
Long-Term Region-Specific Mitochondrial Functionality Changes in Both Cerebral Hemispheres after fMCAo Model of Ischemic Stroke



Abstract
1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Ethics Statement
2.3. Chemicals
2.4. Intraluminal Filament Induced Middle Cerebral Artery Occlusion Model (fMCAo)
2.5. Brain Tissue Homogenate Preparation
2.6. High-Resolution Respirometry
2.7. Measurement of Mitochondrial H2O2 Flow Formation
2.8. Statistical Analysis
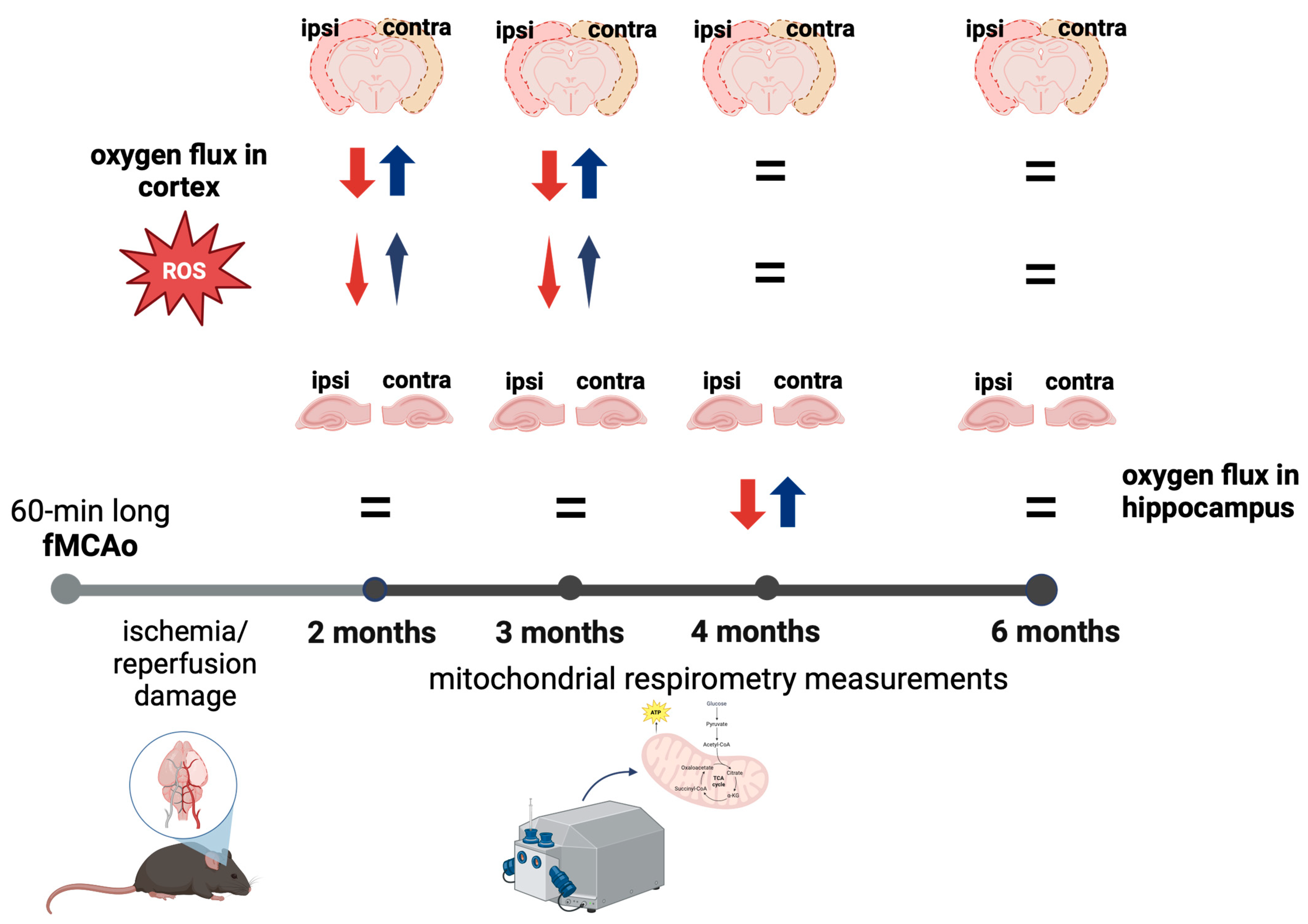
3. Results
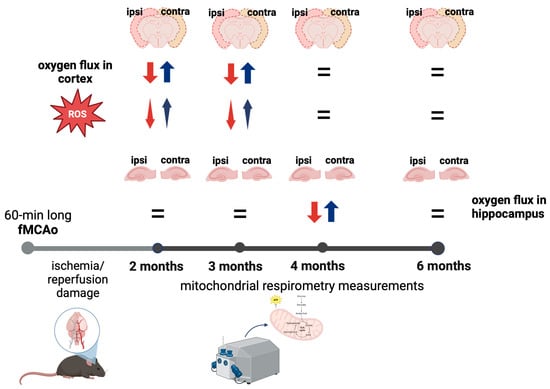
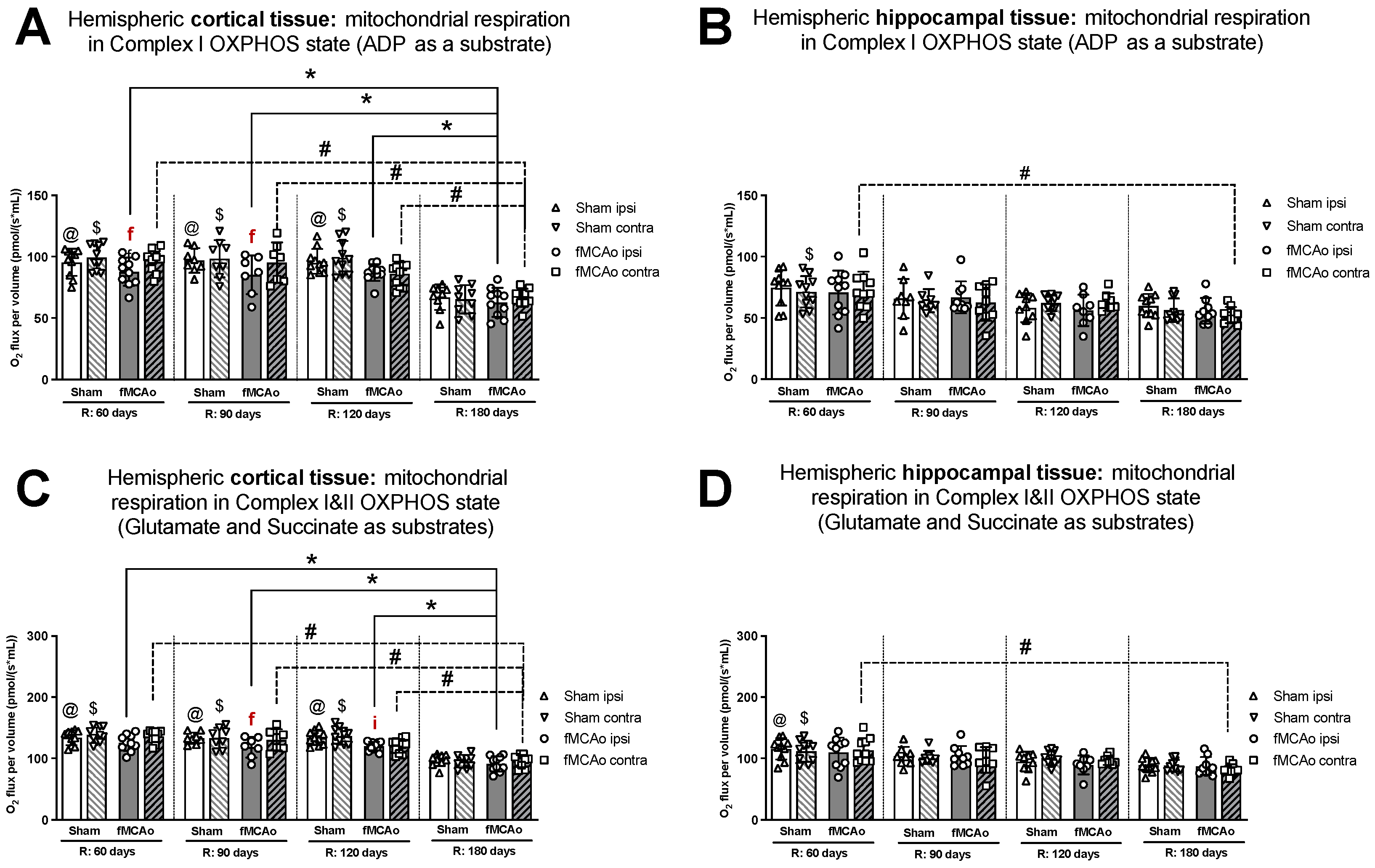
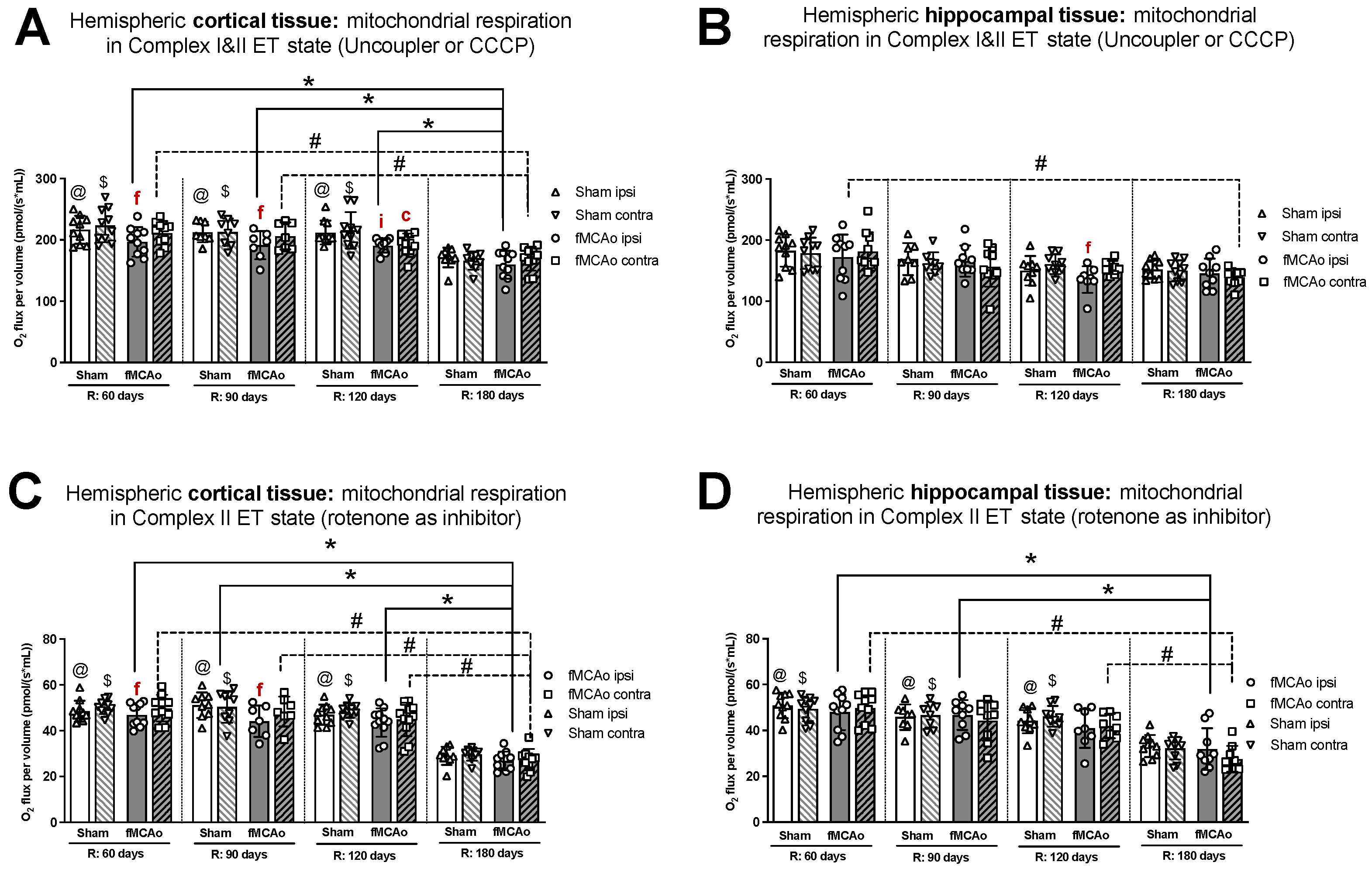
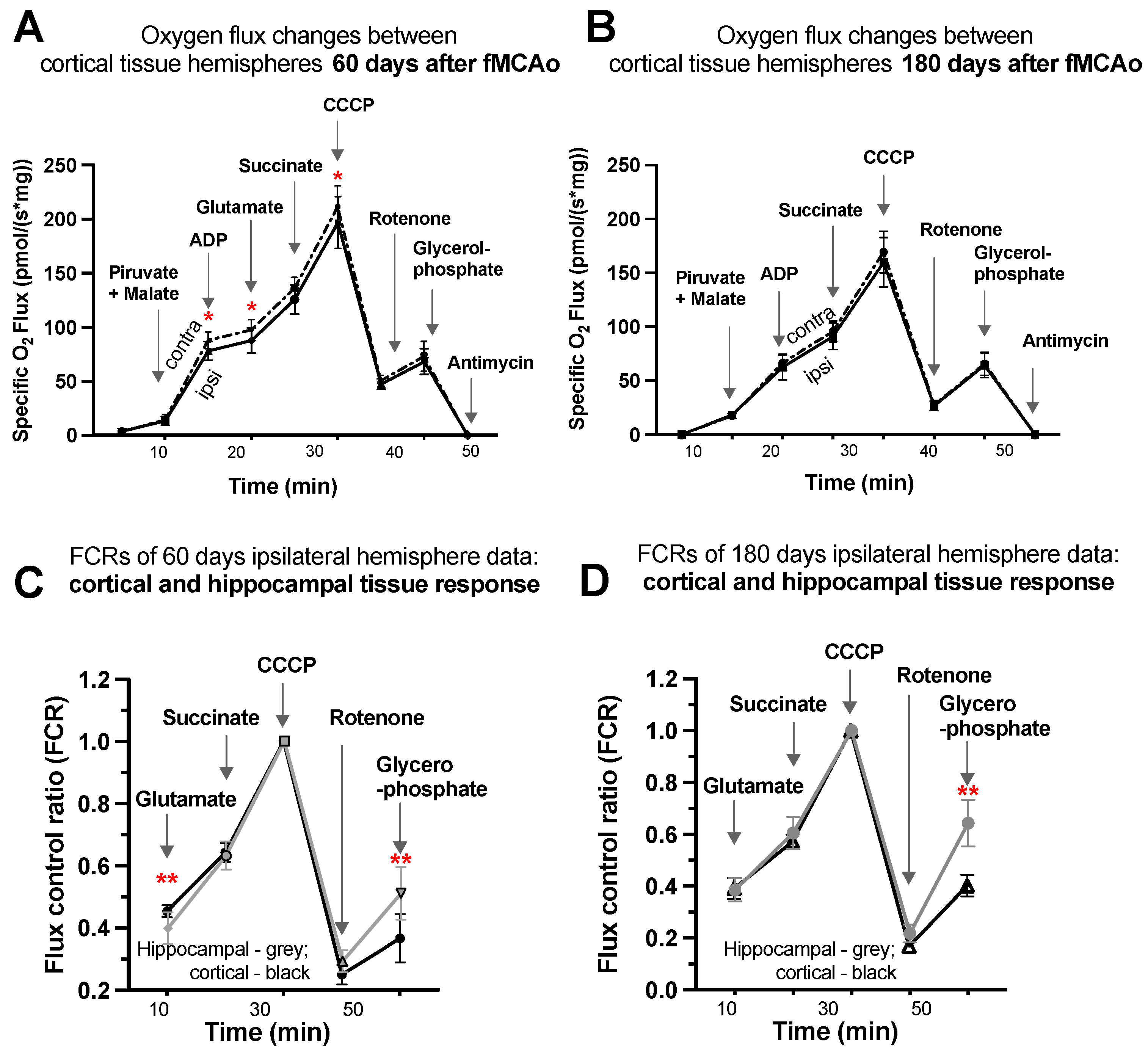
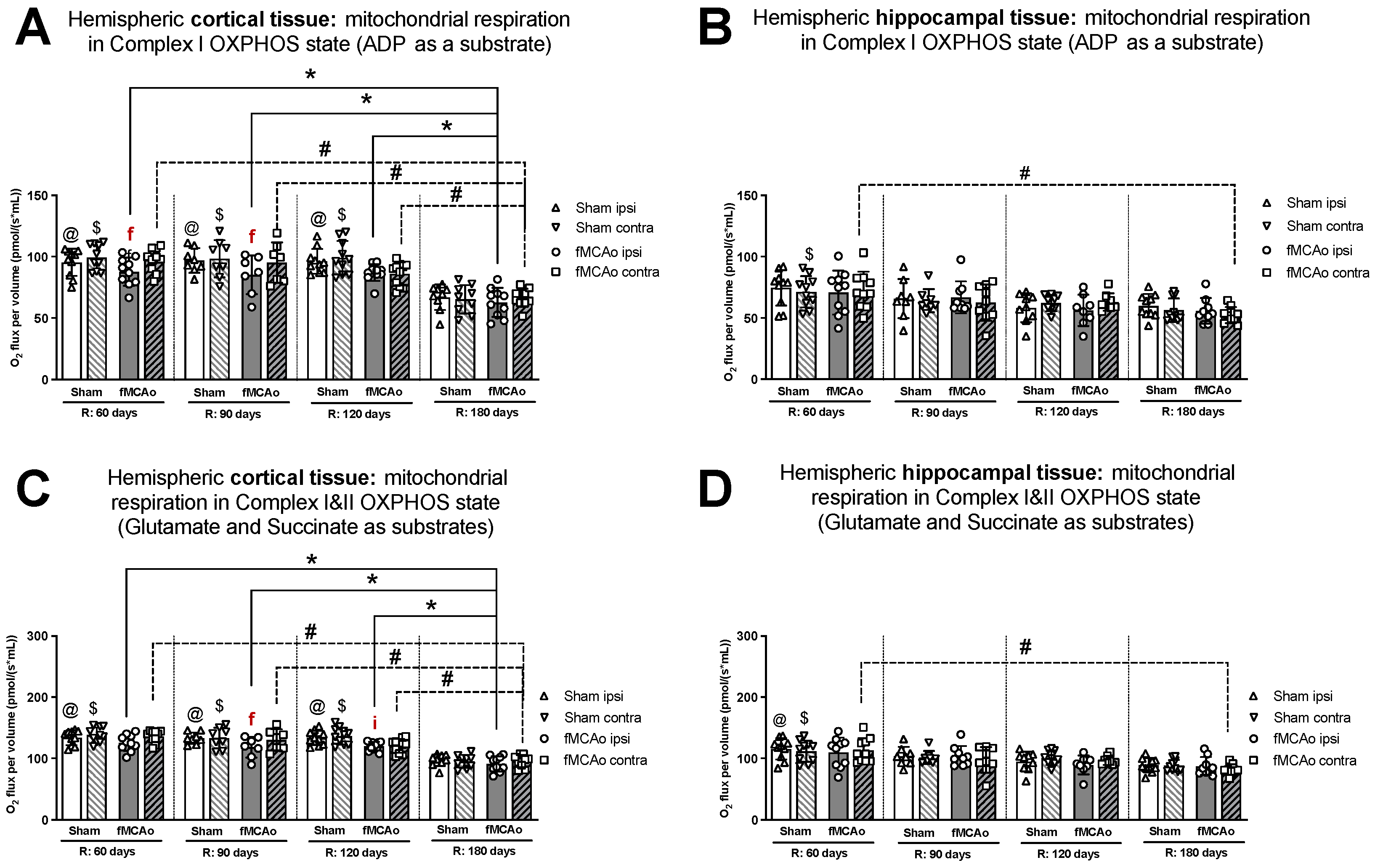
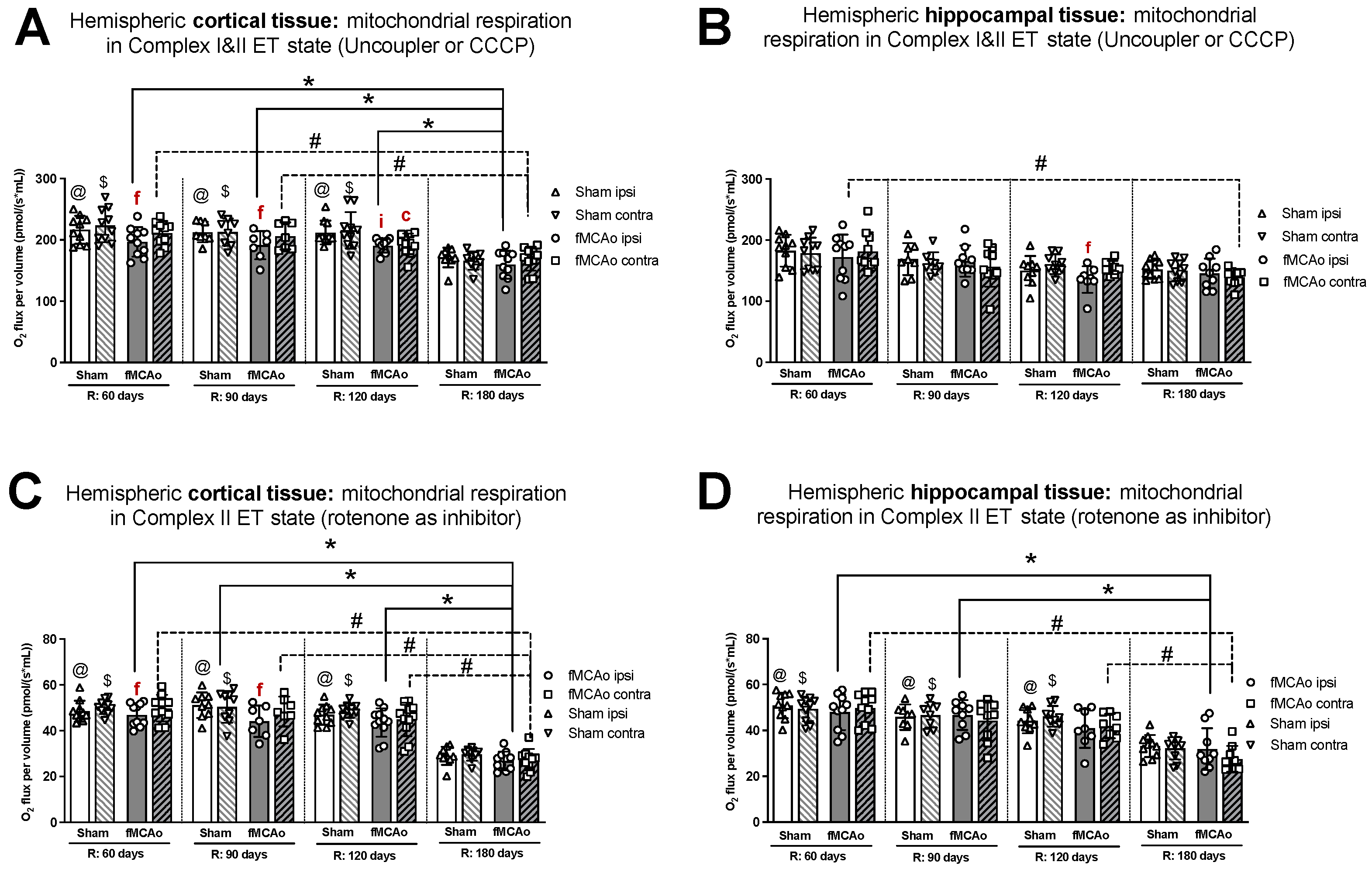
3.1. Mitochondrial Oxygen Consumption
3.1.1. Differences in Tissue Mass-Specific Oxygen Fluxes between Hemispheres in Cortical and Hippocampal Tissue in One Time Period
3.1.2. Differences in Tissue Mass-Specific Oxygen Fluxes between Sham and fMCAo Groups in Hippocampal and Cortical Tissue
3.1.3. Differences in Tissue Mass-Specific Oxygen Fluxes in Cortical Tissue over Time
3.1.4. Differences in Tissue Mass-Specific Oxygen Fluxes in Hippocampal Tissue over Time
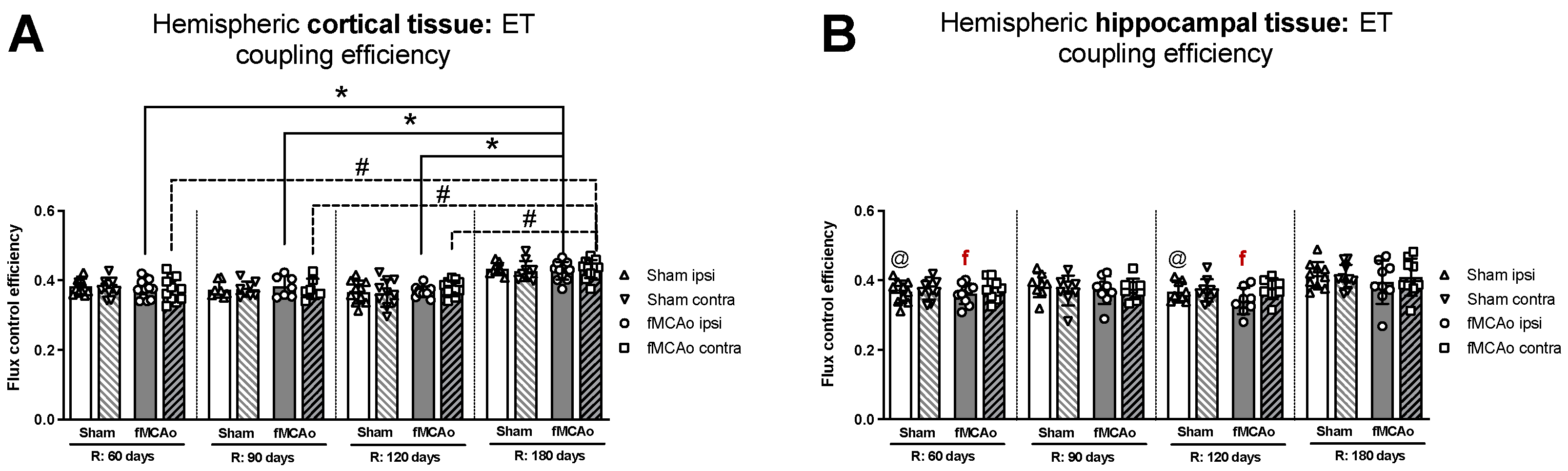
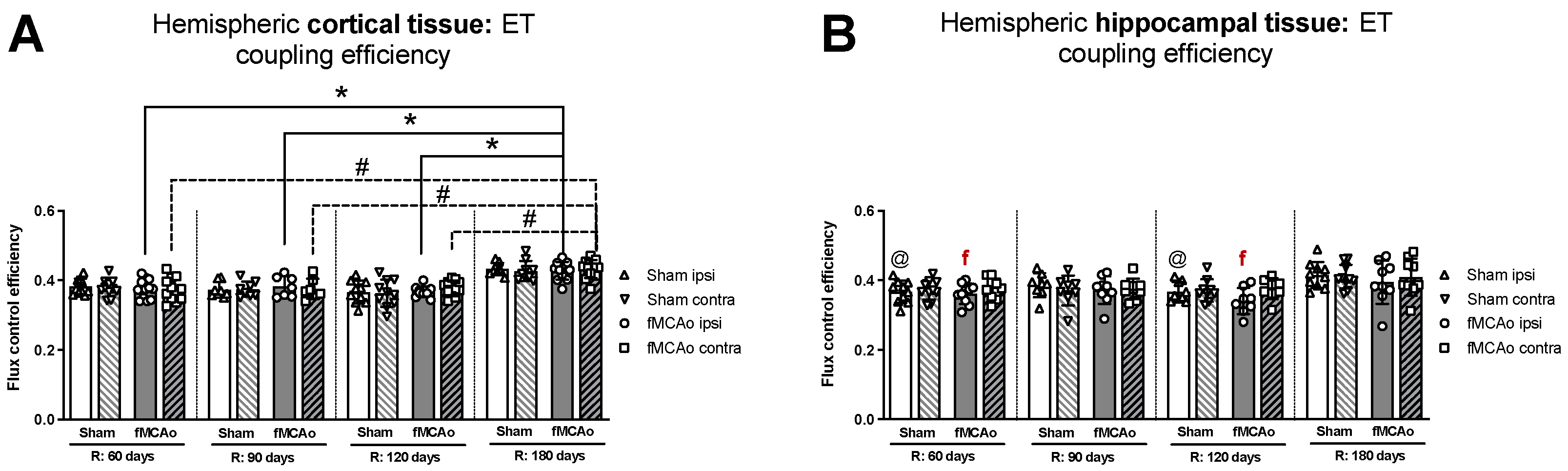
3.1.5. Differences in ET Coupling Efficiency between Hemispheres in Cortical and Hippocampal Tissue
3.1.6. Differences in ET Coupling Efficiency in Cortical and Hippocampal Tissue over Time
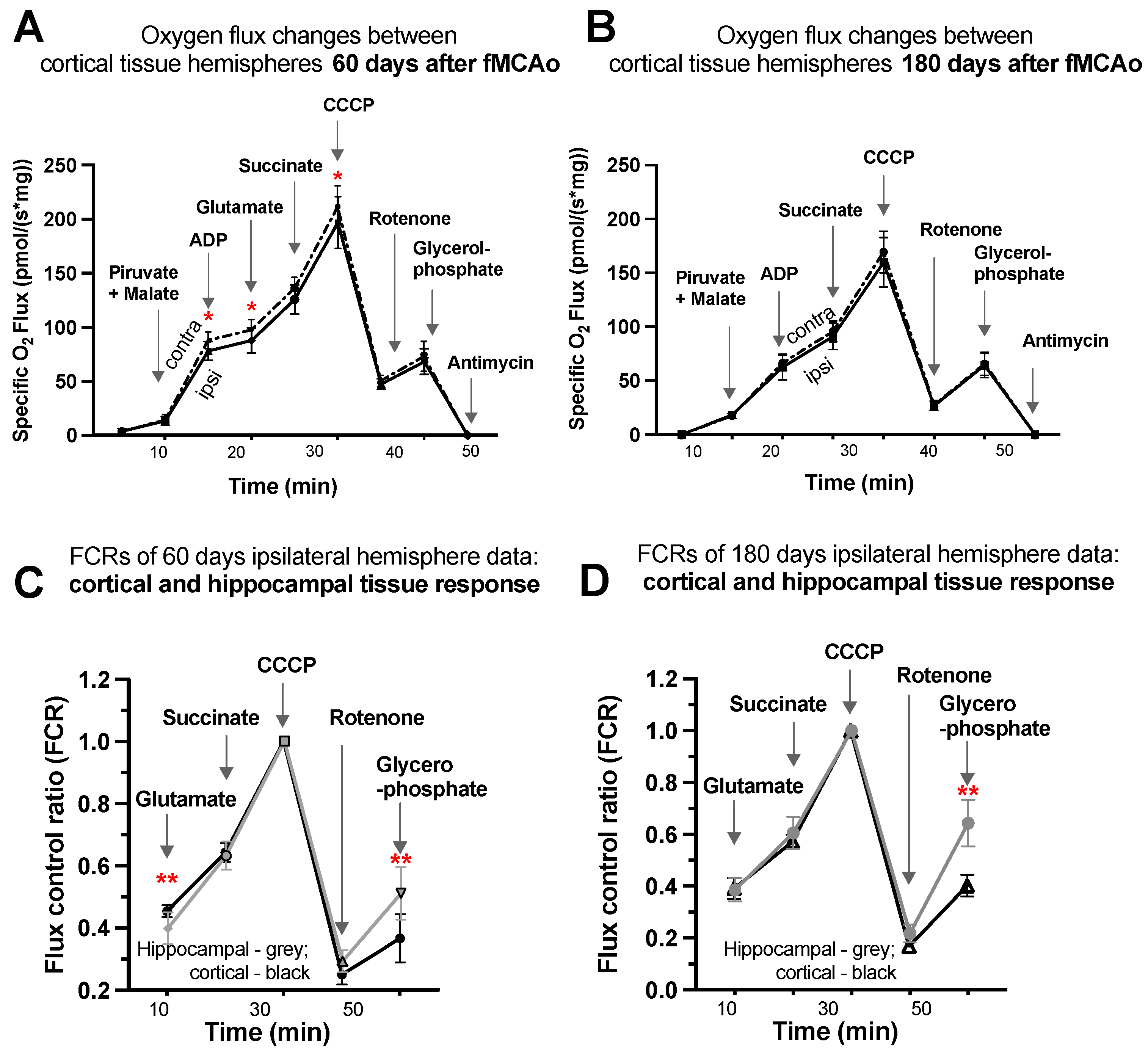
3.1.7. Representative Mitochondrial Measurements following Substrate Additions
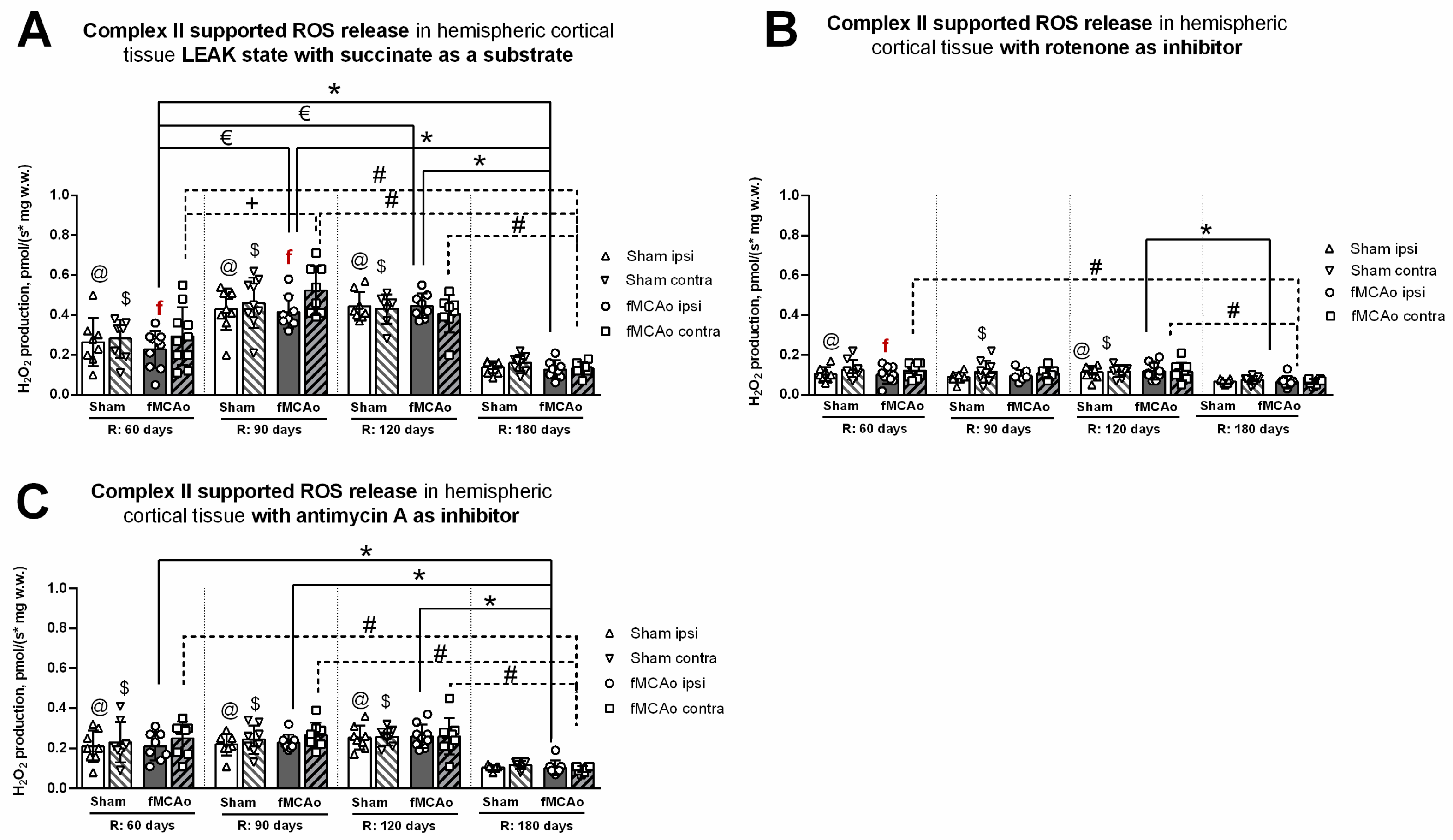
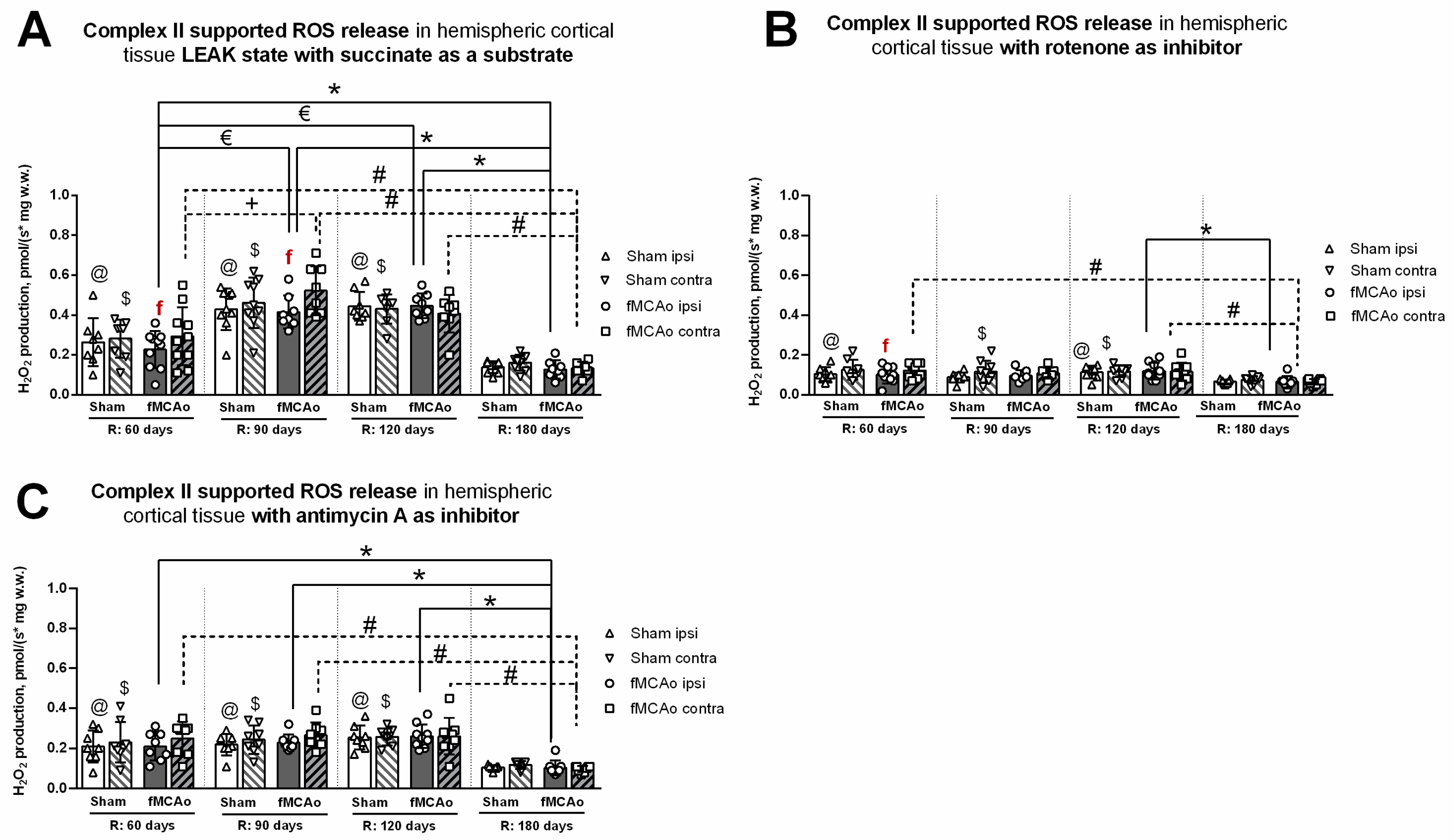
3.2. Mitochondrial ROS Production in Cortical Tissues
3.2.1. Differences in Tissue Mass-Specific H2O2 Emissions between fMCAo Cortical Hemispheres
3.2.2. Differences in Tissue Mass-Specific H2O2 Emissions in Cortical Hemispheres over Time
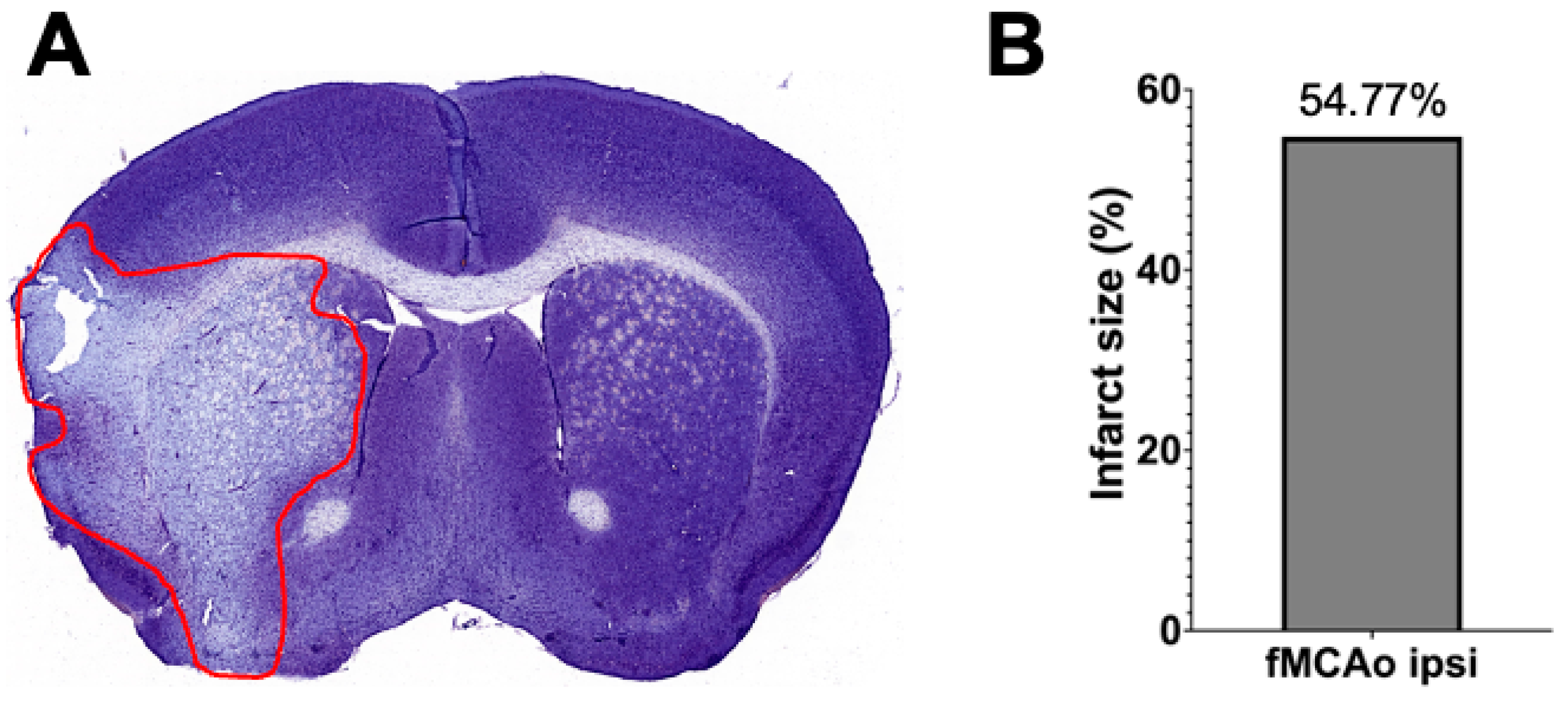
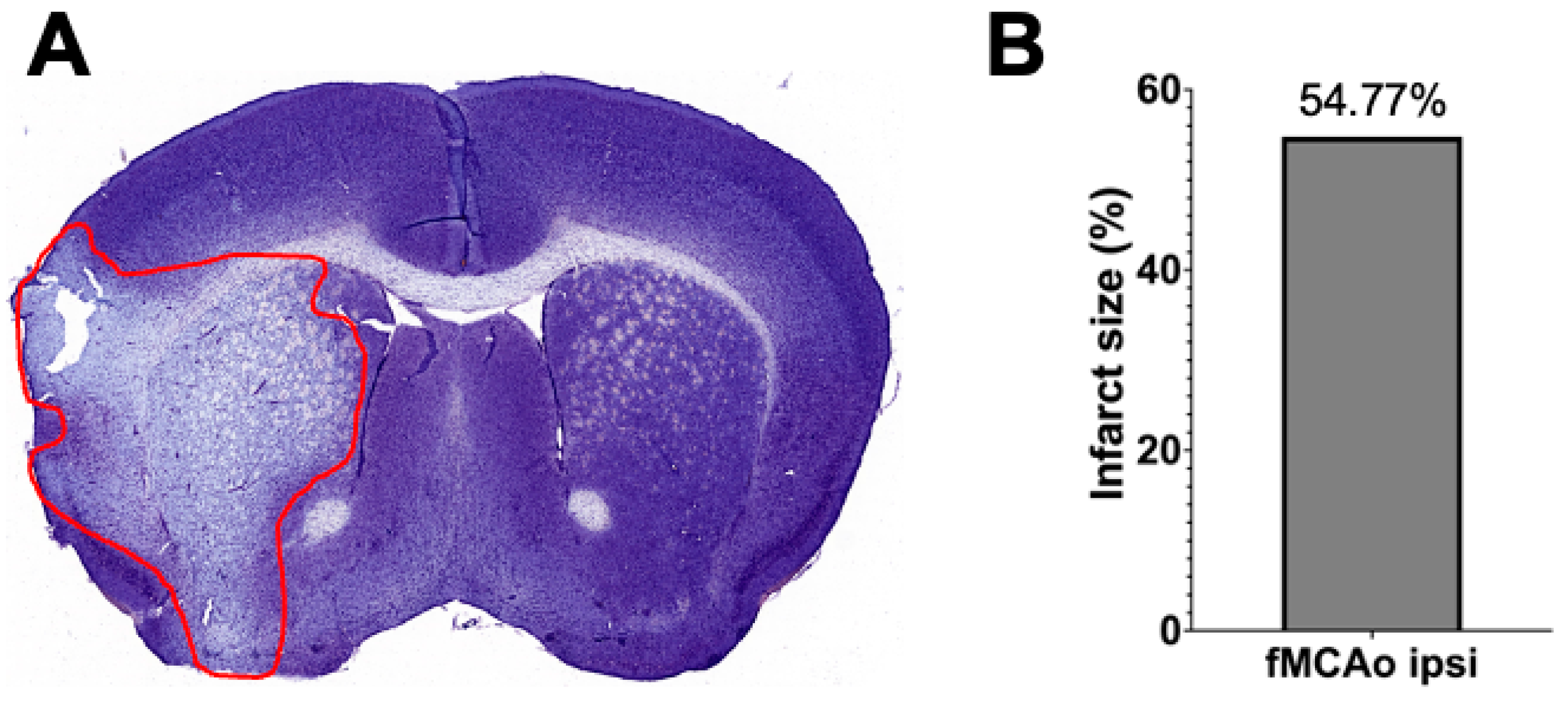
3.3. Representation of Infarct Size following a 60 min Long fMCAo
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Andrabi, S.S.; Parvez, S.; Tabassum, H. Ischemic stroke and mitochondria: Mechanisms and targets. Protoplasma 2020, 257, 335–343. [Google Scholar] [CrossRef]
- Lima, T.; Li, T.Y.; Mottis, A.; Auwerx, J. Pleiotropic effects of mitochondria in aging. Nat. Aging 2022, 2, 199–213. [Google Scholar] [CrossRef] [PubMed]
- Kuznetsov, A.V.; Margreiter, R.; Ausserlechner, M.J.; Hagenbuchner, J. The Complex Interplay between Mitochondria, ROS and Entire Cellular Metabolism. Antioxidants 2022, 11, 1995. [Google Scholar] [CrossRef] [PubMed]
- Chomova, M.; Zitnanova, I. Look into brain energy crisis and membrane pathophysiology in ischemia and reperfusion. Stress. 2016, 19, 341–348. [Google Scholar] [CrossRef] [PubMed]
- Beresewicz-Haller, M.; Krupska, O.; Bochomulski, P.; Dudzik, D.; Checinska, A.; Hilgier, W.; Barbas, C.; Zablocki, K.; Zablocka, B. Mitochondrial Metabolism behind Region-Specific Resistance to Ischemia-Reperfusion Injury in Gerbil Hippocampus. Role of PKCbetaII and Phosphate-Activated Glutaminase. Int. J. Mol. Sci. 2021, 22, 8504. [Google Scholar] [CrossRef] [PubMed]
- Gao, L.; Liu, F.; Hou, P.P.; Manaenko, A.; Xiao, Z.P.; Wang, F.; Xu, T.L.; Hu, Q. Neurons Release Injured Mitochondria as “Help-Me” Signaling After Ischemic Stroke. Front. Aging Neurosci. 2022, 14, 785761. [Google Scholar] [CrossRef] [PubMed]
- Rosen, C.L.; Dinapoli, V.A.; Nagamine, T.; Crocco, T. Influence of age on stroke outcome following transient focal ischemia. J. Neurosurg. 2005, 103, 687–694. [Google Scholar] [CrossRef] [PubMed]
- Li, N.; Kong, X.; Ye, R.; Yang, Q.; Han, J.; Xiong, L. Age-related differences in experimental stroke: Possible involvement of mitochondrial dysfunction and oxidative damage. Rejuvenation Res. 2011, 14, 261–273. [Google Scholar] [CrossRef] [PubMed]
- Qin, C.; Yang, S.; Chu, Y.H.; Zhang, H.; Pang, X.W.; Chen, L.; Zhou, L.Q.; Chen, M.; Tian, D.S.; Wang, W. Signaling pathways involved in ischemic stroke: Molecular mechanisms and therapeutic interventions. Signal Transduct. Target. Ther. 2022, 7, 215. [Google Scholar] [CrossRef] [PubMed]
- Zhu, G.; Wang, X.; Chen, L.; Lenahan, C.; Fu, Z.; Fang, Y.; Yu, W. Crosstalk Between the Oxidative Stress and Glia Cells After Stroke: From Mechanism to Therapies. Front. Immunol. 2022, 13, 852416. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Ma, X.; Yu, W.; Lou, Z.; Mu, D.; Wang, Y.; Shen, B.; Qi, S. Reperfusion promotes mitochondrial dysfunction following focal cerebral ischemia in rats. PLoS ONE 2012, 7, e46498. [Google Scholar] [CrossRef] [PubMed]
- Chouchani, E.T.; Pell, V.R.; Gaude, E.; Aksentijevic, D.; Sundier, S.Y.; Robb, E.L.; Logan, A.; Nadtochiy, S.M.; Ord, E.N.J.; Smith, A.C.; et al. Ischaemic accumulation of succinate controls reperfusion injury through mitochondrial ROS. Nature 2014, 515, 431–435. [Google Scholar] [CrossRef] [PubMed]
- Stepanova, A.; Kahl, A.; Konrad, C.; Ten, V.; Starkov, A.S.; Galkin, A. Reverse electron transfer results in a loss of flavin from mitochondrial complex I: Potential mechanism for brain ischemia reperfusion injury. J. Cereb. Blood Flow. Metab. 2017, 37, 3649–3658. [Google Scholar] [CrossRef] [PubMed]
- Hüttemann, M.; Helling, S.; Sanderson, T.H.; Sinkler, C.; Samavati, L.; Mahapatra, G.; Varughese, A.; Lu, G.; Liu, J.; Ramzan, R.; et al. Regulation of mitochondrial respiration and apoptosis through cell signaling: Cytochrome c oxidase and cytochrome c in ischemia/reperfusion injury and inflammation. Biochim. Biophys. Acta 2012, 1817, 598–609. [Google Scholar] [CrossRef] [PubMed]
- Villa, R.F.; Gorini, A.; Hoyer, S. Effect of ageing and ischemia on enzymatic activities linked to Krebs’ cycle, electron transfer chain, glutamate and aminoacids metabolism of free and intrasynaptic mitochondria of cerebral cortex. Neurochem. Res. 2009, 34, 2102–2116. [Google Scholar] [CrossRef] [PubMed]
- Kuroda, S.; Katsura, K.I.; Tsuchidate, R.; Siesjö, B.K. Secondary bioenergetic failure after transient focal ischaemia is due to mitochondrial injury. Acta Physiol. Scand. 1996, 156, 149–150. [Google Scholar] [CrossRef] [PubMed]
- Anderson, M.F.; Sims, N.R. Mitochondrial respiratory function and cell death in focal cerebral ischemia. J. Neurochem. 1999, 73, 1189–1199. [Google Scholar] [CrossRef] [PubMed]
- Sims, N.R.; Anderson, M.F. Mitochondrial contributions to tissue damage in stroke. Neurochem. Int. 2002, 40, 511–526. [Google Scholar] [CrossRef] [PubMed]
- Olesen, M.A.; Torres, A.K.; Jara, C.; Murphy, M.P.; Tapia-Rojas, C. Premature synaptic mitochondrial dysfunction in the hippocampus during aging contributes to memory loss. Redox Biol. 2020, 34, 101558. [Google Scholar] [CrossRef] [PubMed]
- Racay, P.; Tatarková, Z.; Drgová, A.; Kaplan, P.; Dobrota, D. Ischemia-reperfusion induces inhibition of mitochondrial protein synthesis and cytochrome c oxidase activity in rat hippocampus. Physiol. Res. 2009, 58, 127–138. [Google Scholar] [CrossRef]
- Beresewicz-Haller, M. Hippocampal region-specific endogenous neuroprotection as an approach in the search for new neuroprotective strategies in ischemic stroke. Fiction or fact? Neurochem. Int. 2023, 162, 105455. [Google Scholar] [CrossRef] [PubMed]
- Kawalec, M.; Wojtyniak, P.; Bielska, E.; Lewczuk, A.; Boratynska-Jasinska, A.; Beresewicz-Haller, M.; Frontczak-Baniewicz, M.; Gewartowska, M.; Zablocka, B. Mitochondrial dynamics, elimination and biogenesis during post-ischemic recovery in ischemia-resistant and ischemia-vulnerable gerbil hippocampal regions. Biochim. Biophys. Acta Mol. Basis Dis. 2023, 1869, 166633. [Google Scholar] [CrossRef] [PubMed]
- Bahire, K.L.; Maļuhins, R.; Bello, F.; Freitag, S.V.; Jeļisejevs, I.; Gile, R.; Upīte, J.; Plesnila, N.; Jansone, B. Hemispheric analysis of mitochondrial Complex I and II activity in the mouse model of ischemia-reperfusion-induced injury. Mitochondrion 2023, 69, 147–158. [Google Scholar] [CrossRef] [PubMed]
- Rutkai, I.; Merdzo, I.; Wunnava, S.V.; Curtin, G.T.; Katakam, P.V.; Busija, D.W. Cerebrovascular function and mitochondrial bioenergetics after ischemia-reperfusion in male rats. J. Cereb. Blood Flow. Metab. 2019, 39, 1056–1068. [Google Scholar] [CrossRef] [PubMed]
- Garbuzova-Davis, S.; Haller, E.; Williams, S.N.; Haim, E.D.; Tajiri, N.; Hernandez-Ontiveros, D.G.; Frisina-Deyo, A.; Boffeli, S.M.; Sanberg, P.R.; Borlongan, C.V. Compromised blood-brain barrier competence in remote brain areas in ischemic stroke rats at the chronic stage. J. Comp. Neurol. 2014, 522, 3120–3137. [Google Scholar] [CrossRef] [PubMed]
- Summers, F.A.; Zhao, B.; Ganini, D.; Mason, R.P. Photooxidation of Amplex Red to resorufin: Implications of exposing the Amplex Red assay to light. Methods Enzymol. 2013, 526, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Gröger, M.; Lebesgue, D.; Pruneau, D.; Relton, J.; Kim, S.W.; Nussberger, J.; Plesnila, N. Release of bradykinin and expression of kinin B2 receptors in the brain: Role for cell death and brain edema formation after focal cerebral ischemia in mice. J. Cereb. Blood Flow. Metab. 2005, 25, 978–989. [Google Scholar] [CrossRef] [PubMed]
- Engel, O.; Kolodziej, S.; Dirnagl, U.; Prinz, V. Modeling stroke in mice-middle cerebral artery occlusion with the filament model. J. Vis. Exp. 2011, 47, 1–5. [Google Scholar] [CrossRef]
- Lourbopoulos, A.; Mamrak, U.; Roth, S.; Balbi, M.; Shrouder, J.; Liesz, A.; Hellal, F.; Plesnila, N. Inadequate food and water intake determine mortality following stroke in mice. J. Cereb. Blood Flow. Metab. 2017, 37, 2084–2097. [Google Scholar] [CrossRef]
- Pilipenko, V.; Dzirkale, Z.; Rozkalne, R.; Upite, J.; Hellal, F.; Plesnila, N.; Jansone, B. Focal Cerebral Ischemia Induces Global Subacute Changes in the Number of Neuroblasts and Neurons and the Angiogenic Factor Density in Mice. Medicina 2023, 59, 2168. [Google Scholar] [CrossRef] [PubMed]
- Fernández-Vizarra, E.; Ferrín, G.; Pérez-Martos, A.; Fernández-Silva, P.; Zeviani, M.; Enríquez, J.A. Isolation of mitochondria for biogenetical studies: An update. Mitochondrion 2010, 10, 253–262. [Google Scholar] [CrossRef] [PubMed]
- Gnaiger, E.; Kuznetsov, A.V.; Schneeberger, S.; Seiler, R.; Brandacher, G.; Steurer, W.; Margreiter, R. Mitochondria in the Cold. In Life in the Cold; Springer: Berlin/Heidelberg, Germany, 2000; pp. 431–442. [Google Scholar]
- Gnaiger, E.; Ahn, B.; Alves, M.G.; Amati, F.; Aral, C.; Arandarčikaitė, O.; Frostner, E.Å.; Bailey, D.M.; Silva, A.C.B.S.A.; Battino, M.; et al. Mitochondrial respiratory states and rates: Building blocks of mitochondrial physiology (Part 1). MitoEAGLE Prepr. 2018, 1, 1–35. [Google Scholar] [CrossRef]
- Stepanova, A.; Konrad, C.; Guerrero-Castillo, S.; Manfredi, G.; Vannucci, S.; Arnold, S.; Galkin, A. Deactivation of mitochondrial complex I after hypoxia-ischemia in the immature brain. J. Cereb. Blood Flow. Metab. 2019, 39, 1790–1802. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.; Stepanova, A.; Niatsetskaya, Z.; Sosunov, S.; Arndt, S.; Murphy, M.P.; Galkin, A.; Ten, V.S. Attenuation of oxidative damage by targeting mitochondrial complex I in neonatal hypoxic-ischemic brain injury. Free Radic. Biol. Med. 2018, 124, 517–524. [Google Scholar] [CrossRef] [PubMed]
- Burtscher, J.; Zangrandi, L.; Schwarzer, C.; Gnaiger, E. Differences in mitochondrial function in homogenated samples from healthy and epileptic specific brain tissues revealed by high-resolution respirometry. Mitochondrion 2015, 25, 104–112. [Google Scholar] [CrossRef] [PubMed]
- Pecinová, A.; Drahota, Z.; Nůsková, H.; Pecina, P.; Houštěk, J. Evaluation of basic mitochondrial functions using rat tissue homogenates. Mitochondrion 2011, 11, 722–728. [Google Scholar] [CrossRef] [PubMed]
- Gnaiger, E. Mitochondrial pathways and respiratory control. An introduction to OXPHOS analysis. Bioenerg. Commun. 2020, 2020, 2. [Google Scholar] [CrossRef]
- Butler, T.L.; Kassed, C.A.; Sanberg, P.R.; Willing, A.E.; Pennypacker, K.R. Neurodegeneration in the rat hippocampus and striatum after middle cerebral artery occlusion. Brain Res. 2002, 929, 252–260. [Google Scholar] [CrossRef] [PubMed]
- Ronnback, A.; Dahlqvist, P.; Svensson, P.A.; Jernas, M.; Carlsson, B.; Carlsson, L.M.; Olsson, T. Gene expression profiling of the rat hippocampus one month after focal cerebral ischemia followed by enriched environment. Neurosci. Lett. 2005, 385, 173–178. [Google Scholar] [CrossRef] [PubMed]
- Du, J.; Ma, M.; Zhao, Q.; Fang, L.; Chang, J.; Wang, Y.; Fei, R.; Song, X. Mitochondrial bioenergetic deficits in the hippocampi of rats with chronic ischemia-induced vascular dementia. Neuroscience 2013, 231, 345–352. [Google Scholar] [CrossRef] [PubMed]
- Schaapsmeerders, P.; van Uden, I.W.; Tuladhar, A.M.; Maaijwee, N.A.; van Dijk, E.J.; Rutten-Jacobs, L.C.; Arntz, R.M.; Schoonderwaldt, H.C.; Dorresteijn, L.D.; de Leeuw, F.E.; et al. Ipsilateral hippocampal atrophy is associated with long-term memory dysfunction after ischemic stroke in young adults. Hum. Brain Mapp. 2015, 36, 2432–2442. [Google Scholar] [CrossRef] [PubMed]
- Kula-Alwar, D.; Prag, H.A.; Krieg, T. Targeting Succinate Metabolism in Ischemia/Reperfusion Injury. Circulation 2019, 140, 1968–1970. [Google Scholar] [CrossRef] [PubMed]
- Korshunov, S.S.; Skulachev, V.P.; Starkov, A.A. High protonic potential actuates a mechanism of production of reactive oxygen species in mitochondria. FEBS Lett. 1997, 416, 15–18. [Google Scholar] [CrossRef] [PubMed]
- Ferrari, F.; Gorini, A.; Hoyer, S.; Villa, R.F. Glutamate metabolism in cerebral mitochondria after ischemia and post-ischemic recovery during aging: Relationships with brain energy metabolism. J. Neurochem. 2018, 146, 416–428. [Google Scholar] [CrossRef] [PubMed]
- Ketheeswaranathan, P.; Turner, N.A.; Spary, E.J.; Batten, T.F.; McColl, B.W.; Saha, S. Changes in glutamate transporter expression in mouse forebrain areas following focal ischemia. Brain Res. 2011, 1418, 93–103. [Google Scholar] [CrossRef] [PubMed]
- Galkin, A. Brain Ischemia/Reperfusion Injury and Mitochondrial Complex I Damage. Biochemistry 2019, 84, 1411–1423. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, N.H.; Gonzalez, S.V.; Hassel, B. Formation of glycerol from glucose in rat brain and cultured brain cells. Augmentation with kainate or ischemia. J. Neurochem. 2007, 101, 1694–1700. [Google Scholar] [CrossRef] [PubMed]
- Gaignard, P.; Liere, P.; Thérond, P.; Schumacher, M.; Slama, A.; Guennoun, R. Role of Sex Hormones on Brain Mitochondrial Function, with Special Reference to Aging and Neurodegenerative Diseases. Front. Aging Neurosci. 2017, 9, 406. [Google Scholar] [CrossRef]
- Gaignard, P.; Savouroux, S.; Liere, P.; Pianos, A.; Thérond, P.; Schumacher, M.; Slama, A.; Guennoun, R. Effect of Sex Differences on Brain Mitochondrial Function and Its Suppression by Ovariectomy and in Aged Mice. Endocrinology 2015, 156, 2893–2904. [Google Scholar] [CrossRef] [PubMed]
- Khalifa, A.R.; Abdel-Rahman, E.A.; Mahmoud, A.M.; Ali, M.H.; Noureldin, M.; Saber, S.H.; Mohsen, M.; Ali, S.S. Sex-specific differences in mitochondria biogenesis, morphology, respiratory function, and ROS homeostasis in young mouse heart and brain. Physiol. Rep. 2017, 5, 1–19. [Google Scholar] [CrossRef]
- Cui, H.; Kong, Y.; Zhang, H. Oxidative stress, mitochondrial dysfunction, and aging. J. Signal Transduct. 2012, 2012, 646354. [Google Scholar] [CrossRef] [PubMed]
- Giorgi, C.; Marchi, S.; Simoes, I.C.M.; Ren, Z.; Morciano, G.; Perrone, M.; Patalas-Krawczyk, P.; Borchard, S.; Jędrak, P.; Pierzynowska, K.; et al. Mitochondria and Reactive Oxygen Species in Aging and Age-Related Diseases. Int. Rev. Cell Mol. Biol. 2018, 340, 209–344. [Google Scholar] [CrossRef] [PubMed]
- Lores-Arnaiz, S.; Lombardi, P.; Karadayian, A.G.; Orgambide, F.; Cicerchia, D.; Bustamante, J. Brain cortex mitochondrial bioenergetics in synaptosomes and non-synaptic mitochondria during aging. Neurochem. Res. 2016, 41, 353–363. [Google Scholar] [CrossRef]
- Pandya, J.D.; Royland, J.E.; MacPhail, R.C.; Sullivan, P.G.; Kodavanti, P.R. Age- and brain region-specific differences in mitochondrial bioenergetics in Brown Norway rats. Neurobiol. Aging 2016, 42, 25–34. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment Group | Hemisphere | Measurement Day (R) | Induced Respiratory State | |||
|---|---|---|---|---|---|---|
| CI OXPHOS (Figure 1A) | CI&II OXPHOS (Figure 1C) | CI&II ET (Figure 2A) | CII ET (Figure 2C) | |||
| Stroke Group | Ipsilateral | 60 | * p < 0.0001 | * p < 0.0001 | * p = 0.0044 | * p < 0.0001 |
| 90 | * p = 0.0037 | * p = 0.0018 | * p = 0.0110 | * p < 0.0001 | ||
| 120 | * p = 0.0003 | * p = 0.0001 | * p = 0.0214 | * p < 0.0001 | ||
| Contralateral | 60 | # p = 0.0002 | #p < 0.0001 | #p = 0.0003 | # p < 0.0001 | |
| 90 | # p = 0.0023 | # p = 0.0010 | # p = 0.0206 | # p < 0.0001 | ||
| 120 | # p = 0.0306 | # p = 0.0116 | - | # p < 0.0001 | ||
| Sham Group | Ipsilateral | 60 | @ p < 0.0001 | @ p < 0.0001 | @ p = 0.0006 | @ p < 0.0001 |
| 90 | @ p < 0.0001 | @ p < 0.0001 | @ p = 0.0216 | @ p < 0.0001 | ||
| 120 | @ p < 0.0001 | @ p < 0.0001 | @ p = 0.0046 | @ p < 0.0001 | ||
| Contralateral | 60 | - | $ p < 0.0001 | $ p < 0.0001 | $ p < 0.0001 | |
| 90 | - | $ p < 0.0001 | $ p = 0.0391 | $ p < 0.0001 | ||
| 120 | - | $ p < 0.0001 | $ p = 0.0012 | $ p < 0.0001 | ||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bahire, K.L.; Maļuhins, R.; Bello, F.; Upīte, J.; Makarovs, A.; Jansone, B. Long-Term Region-Specific Mitochondrial Functionality Changes in Both Cerebral Hemispheres after fMCAo Model of Ischemic Stroke. Antioxidants 2024, 13, 416. https://doi.org/10.3390/antiox13040416
Bahire KL, Maļuhins R, Bello F, Upīte J, Makarovs A, Jansone B. Long-Term Region-Specific Mitochondrial Functionality Changes in Both Cerebral Hemispheres after fMCAo Model of Ischemic Stroke. Antioxidants. 2024; 13(4):416. https://doi.org/10.3390/antiox13040416
Chicago/Turabian StyleBahire, Ksenija Lūcija, Reinis Maļuhins, Fiona Bello, Jolanta Upīte, Aleksandrs Makarovs, and Baiba Jansone. 2024. "Long-Term Region-Specific Mitochondrial Functionality Changes in Both Cerebral Hemispheres after fMCAo Model of Ischemic Stroke" Antioxidants 13, no. 4: 416. https://doi.org/10.3390/antiox13040416
APA StyleBahire, K. L., Maļuhins, R., Bello, F., Upīte, J., Makarovs, A., & Jansone, B. (2024). Long-Term Region-Specific Mitochondrial Functionality Changes in Both Cerebral Hemispheres after fMCAo Model of Ischemic Stroke. Antioxidants, 13(4), 416. https://doi.org/10.3390/antiox13040416