
Apomorphine Suppresses the Progression of Steatohepatitis by Inhibiting Ferroptosis

 , , , ,
, , , , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Reagents
2.2. Animals
2.3. Cell Culture and Cell Death Assay
2.4. Histology
2.5. Quantitative Real-Time Polymerase Chain Reaction (PCR)
2.6. Measurements
2.7. Western Blotting
2.8. Immunocytochemistry
2.9. Statistical Analysis
3. Results
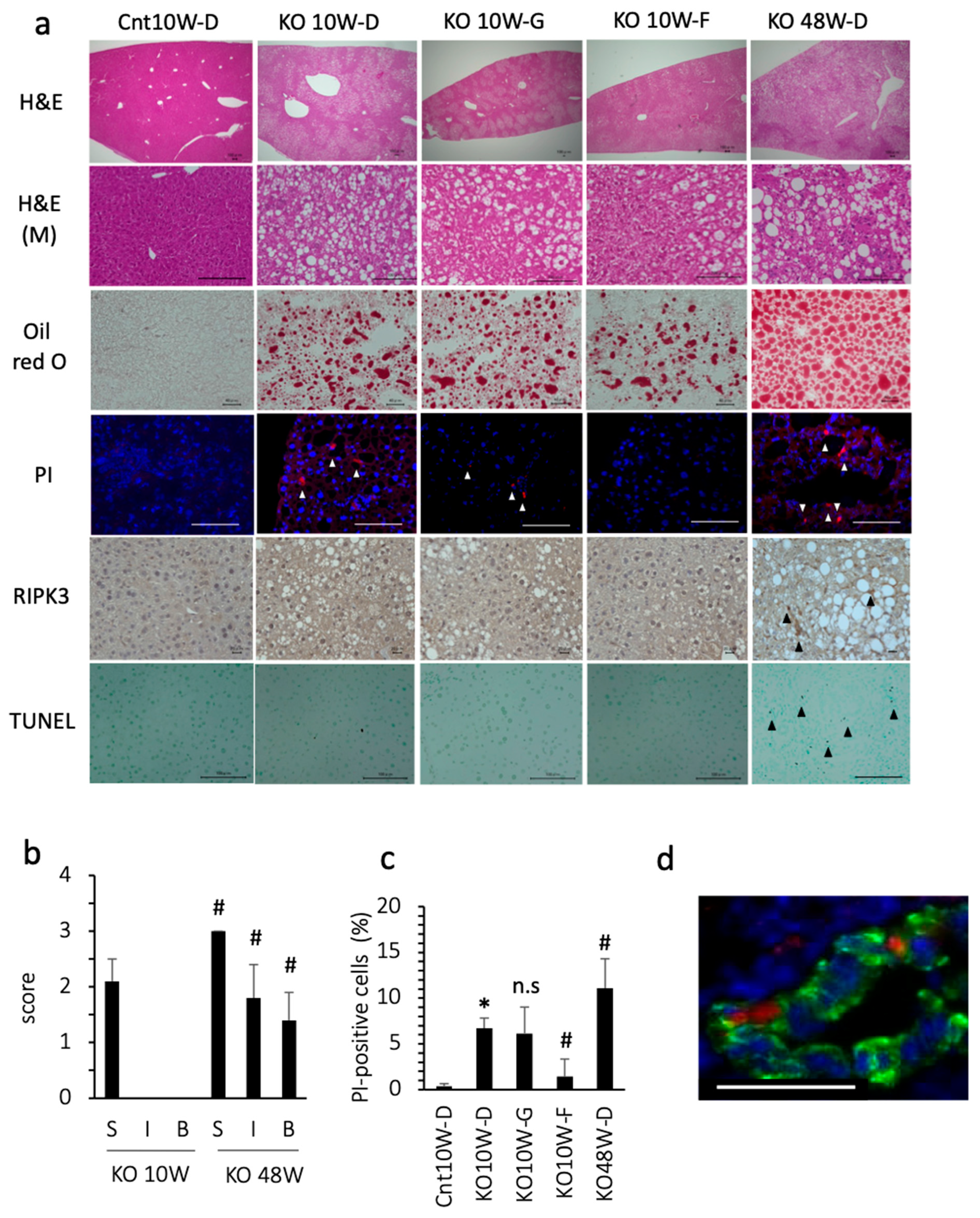
3.1. Ferroptosis Occurred in PTEN KO Mice on a Standard Diet
3.2. Ferroptosis Occurred in the Early Stage of Steatohepatitis
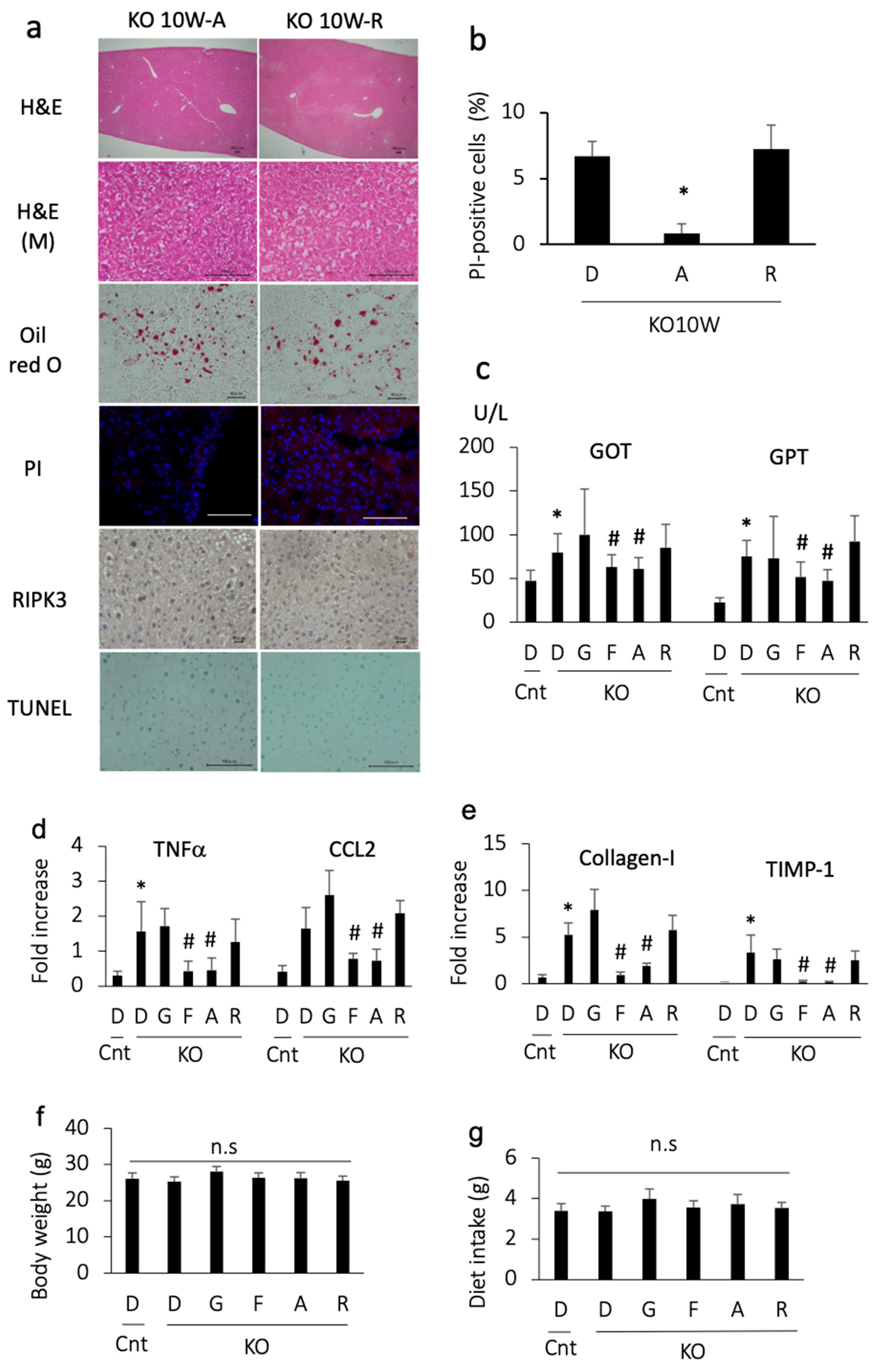
3.3. Apomorphine Ameliorated Liver Injury
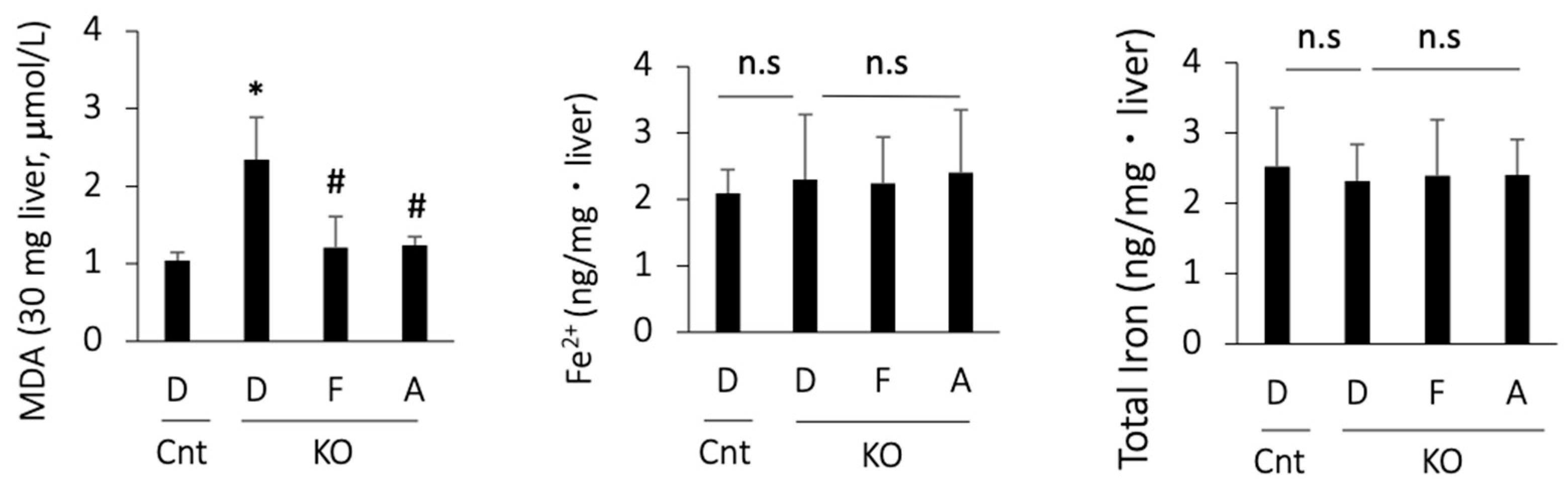
3.4. Apomorphine Reduced the Oxidative Stress Marker but Not Iron Content in the Liver
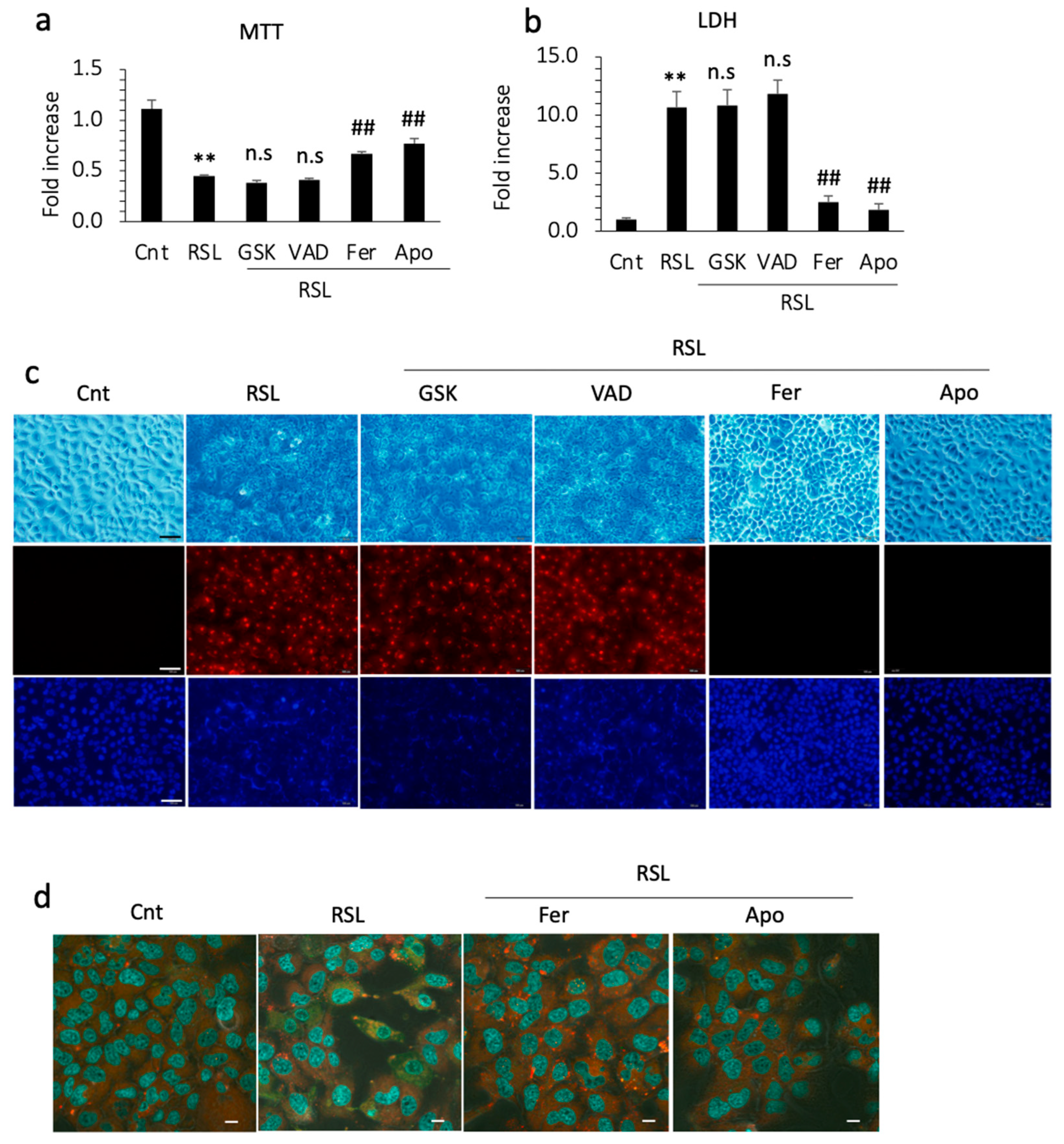
3.5. Apomorphine Suppressed Cell Death through Ferroptosis
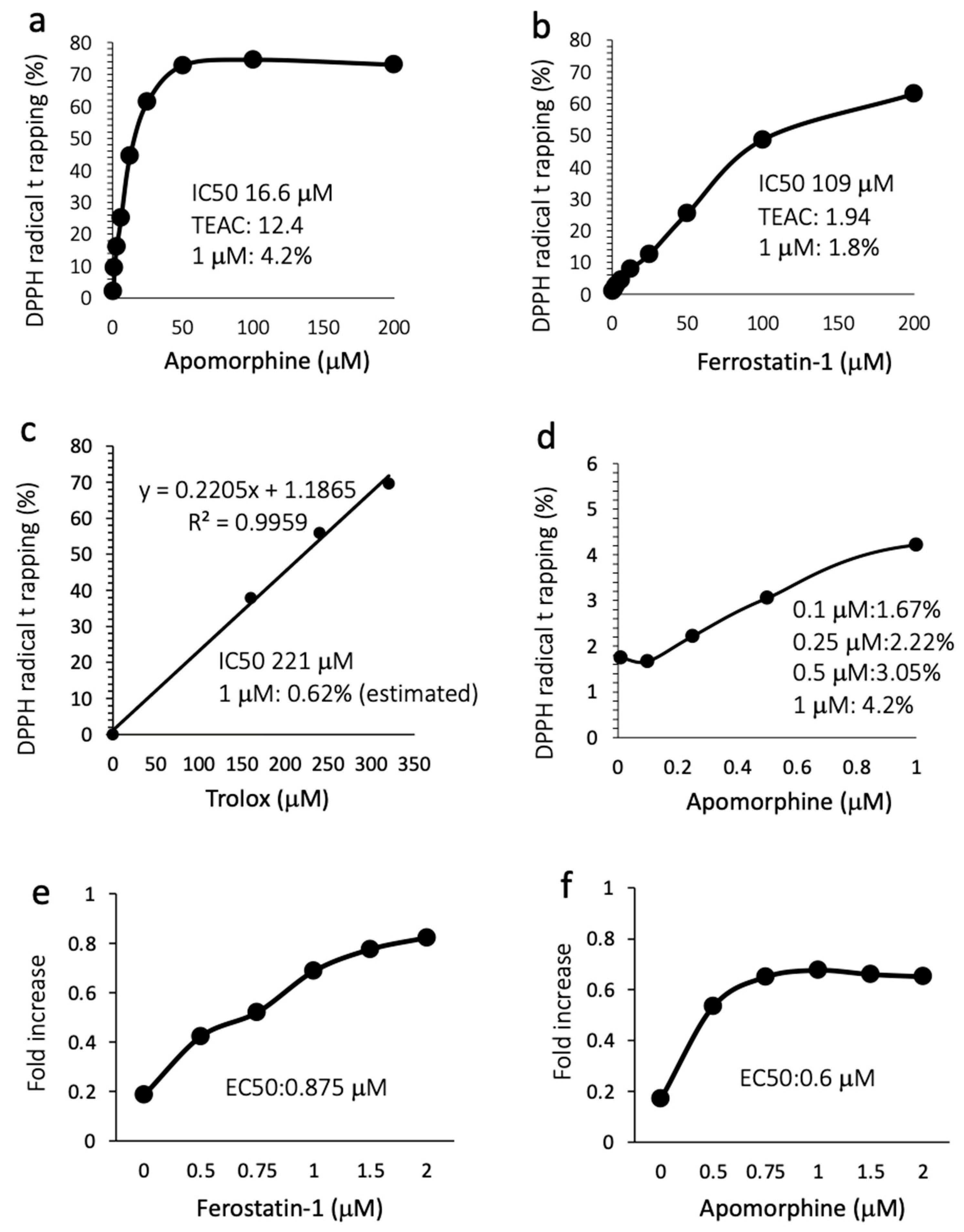
3.6. Apomorphine Had Potent Radical Trapping Activity with an Inhibitory Effect of Cell Death
3.7. Apomorphine Activates nrf2
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Rinella, M.E.; Lazarus, J.V.; Ratziu, V.; Francque, S.M.; Sanyal, A.J.; Kanwal, F.; Romero, D.; Abdelmalek, M.F.; Anstee, Q.M.; Arab, J.P.; et al. A multisociety Delphi consensus statement on new fatty liver disease nomenclature. J. Hepatol. 2023, 79, 1542–1556. [Google Scholar] [CrossRef] [PubMed]
- Younossi, Z.M.; Golabi, P.; Paik, J.M.; Henry, A.; Van Dongen, C.; Henry, L. The global epidemiology of nonalcoholic fatty liver disease (NAFLD) and nonalcoholic steatohepatitis (NASH): A systematic review. Hepatology 2023, 77, 1335–1347. [Google Scholar] [CrossRef]
- Shojaie, L.; Iorga, A.; Dara, L. Cell Death in Liver Diseases: A Review. Int. J. Mol. Sci. 2020, 21, 9682. [Google Scholar] [CrossRef] [PubMed]
- Feldstein, A.E.; Canbay, A.; Angulo, P.; Taniai, M.; Burgart, L.J.; Lindor, K.D.; Gores, G.J. Hepatocyte apoptosis and fas expression are prominent features of human nonalcoholic steatohepatitis. Gastroenterology 2003, 125, 437–443. [Google Scholar] [CrossRef] [PubMed]
- Thapaliya, S.; Wree, A.; Povero, D.; Inzaugarat, M.E.; Berk, M.; Dixon, L.; Papouchado, B.G.; Feldstein, A.E. Caspase 3 inactivation protects against hepatic cell death and ameliorates fibrogenesis in a diet-induced NASH model. Dig. Dis. Sci. 2014, 59, 1197–1206. [Google Scholar] [CrossRef] [PubMed]
- Hatting, M.; Zhao, G.; Schumacher, F.; Sellge, G.; Masaoudi, M.A.; Gaβler, N.; Boekschoten, M.; Müller, M.; Liedtke, C.; Cubero, F.J.; et al. Hepatocyte caspase-8 is an essential modulator of steatohepatitis in rodents. Hepatology 2013, 57, 2189–2201. [Google Scholar] [CrossRef] [PubMed]
- Harrison, S.A.; Goodman, Z.; Jabbar, A.; Vemulapalli, R.; Younes, Z.H.; Freilich, B.; Sheikh, M.Y.; Schattenberg, J.M.; Kayali, Z.; Zivony, A.; et al. A randomized, placebo-controlled trial of emricasan in patients with NASH and F1-F3 fibrosis. J. Hepatol. 2020, 72, 816–827. [Google Scholar] [CrossRef] [PubMed]
- Inaba, Y.; Hashiuchi, E.; Watanabe, H.; Kimura, K.; Oshima, Y.; Tsuchiya, K.; Murai, S.; Takahashi, C.; Matsumoto, M.; Kitajima, S.; et al. The transcription factor ATF3 switches cell death from apoptosis to necroptosis in hepatic steatosis in male mice. Nat. Commun. 2023, 14, 167. [Google Scholar] [CrossRef]
- Gautheron, J.; Vucur, M.; Reisinger, F.; Cardenas, D.V.; Roderburg, C.; Koppe, C.; Kreggenwinkel, K.; Schneider, A.T.; Bartneck, M.; Neumann, U.P.; et al. A positive feedback loop between RIP3 and JNK controls non-alcoholic steatohepatitis. EMBO Mol. Med. 2014, 6, 1062–1074. [Google Scholar] [CrossRef]
- Dixon, S.J.; Lemberg, K.M.; Lamprecht, M.R.; Skouta, R.; Zaitsev, E.M.; Gleason, C.E.; Patel, D.N.; Bauer, A.J.; Cantley, A.M.; Yang, W.S.; et al. Ferroptosis: An iron-dependent form of nonapoptotic cell death. Cell 2012, 149, 1060–1072. [Google Scholar] [CrossRef]
- Chen, J.; Li, X.; Ge, C.; Min, J.; Wang, F. The multifaceted role of ferroptosis in liver disease. Cell Death Differ. 2022, 29, 467–480. [Google Scholar] [CrossRef] [PubMed]
- Friedman, S.L.; Neuschwander-Tetri, B.A.; Rinella, M.; Sanyal, A.J. Mechanisms of NAFLD development and therapeutic strategies. Nat. Med. 2018, 24, 908–922. [Google Scholar] [CrossRef] [PubMed]
- Ji, H.F.; Sun, Y.; Shen, L. Effect of vitamin E supplementation on aminotransferase levels in patients with NAFLD, NASH, and CHC: Results from a meta-analysis. Nutrition 2014, 30, 986–991. [Google Scholar] [CrossRef] [PubMed]
- Jaruvongvanich, V.; Riangwiwat, T.; Sanguankeo, A.; Upala, S. Outcome of phlebotomy for treating nonalcoholic fatty liver disease: A systematic review and meta-analysis. Saudi J. Gastroenterol. 2016, 22, 407–414. [Google Scholar] [PubMed]
- Qi, J.; Kim, J.W.; Zhou, Z.; Lim, C.W.; Kim, B. Ferroptosis Affects the Progression of Nonalcoholic Steatohepatitis via the Modulation of Lipid Peroxidation-Mediated Cell Death in Mice. Am. J. Pathol. 2020, 190, 68–81. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Wang, T.; Huang, X.; Li, Y.; Sun, T.; Zang, S.; Guan, K.; Xiong, Y.; Liu, J.; Yuan, H. Targeting ferroptosis alleviates methionine-choline deficient (MCD)-diet induced NASH by suppressing liver lipotoxicity. Liver Int. 2020, 40, 1378–1394. [Google Scholar] [CrossRef] [PubMed]
- Tsurusaki, S.; Tsuchiya, Y.; Koumura, T.; Nakasone, M.; Sakamoto, T.; Matsuoka, M.; Imai, H.; Kok, C.Y.-Y.; Okochi, H.; Nakano, H.; et al. Hepatic ferroptosis plays an important role as the trigger for initiating inflammation in nonalcoholic steatohepatitis. Cell Death Dis. 2019, 10, 449. [Google Scholar] [CrossRef]
- Pfeiffer, R.F.; Gutmann, L.; Hull, K.L., Jr.; Bottini, P.B.; Sherry, J.H. Continued efficacy and safety of subcutaneous apomorphine in patients with advanced Parkinson’s disease. Park. Relat. Disord. 2007, 13, 93–100. [Google Scholar] [CrossRef]
- Carbone, F.; Djamshidian, A.; Seppi, K.; Poewe, W. Apomorphine for Parkinson’s Disease: Efficacy and Safety of Current and New Formulations. CNS Drugs 2019, 33, 905–918. [Google Scholar] [CrossRef]
- Gassen MG, Y.; Pinchasi, B.; Youdim, M.B. Apomorphine is a highly potent free radical scavenger in rat brain mitochondrial fraction. Eur. J. Pharmacol. 1996, 308, 219–225. [Google Scholar] [CrossRef]
- Heurtaux, T.; Kirchmeyer, M.; Koncina, E.; Felten, P.; Richart, L.; Huarte, O.U.; Schohn, H.; Mittelbronn, M. Apomorphine Reduces A53T alpha-Synuclein-Induced Microglial Reactivity Through Activation of NRF2 Signalling Pathway. Cell. Mol. Neurobiol. 2022, 42, 2673–2695. [Google Scholar] [CrossRef] [PubMed]
- Miyauchi, A.; Kouga, T.; Jimbo, E.F.; Matsuhashi, T.; Abe, T.; Yamagata, T.; Osaka, T. Apomorphine rescues reactive oxygen species-induced apoptosis of fibroblasts with mitochondrial disease. Mitochondrion 2019, 49, 111–120. [Google Scholar] [CrossRef] [PubMed]
- Miyauchi, A.; Watanabe, C.; Yamada, N.; Jimbo, E.J.; Kobayashi, M.; Ohishi, N.; Nagayoshi, A.; Aoki, S.; Kishita, Y.; Ohtake, A.; et al. Apomorphine is a potent inhibitor of ferroptosis independent of dopaminergic receptors. Sci. Rep. 2024, 14, 4820. [Google Scholar] [CrossRef] [PubMed]
- Arai, N.; Miura, K.; Aizawa, K.; Sekiya, M.; Nagayama, M.; Sakamoto, H.; Maeda, H.; Morimoto, N.; Iwamoto, S.; Yamamoto, H. Probiotics suppress nonalcoholic steatohepatitis and carcinogenesis progression in hepatocyte-specific PTEN knockout mice. Sci. Rep. 2022, 12, 16206. [Google Scholar] [CrossRef]
- Yue, S.; Wang, T.; Yang, Y.; Fan, Y.; Zhou, L.; Li, M.; Fu, F. Lipopolysaccharide/D-galactosamine-induced acute liver injury could be attenuated by dopamine receptor agonist rotigotine via regulating NF-kappaB signaling pathway. Int. Immunopharmacol. 2021, 96, 107798. [Google Scholar] [CrossRef] [PubMed]
- National Research Council (US) Institute for Laboratory Animal Research. Guide for the Care and Use of Laboratory Animals; National Academy of Science: Washington, DC, USA, 1996. [Google Scholar]
- Miura, K.; Kodama, Y.; Inokuchi, S.; Schnabl, B.; Aoyama, T.; Ohnishi, H.; Olefsky, J.M.; Brenner, D.A.; Seki, E. Toll-like receptor 9 promotes steatohepatitis by induction of interleukin-1beta in mice. Gastroenterology 2010, 139, 323–334.e7. [Google Scholar] [CrossRef]
- Kleiner, D.K.; Brunt, E.; Natta, M.V.; Behling, C.; Contos, M.J.; Cummings, O.W.; Ferrell, L.D.; Liu, Y.C.; Torbenson, M.S.; Unalp-Arida, A.; et al. Design and validation of a histological scoring system for nonalcoholic fatty liver disease. Hepatology 2005, 41, 1313–1321. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Z.; Zhang, Y.; Huang, X.; Can, L.; Zhao, X.; Wang, Y.; Xue, J.; Cheng, M.; Zhu, L. Thymosin beta 4 alleviates non-alcoholic fatty liver by inhibiting ferroptosis via up-regulation of GPX4. Eur. J. Pharmacol. 2021, 908, 174351. [Google Scholar] [CrossRef]
- Guan, Q.; Wang, Z.; Hu, K.; Cao, J.; Dong, Y.; Chen, Y. Melatonin Ameliorates Hepatic Ferroptosis in NAFLD by Inhibiting ER Stress via the MT2/cAMP/PKA/IRE1 Signaling Pathway. Int. J. Biol. Sci. 2023, 19, 3937–3950. [Google Scholar] [CrossRef]
- Zhou, H.; Tang, L.; Yang, Y.; Lin, L.; Dai, J.; Ge, P.; Ai, Q.; Jiang, R.; Zhang, L. Dopamine alleviated acute liver injury induced by lipopolysaccharide/d-galactosamine in mice. Int. Immunopharmacol. 2018, 61, 249–255. [Google Scholar] [CrossRef]
- Qing, J.; Ren, Y.; Zhang, Y.; Yan, M.; Zhang, H.; Wu, D.; Ma, Y.; Chen, Y.; Huang, X.; Wu, Q.; et al. Dopamine receptor D2 antagonism normalizes profibrotic macrophage-endothelial crosstalk in non-alcoholic steatohepatitis. J. Hepatol. 2022, 76, 394–406. [Google Scholar] [CrossRef] [PubMed]
- Hara, H.; Ohta, M.; Adachi, T. Apomorphine protects against 6-hydroxydopamine-induced neuronal cell death through activation of the Nrf2-ARE pathway. J. Neurosci. Res. 2006, 84, 860–866. [Google Scholar] [CrossRef]
- Cho, S.S.; Yang, J.H.; Lee, J.H.; Baek, J.S.; Ku, S.K.; Cho, I.J.; Kim, K.M.; Ki, S.H. Ferroptosis contribute to hepatic stellate cell activation and liver fibrogenesis. Free Radic. Biol. Med. 2022, 193 Pt 2, 620–637. [Google Scholar] [CrossRef] [PubMed]
- He, F.; Zhang, P.; Liu, J.; Wang, R.; Kaufman, R.J.; Yaden, B.C.; Karin, M. ATF4 suppresses hepatocarcinogenesis by inducing SLC7A11 (xCT) to block stress-related ferroptosis. J. Hepatol. 2023, 79, 362–377. [Google Scholar] [CrossRef] [PubMed]
- Nishizawa, H.; Matsumoto, M.; Chen, G.; Ishii, Y.; Tada, K.; Onodera, M.; Kato, H.; Muto, A.; Tanaka, K.; Igarashi, K.; et al. Lipid peroxidation and the subsequent cell death transmitting from ferroptotic cells to neighboring cells. Cell Death Dis. 2021, 12, 332. [Google Scholar] [CrossRef] [PubMed]
- Kong, Z.; Liu, R.; Cheng, Y. Artesunate alleviates liver fibrosis by regulating ferroptosis signaling pathway. Biomed. Pharmacother. 2019, 109, 2043–2053. [Google Scholar] [CrossRef] [PubMed]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Maeda, H.; Miura, K.; Aizawa, K.; Bat-Erdene, O.; Sashikawa-Kimura, M.; Noguchi, E.; Watanabe, M.; Yamada, N.; Osaka, H.; Morimoto, N.; et al. Apomorphine Suppresses the Progression of Steatohepatitis by Inhibiting Ferroptosis. Antioxidants 2024, 13, 805. https://doi.org/10.3390/antiox13070805
Maeda H, Miura K, Aizawa K, Bat-Erdene O, Sashikawa-Kimura M, Noguchi E, Watanabe M, Yamada N, Osaka H, Morimoto N, et al. Apomorphine Suppresses the Progression of Steatohepatitis by Inhibiting Ferroptosis. Antioxidants. 2024; 13(7):805. https://doi.org/10.3390/antiox13070805
Chicago/Turabian StyleMaeda, Hiroshi, Kouichi Miura, Kenichi Aizawa, Oyunjargal Bat-Erdene, Miho Sashikawa-Kimura, Eri Noguchi, Masako Watanabe, Naoya Yamada, Hitoshi Osaka, Naoki Morimoto, and et al. 2024. "Apomorphine Suppresses the Progression of Steatohepatitis by Inhibiting Ferroptosis" Antioxidants 13, no. 7: 805. https://doi.org/10.3390/antiox13070805
APA StyleMaeda, H., Miura, K., Aizawa, K., Bat-Erdene, O., Sashikawa-Kimura, M., Noguchi, E., Watanabe, M., Yamada, N., Osaka, H., Morimoto, N., & Yamamoto, H. (2024). Apomorphine Suppresses the Progression of Steatohepatitis by Inhibiting Ferroptosis. Antioxidants, 13(7), 805. https://doi.org/10.3390/antiox13070805