


A Mechanistic Study of the Osteogenic Effect of Arecoline in an Osteoporosis Model: Inhibition of Iron Overload-Induced Osteogenesis by Promoting Heme Oxygenase-1 Expression

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Methods and Materials
2.1. Materials and Reagents
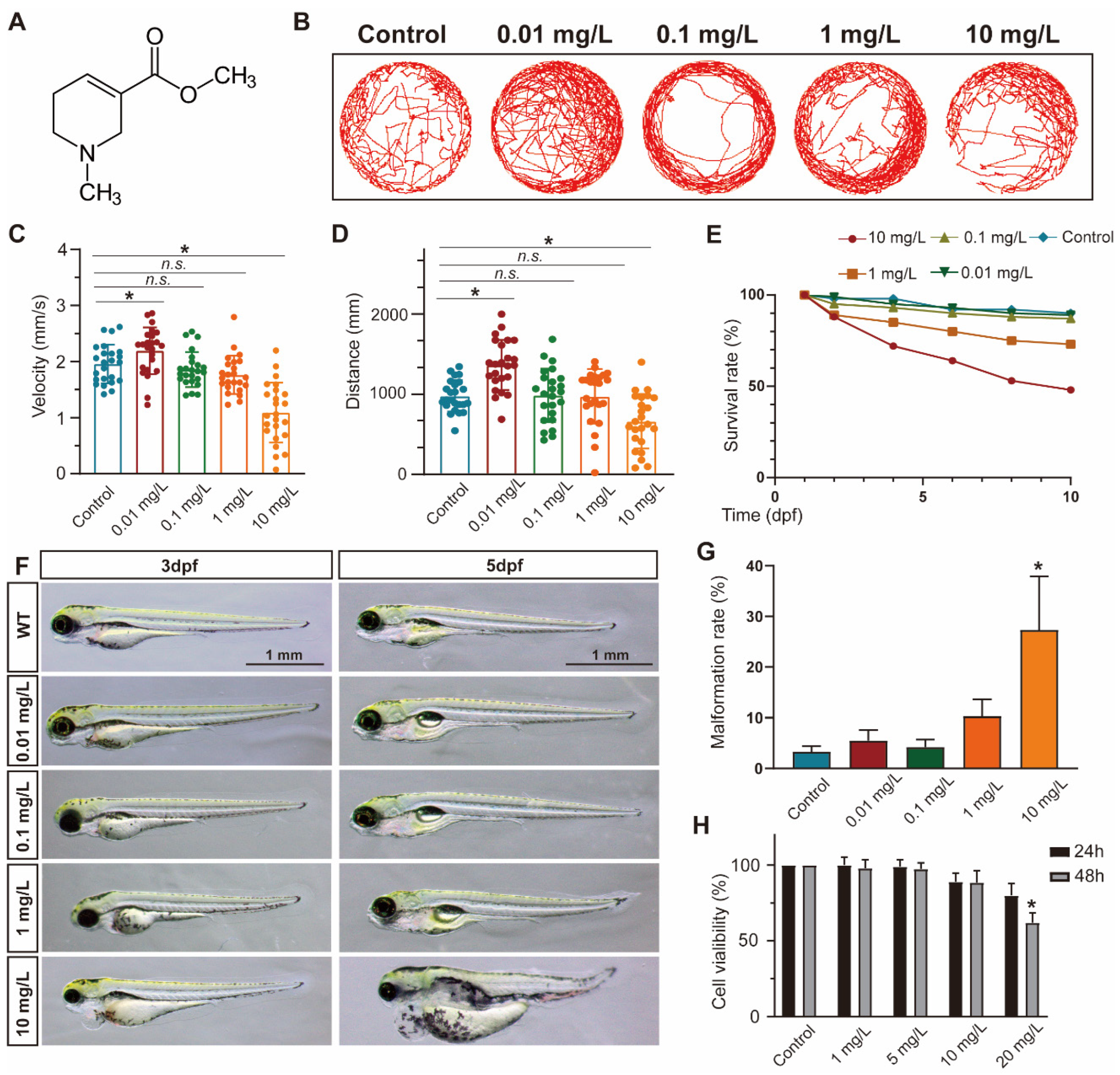
2.2. Zebrafish Husbandry, Treatment and Locomotion Analysis
2.3. Generation of the Transgenic Zebrafish Line
2.4. Calcein Staining
2.5. qPCR Analysis
2.6. Cell Proliferation Assay
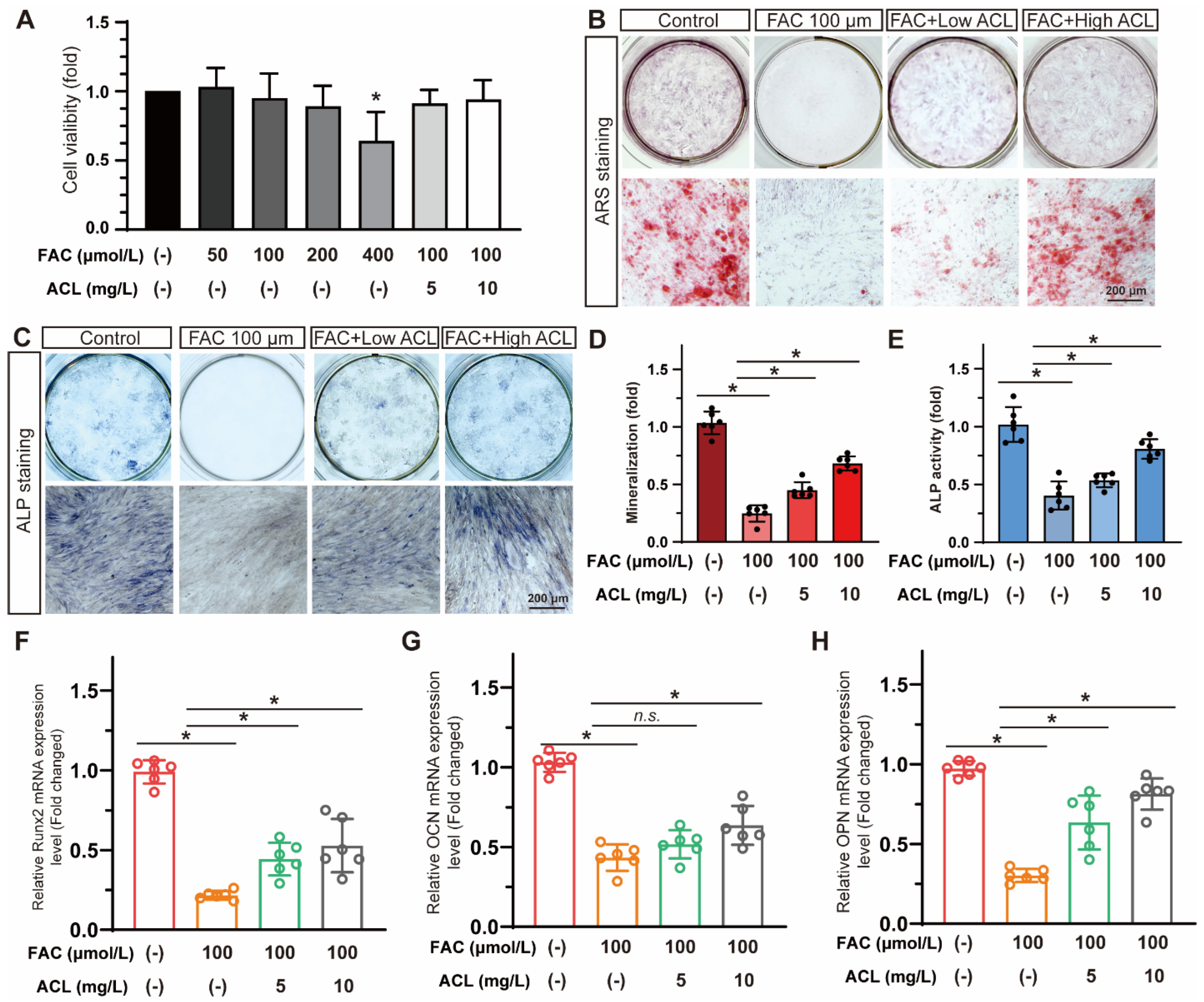
2.7. Alizarin Red and Alkaline Phosphatase Staining
2.8. RNA Sequencing and Data Analysis
2.9. ROS Production Measurement
2.10. Quantification of Iron, Malondialdehyde (MDA), and Glutathione (GSH) Levels in Zebrafish Larvae
2.11. Transmission Electron Microscopy (TEM)
2.12. Oxidative Stress Analysis
2.13. Intracellular ROS, Iron, Lipid Peroxidation Levels Determination
2.14. siRNA Transfection
2.15. Western Blot Analysis
2.16. Statistical Analysis
3. Results
3.1. ACL Exposure Caused Concentration-Dependent Developmental Malformations in Zebrafish
3.2. ACL Alleviated FAC-Induced Inhibition of Osteogenesis in Zebrafish
3.3. ACL Improved the Osteogenic Differentiation of MC3T3-E1 Cells under FAC Treatment
3.4. Ferroptosis May Play a Role in the Anti-Osteoporotic Effects of ACL under FAC Exposure
3.5. Ferroptosis Was Inhibited by ACL Treatment under FAC Exposure
3.6. Inhibition of Ferroptosis by ACL Can Be Reversed by HO-1 Inhibition in MC3T3-E1 Cells
3.7. The Protective Effect of ACL Can Be Inhibited by hmox1a Knockout in Zebrafish
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Zhang, P.; Chen, L.; Zhao, Q.; Du, X.; Bi, M.; Li, Y.; Jiao, Q.; Jiang, H. Ferroptosis was more initial in cell death caused by iron overload and its underlying mechanism in Parkinson’s disease. Free. Radic. Biol. Med. 2020, 152, 227–234. [Google Scholar] [CrossRef] [PubMed]
- Wang, F.; Weng, Z.; Song, H.; Bao, Y.; Sui, H.; Fang, Y.; Tang, X.; Shen, X. Ferric ammonium citrate (FAC)-induced inhibition of osteoblast proliferation/differentiation and its reversal by soybean-derived peptides (SDP). Food Chem. Toxicol. Int. J. Publ. Br. Ind. Biol. Res. Assoc. 2021, 156, 112527. [Google Scholar] [CrossRef] [PubMed]
- Luo, C.; Xu, W.; Tang, X.; Liu, X.; Cheng, Y.; Wu, Y.; Xie, Z.; Wu, X.; He, X.; Wang, Q.; et al. Canonical Wnt signaling works downstream of iron overload to prevent ferroptosis from damaging osteoblast differentiation. Free. Radic. Biol. Med. 2022, 188, 337–350. [Google Scholar] [CrossRef] [PubMed]
- Khosla, S.; Wright, N.C.; Elderkin, A.L.; Kiel, D.P. Osteoporosis in the USA: Prevention and unmet needs. Lancet Diabetes Endocrinol. 2023, 11, 19–20. [Google Scholar] [CrossRef] [PubMed]
- Xu, Z.; Sun, W.; Li, Y.; Ling, S.; Zhao, C.; Zhong, G.; Zhao, D.; Song, J.; Song, H.; Li, J.; et al. The regulation of iron metabolism by hepcidin contributes to unloading-induced bone loss. Bone 2017, 94, 152–161. [Google Scholar] [CrossRef] [PubMed]
- Lv, H.; Wang, Y.; Zhen, C.; Liu, J.; Chen, X.; Zhang, G.; Yao, W.; Guo, H.; Wei, Y.; Wang, S.; et al. A static magnetic field improves bone quality and balances the function of bone cells with regulation on iron metabolism and redox status in type 1 diabetes. FASEB J. Off. Publ. Fed. Am. Soc. Exp. Biol. 2023, 37, e22985. [Google Scholar] [CrossRef] [PubMed]
- Liu, F.L.; Chen, C.L.; Lai, C.C.; Lee, C.C.; Chang, D.M. Arecoline suppresses RANKL-induced osteoclast differentiation in vitro and attenuates LPS-induced bone loss in vivo. Phytomedicine Int. J. Phytother. Phytopharm. 2020, 69, 153195. [Google Scholar] [CrossRef] [PubMed]
- Liu, P.F.; Chang, Y.F. The Controversial Roles of Areca Nut: Medicine or Toxin? Int. J. Mol. Sci. 2023, 24, 8996. [Google Scholar] [CrossRef] [PubMed]
- Lin, B.; Xu, P.; Zheng, J.; Deng, X.; Ye, Q.; Huang, Z.; Wang, N. Effects and mechanisms of natural alkaloids for prevention and treatment of osteoporosis. Front. Pharmacol. 2022, 13, 1014173. [Google Scholar] [CrossRef]
- Ko, A.M.; Tu, H.P.; Ko, Y.C. Systematic Review of Roles of Arecoline and Arecoline N-Oxide in Oral Cancer and Strategies to Block Carcinogenesis. Cells 2023, 12, 1208. [Google Scholar] [CrossRef]
- Sartinah, A.; Nugrahani, I.; Ibrahim, S.; Anggadiredja, K. Potential metabolites of Arecaceae family for the natural anti-osteoarthritis medicine: A review. Heliyon 2022, 8, e12039. [Google Scholar] [CrossRef] [PubMed]
- Ru, Q.; Li, Y.; Xie, W.; Ding, Y.; Chen, L.; Xu, G.; Wu, Y.; Wang, F. Fighting age-related orthopedic diseases: Focusing on ferroptosis. Bone Res. 2023, 11, 12. [Google Scholar] [CrossRef] [PubMed]
- Duan, J.Y.; Lin, X.; Xu, F.; Shan, S.K.; Guo, B.; Li, F.X.; Wang, Y.; Zheng, M.H.; Xu, Q.S.; Lei, L.M.; et al. Ferroptosis and Its Potential Role in Metabolic Diseases: A Curse or Revitalization? Front. Cell Dev. Biol. 2021, 9, 701788. [Google Scholar] [CrossRef]
- Zhang, H.; Wang, A.; Li, G.; Zhai, Q.; Huang, Z.; Wang, X.; Cao, Z.; Liu, L.; Liu, G.; Chen, B.; et al. Osteoporotic bone loss from excess iron accumulation is driven by NOX4-triggered ferroptosis in osteoblasts. Free. Radic. Biol. Med. 2023, 198, 123–136. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Xu, J.; Qiu, J.; Xing, C.; Li, X.; Leng, B.; Su, Y.; Lin, J.; Lin, J.; Mei, X.; et al. Novel and rapid osteoporosis model established in zebrafish using high iron stress. Biochem. Biophys. Res. Commun. 2018, 496, 654–660. [Google Scholar] [CrossRef] [PubMed]
- Peng, W.H.; Lee, Y.C.; Chau, Y.P.; Lu, K.S.; Kung, H.N. Short-term exposure of zebrafish embryos to arecoline leads to retarded growth, motor impairment, and somite muscle fiber changes. Zebrafish 2015, 12, 58–70. [Google Scholar] [CrossRef]
- Serikuly, N.; Alpyshov, E.T.; Wang, D.; Wang, J.; Yang, L.; Hu, G.; Yan, D.; Demin, K.A.; Kolesnikova, T.O.; Galstyan, D.; et al. Effects of acute and chronic arecoline in adult zebrafish: Anxiolytic-like activity, elevated brain monoamines and the potential role of microglia. Prog. Neuro-Psychopharmacol. Biol. Psychiatry 2021, 104, 109977. [Google Scholar] [CrossRef] [PubMed]
- Siregar, P.; Audira, G.; Feng, L.Y.; Lee, J.H.; Santoso, F.; Yu, W.H.; Lai, Y.H.; Li, J.H.; Lin, Y.T.; Chen, J.R.; et al. Pharmaceutical Assessment Suggests Locomotion Hyperactivity in Zebrafish Triggered by Arecoline Might Be Associated with Multiple Muscarinic Acetylcholine Receptors Activation. Toxins 2021, 13, 259. [Google Scholar] [CrossRef]
- Siregar, P.; Audira, G.; Castillo, A.L.; Roldan, M.J.M.; Suryanto, M.E.; Liu, R.X.; Lin, Y.T.; Lai, Y.H.; Hsiao, C.D. Comparison of the psychoactive activity of four primary Areca nut alkaloids in zebrafish by behavioral approach and molecular docking. Biomed. Pharmacother. Biomed. Pharmacother. 2022, 155, 113809. [Google Scholar] [CrossRef]
- Ryter, S.W. Heme Oxygenase-1: An Anti-Inflammatory Effector in Cardiovascular, Lung, and Related Metabolic Disorders. Antioxidants 2022, 11, 555. [Google Scholar] [CrossRef]
- Stockwell, B.R. A powerful cell-protection system prevents cell death by ferroptosis. Nature 2019, 575, 597–598. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Lin, Y.; Wang, M.; Yuan, K.; Wang, Q.; Mu, P.; Du, J.; Yu, Z.; Yang, S.; Huang, K.; et al. Targeting ferroptosis suppresses osteocyte glucolipotoxicity and alleviates diabetic osteoporosis. Bone Res. 2022, 10, 26. [Google Scholar] [CrossRef] [PubMed]
- Chau, L.Y. Heme oxygenase-1: Emerging target of cancer therapy. J. Biomed. Sci. 2015, 22, 22. [Google Scholar] [CrossRef] [PubMed]
- Chiabrando, D.; Vinchi, F.; Fiorito, V.; Mercurio, S.; Tolosano, E. Heme in pathophysiology: A matter of scavenging, metabolism and trafficking across cell membranes. Front. Pharmacol. 2014, 5, 61. [Google Scholar] [CrossRef] [PubMed]
- Pan, Z.; He, Q.; Zeng, J.; Li, S.; Li, M.; Chen, B.; Yang, J.; Xiao, J.; Zeng, C.; Luo, H.; et al. Naringenin protects against iron overload-induced osteoarthritis by suppressing oxidative stress. Phytomedicine Int. J. Phytother. Phytopharm. 2022, 105, 154330. [Google Scholar] [CrossRef] [PubMed]
- Tang, Z.; Ju, Y.; Dai, X.; Ni, N.; Liu, Y.; Zhang, D.; Gao, H.; Sun, H.; Zhang, J.; Gu, P. HO-1-mediated ferroptosis as a target for protection against retinal pigment epithelium degeneration. Redox Biol. 2021, 43, 101971. [Google Scholar] [CrossRef] [PubMed]
- Ryter, S.W. Heme Oxgenase-1, a Cardinal Modulator of Regulated Cell Death and Inflammation. Cells 2021, 10, 515. [Google Scholar] [CrossRef] [PubMed]
- Che, J.; Yang, J.; Zhao, B.; Shang, P. HO-1: A new potential therapeutic target to combat osteoporosis. Eur. J. Pharmacol. 2021, 906, 174219. [Google Scholar] [CrossRef]
- Otterbein, L.E.; Foresti, R.; Motterlini, R. Heme Oxygenase-1 and Carbon Monoxide in the Heart: The Balancing Act Between Danger Signaling and Pro-Survival. Circ. Res. 2016, 118, 1940–1959. [Google Scholar] [CrossRef]
- Chiang, S.K.; Chen, S.E.; Chang, L.C. The Role of HO-1 and Its Crosstalk with Oxidative Stress in Cancer Cell Survival. Cells 2021, 10, 2401. [Google Scholar] [CrossRef]
- Wlodkowic, D.; Campana, O. Toward High-Throughput Fish Embryo Toxicity Tests in Aquatic Toxicology. Environ. Sci. Technol. 2021, 55, 3505–3513. [Google Scholar] [CrossRef] [PubMed]
- Rosa, J.T.; Laizé, V.; Gavaia, P.J.; Cancela, M.L. Fish Models of Induced Osteoporosis. Front. Cell Dev. Biol. 2021, 9, 672424. [Google Scholar] [CrossRef] [PubMed]
- Miao, W.; He, L.; Zhang, Y.; Zhu, X.; Jiang, Y.; Liu, P.; Zhang, T.; Li, C. Ferroptosis is partially responsible for dexamethasone-induced T cell ablation, but not osteoporosis in larval zebrafish. Ecotoxicol. Environ. Saf. 2022, 242, 113872. [Google Scholar] [CrossRef] [PubMed]
- Luo, K.; Stocker, R.; Britton, W.J.; Kikuchi, K.; Oehlers, S.H. Haem oxygenase limits Mycobacterium marinum infection-induced detrimental ferrostatin-sensitive cell death in zebrafish. FEBS J. 2022, 289, 671–681. [Google Scholar] [CrossRef] [PubMed]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jiang, Z.; Deng, L.; Xiang, G.; Xu, X.; Wang, Y. A Mechanistic Study of the Osteogenic Effect of Arecoline in an Osteoporosis Model: Inhibition of Iron Overload-Induced Osteogenesis by Promoting Heme Oxygenase-1 Expression. Antioxidants 2024, 13, 430. https://doi.org/10.3390/antiox13040430
Jiang Z, Deng L, Xiang G, Xu X, Wang Y. A Mechanistic Study of the Osteogenic Effect of Arecoline in an Osteoporosis Model: Inhibition of Iron Overload-Induced Osteogenesis by Promoting Heme Oxygenase-1 Expression. Antioxidants. 2024; 13(4):430. https://doi.org/10.3390/antiox13040430
Chicago/Turabian StyleJiang, Zhongjing, Linhua Deng, Gang Xiang, Xia Xu, and Yunjia Wang. 2024. "A Mechanistic Study of the Osteogenic Effect of Arecoline in an Osteoporosis Model: Inhibition of Iron Overload-Induced Osteogenesis by Promoting Heme Oxygenase-1 Expression" Antioxidants 13, no. 4: 430. https://doi.org/10.3390/antiox13040430
APA StyleJiang, Z., Deng, L., Xiang, G., Xu, X., & Wang, Y. (2024). A Mechanistic Study of the Osteogenic Effect of Arecoline in an Osteoporosis Model: Inhibition of Iron Overload-Induced Osteogenesis by Promoting Heme Oxygenase-1 Expression. Antioxidants, 13(4), 430. https://doi.org/10.3390/antiox13040430