
Changes in Faecal Microbiota Profile and Plasma Biomarkers following the Administration of an Antioxidant Oleuropein-Rich Leaf Extract in a Rat Model Mimicking Colorectal Cancer
 ,
,  ,
,  , , , ,
, , , ,  ,
,  and
and 
Abstract
1. Introduction
2. Materials and Methods
2.1. Preparation of the Olive Leaf Extract and Antioxidant Activity
2.2. Animal Experimental Design and Diets
2.3. Bacterial DNA Extraction, 16S rRNA Gene Sequencing, and Sequencing Data Analysis
2.4. Faecal Water Preparation and Quantification of Short- and Medium-Chain Fatty Acids
2.5. Extraction of Metabolites from Plasma Samples
2.6. Untargeted Metabolomic Profiling by High-Resolution Mass Spectrometry
2.7. Statistical Analyses of Metabolomics and Metagenomics Data
2.8. Pathway Analysis and Plasma Metabolites as Biomarkers
3. Results and Discussion
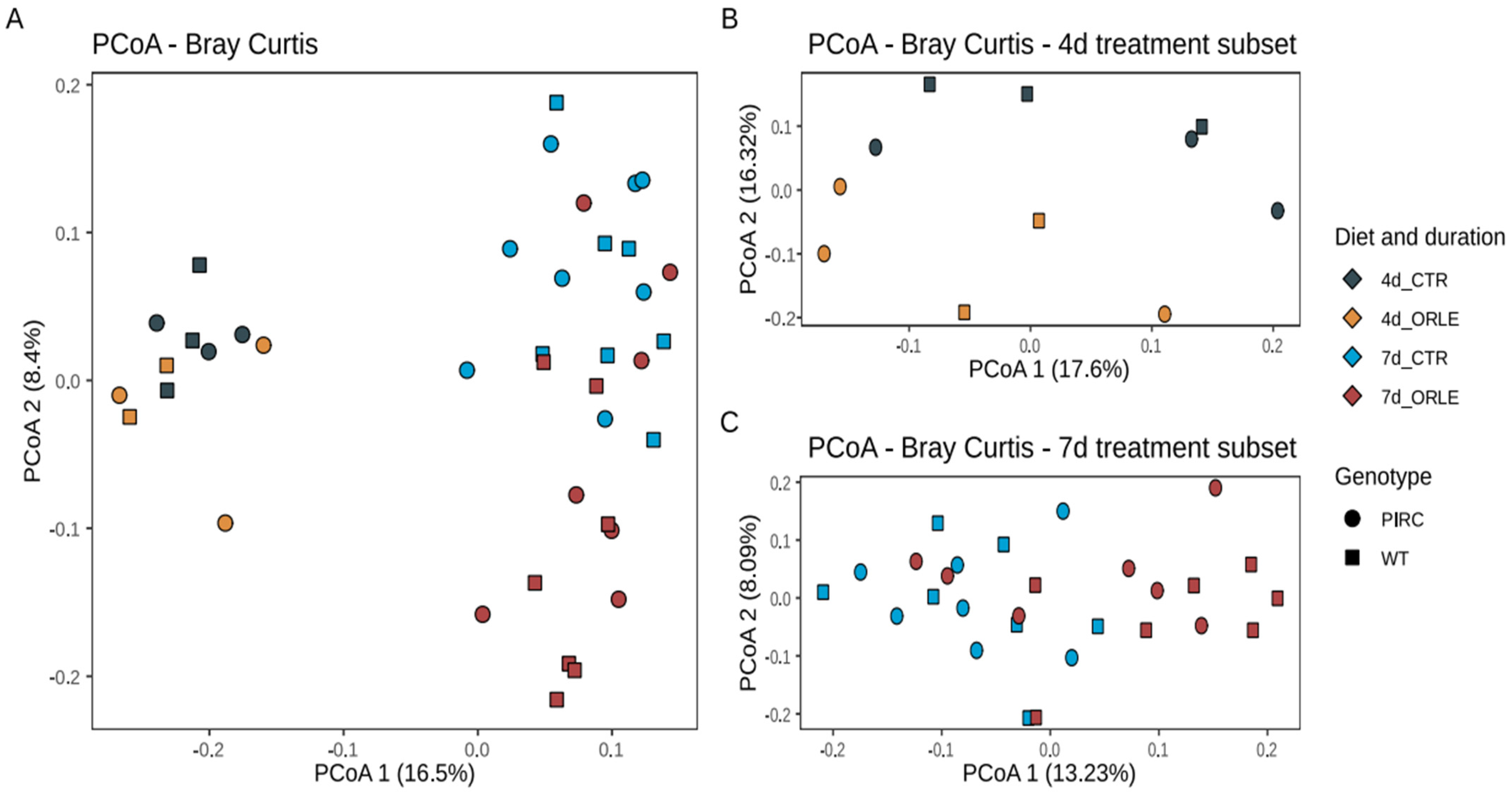
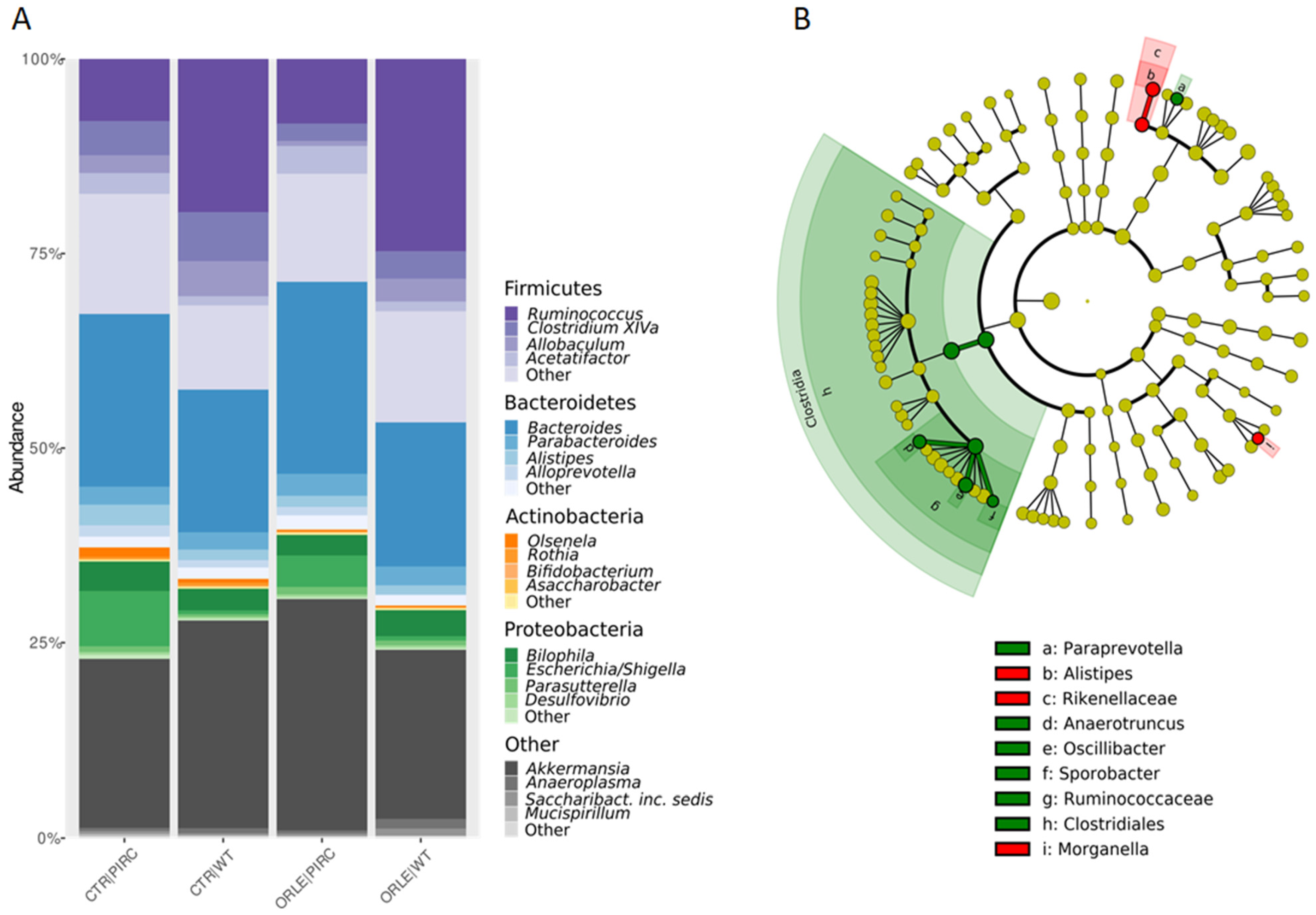
3.1. Intestinal Microbiota Profiles
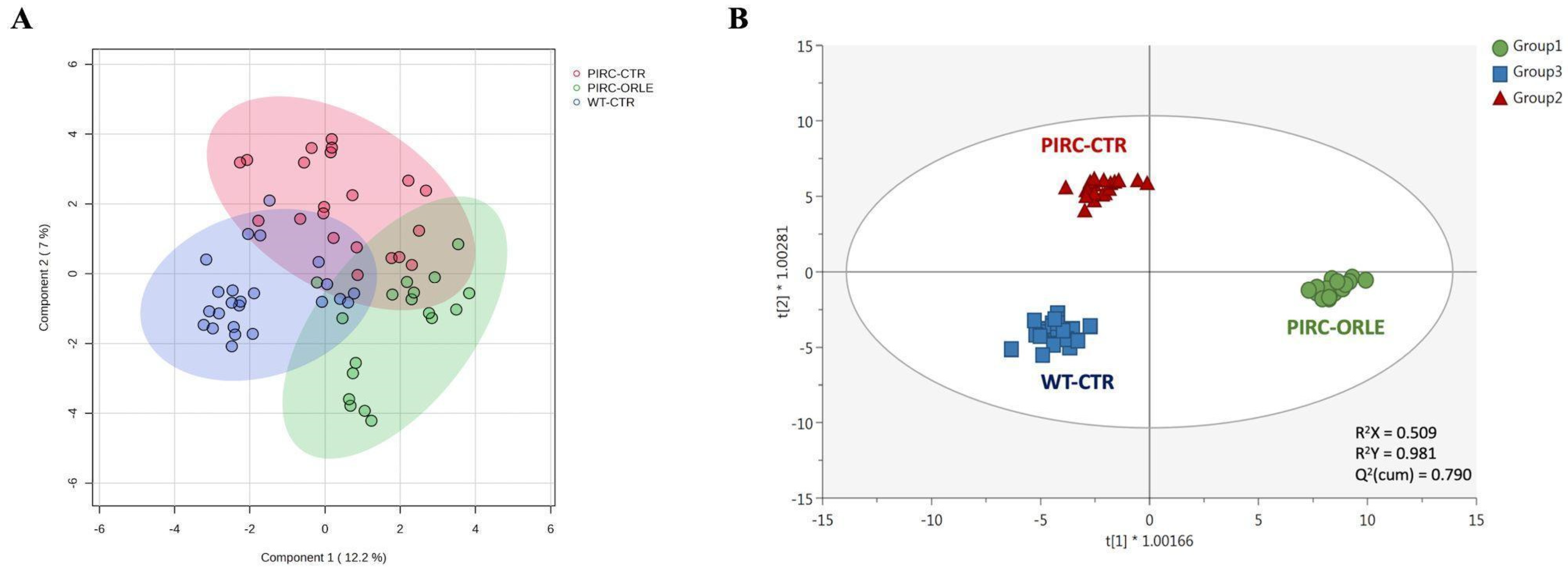
3.2. Metabolomic Profile and Multivariate Discrimination of Plasma Samples
3.3. Pathway Analyses to Highlight the Impact of ORLE on Plasma Metabolome
3.4. Importance of Lipid-like Molecules as Related to the ORLE Effect on Plasma Metabolome
3.5. Limitations of the Study
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Peršurić, Ž.; Saftić, L.; Klisović, D.; Pavelić, S.K. Polyphenol-based design of functional olive leaf infusions. Food Technol. Biotechnol. 2019, 57, 171–182. [Google Scholar] [CrossRef] [PubMed]
- Debs, E.; Abi-Khattar, A.-M.; Rajha, H.N.; Abdel-Massih, R.M.; Assaf, J.-C.; Koubaa, M.; Maroun, R.G.; Louka, N. Valorization of olive leaves through polyphenol recovery using innovative pretreatments and extraction techniques: An updated review. Separations 2023, 10, 587. [Google Scholar] [CrossRef]
- Rocchetti, G.; Luisa Callegari, M.; Senizza, A.; Giuberti, G.; Ruzzolini, J.; Romani, A.; Urciuoli, S.; Nediani, C.; Lucini, L. Oleuropein from olive leaf extracts and extra-virgin olive oil provides distinctive phenolic profiles and modulation of microbiota in the large intestine. Food Chem. 2022, 380, 132187. [Google Scholar] [CrossRef] [PubMed]
- Talhaoui, N.; Taamalli, A.; Gómez-Caravaca, A.M.; Fernández-Gutiérrez, A.; Segura-Carretero, A. Phenolic compounds in olive leaves: Analytical determination, biotic and abiotic influence, and health benefits. Food Res. Int. 2015, 77, 92–108. [Google Scholar] [CrossRef]
- Zhang, C.; Xin, X.; Zhang, J.; Zhu, S.; Niu, E.; Zhou, Z.; Liu, D. Comparative evaluation of the phytochemical profiles and antioxidant potentials of olive leaves from 32 cultivars grown in China. Molecules 2022, 27, 1292. [Google Scholar] [CrossRef] [PubMed]
- de Bock, M.; Derraik, J.G.; Brennan, C.M.; Biggs, J.B.; Morgan, P.E.; Hodgkinson, S.C.; Hofman, P.L.; Cutfield, W.S. Olive (Olea europaea L.) leaf polyphenols improve insulin sensitivity in middle-aged overweight men: A randomized, placebo-controlled, crossover trial. PLoS ONE 2013, 8, e57622. [Google Scholar] [CrossRef] [PubMed]
- Morgan, E.; Arnold, M.; Gini, A.; Lorenzoni, V.; Cabasag, C.J.; Laversanne, M.; Vignat, J.; Ferlay, J.; Murphy, N.; Bray, F. Global burden of colorectal cancer in 2020 and 2040: Incidence and mortality estimates from GLOBOCAN. Gut 2023, 72, 338–344. [Google Scholar] [CrossRef] [PubMed]
- Brenner, H.; Kloor, M.; Pox, C.P. Colorectal cancer. Lancet 2014, 383, 1490–1502. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Yu, J. The association of diet, gut microbiota and colorectal cancer: What we eat may imply what we get. Protein Cell 2018, 9, 474–487. [Google Scholar] [CrossRef]
- Long, J.; Guan, P.; Hu, X.; Yang, L.; He, L.; Lin, Q.; Luo, F.; Li, J.; He, X.; Du, Z.; et al. Natural polyphenols as targeted modulators in colon cancer: Molecular mechanisms and applications. Front. Immunol. 2021, 12, 635484. [Google Scholar] [CrossRef]
- Liu, Z.; Hu, M. Natural polyphenol disposition via coupled metabolic pathways. Expert Opin. Drug Metab. Toxicol. 2007, 3, 389–406. [Google Scholar] [CrossRef] [PubMed]
- Ruzzolini, J.; Chioccioli, S.; Monaco, N.; Peppicelli, S.; Andreucci, E.; Urciuoli, S.; Romani, A.; Luceri, C.; Tortora, K.; Calorini, L.; et al. Oleuropein-Rich Leaf Extract as a Broad Inhibitor of Tumour and Macrophage iNOS in an Apc Mutant Rat Model. Antioxidants 2021, 10, 1577. [Google Scholar] [CrossRef] [PubMed]
- Romani, A.; Campo, M.; Urciuoli, S.; Marrone, G.; Noce, A.; Bernini, R. An industrial and sustainable platform for the production of bioactive micronized powders and extracts enriched in polyphenols from Olea europaea L. and Vitis vinifera L. wastes. Front. Nutr. 2020, 7, 120. [Google Scholar] [CrossRef] [PubMed]
- López-Alarcón, C.; Denicola, A. Evaluating the antioxidant capacity of natural products: A review on chemical and cellular-based assays. Anal. Chim. Acta 2013, 763, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Heimler, D.; Vignolini, P.; Dini, M.G.; Romani, A. Rapid tests to assess the antioxidant activity of Phaseolus vulgaris L. dry beans. J. Agric. Food Chem. 2005, 53, 3053–3056. [Google Scholar] [CrossRef]
- Reagan-Shaw, S.; Nihal, M.; Ahmad, N. Dose translation from animal to human studies revisited. FASEB J. 2008, 22, 659. [Google Scholar] [CrossRef]
- Vitali, F.; Tortora, K.; Di Paola, M.; Bartolucci, G.; Menicatti, M.; De Filippo, C.; Caderni, G. Intestinal microbiota profiles in a genetic model of colon tumorigenesis correlates with colon cancer biomarkers. Sci. Rep. 2022, 12, 1432. [Google Scholar] [CrossRef]
- Saigusa, D.; Okamura, Y.; Motoike, I.N.; Katoh, Y.; Kurosawa, Y.; Saijyo, R.; Koshiba, S.; Yasuda, J.; Motohashi, H.; Sugawara, J.; et al. Establishment of protocols for global metabolomics by LC-MS for biomarker discovery. PLoS ONE 2016, 11, e0160555. [Google Scholar] [CrossRef]
- Rocchetti, G.; Rebecchi, A.; Maria Lopez, C.; Dallolio, M.; Dallolio, G.; Trevisan, M.; Lucini, L. Impact of axenic and mixed starter cultures on metabolomic and sensory profiles of ripened Italian salami. Food Chem. 2023, 402, 134182. [Google Scholar] [CrossRef]
- Fu, Q.; Zhou, S.; Yu, M.; Lu, Y.; He, G.; Huang, X.; Huang, Y. Portulaca oleracea polysaccharides modulate intestinal microflora in aged rats in vitro. Front. Microbiol. 2022, 13, 841397. [Google Scholar] [CrossRef]
- Leung, H.; Xiong, L.; Ni, Y.; Busch, A.; Bauer, M.; Press, A.T.; Panagiotou, G. Impaired flux of bile acids from the liver to the gut reveals microbiome-immune interactions associated with liver damage. npj Biofilms Microbiomes 2023, 9, 35. [Google Scholar] [CrossRef] [PubMed]
- Lopetuso, L.R.; Scaldaferri, F.; Petito, V.; Gasbarrini, A. Commensal Clostridia: Leading players in the maintenance of gut homeostasis. Gut Pathog. 2013, 5, 23. [Google Scholar] [CrossRef] [PubMed]
- Mo, Q.; Fu, A.; Deng, L.; Zhao, M.; Li, Y.; Zhang, H.; Feng, F. High-dose glycerol monolaurate up-regulated beneficial indigenous microbiota without inducing metabolic dysfunction and systemic inflammation: New insights into its antimicrobial potential. Nutrients 2019, 11, 1981. [Google Scholar] [CrossRef]
- Urzúa, C.; González, E.; Dueik, V.; Bouchon, P.; Giménez, B.; Robert, P. Olive leaves extract encapsulated by spray-drying in vacuum fried starch–gluten doughs. Food Bioprod. Process. 2017, 106, 171–180. [Google Scholar] [CrossRef]
- Wishart, D.S.; Guo, A.; Oler, E.; Wang, F.; Anjum, A.; Peters, H.; Dizon, R.; Sayeeda, Z.; Tian, S.; Lee, B.L.; et al. HMDB 5.0: The Human metabolome database for 2022. Nucleic Acids Res. 2022, 50, D622–D631. [Google Scholar] [CrossRef] [PubMed]
- Mosele, J.I.; Macià, A.; Motilva, M.J. Metabolic and microbial modulation of the large intestine ecosystem by non-absorbed siet phenolic compounds: A review. Molecules 2015, 20, 17429–17468. [Google Scholar] [CrossRef]
- Milovic, V.; Turchanowa, L. Polyamines and colon cancer. Biochem. Soc. Trans. 2003, 31, 381–383. [Google Scholar] [CrossRef]
- Kurabe, N.; Hayasaka, T.; Ogawa, M.; Masaki, N.; Ide, Y.; Waki, M.; Nakamura, T.; Kurachi, K.; Kahyo, T.; Shinmura, K.; et al. Accumulated phosphatidylcholine (16:0/16:1) in human colorectal cancer; possible involvement of LPCAT4. Cancer Sci. 2013, 104, 1295–1302. [Google Scholar] [CrossRef]
- Dobrzyńska, I.; Szachowicz-Petelska, B.; Sulkowski, S.; Figaszewski, Z. Changes in electric charge and phospholipids composition in human colorectal cancer cells. Mol. Cell. Biochem. 2005, 276, 113–119. [Google Scholar] [CrossRef]
- da Silva, S.L.; Calgarotto, A.K.; Maso, V.; Damico, D.C.; Baldasso, P.; Veber, C.L.; Villar, J.A.; Oliveira, A.R.; Comar, M., Jr.; Oliveira, K.M.; et al. Molecular modeling and inhibition of phospholipase A2 by polyhydroxy phenolic compounds. Eur. J. Med. Chem. 2009, 44, 312–321. [Google Scholar] [CrossRef]
- Wishart, D.S.; Mandal, R.; Stanislaus, A.; Ramirez-Gaona, M. Cancer metabolomics and the human metabolome database. Metabolites 2016, 6, 10. [Google Scholar] [CrossRef] [PubMed]
- Geijsen, A.J.M.R.; van Roekel, E.H.; van Duijnhoven, F.J.B.; Achaintre, D.; Bachleitner-Hofmann, T.; Baierl, A.; Bergmann, M.M.; Boehm, J.; Bours, M.J.L.; Brenner, H.; et al. Plasma metabolites associated with colorectal cancer stage: Findings from an international consortium. Int. J. Cancer Res. 2020, 146, 3256–3266. [Google Scholar] [CrossRef] [PubMed]
- Chevolleau, S.; Noguer-Meireles, M.H.; Mervant, L.; Martin, J.F.; Jouanin, I.; Pierre, F.; Naud, N.; Guéraud, F.; Debrauwer, L. Towards aldehydomics: Untargeted trapping and analysis of reactive diet-related carbonyl compounds formed in the intestinal lumen. Antioxidants 2021, 10, 1261. [Google Scholar] [CrossRef] [PubMed]
- Domínguez, R.; Pateiro, M.; Gagaoua, M.; Barba, F.J.; Zhang, W.; Lorenzo, J.M. A comprehensive review on lipid oxidation in meat and meat products. Antioxidants 2019, 8, 429. [Google Scholar] [CrossRef] [PubMed]
- Jacobs, R.J.; Voorneveld, P.W.; Kodach, L.L.; Hardwick, J.C. Cholesterol metabolism and colorectal cancers. Curr. Opin. Pharmacol. 2012, 12, 690–695. [Google Scholar] [CrossRef] [PubMed]
- Yan, G.; Li, L.; Zhu, B.; Li, Y. Lipidome in colorectal cancer. Oncotarget 2016, 7, 33429–33439. [Google Scholar] [CrossRef] [PubMed]
- Roopashree, P.G.; Shetty, S.S.; Kumari, N.S. Effect of medium chain fatty acid in human health and disease. J. Funct. Foods 2021, 87, 104724. [Google Scholar] [CrossRef]
- Crotti, S.; Agnoletto, E.; Cancemi, G.; Di Marco, V.; Traldi, P.; Pucciarelli, S.; Nitti, D.; Agostini, M. Altered plasma levels of decanoic acid in colorectal cancer as a new diagnostic biomarker. Anal. Bioanal. Chem. 2016, 408, 6321–6328. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
| Variables | Df | Sum of SQS | R2 | F-test | p-value |
| ORLE treatment | 1 | 0.2461 | 0.05346 | 2.5224 | 0.002 * |
| Genotype | 1 | 0.1273 | 0.02767 | 1.3054 | 0.105 |
| Length | 1 | 0.7176 | 0.15589 | 7.3556 | 0.001 * |
| Residual | 36 | 3.5119 | 0.76298 | ||
| Total | 39 | 4.6029 | 1.00000 | ||
| 4-day Intervention Subset | |||||
| Variables | Df | Sum of SQS | R2 | F-test | p-value |
| ORLE treatment | 1 | 0.14121 | 0.14525 | 1.5535 | 0.020 * |
| Genotype | 1 | 0.08233 | 0.08468 | 0.9057 | 0.644 |
| Interaction | 1 | 0.11236 | 0.11588 | 1.2361 | 0.143 |
| Residual | 7 | 0.63627 | 0.65449 | ||
| Total | 10 | 0.97216 | 1.00000 | ||
| 7-day Intervention Subset | |||||
| Variables | Df | Sum of SQS | R2 | F-test | p-value |
| ORLE treatment | 1 | 0.21495 | 0.07866 | 2.2510 | 0.001 * |
| Genotype | 1 | 0.12942 | 0.04736 | 1.3553 | 0.047 * |
| Interaction | 1 | 0.09656 | 0.03534 | 1.0113 | 0.457 |
| Residual | 24 | 2.29174 | 0.83864 | ||
| Total | 27 | 2.73268 | 1.00000 | ||
| Chemical Class | Sig. (p-Value) | PIRC ORLE vs. CTR | Plasma Biomarker (OPLS-DA) |
|---|---|---|---|
| Polyamines | 4.11 × 10−3 | LogFC: 1.24 | Spermidine (VIP score = 1.82) |
| Amino acids and peptides | 6.48 × 10−3 | LogFC: −5.00 | Tryptophan (VIP score = 1.37) |
| Fatty acids and conjugates | 1.60 × 10−2 | LogFC: 2.36 | 13,16,19-Docosatrienoic acid (VIP score = 1.89) |
| Glycerophospholipids | 5.54 × 10−3 | LogFC: −0.51 | LysoPC(16:0) (VIP score = 1.83) |
| Medium-chain aldehydes | 1.99 × 10−2 | LogFC: 1.99 | 3,6-Undecadienal (VIP score = 1.02) |
| Phenolic metabolites | 1.44 × 10−2 | LogFC: 3.72 | Dihydroferulic acid, 4-O-glucuronide (VIP score = 1.72) |
| Pyridines and derivatives | 1.89 × 10−2 | LogFC: −1.93 | 3-Hexyl-pyridine (VIP score = 1.34) |
| Pyrimidines and derivatives | 2.21 × 10−2 | LogFC: 0.03 | Thymine (VIP score = 1.41) |
| Terpenoids | 9.40 × 10−3 | LogFC: −1.01 | Terpinyl-isovalerate (VIP score = 1.45) |
| Carbohydrates and conjugates | 2.44 × 10−2 | LogFC: 2.25 | Ribose 5-phosphate (VIP score = 1.32) |
| Other metabolites | 1.06 × 10−2 | LogFC: 0.30 | 13′-Hydroxy-tocopherol (VIP score = 1.23) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chioccioli, S.; Rocchetti, G.; Ruzzolini, J.; Urciuoli, S.; Vitali, F.; Bartolucci, G.; Pallecchi, M.; Caderni, G.; De Filippo, C.; Nediani, C.; et al. Changes in Faecal Microbiota Profile and Plasma Biomarkers following the Administration of an Antioxidant Oleuropein-Rich Leaf Extract in a Rat Model Mimicking Colorectal Cancer. Antioxidants 2024, 13, 724. https://doi.org/10.3390/antiox13060724
Chioccioli S, Rocchetti G, Ruzzolini J, Urciuoli S, Vitali F, Bartolucci G, Pallecchi M, Caderni G, De Filippo C, Nediani C, et al. Changes in Faecal Microbiota Profile and Plasma Biomarkers following the Administration of an Antioxidant Oleuropein-Rich Leaf Extract in a Rat Model Mimicking Colorectal Cancer. Antioxidants. 2024; 13(6):724. https://doi.org/10.3390/antiox13060724
Chicago/Turabian StyleChioccioli, Sofia, Gabriele Rocchetti, Jessica Ruzzolini, Silvia Urciuoli, Francesco Vitali, Gianluca Bartolucci, Marco Pallecchi, Giovanna Caderni, Carlotta De Filippo, Chiara Nediani, and et al. 2024. "Changes in Faecal Microbiota Profile and Plasma Biomarkers following the Administration of an Antioxidant Oleuropein-Rich Leaf Extract in a Rat Model Mimicking Colorectal Cancer" Antioxidants 13, no. 6: 724. https://doi.org/10.3390/antiox13060724
APA StyleChioccioli, S., Rocchetti, G., Ruzzolini, J., Urciuoli, S., Vitali, F., Bartolucci, G., Pallecchi, M., Caderni, G., De Filippo, C., Nediani, C., & Lucini, L. (2024). Changes in Faecal Microbiota Profile and Plasma Biomarkers following the Administration of an Antioxidant Oleuropein-Rich Leaf Extract in a Rat Model Mimicking Colorectal Cancer. Antioxidants, 13(6), 724. https://doi.org/10.3390/antiox13060724