 95, 103966. [Google Scholar] [CrossRef]
- Yuan, K.; Liu, Y.; Chen, H.N.; Zhang, L.; Lan, J.; Gao, W.; Dou, Q.; Nice, E.C.; Huang, C. Thiol-based redox proteomics in cancer research. Proteomics 2015, 15, 287–299. [Google Scholar] [CrossRef] [PubMed]
- Kobayashi, Y.; Suzuki, R.; Yasukawa, K.; Oba, K.; Yamauchi, T.; Yatomi, Y.; Kadowaki, T. Oxidized albumin in blood reflects the severity of multiple vascular complications in diabetes mellitus. Metabol. Open 2020, 6, 100032. [Google Scholar] [CrossRef]
- Paramasivan, S.; Adav, S.S.; Ngan, S.C.; Dalan, R.; Leow, M.K.; Ho, H.H.; Sze, S.K. Serum albumin cysteine trioxidation is a potential oxidative stress biomarker of type 2 diabetes mellitus. Sci. Rep. 2020, 10, 6475. [Google Scholar] [CrossRef]
- Zhu, H.; Blake, S.; Chan, K.T.; Pearson, R.B.; Kang, J. Cystathionine beta-Synthase in Physiology and Cancer. Biomed. Res. Int. 2018, 2018, 3205125. [Google Scholar] [CrossRef]
- Zhang, J.J.; Zheng, Y.; Vermeulen, R.; Liu, X.L.; Dai, Y.; Hu, W.; He, L.; Lin, Y.; Ren, D.; Duan, H.; et al. Urinary Amino-PAHs in relation to diesel engine emissions and urinary mutagenicity. Int. J. Hyg. Environ. Health 2023, 253, 114223. [Google Scholar] [CrossRef]
- Campbell, K.; Vowinckel, J.; Keller, M.A.; Ralser, M. Methionine Metabolism Alters Oxidative Stress Resistance via the Pentose Phosphate Pathway. Antioxid. Redox Signal. 2016, 24, 543–547. [Google Scholar] [CrossRef] [PubMed]
- Keller, B.O.; Sui, J.; Young, A.B.; Whittal, R.M. Interferences and contaminants encountered in modern mass spectrometry. Anal. Chim. Acta 2008, 627, 71–81. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.; Zhang, J.; Zhang, L.; Liu, W.; Weisel, C.P. Selective detection of monohydroxy metabolites of polycyclic aromatic hydrocarbons in urine using liquid chromatography/triple quadrupole tandem mass spectrometry. Rapid Commun. Mass Spectrom. 2004, 18, 2299–2308. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variable | HC Cases (n = 206) | LUAD Cases (n = 148) | Controls (n = 293) | p-Value (HC) 1 | p-Value (LUAD) 1 | ||
|---|---|---|---|---|---|---|---|
| BMI (kg/m2) | Min | 16.51 | 16.59 | Min | 16.45 | 0.001 * | 0.005 * |
| Mean | 23.82 | 23.82 | Mean | 24.90 | |||
| Median | 23.82 | 23.90 | Median | 24.44 | |||
| Max | 35.42 | 35.42 | Max | 36.18 | |||
| Not available | 0 | 0 | Not available | 1 | |||
| Amino-PAHs 2 (pg/mL) | Min | 555.8 | 591.6 | Min | 288.1 | 0.226 | 0.449 |
| Mean | 4783 | 4949 | Mean | 3916 | |||
| Median | 3544 | 3508 | Median | 2951 | |||
| Max | 57,121 | 57,121 | Max | 16,024 | |||
| Not available | 116 | 85 | Not available | 178 | |||
| Hydroxy-PAHs 3 (pg/mL) | Min | 95.87 | 95.87 | Min | 105.5 | 0.820 | 0.840 |
| Mean | 1040 | 1010 | Mean | 978.0 | |||
| Median | 872.0 | 854.4 | Median | 799.9 | |||
| Max | 3914 | 3914 | Max | 3578 | |||
| Not available | 126 | 95 | Not available | 189 | |||
| Education | College | 30 | 21 | College | 34 | 0.024 * | 0.003 * |
| High School | 60 | 50 | High School | 60 | |||
| Middle School | 57 | 39 | Middle School | 80 | |||
| Elementary | 59 | 38 | Elementary | 119 | |||
| Age (years) | Min | 40 | 40 | Min | 40 | 0.002 * | 0.0003 * |
| Mean | 56.2 | 55.5 | Mean | 58.5 | |||
| Median | 57 | 56.5 | Median | 62 | |||
| Max | 70 | 70 | Max | 70 | |||
| All Soy Consumption (g/d) | Min | 6.29 | 14.42 | Min | 2.28 | 0.014 * | 0.270 |
| Mean | 211.3 | 236.8 | Mean | 249.6 | |||
| Median | 89.10 | 100.3 | Median | 151.3 | |||
| Max | 1099 | 1099 | Max | 1277 | |||
| All Vegetable Consumption (g/d) | Min | 52.2 | 59.5 | Min | 23.8 | 0.487 | 0.075 |
| Mean | 308.7 | 320.7 | Mean | 298.1 | |||
| Median | 265.0 | 294.7 | Median | 258.7 | |||
| Max | 1050 | 983.6 | Max | 878.6 | |||
| Methionine (g/d) | Min | 0.414 | 0.414 | Min | 0.630 | 0.781 | 0.659 |
| Mean | 1.501 | 1.522 | Mean | 1.505 | |||
| Median | 1.393 | 1.440 | Median | 1.454 | |||
| Max | 3.986 | 3.826 | Max | 3.167 | |||
| Folate (μg/d) | Min | 45.3 | 45.3 | Min | 68.4 | 0.675 | 0.182 |
| Mean | 263.6 | 269.8 | Mean | 264.3 | |||
| Median | 250.0 | 262.8 | Median | 244.0 | |||
| Max | 692.2 | 658.3 | Max | 1155 | |||
| ETS (Ever Exposed) | No | 44 | 34 | No | 75 | 0.522 | 0.624 |
| Yes | 141 | 133 | Yes | 204 | |||
| Not available | 21 | 15 | Not available | 14 | |||
| Time to Diagnosis (years) | Min | 0.183 | 0.183 | ||||
| Mean | 7.533 | 7.568 | |||||
| Median | 8.355 | 8.475 | |||||
| Max | 13.503 | 13.336 | |||||
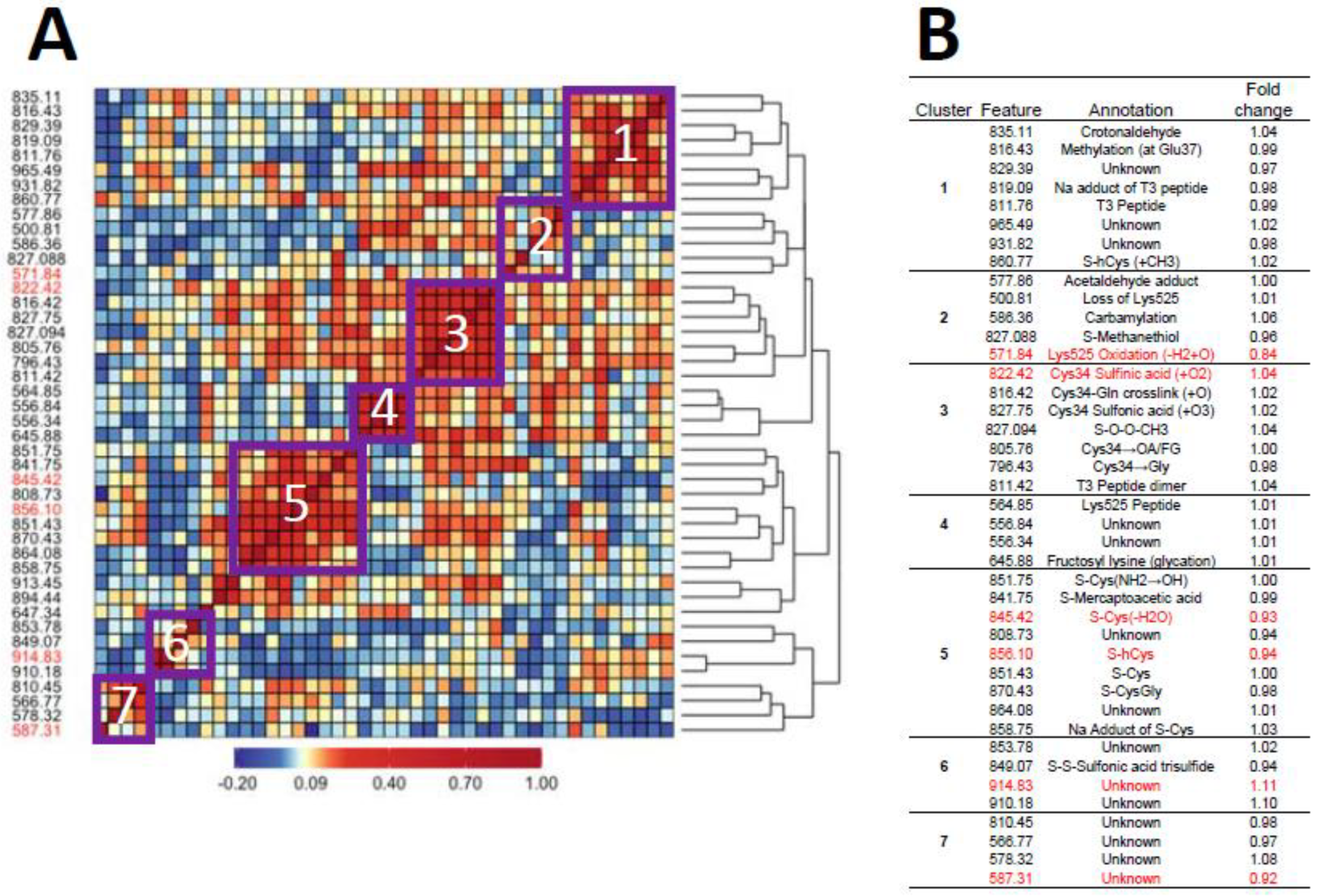
| Feature | RT, min | MIM Observed (m/z, +3) | MIM Theoret. (m/z, +3) | ΔMass (ppm) | Added Mass (+H, Da) | Elemental Composition | Annotation | Mean Peak Area (×106) |
|---|---|---|---|---|---|---|---|---|
| 796.43 a,b,c,f,g,h | 27.717 | 796.4298 | 796.4301 | −0.3767 | −45.983 | -CH2S | Cys34→Gly | 6.16 |
| 805.76 a,b,c,f,h | 27.365 | 805.7614 | 805.7618 | −0.4964 | −17.989 | -SH2, +O | Cys34→oxoalanine or fGly | 0.182 |
| 808.73 a,b,c,d,e,f,g,h | 27.868 | 808.7281 | −9.088 | Not Cys34 adduct | 42.9 | |||
| 810.45 a,d,f | 28.263 | 810.4521 | −3.916 | Not Cys34 adduct | 0.168 | |||
| 811.42 a,b,c,d,e,f,g,h | 30.803 | 811.4242 | 811.4234 | 0.9859 | −1.000 | T3 dimer * | 3.45 | |
| 811.76 a,b,c,d,e,f,g,h | 28.682 | 811.7576 | 811.7594 | −2.2174 | 0.000 | Unmodified T3 * | 9.25 | |
| 816.42 a,b,c,d,e,f,g,h | 27.390 | 816.4188 | 816.4191 | −0.3675 | 13.984 | -H2, +O | Cys34-Gln crosslink * | 1.39 |
| 816.43 a,b,c,d,e,f,g,h | 29.102 | 816.4306 | 816.4312 | −0.7349 | 15.027 | +CH3 | Methylation (at Glu37) | 3.94 |
| 819.09 b,e,g | 28.514 | 819.0863 | 22.994 | Na adduct of T3 | 0.469 | |||
| 822.42 a,b,c,d,e,f,g,h | 27.207 | 822.4222 | 822.4226 | −0.4864 | 33.002 | +HO2 | Cys34 sulfinic acid * | 16.4 |
| 827.088 b,c,f,g,h | 29.566 | 827.0883 | 827.0886 | −0.3627 | 47.000 | +CH3S | S-Methanethiol * | 40.9 |
| 827.094 c,d,f,g,h | 27.790 | 827.0942 | 827.0945 | −0.3627 | 47.018 | +CH3O2 | S-(O)-O-CH3 | 1.23 |
| 827.75 a,b,c,d,e,f,g,h | 27.361 | 827.7537 | 827.7543 | −0.7249 | 48.996 | +HO3 | Cys34 Sulfonic acid * | 2.08 |
| 829.396 a,b,g,h | 28.575 | 829.3959 | 53.923 | Not Cys34 adduct | 0.321 | |||
| 835.11 a,c,e,g,f,h | 28.809 | 835.1062 | 835.1066 | −0.4790 | 71.054 | +C4H7O | Crotonaldehyde * | 1.58 |
| 841.75 a,b,c,d,e,f,g,h | 28.411 | 841.7513 | 841.7519 | −0.7128 | 90.989 | +C2H3O2S | S-Mercaptoacetic acid | 0.061 |
| 842.07 | 28.121 | 842.0734 | 91.955 | Unknown | NA | |||
| 845.42 a,b,c,d,e,f,g,h | 27.383 | 845.4238 | 845.4239 | −0.1183 | 102.006 | +C3H4NOS | S-Cys (-H2O) | 0.487 |
| 849.07 a,b,c,h | 28.440 | 849.0684 | 849.0689 | −0.5889 | 112.940 | +HO3S2 | S-Sulfonic acid trisulfide | 0.880 |
| 851.43 a,b,c,d,e,f,g,h | 26.980 | 851.4266 | 851.4274 | −0.9396 | 120.015 | +C3H6NO2S | S-Cys * | 67.2 |
| 851.76 a,b,c,d,e,f,g | 27.632 | 851.7554 | 851.7554 | 0.0000 | 121.000 | +C3H5O3S | S-Cys (NH2→OH) | 1.23 |
| 853.78 a,b,f | 27.503 | 853.7823 | 127.082 | Unknown | 9.20 | |||
| 856.10 a,b,c,d,e,f,g,h | 26.746 | 856.0988 | 856.0993 | −0.5840 | 134.032 | +C4H8NO2S | S-hCys * | 200 |
| 858.75 a,b,c,d,e,g,h | 26.391 | 858.7546 | 141.999 | +C3H5NO2SNa | Na adduct of S-Cys | 1.52 | ||
| 860.77 b,d,e,f,g | 26.866 | 860.7703 | 860.7712 | −1.0456 | 148.046 | +C5H10NO2S | S-hCys (+CH3) | 5.10 |
| 864.08 a,b,c,e,f,g | 26.305 | 864.0763 | 157.964 | Not Cys34 adduct | 0.760 | |||
| 870.43 a,b,c,d,e,f,g,h | 26.031 | 870.4342 | 870.4345 | −0.3447 | 177.038 | +C5H9N2O3S | S-CysGly * | 0.751 |
| 894.44 a,b,c,d,e,f,g,h | 26.684 | 894.4414 | 894.4416 | −0.2236 | 249.059 | +C8H13N2O5S | S-γ-GluCys * | 0.092 |
| 910.18 g,h | 32.954 | 910.1767 | 296.265 | +C18H34NO2 | Unknown | 0.009 | ||
| 913.45 a,b,c,d,e,f,g,h | 26.554 | 913.4487 | 913.4487 | 0.0000 | 306.081 | +C10H16N3O6S | S-Glutathione * | 0.250 |
| 914.84 h | 32.931 | 914.8359 | 310.243 | Unknown | 0.552 | |||
| 931.82 b,c,d,f,g | 24.921 | 931.8204 | 361.196 | Unknown | 0.685 | |||
| 965.49 b,c,d,e,f,h | 25.207 | 965.4910 | 462.208 | Unknown | 2.24 | |||
| 970.16 d,f,h | 25.473 | 970.1631 | 476.224 | Unknown | NA |
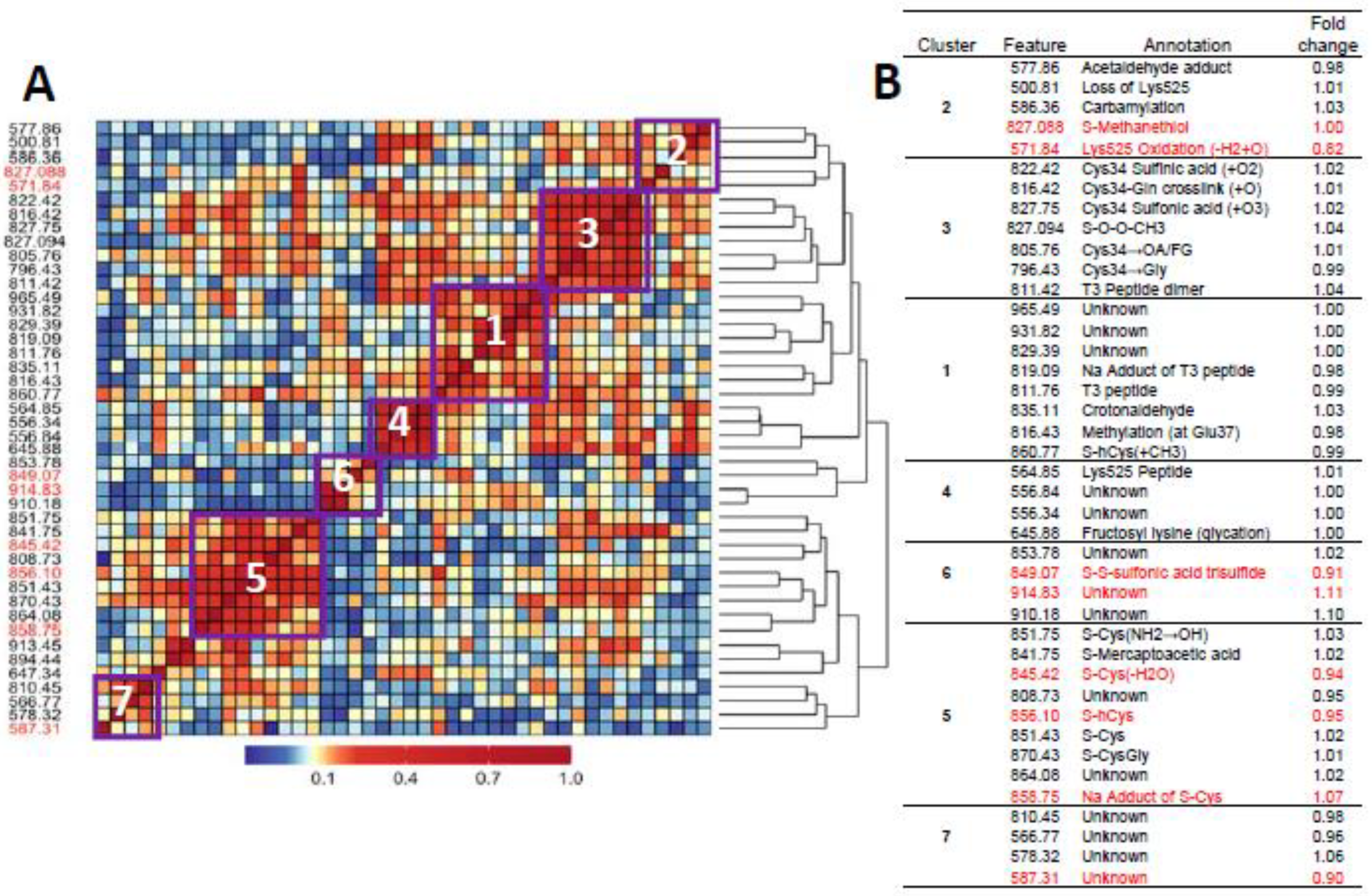
| Feature | RT, min | MIM Observed (m/z, +3) | MIM Theoretical (m/z, +3) | ΔMass (ppm) | Added Mass (+H, Da) | Elemental Composition (+H) | Annotation | Mean Peak Area (×108) |
|---|---|---|---|---|---|---|---|---|
| 500.81 a,b,c | 12.063 | 500.8045 | 500.8055 | −1.9968 | −128.095 | -C6H12N2O | Loss of Lysine | 156 |
| 556.34 a,b,c | 10.701 | 556.3396 | 556.3397 | −0.1797 | −17.024 | -NH3 | Loss of ammonia | 0.037 |
| 556.84 | 10.995 | 556.8430 | 556.8436 | −1.0775 | −16.017 | -NH2 | Deamination | 0.309 |
| 564.85 a,b,c | 10.822 | 564.8518 | 564.8529 | −1.8474 | 0.000 | Lys525 containing peptide | 7.96 | |
| 566.77 a,c | 10.684 | 566.7741 | 4.853 | Unknown | 0.014 | |||
| 571.84 c | 11.074 | 571.8423 | 571.8426 | −0.5246 | 14.989 | -H2, +O | Lys525 oxidation product | 3.87 |
| 577.86 a | 12.454 | 577.8607 | 577.8608 | −0.1731 | 27.026 | +C2H3 | Acetaldehyde | 1.91 |
| 578.32 | 12.541 | 578.3164 | 27.937 | Unknown | 0.036 | |||
| 580.85 c | 11.110 | 580.8476 | 580.8479 | −0.5165 | 32.999 | +O2H | Lys525 oxidation product | NA |
| 586.36 a,b,c | 13.265 | 586.3557 | 586.3559 | −0.3411 | 44.016 | +CH2NO | Carbamylation | 9.59 |
| 587.31 c | 13.362 | 587.3109 | 45.926 | Unknown | 1.01 | |||
| 645.88 a,b,c | 10.904 | 645.8791 | 645.8794 | −0.4645 | 163.063 | +C6H11O5 | Fructosyl lysine (glycation)/Hexose | 18.4 |
| 647.34 c | 10.778 | 647.3378 | 165.980 | Unknown | 1.03 |
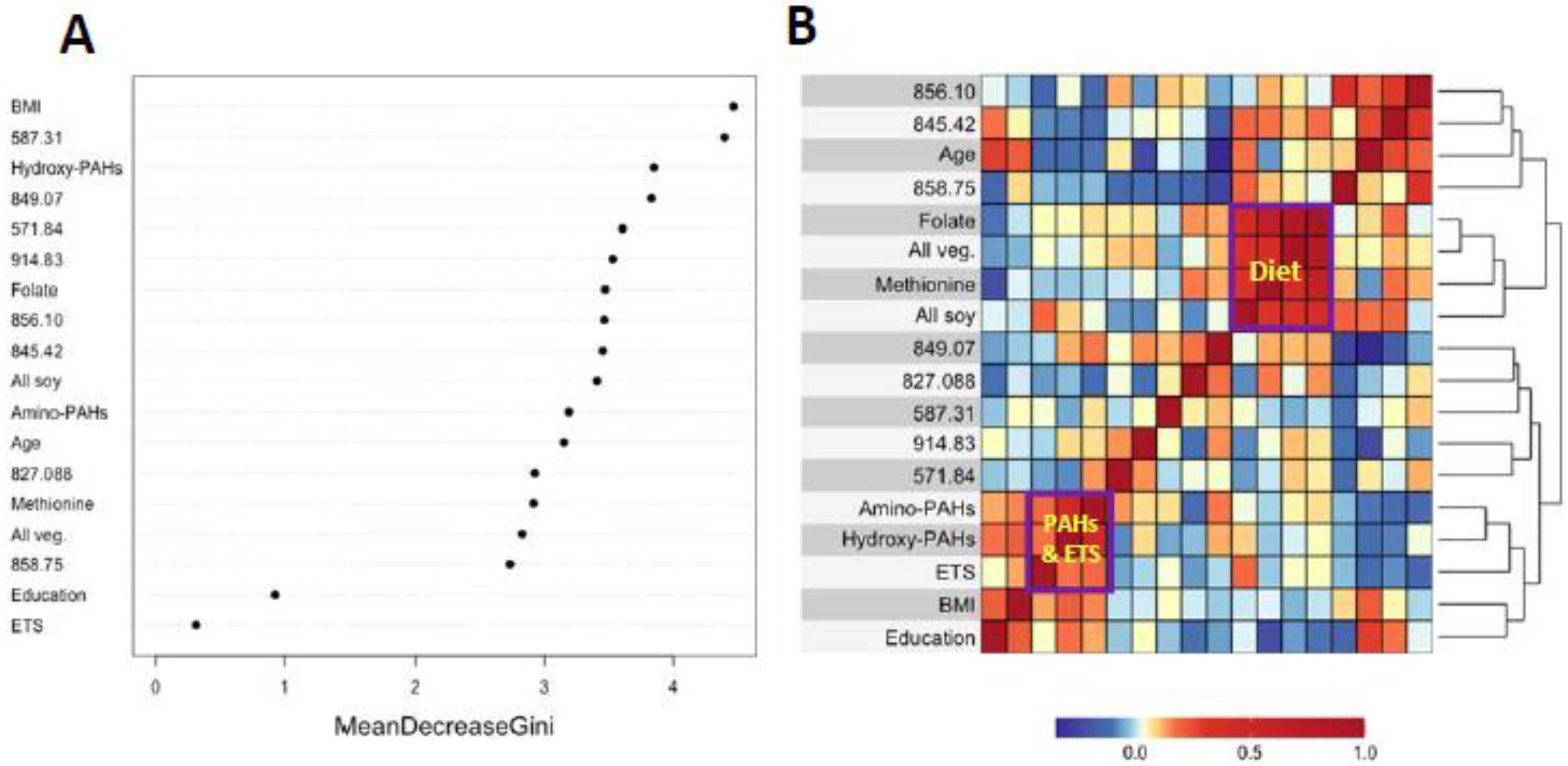
| Histologically Confirmed | LUAD | ||||||||
|---|---|---|---|---|---|---|---|---|---|
| Feature | Annotation | FC | p-Value | LASSO | RF | FC | p-Value | LASSO | RF |
| 571.84 | Lys525 oxidation (-H2+O) | 0.842 | 0.040 | Yes | Yes | 0.823 | 0.095 | Yes | Yes |
| 587.31 | Unknown | 0.921 | 0.081 | Yes | No | 0.896 | 0.040 | Yes | No |
| 822.42 | Cys34 sulfinic acid | 1.042 | 0.096 | Yes | Yes | 1.018 | 0.504 | No | No |
| 827.088 | S-Methanethiol | 0.963 | 0.453 | No | No | 0.999 | 0.922 | No | Yes |
| 845.42 | S-Cys(-H2O) | 0.932 | 0.014 | Yes | Yes | 0.937 | 0.047 | Yes | Yes |
| 849.07 | S-Sulfonic acid trisulfide | 0.935 | 0.079 | No | No | 0.912 | 0.028 | Yes | No |
| 856.10 | S-hCys | 0.942 | 0.044 | Yes | Yes | 0.946 | 0.087 | Yes | Yes |
| 858.75 | Na adduct of S-Cys | 1.029 | 0.400 | No | No | 1.070 | 0.087 | Yes | No |
| 914.83 | Unknown | 1.114 | 0.047 | Yes | Yes | 1.113 | 0.073 | Yes | Yes |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Imani, P.; Grigoryan, H.; Dudoit, S.; Shu, X.-O.; Wong, J.; Zhang, L.; Zhang, J.; Hu, W.; Cai, Q.; Gao, Y.; et al. HSA Adductomics in the Shanghai Women’s Health Study Links Lung Cancer in Never-Smokers with Air Pollution, Redox Biology, and One-Carbon Metabolism. Antioxidants 2025, 14, 335. https://doi.org/10.3390/antiox14030335
Imani P, Grigoryan H, Dudoit S, Shu X-O, Wong J, Zhang L, Zhang J, Hu W, Cai Q, Gao Y, et al. HSA Adductomics in the Shanghai Women’s Health Study Links Lung Cancer in Never-Smokers with Air Pollution, Redox Biology, and One-Carbon Metabolism. Antioxidants. 2025; 14(3):335. https://doi.org/10.3390/antiox14030335
Chicago/Turabian StyleImani, Partow, Hasmik Grigoryan, Sandrine Dudoit, Xiao-Ou Shu, Jason Wong, Luoping Zhang, Junfeng Zhang, Wei Hu, Qiuyin Cai, Yutang Gao, and et al. 2025. "HSA Adductomics in the Shanghai Women’s Health Study Links Lung Cancer in Never-Smokers with Air Pollution, Redox Biology, and One-Carbon Metabolism" Antioxidants 14, no. 3: 335. https://doi.org/10.3390/antiox14030335
APA StyleImani, P., Grigoryan, H., Dudoit, S., Shu, X.-O., Wong, J., Zhang, L., Zhang, J., Hu, W., Cai, Q., Gao, Y., Blechter, B., Rahman, M., Zheng, W., Rothman, N., Lan, Q., & Rappaport, S. M. (2025). HSA Adductomics in the Shanghai Women’s Health Study Links Lung Cancer in Never-Smokers with Air Pollution, Redox Biology, and One-Carbon Metabolism. Antioxidants, 14(3), 335. https://doi.org/10.3390/antiox14030335