
Polygonatum sibiricum Polysaccharides Extracted with Ultrasound-Assisted Deep Eutectic Solvents Protect L6 Cells Against Oxidative Stress in a Cellular Model of Sarcopenic Obesity

 ,
,  ,
,
Abstract
1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Main Instruments and Equipment
2.3. Preparations of the PsP Solutions of Different Dosages and Positive Control Solution
2.4. Cell Culture and Treatment
2.5. Cell Viability Assay
2.6. Triglyceride Assay
2.7. Oil Red O Staining
2.8. ROS Assay
2.9. Glutathione Peroxidase Activity Assay
2.10. Superoxide Dismutase Activity Assay
2.11. Catalase Activity Assay
2.12. MDA Assay
2.13. Diameter of Myotube Cells Assay
2.14. Myosin Heavy Chain Protein Expression-Level Assay
2.15. Myoblast Differentiation Protein Expression-Level Assay
2.16. Statistical Analysis
3. Results
3.1. Cell Proliferation Activity Assay
3.2. TG Content Determination
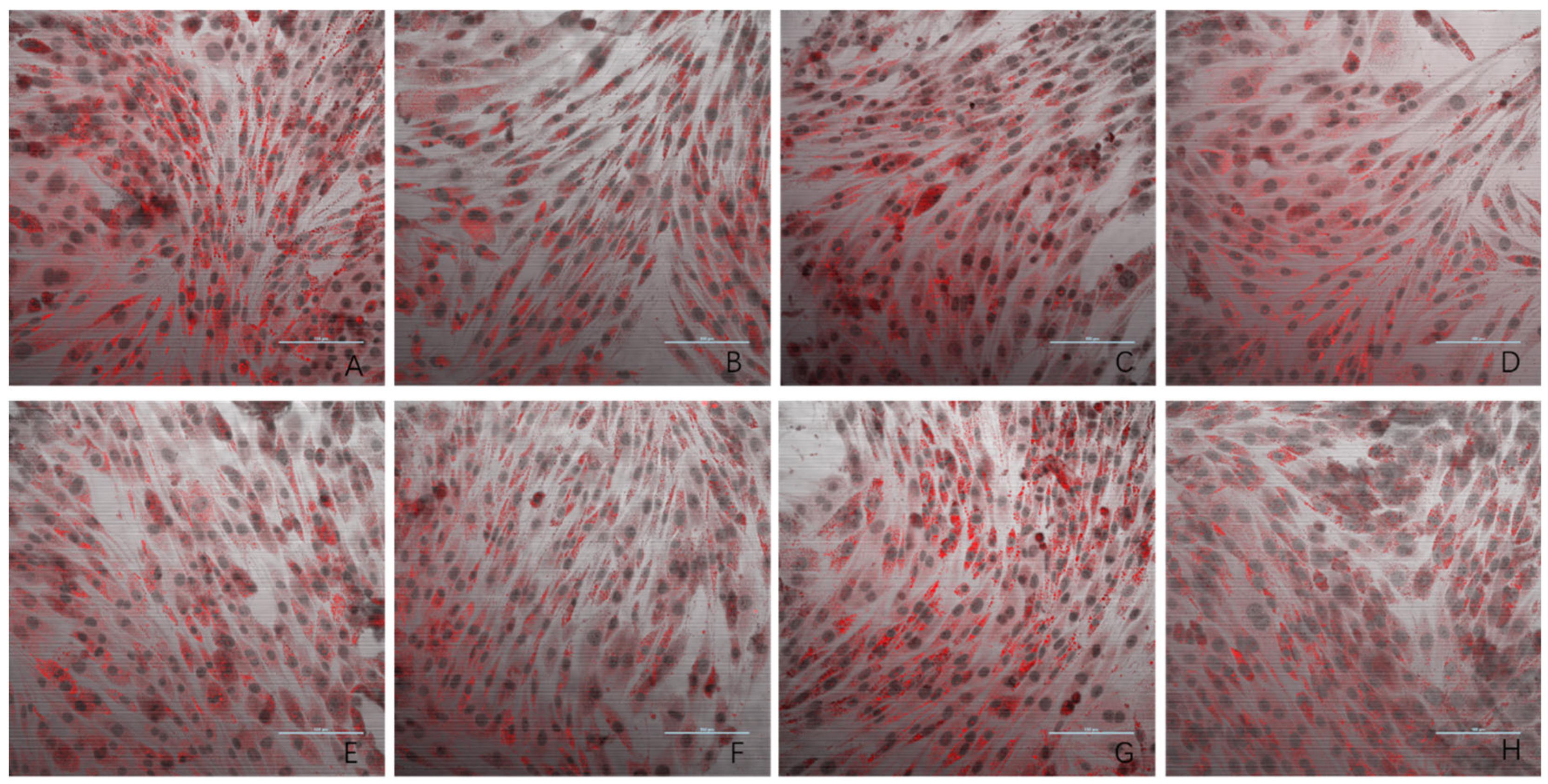
3.3. Oil Red O Staining Result
3.4. Determination of ROS Levels
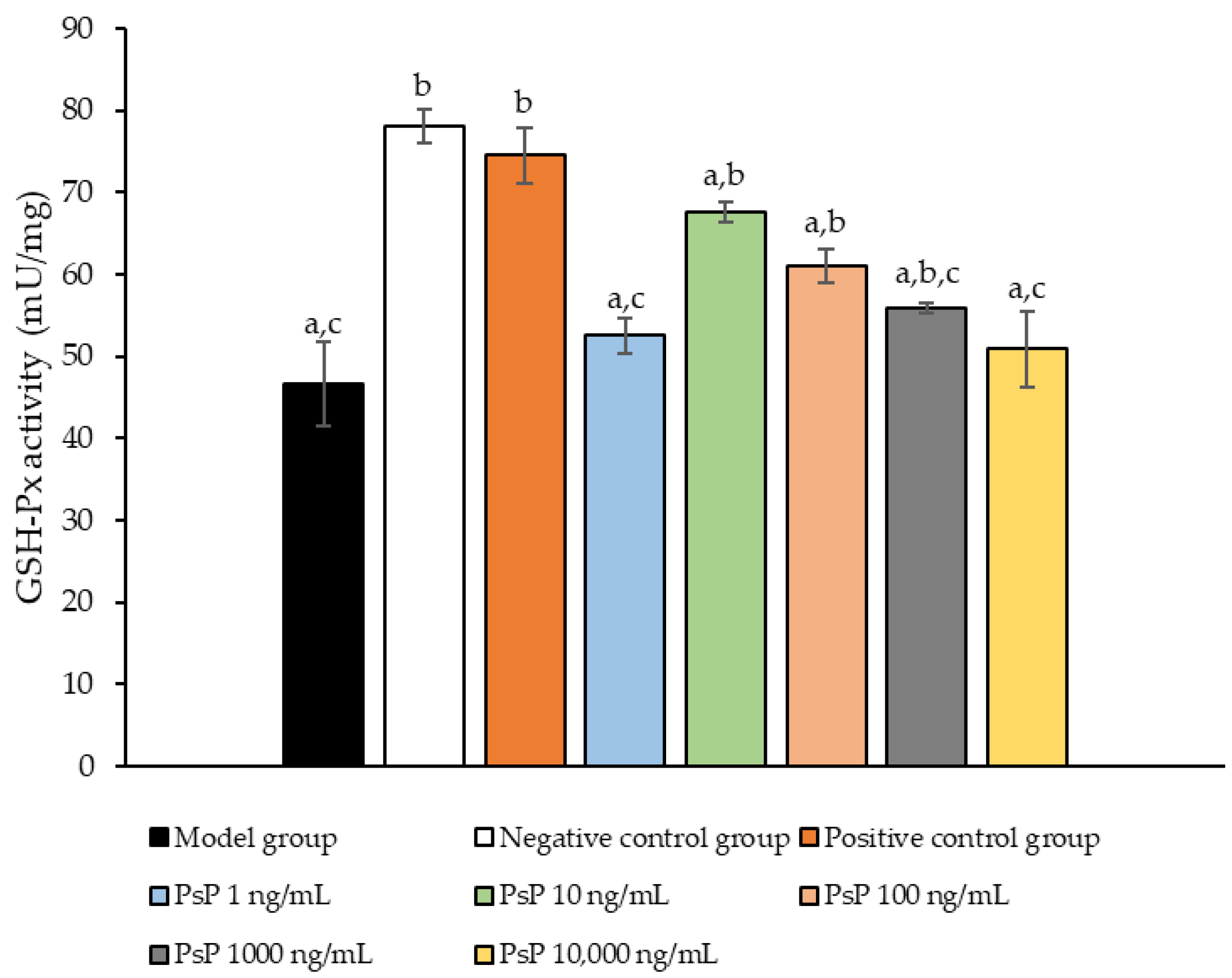
3.5. Determination of GSH-Px Activity
3.6. Determination of SOD Activity
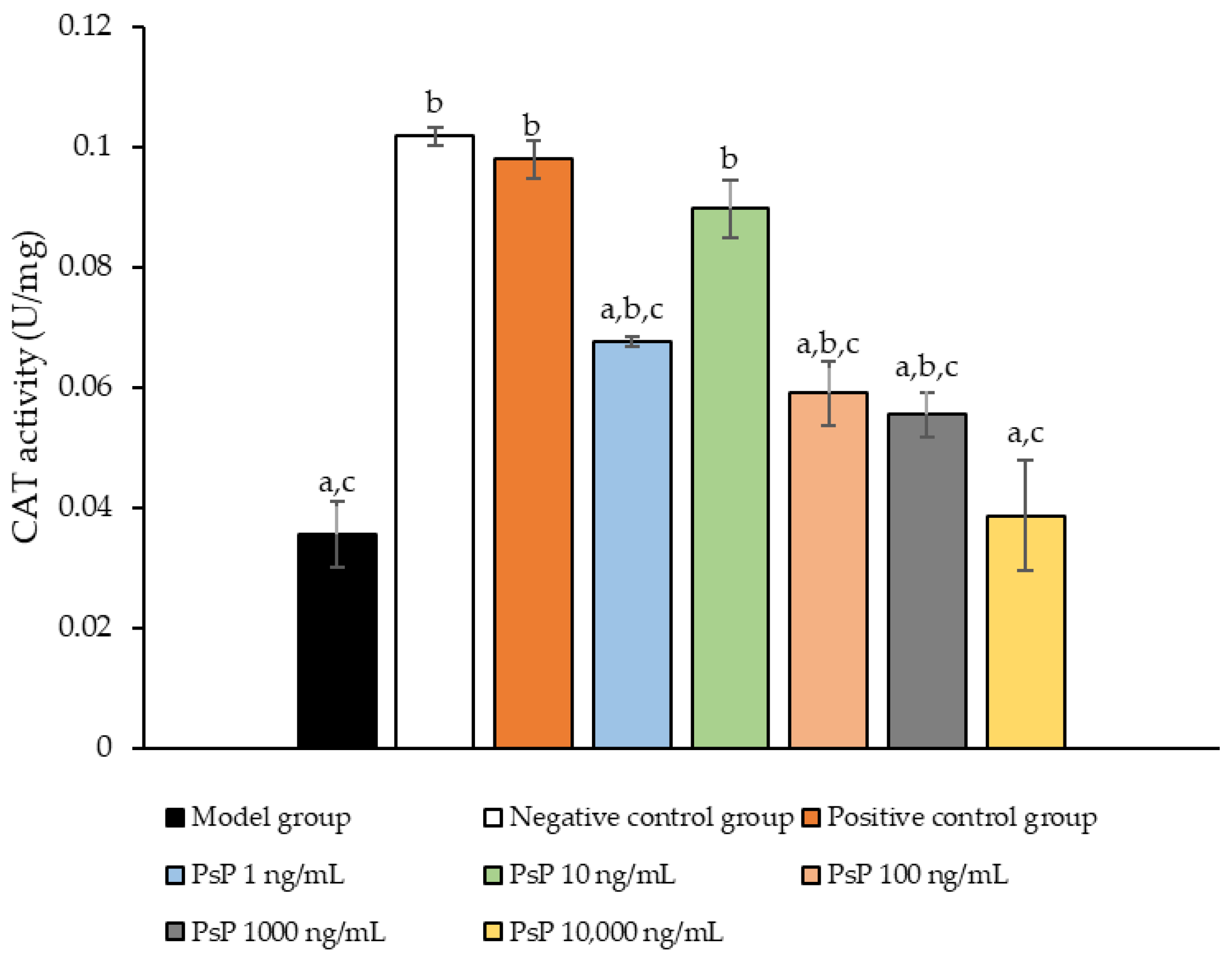
3.7. Determination of CAT Activity
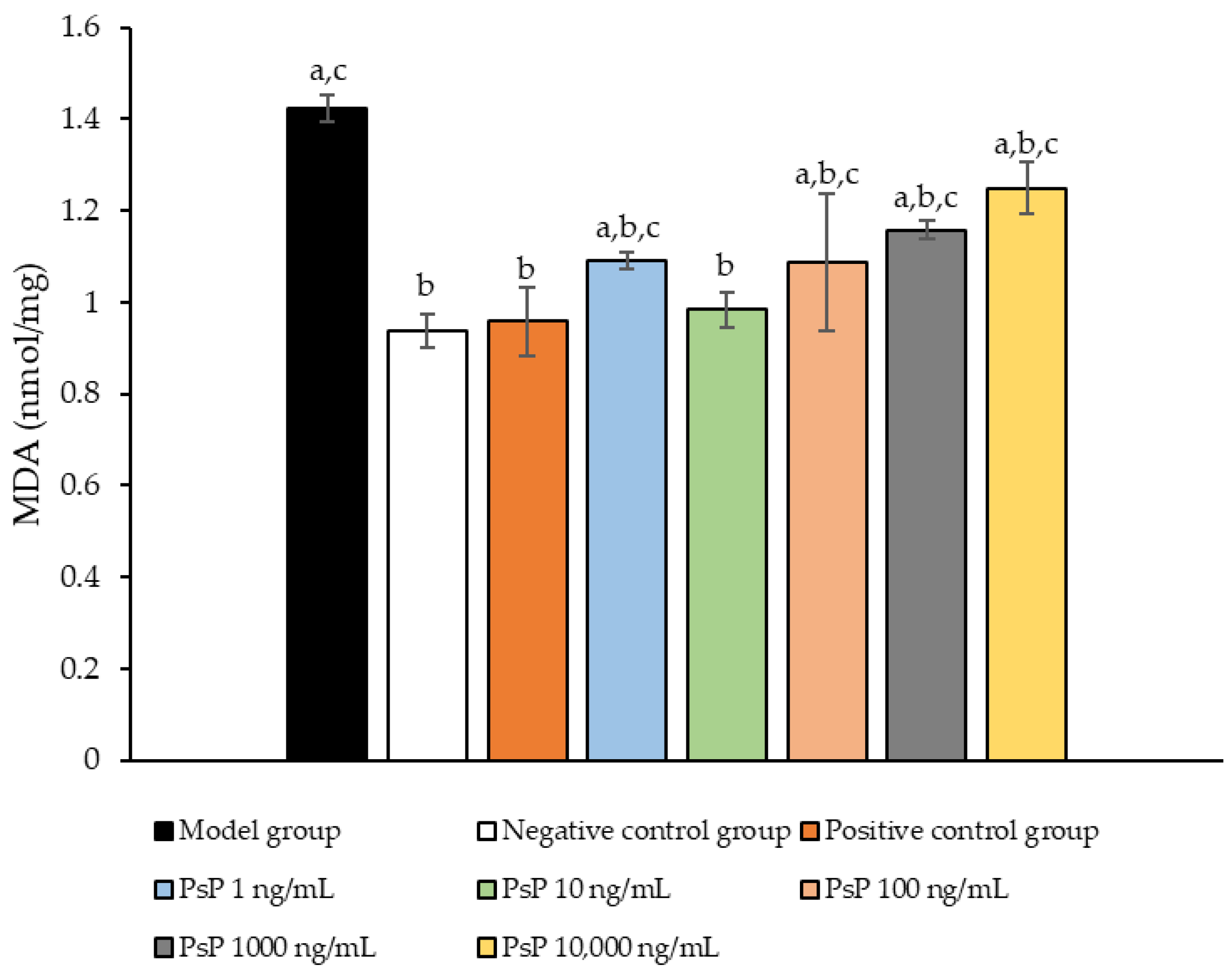
3.8. Determination of MDA Content
3.9. Measurement of Myotube Cell Diameter
3.10. Determination of MHC Protein Expression Level
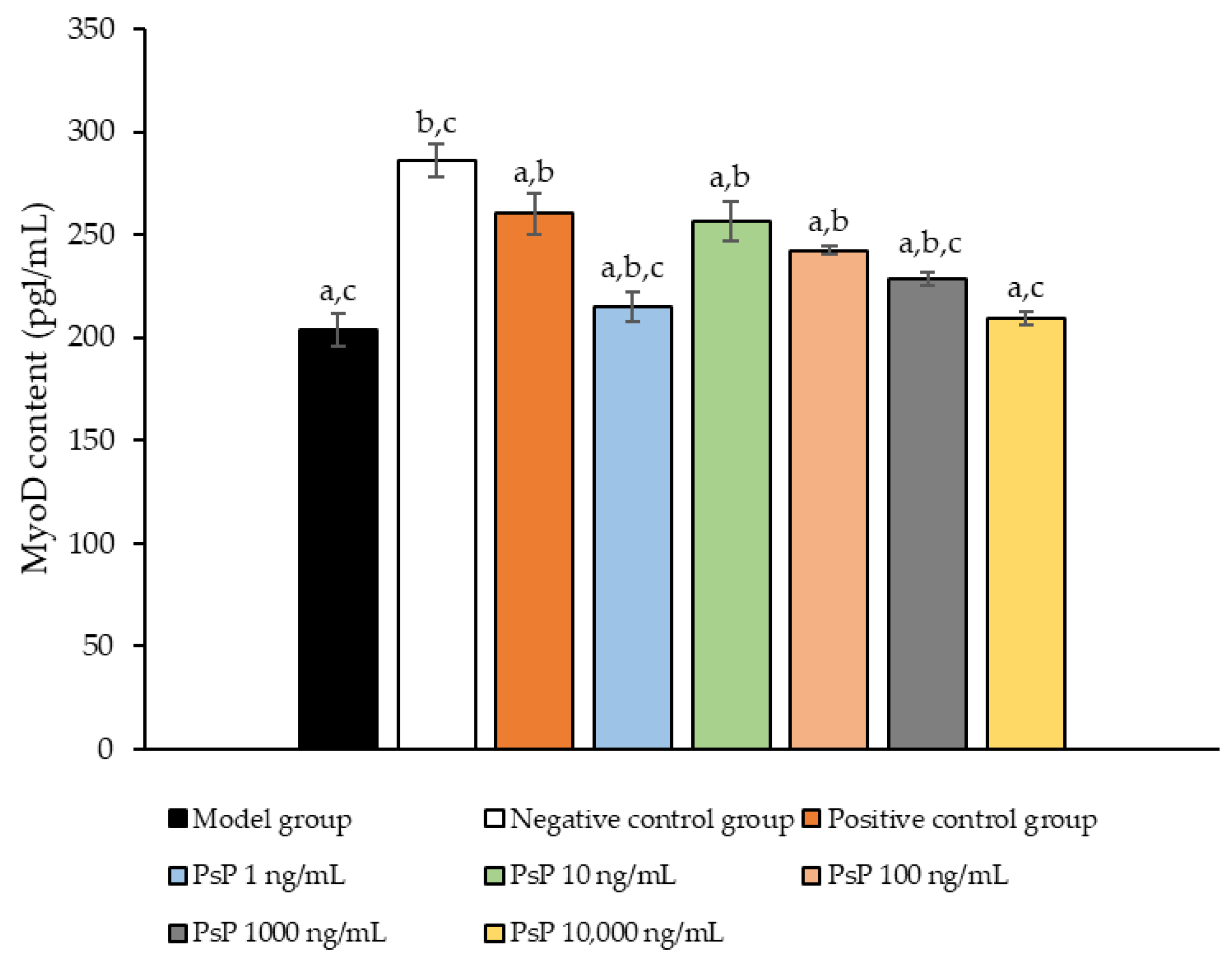
3.11. Determination of MyoD Protein Expression Level
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Sun, T.; Zhang, H.; Li, Y.; Liu, Y.; Dai, W.; Fang, J.; Cao, C.; Die, Y.; Liu, Q.; Wang, C.; et al. Physicochemical properties and immunological activities of polysaccharides from both crude and wine-processed Polygonatum sibiricum. Int. J. Biol. Macromol. 2020, 143, 255–264. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Cai, X.T.; Tian, Q.H.; Xiao, L.X.; Zeng, Z.; Cai, X.T.; Yan, J.Z.; Li, Q.Y. Microwave-Assisted Degradation of Polysaccharide from Polygonatum sibiricum and Antioxidant Activity. J. Food Sci. 2019, 84, 754–761. [Google Scholar] [CrossRef] [PubMed]
- Horng, C.T.; Huang, J.K.; Wang, H.Y.; Huang, C.C.; Chen, F.A. Antioxidant and antifatigue activities of Polygonatum alte-lobatum Hayata rhizomes in rats. Nutrients 2014, 6, 5327–5337. [Google Scholar] [CrossRef] [PubMed]
- Ma, W.; Wei, S.; Peng, W.; Sun, T.; Huang, J.; Yu, R.; Zhang, B.; Li, W. Antioxidant Effect of Polygonatum sibiricum Polysaccharides in D-Galactose-Induced Heart Aging Mice. Biomed. Res. Int. 2021, 2021, 6688855. [Google Scholar] [CrossRef]
- Luo, S.; Zhang, X.; Huang, S.; Feng, X.; Zhang, X.; Xiang, D. A monomeric polysaccharide from Polygonatum sibiricum improves cognitive functions in a model of Alzheimer’s disease by reshaping the gut microbiota. Int. J. Biol. Macromol. 2022, 213, 404–415. [Google Scholar] [CrossRef]
- Zhang, H.; Cao, Y.; Chen, L.; Wang, J.; Tian, Q.; Wang, N.; Liu, Z.; Li, J.; Wang, N.; Wang, X.; et al. A polysaccharide from Polygonatum sibiricum attenuates amyloid-β-induced neurotoxicity in PC12 cells. Carbohydr. Polym. 2015, 117, 879–886. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.; Luo, J.; Jia, M.; Chai, Y.; Bao, Y. Polygonatum sibiricum saponin Exerts Beneficial Hypoglycemic Effects in Type 2 Diabetes Mice by Improving Hepatic Insulin Resistance and Glycogen Synthesis-Related Proteins. Nutrients 2022, 14, 5222. [Google Scholar] [CrossRef]
- Wang, Y.; Qin, S.; Pen, G.; Chen, D.; Han, C.; Miao, C.; Lu, B.; Su, C.; Feng, S.; Li, W.; et al. Original Research: Potential ocular protection and dynamic observation of Polygonatum sibiricum polysaccharide against streptozocin-induced diabetic rats’ model. Exp. Biol. Med. 2017, 242, 92–101. [Google Scholar] [CrossRef]
- Ma, Y.L.; Zhang, Y.S.; Zhang, F.; Zhang, Y.Y.; Thakur, K.; Zhang, J.G.; Wei, Z.J. Methyl protodioscin from Polygonatum sibiricum inhibits cervical cancer through cell cycle arrest and apoptosis induction. Food Chem. Toxicol. 2019, 132, 110655. [Google Scholar] [CrossRef]
- Zhou, D.; Li, X.; Chang, W.; Han, Y.; Liu, B.; Chen, G.; Li, N. Antiproliferative steroidal glycosides from rhizomes of Polygonatum sibiricum. Phytochemistry 2019, 164, 172–183. [Google Scholar] [CrossRef]
- Zhao, H.; Wang, Q.L.; Hou, S.B.; Chen, G. Chemical constituents from the rhizomes of Polygonatum sibiricum Red. and anti-inflammatory activity in RAW264.7 macrophage cells. Nat. Prod. Res. 2019, 33, 2359–2362. [Google Scholar] [CrossRef] [PubMed]
- Liu, D.; Tang, W.; Han, C.; Nie, S. Advances in Polygonatum sibiricum polysaccharides: Extraction, purification, structure, biosynthesis, and bioactivity. Front. Nutr. 2022, 9, 1074671. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.; Patil, S.; Qian, A.; Zhao, C. Bioactive Compounds of Polygonatum sibiricum—Therapeutic Effect and Biological Activity. Endocr. Metab. Immune Disord. Drug Targets 2022, 22, 26–37. [Google Scholar] [CrossRef]
- Shu, G.; Xu, D.; Zhao, J.; Yin, L.; Lin, J.; Fu, H.; Tang, H.; Fang, J.; Peng, X.; Zhao, X. Protective effect of Polygonatum sibiricum polysaccharide on cyclophosphamide-induced immunosuppression in chickens. Res. Vet. Sci. 2021, 135, 96–105. [Google Scholar] [CrossRef] [PubMed]
- Cui, X.; Wang, S.; Cao, H.; Guo, H.; Li, Y.; Xu, F.; Zheng, M.; Xi, X.; Han, C. A Review: The Bioactivities and Pharmacological Applications of Polygonatum sibiricum polysaccharides. Molecules 2018, 23, 1170. [Google Scholar] [CrossRef]
- Tural, R.; Karakaya, C.; Erdem, M.; Aykol, Z.; Karabacak, R.O.; Kavutçu, M. Investigation of oxidative stress status in cumulus cells in patients with in vitro fertilization. Turk. J. Med. Sci. 2021, 51, 1969–1975. [Google Scholar] [CrossRef]
- Li, L.X.; Feng, X.; Tao, M.T.; Paulsen, B.S.; Huang, C.; Feng, B.; Liu, W.; Yin, Z.Q.; Song, X.; Zhao, X.; et al. Benefits of neutral polysaccharide from rhizomes of Polygonatum sibiricum to intestinal function of aged mice. Front. Nutr. 2022, 9, 992102. [Google Scholar] [CrossRef]
- Wang, S.; Li, G.; Zhang, X.; Wang, Y.; Qiang, Y.; Wang, B.; Zou, J.; Niu, J.; Wang, Z. Structural characterization and antioxidant activity of Polygonatum sibiricum polysaccharides. Carbohydr. Polym. 2022, 291, 119524. [Google Scholar] [CrossRef]
- Filomeni, G.; De Zio, D.; Cecconi, F. Oxidative stress and autophagy: The clash between damage and metabolic needs. Cell Death Differ. 2015, 22, 377–388. [Google Scholar] [CrossRef]
- Sies, H. Oxidative stress: A concept in redox biology and medicine. Redox Biol. 2015, 4, 180–183. [Google Scholar] [CrossRef]
- Kalinkovich, A.; Livshits, G. Sarcopenic obesity or obese sarcopenia: A cross talk between age-associated adipose tissue and skeletal muscle inflammation as a main mechanism of the pathogenesis. Ageing Res. Rev. 2017, 35, 200–221. [Google Scholar] [CrossRef] [PubMed]
- Guo, A.; Li, K.; Xiao, Q. Fibroblast growth factor 19 alleviates palmitic acid-induced mitochondrial dysfunction and oxidative stress via the AMPK/PGC-1α pathway in skeletal muscle. Biochem. Biophys. Res. Commun. 2020, 526, 1069–1076. [Google Scholar] [CrossRef] [PubMed]
- Beetham, K.S.; Howden, E.J.; Small, D.M.; Briskey, D.R.; Rossi, M.; Isbel, N.; Coombes, J.S. Oxidative stress contributes to muscle atrophy in chronic kidney disease patients. Redox Rep. 2015, 20, 126–132. [Google Scholar] [CrossRef] [PubMed]
- Ferguson, C.; Pini, N.; Du, X.; Farina, M.; Hwang, J.M.C.; Pietrangelo, T.; Cheng, X. Broadband electrical impedance as a novel characterization of oxidative stress in single L6 skeletal muscle cells. Anal. Chim. Acta 2021, 1173, 338678. [Google Scholar] [CrossRef]
- Aldahhan, R.A.; Motawei, K.H.; Al-Hariri, M.T. Lipotoxicity-related sarcopenia: A review. J. Med. Life 2022, 15, 1334–1339. [Google Scholar] [CrossRef]
- Huang, Y.; Zhu, X.; Chen, K.; Lang, H.; Zhang, Y.; Hou, P.; Ran, L.; Zhou, M.; Zheng, J.; Yi, L.; et al. Resveratrol prevents sarcopenic obesity by reversing mitochondrial dysfunction and oxidative stress via the PKA/LKB1/AMPK pathway. Aging 2019, 11, 2217–2240. [Google Scholar] [CrossRef]
- Jovanović, M.S.; Krgović, N.; Živković, J.; Stević, T.; Zdunić, G.; Bigović, D.; Šavikin, K. Ultrasound-Assisted Natural Deep Eutectic Solvents Extraction of Bilberry Anthocyanins: Optimization, Bioactivities, and Storage Stability. Plants 2022, 11, 2680. [Google Scholar] [CrossRef]
- Siddiqui, S.A.; Ali Redha, A.; Salauddin, M.; Harahap, I.A.; Rupasinghe, H.P.V. Factors Affecting the Extraction of (Poly)Phenols from Natural Resources Using Deep Eutectic Solvents Combined with Ultrasound-Assisted Extraction. Crit. Rev. Anal. Chem. 2025, 55, 139–160. [Google Scholar] [CrossRef]
- Xing, C.; Cui, W.Q.; Zhang, Y.; Zou, X.S.; Hao, J.Y.; Zheng, S.D.; Wang, T.T.; Wang, X.Z.; Wu, T.; Liu, Y.Y.; et al. Ultrasound-assisted deep eutectic solvents extraction of glabridin and isoliquiritigenin from Glycyrrhiza glabra: Optimization, extraction mechanism and in vitro bioactivities. Ultrason. Sonochem. 2022, 83, 105946. [Google Scholar] [CrossRef]
- Sun, C.; Wang, G.; Sun, J.; Yin, J.; Huang, J.; Li, Z.; Mu, D.; He, M.; Liu, T.; Cheng, J.; et al. A New Method of Extracting Polygonatum sibiricum Polysaccharide with Antioxidant Function: Ultrasound-Assisted Extraction-Deep Eutectic Solvents Method. Foods 2023, 12, 3438. [Google Scholar] [CrossRef]
- Tai, H.; Wang, Z.; Gong, H.; Han, X.; Zhou, J.; Wang, X.; Wei, X.; Ding, Y.; Huang, N.; Qin, J.; et al. Autophagy impairment with lysosomal and mitochondrial dysfunction is an important characteristic of oxidative stress-induced senescence. Autophagy 2017, 13, 99–113. [Google Scholar] [CrossRef]
- Corcoran, M.P.; Lamon-Fava, S.; Fielding, R.A. Skeletal muscle lipid deposition and insulin resistance: Effect of dietary fatty acids and exercise. Am. J. Clin. Nutr. 2007, 85, 662–677. [Google Scholar] [CrossRef]
- Huang, G.; Mei, X.; Hu, J. The Antioxidant Activities of Natural Polysaccharides. Curr. Drug Targets 2017, 18, 1296–1300. [Google Scholar] [CrossRef]
- Liu, Y.; Ye, Y.; Hu, X.; Wang, J. Structural characterization and anti-inflammatory activity of a polysaccharide from the lignified okra. Carbohydr. Polym. 2021, 265, 118081. [Google Scholar] [CrossRef]
- Cui, M.; Wang, Y.; Liu, K. Structure Analysis and Anti-Inflammatory Effect of a Polysaccharide from Hippocampus mohnikei. Chem. Biodivers. 2022, 19, e202100739. [Google Scholar] [CrossRef]
- Tumova, J.; Andel, M.; Trnka, J. Excess of free fatty acids as a cause of metabolic dysfunction in skeletal muscle. Physiol. Res. 2016, 65, 193–207. [Google Scholar] [CrossRef]
- Hong, B.S.; Li, Y.; Lai, S.; Liu, J.; Guan, H.; Ke, W.; He, X.; Li, Y. Ectopic Fat Deposition on Insulin Sensitivity: Correlation of Hepatocellular Lipid Content and M Value. J. Diabetes Res. 2016, 2016, 3684831. [Google Scholar] [CrossRef]
- Sciorati, C.; Clementi, E.; Manfredi, A.A.; Rovere-Querini, P. Fat deposition and accumulation in the damaged and inflamed skeletal muscle: Cellular and molecular players. Cell Mol. Life Sci. 2015, 72, 2135–2156. [Google Scholar] [CrossRef]
- Sin, T.K.; Tam, B.T.; Yu, A.P.; Yip, S.P.; Yung, B.Y.; Chan, L.W.; Wong, C.S.; Rudd, J.A.; Siu, P.M. Acute Treatment of Resveratrol Alleviates Doxorubicin-Induced Myotoxicity in Aged Skeletal Muscle Through SIRT1-Dependent Mechanisms. J. Gerontol. Ser. A Biol. Sci. Med. Sci. 2016, 71, 730–739. [Google Scholar] [CrossRef]
- Yan, F.; Mu, Y.; Yan, G.; Liu, J.; Shen, J.; Luo, G. Antioxidant enzyme mimics with synergism. Mini Rev. Med. Chem. 2010, 10, 342–356. [Google Scholar] [CrossRef]
- Glorieux, C.; Calderon, P.B. Catalase, a remarkable enzyme: Targeting the oldest antioxidant enzyme to find a new cancer treatment approach. Biol. Chem. 2017, 398, 1095–1108. [Google Scholar] [CrossRef]
- Sun, Y.; Liu, J.; Wan, L.; Wang, F.; Zhang, X.J.; Qi, Y.J. Improving effects of Astragalus Polysaccharides on cardiac function via Keap1/Nrf2-ARE signal pathway in adjuvant arthritis rats. Chin. Herbal. Med. 2016, 8, 143–153. [Google Scholar] [CrossRef]
- Gaweł, S.; Wardas, M.; Niedworok, E.; Wardas, P. Dialdehyd malonowy (MDA) jako wskaźnik procesów peroksydacji lipidów w organizmie [Malondialdehyde (MDA) as a lipid peroxidation marker]. Wiad. Lek. 2004, 57, 453–455. (In Polish) [Google Scholar] [PubMed]
- Powers, S.K.; Smuder, A.J.; Criswell, D.S. Mechanistic links between oxidative stress and disuse muscle atrophy. Antioxid. Redox Signal. 2011, 15, 2519–2528. [Google Scholar] [CrossRef]
- Dodd, S.L.; Gagnon, B.J.; Senf, S.M.; Hain, B.A.; Judge, A.R. Ros-mediated activation of NF-kappaB and Foxo during muscle disuse. Muscle Nerve 2010, 41, 110–113. [Google Scholar] [CrossRef]
- Choi, W.H.; Son, H.J.; Jang, Y.J.; Ahn, J.; Jung, C.H.; Ha, T.Y. Apigenin Ameliorates the Obesity-Induced Skeletal Muscle Atrophy by Attenuating Mitochondrial Dysfunction in the Muscle of Obese Mice. Mol. Nutr. Food Res. 2017, 61, 1700218. [Google Scholar] [CrossRef]
- Miwa, S.; Kashyap, S.; Chini, E.; von Zglinicki, T. Mitochondrial dysfunction in cell senescence and aging. J. Clin. Investig. 2022, 132, e158447. [Google Scholar] [CrossRef]
- Azizi, S.; Nematollahi, M.A.; Mojazi Amiri, B.; Vélez, E.J.; Salmerón, C.; Chan, S.J.; Navarro, I.; Capilla, E.; Gutiérrez, J. IGF-I and IGF-II effects on local IGF system and signaling pathways in gilthead sea bream (Sparus aurata) cultured myocytes. Gen. Comp. Endocrinol. 2016, 232, 7–16. [Google Scholar] [CrossRef]
- Zhu, Z.; Li, C.; Gu, X.; Wang, X.; Zhang, G.; Fan, M.; Zhao, Y.; Liu, X.; Zhang, X. Paeoniflorin alleviated muscle atrophy in cancer cachexia through inhibiting TLR4/NF-κB signaling and activating AKT/mTOR signaling. Toxicol. Appl. Pharmacol. 2024, 484, 116846. [Google Scholar] [CrossRef]
- Jing, Y.; Yan, M.; Zhang, H.; Liu, D.; Qiu, X.; Hu, B.; Zhang, D.; Zheng, Y.; Wu, L. Effects of Extraction Methods on the Physicochemical Properties and Biological Activities of Polysaccharides from Polygonatum sibiricum. Foods 2023, 12, 2088. [Google Scholar] [CrossRef]
- Chen, Y.; Liu, J.; Xu, Y.; Sun, C.; Qu, W.; Du, H.; He, M.; Huo, J.; Sun, J.; Huang, J.; et al. Comparison of Polygonatum sibiricum Polysaccharides Found in Young and Mature Rhizomes. Foods 2024, 13, 2010. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name of Product | Product Code | Company (City, State, Country) |
|---|---|---|
| DEME culture medium | SH30243.01 | Thermo Fisher Scientific, Inc. (Waltham, MA, USA) |
| Fetal bovine serum | 10091148 | Thermo Fisher Scientific, Inc. (Waltham, MA, USA) |
| Horse serum | 16050122 | Thermo Fisher Scientific, Inc. (Waltham, MA, USA) |
| Dual antibodies (penicillin + streptomycin) | 15140122 | Thermo Fisher Scientific, Inc. (Waltham, MA, USA) |
| Palmitic acid | P0500 | Sigma Co., Ltd. (Louis, MO, USA) |
| N-Acetyl-L-cysteine | A7250 | Sigma Co., Ltd. (Louis, MO, USA) |
| CCK-8 cell activity detection kit | C0037 | Beyotime, Inc. (Shanghai, China) |
| Reactive oxygen species detection kit | S0033S | Beyotime, Inc. (Shanghai, China) |
| Glutathione peroxidase activity detection kit | BC1195 | Beijing Solarbio Life Science, Inc. (Beijing, China) |
| Total superoxide dismutase activity detection kit | S0101S | Beyotime, Inc. (Shanghai, China) |
| Catalase activity detection kit | BC0205 | Beijing Solarbio Life Science, Inc. (Beijing, China) |
| Lipid oxidation detection kit | S0131M | Beyotime, Inc. (Shanghai, China) |
| Triglyceride determination kit | A110-1-1 | Nanjing Jiacheng Bioengineering Institute, Inc. (Nanjing, China) |
| Oil red O dyeing solution | D027-1-3 | Nanjing Jiacheng Bioengineering Institute, Inc. (Nanjing, China) |
| Rat myosin heavy chain enzyme linked immunosorbent assay kit | LV20904 | Animaluni, Inc. (Shanghai, China) |
| Rat myogenic differentiation protein enzyme-linked immunosorbent assay kit | ELK6019-96 | ELK Biotechnology Co., Ltd. (Denver, CO, USA) |
| Group | Intervention Scheme | Modeling Scheme |
|---|---|---|
| Model group | Differentiation medium | Differentiation medium containing 0.75 mMPA |
| Negative control group (Blank control group) | Differentiation medium | Differentiation medium |
| Positive control group | Differentiation medium containing 5 mM NAC | Differentiation medium containing 0.75 mM PA |
| Dose group 1 | Differentiation medium containing 1 ng/mL of PsP extracted with the UAE-DESs method | Differentiation medium containing 0.75 mM PA |
| Dose group 2 | Differentiation medium containing 10 ng/mL of PsP extracted with the UAE-DESs method | Differentiation medium containing 0.75 mM PA |
| Dose group 3 | Differentiation medium containing 100 ng/mL of PsP extracted with the UAE-DESs method | Differentiation medium containing 0.75 mM PA |
| Dose group 4 | Differentiation medium containing 1000 ng/mL of PsP extracted with the UAE-DESs method | Differentiation medium containing 0.75 mM PA |
| Dose group 5 | Differentiation medium containing 10,000 ng/mL of PsP extracted with the UAE-DESs method | Differentiation medium containing 0.75 mM PA |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sun, C.; Xu, Y.; Du, H.; Chen, Y.; Qu, W.; He, M.; Liu, Z.; Huang, J.; Huo, J.; Yin, J.; et al. Polygonatum sibiricum Polysaccharides Extracted with Ultrasound-Assisted Deep Eutectic Solvents Protect L6 Cells Against Oxidative Stress in a Cellular Model of Sarcopenic Obesity. Antioxidants 2025, 14, 315. https://doi.org/10.3390/antiox14030315
Sun C, Xu Y, Du H, Chen Y, Qu W, He M, Liu Z, Huang J, Huo J, Yin J, et al. Polygonatum sibiricum Polysaccharides Extracted with Ultrasound-Assisted Deep Eutectic Solvents Protect L6 Cells Against Oxidative Stress in a Cellular Model of Sarcopenic Obesity. Antioxidants. 2025; 14(3):315. https://doi.org/10.3390/antiox14030315
Chicago/Turabian StyleSun, Chaoqun, Yifan Xu, Hanchen Du, Yan Chen, Wenjie Qu, Menglu He, Zhengyi Liu, Jian Huang, Junsheng Huo, Jiyong Yin, and et al. 2025. "Polygonatum sibiricum Polysaccharides Extracted with Ultrasound-Assisted Deep Eutectic Solvents Protect L6 Cells Against Oxidative Stress in a Cellular Model of Sarcopenic Obesity" Antioxidants 14, no. 3: 315. https://doi.org/10.3390/antiox14030315
APA StyleSun, C., Xu, Y., Du, H., Chen, Y., Qu, W., He, M., Liu, Z., Huang, J., Huo, J., Yin, J., & Liu, J. (2025). Polygonatum sibiricum Polysaccharides Extracted with Ultrasound-Assisted Deep Eutectic Solvents Protect L6 Cells Against Oxidative Stress in a Cellular Model of Sarcopenic Obesity. Antioxidants, 14(3), 315. https://doi.org/10.3390/antiox14030315