
The Impact of Oxidative Stress on the Epigenetics of Fetal Alcohol Spectrum Disorders

 , ,
, ,  ,
,  ,
,  and
and 
Abstract
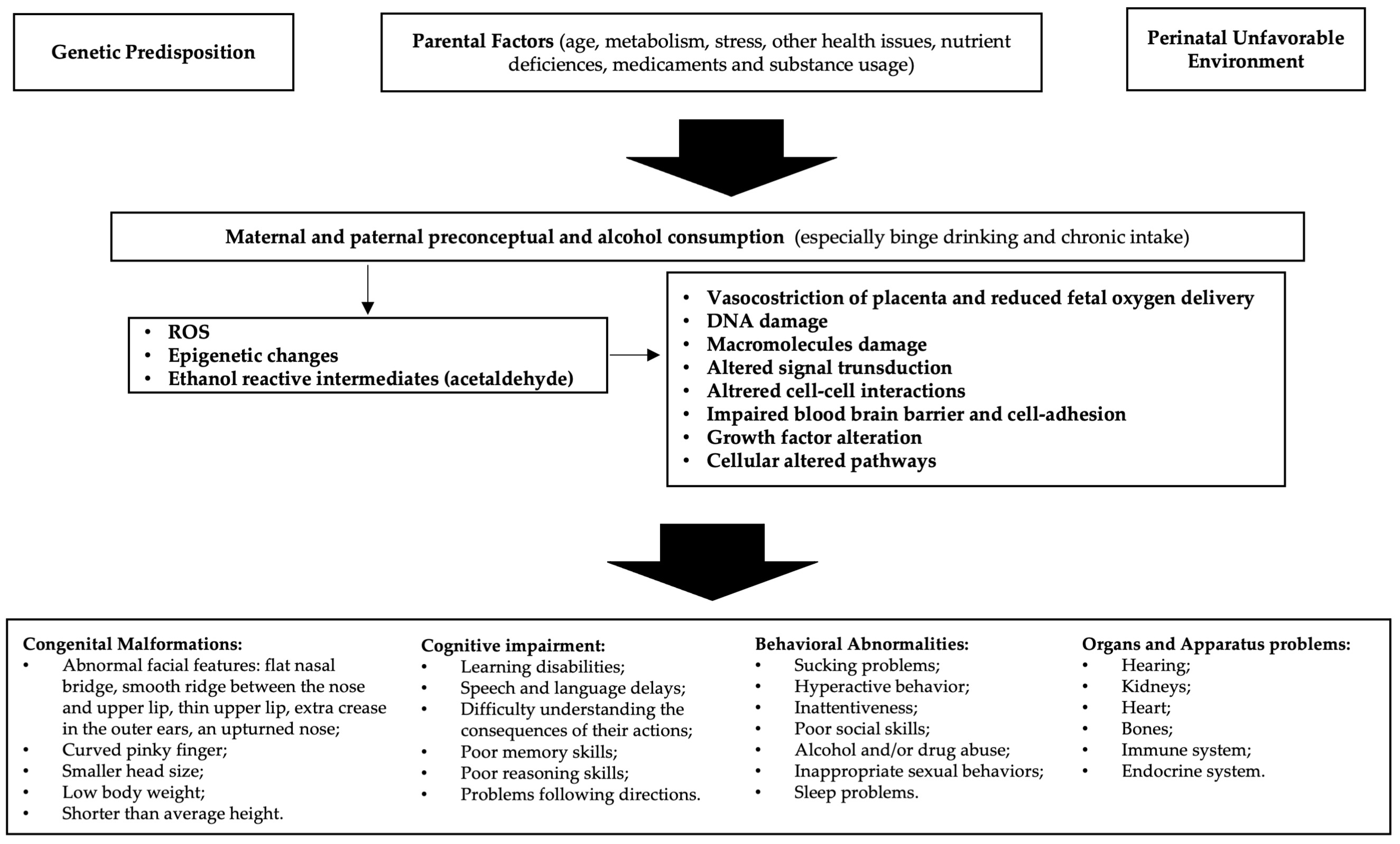
1. Fetal Alcohol Spectrum Disorders (FASD)
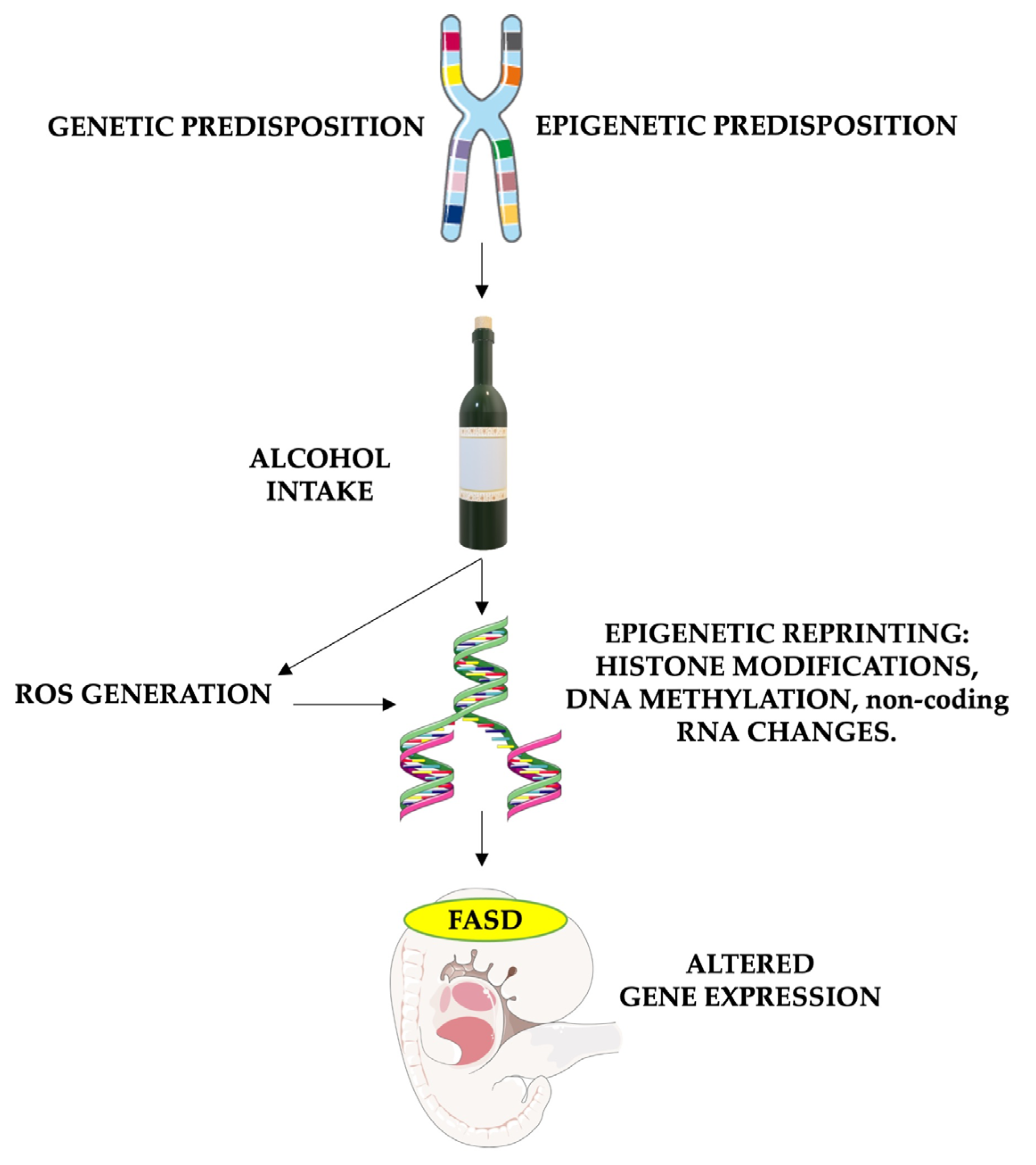
2. FASD Epigenetics
3. Oxidative Stress and FASD
4. Epigenetics and Oxidative Stress
5. FASD Current Treatment
6. Therapeutic Implications
7. Discussion
8. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Mattson, S.N.; Crocker, N.; Nguyen, T.T. Fetal alcohol spectrum disorders: Neuropsychological and behavioral features. Neuropsychol. Rev. 2011, 21, 81–101. [Google Scholar] [CrossRef] [PubMed]
- Greenmyer, J.R.; Popova, S.; Klug, M.G.; Burd, L. Fetal alcohol spectrum disorder: A systematic review of the cost of and savings from prevention in the United States and Canada. Addiction 2020, 115, 409–417. [Google Scholar] [CrossRef] [PubMed]
- Vorgias, D.; Bernstein, B. Fetal Alcohol Syndrome; StatPearls Publishing: Treasure Island, FL, USA, 2021. [Google Scholar]
- Terracina, S.; Ferraguti, G.; Tarani, L.; Messina, M.P.; Lucarelli, M.; Vitali, M.; De Persis, S.; Greco, A.; Minni, A.; Polimeni, A.; et al. Transgenerational Abnormalities Induced by Paternal Preconceptual Alcohol Drinking. Findings from Humans and Animal Models. Curr. Neuropharmacol. 2021, 19, 1158–1173. [Google Scholar] [CrossRef] [PubMed]
- Kalberg, W.O.; Buckley, D. FASD: What types of intervention and rehabilitation are useful? Neurosci. Biobehav. Rev. 2007, 31, 278–285. [Google Scholar] [CrossRef]
- Center for Disease Control and Prevention. Data & Statistics: Prevalence of FASDs. 2023. Available online: https://www.cdc.gov/ncbddd/fasd/data.html (accessed on 21 March 2024).
- Haycock, P.C. Fetal alcohol spectrum disorders: The epigenetic perspective. Biol. Reprod. 2009, 81, 607–617. [Google Scholar] [CrossRef]
- Ciafrè, S.; Ferraguti, G.; Greco, A.; Polimeni, A.; Ralli, M.; Ceci, F.M.; Ceccanti, M.; Fiore, M. Alcohol as an early life stressor: Epigenetics, metabolic, neuroendocrine and neurobehavioral implications. Neurosci. Biobehav. Rev. 2020, 118, 654–668. [Google Scholar] [CrossRef] [PubMed]
- Popova, S.; Lange, S.; Shield, K.; Mihic, A.; Chudley, A.E.; Mukherjee, R.A.S.; Bekmuradov, D.; Rehm, J. Comorbidity of fetal alcohol spectrum disorder: A systematic review and meta-analysis. Lancet 2016, 387, 978–987. [Google Scholar] [CrossRef] [PubMed]
- Rasmussen, C.; Andrew, G.; Zwaigenbaum, L.; Tough, S. Neurobehavioural outcomes of children with fetal alcohol spectrum disorders: A Canadian perspective. Paediatr. Child. Health 2008, 13, 185–191. [Google Scholar] [PubMed]
- Mead, E.A.; Sarkar, D.K. Fetal alcohol spectrum disorders and their transmission through genetic and epigenetic mechanisms. Front. Genet. 2014, 5, 154. [Google Scholar] [CrossRef]
- Burd, L.; Blair, J.; Dropps, K. Prenatal alcohol exposure, blood alcohol concentrations and alcohol elimination rates for the mother, fetus and newborn. J. Perinatol. 2012, 32, 652–659. [Google Scholar] [CrossRef]
- Keen, C.L.; Uriu-Adams, J.Y.; Skalny, A.; Grabeklis, A.; Grabeklis, S.; Green, K.; Yevtushok, L.; Wertelecki, W.W.; Chambers, C.D. The plausibility of maternal nutritional status being a contributing factor to the risk for fetal alcohol spectrum disorders: The potential influence of zinc status as an example. BioFactors 2010, 36, 125–135. [Google Scholar] [CrossRef]
- King, J.C.; Fabro, S. Alcohol consumption and cigarette smoking: Effect on pregnancy. Clin. Obstet. Gynecol. 1983, 26, 437–448. [Google Scholar] [CrossRef]
- Gupta, K.K.; Gupta, V.K.; Shirasaka, T. An Update on Fetal Alcohol Syndrome—Pathogenesis, Risks, and Treatment. Alcohol. Clin. Exp. Res. 2016, 40, 1594–1602. [Google Scholar] [CrossRef]
- Miller, L.; Shapiro, A.M.; Wells, P.G. Embryonic catalase protects against ethanol-initiated DNA oxidation and teratogenesis in acatalasemic and transgenic human catalase-expressing mice. Toxicol. Sci. 2013, 134, 400–411. [Google Scholar] [CrossRef]
- McCarver, D.G.; Thomasson, H.R.; Martier, S.S.; Sokol, R.J.; Li, T. Alcohol dehydrogenase-2*3 allele protects against alcohol-related birth defects among African Americans. J. Pharmacol. Exp. Ther. 1997, 283, 1095–1101. [Google Scholar]
- Tunc-Ozcan, E.; Sittig, L.J.; Harper, K.M.; Graf, E.N.; Redei, E.E. Hypothesis: Genetic and epigenetic risk factors interact to modulate vulnerability and resilience to FASD. Front. Genet. 2014, 5, 261. [Google Scholar] [CrossRef]
- Popova, S.; Charness, M.E.; Burd, L.; Crawford, A.; Hoyme, H.E.; Mukherjee, R.A.S.; Riley, E.P.; Elliott, E.J. Fetal alcohol spectrum disorders. Nat. Rev. Dis. Prim. 2023, 9, 11. [Google Scholar] [CrossRef]
- Breton-Larrivée, M.; Elder, E.; Legault, L.; Langford-Avelar, A.; MacFarlane, A.J.; McGraw, S. Mitigating the detrimental developmental impact of early fetal alcohol exposure using a maternal methyl donor-enriched diet. FASEB J. 2023, 37, e22829. [Google Scholar] [CrossRef]
- Ceccanti, M.; Hamilton, D.; Coriale, G.; Carito, V.; Aloe, L.; Chaldakov, G.; Romeo, M.; Ceccanti, M.; Iannitelli, A.; Fiore, M. Spatial learning in men undergoing alcohol detoxification. Physiol. Behav. 2015, 149, 324–330. [Google Scholar] [CrossRef]
- Novick Brown, N.; Greenspan, S. Diminished culpability in fetal alcohol spectrum disorders (FASD). Behav. Sci. Law. 2022, 40, 1–13. [Google Scholar] [CrossRef]
- Ismail, S.; Buckley, S.; Budacki, R.; Jabbar, A.; Gallicano, G.I. Screening, Diagnosing and Prevention of Fetal Alcohol Syndrome: Is This Syndrome Treatable. Dev. Neurosci. 2010, 32, 91–100. [Google Scholar] [CrossRef]
- Fuso, A.; Lucarelli, M. CpG and non-CpG methylation in the diet-epigenetics-neurodegeneration connection. Curr. Nutr. Rep. 2019, 8, 74–82. [Google Scholar] [CrossRef]
- Nicolia, V.; Lucarelli, M.; Fuso, A. Environment, epigenetics and neurodegeneration: Focus on nutrition in Alzheimer’s disease. Exp. Gerontol. 2015, 68, 8–12. [Google Scholar] [CrossRef]
- Crouch, J.; Shvedova, M.; Thanapaul, R.J.R.S.; Botchkarev, V.; Roh, D. Epigenetic Regulation of Cellular Senescence. Cells 2022, 11, 672. [Google Scholar] [CrossRef]
- Li, Y. Modern epigenetics methods in biological research. Methods 2021, 187, 104–113. [Google Scholar] [CrossRef]
- Fuso, A.; Ferraguti, G.; Grandoni, F.; Ruggeri, R.; Scarpa, S.; Strom, R.; Lucarelli, M. Early demethylation of non-CpG, CpC-rich, elements in the myogenin 5′-flanking region: A priming effect on the spreading of active demethylation? Cell Cycle 2010, 9, 3965–3976. [Google Scholar] [CrossRef]
- Mitro, N.; Verdeguer, F.; Perissi, V. Editorial: Epigenetics and metabolism. Front. Endocrinol. (Lausanne) 2024, 15, 1373368. [Google Scholar] [CrossRef]
- Smith, I.M.; Mydlarz, W.K.; Mithani, S.K.; Califano, J.A. DNA global hypomethylation in squamous cell head and neck cancer associated with smoking, alcohol consumption and stage. Int. J. Cancer 2007, 121, 1724–1728. [Google Scholar] [CrossRef]
- Finegersh, A.; Rompala, G.R.; Martin, D.I.; Homanics, G.E. Drinking beyond a lifetime: New and emerging insights into paternal alcohol exposure on subsequent generations. Alcohol 2015, 49, 461–470. [Google Scholar] [CrossRef]
- Raia, T.; Armeli, F.; Cavallaro, R.A.; Ferraguti, G.; Businaro, R.; Lucarelli, M.; Fuso, A. Perinatal S-Adenosylmethionine Supplementation Represses PSEN1 Expression by the Cellular Epigenetic Memory of CpG and Non-CpG Methylation in Adult TgCRD8 Mice. Int. J. Mol. Sci. 2023, 24, 11675. [Google Scholar] [CrossRef]
- Sarkar, D.K. Male germline transmits fetal alcohol epigenetic marks for multiple generations: A review. Addict. Biol. 2016, 21, 23–34. [Google Scholar] [CrossRef]
- Monti, N.; Cavallaro, R.A.; Stoccoro, A.; Nicolia, V.; Scarpa, S.; Kovacs, G.G.; Fiorenza, M.T.; Lucarelli, M.; Aronica, E.; Ferrer, I.; et al. CpG and non-CpG Presenilin1 methylation pattern in course of neurodevelopment and neurodegeneration is associated with gene expression in human and murine brain. Epigenetics 2020, 15, 781–799. [Google Scholar] [CrossRef] [PubMed]
- Pierandrei, S.; Truglio, G.; Ceci, F.; Del Porto, P.; Bruno, S.M.; Castellani, S.; Conese, M.; Ascenzioni, F.; Lucarelli, M. Dna methylation patterns correlate with the expression of scnn1a, scnn1b, and scnn1g (Epithelial sodium channel, enac) genes. Int. J. Mol. Sci. 2021, 22, 3754. [Google Scholar] [CrossRef]
- Ho, S.M.; Johnson, A.; Tarapore, P.; Janakiram, V.; Zhang, X.; Leung, Y.K. Environmental epigenetics and its implication on disease risk and health outcomes. ILAR J. 2012, 53, 289–305. [Google Scholar] [CrossRef]
- Blaconà, G.; Raso, R.; Castellani, S.; Pierandrei, S.; Del Porto, P.; Ferraguti, G.; Ascenzioni, F.; Conese, M.; Lucarelli, M. Downregulation of epithelial sodium channel (ENaC) activity in cystic fibrosis cells by epigenetic targeting. Cell. Mol. Life Sci. 2022, 79, 257. [Google Scholar] [CrossRef] [PubMed]
- Lomberk, G.A.; Iovanna, J.; Urrutia, R. The promise of epigenomic therapeutics in pancreatic cancer. Epigenomics 2016, 8, 831–842. [Google Scholar] [CrossRef]
- Santaló, J.; Berdasco, M. Ethical implications of epigenetics in the era of personalized medicine. Clin. Epigenetics 2022, 14, 44. [Google Scholar] [CrossRef] [PubMed]
- Ramsay, M. Genetic and epigenetic insights into fetal alcohol spectrum disorders. Genome Med. 2010, 2, 27. [Google Scholar] [CrossRef] [PubMed]
- Basavarajappa, B.S. Epigenetics in fetal alcohol spectrum disorder. Prog. Mol. Biol. Transl. Sci. 2023, 197, 211–239. [Google Scholar] [CrossRef]
- Legault, L.M.; Bertrand-Lehouillier, V.; McGraw, S. Pre-implantation alcohol exposure and developmental programming of FASD: An epigenetic perspective. Biochem. Cell Biol. 2018, 96, 117–130. [Google Scholar] [CrossRef]
- Basavarajappa, B.S.; Subbanna, S. Epigenetic mechanisms in developmental alcohol-induced neurobehavioral deficits. Brain Sci. 2016, 6, 12. [Google Scholar] [CrossRef]
- Kaminen-Ahola, N. Fetal alcohol spectrum disorders: Genetic and epigenetic mechanisms. Prenat. Diagn. 2020, 40, 1185–1192. [Google Scholar] [CrossRef]
- Mahnke, A.H.; Miranda, R.C.; Homanics, G.E. Epigenetic mediators and consequences of excessive alcohol consumption. Alcohol 2017, 60, 1–6. [Google Scholar] [CrossRef]
- Okazaki, S.; Otsuka, I.; Shinko, Y.; Horai, T.; Hirata, T.; Yamaki, N.; Sora, I.; Hishimoto, A. Epigenetic Clock Analysis in Children with Fetal Alcohol Spectrum Disorder. Alcohol. Clin. Exp. Res. 2021, 45, 329–337. [Google Scholar] [CrossRef]
- Liyanage, V.; Curtis, K.; Zachariah, R.; Chudley, A.; Rastegar, M. Overview of the Genetic Basis and Epigenetic Mechanisms that Contribute to FASD Pathobiology. Curr. Top. Med. Chem. 2016, 17, 808–828. [Google Scholar] [CrossRef]
- Ungerer, M.; Knezovich, J.; Ramsay, M. In utero alcohol exposure, epigenetic changes, and their consequences. Alcohol. Res. Curr. Rev. 2013, 35, 37–46. [Google Scholar]
- Oei, J.L. Alcohol use in pregnancy and its impact on the mother and child. Addiction 2020, 115, 2148–2163. [Google Scholar] [CrossRef]
- Ouko, L.A.; Shantikumar, K.; Knezovich, J.; Haycock, P.; Schnugh, D.J.; Ramsay, M. Effect of alcohol consumption on CpG methylation in the differentially methylated regions of H19 and IG-DMR in male gametes—Implications for fetal alcohol spectrum disorders. Alcohol. Clin. Exp. Res. 2009, 33, 1615–1627. [Google Scholar] [CrossRef]
- Chang, R.C.; Skiles, W.M.; Chronister, S.S.; Wang, H.; Sutton, G.I.; Bedi, Y.S.; Snyder, M.; Long, C.R.; Golding, M.C. DNA methylation-independent growth restriction and altered developmental programming in a mouse model of preconception male alcohol exposure. Epigenetics 2017, 12, 841–853. [Google Scholar] [CrossRef]
- Haycock, P.C.; Ramsay, M. Exposure of mouse embryos to ethanol during preimplantation development: Effect on DNA methylation in the h19 imprinting control region. Biol. Reprod. 2009, 81, 618–627. [Google Scholar] [CrossRef]
- Jasiulionis, M.G. Abnormal epigenetic regulation of immune system during aging. Front. Immunol. 2018, 9, 197. [Google Scholar] [CrossRef] [PubMed]
- Lussier, A.A.; Bodnar, T.S.; Weinberg, J. Intersection of Epigenetic and Immune Alterations: Implications for Fetal Alcohol Spectrum Disorder and Mental Health. Front. Neurosci. 2021, 15, 788630. [Google Scholar] [CrossRef] [PubMed]
- Rompala, G.R.; Mounier, A.; Wolfe, C.M.; Lin, Q.; Lefterov, I.; Homanics, G.E. Heavy chronic intermittent ethanol exposure alters small noncoding RNAs in mouse sperm and epididymosomes. Front. Genet. 2018, 9, 32. [Google Scholar] [CrossRef] [PubMed]
- Bedi, Y.; Chang, R.C.; Gibbs, R.; Clement, T.M.; Golding, M.C. Alterations in sperm-inherited noncoding RNAs associate with late-term fetal growth restriction induced by preconception paternal alcohol use. Reprod. Toxicol. 2019, 87, 11–20. [Google Scholar] [CrossRef] [PubMed]
- Fuso, A.; Raia, T.; Orticello, M.; Lucarelli, M. The complex interplay between DNA methylation and miRNAs in gene expression regulation. Biochimie 2020, 173, 12–16. [Google Scholar] [CrossRef] [PubMed]
- Roozen, S.; Ehrhart, F. Fetal alcohol spectrum disorders and the risk of crime. Handb. Clin. Neurol. 2023, 197, 197–204. [Google Scholar]
- Wentzel, P.; Rydberg, U.; Eriksson, U.J. Antioxidative treatment diminishes ethanol-induced congenital malformations in the rat. Alcohol. Clin. Exp. Res. 2006, 30, 1752–1760. [Google Scholar] [CrossRef] [PubMed]
- Derme, M.; Piccioni, M.G.; Brunelli, R.; Crognale, A.; Denotti, M.; Ciolli, P.; Scomparin, D.; Tarani, L.; Paparella, R.; Terrin, G.; et al. Oxidative Stress in a Mother Consuming Alcohol during Pregnancy and in Her Newborn: A Case Report. Antioxidants 2023, 12, 1216. [Google Scholar] [CrossRef] [PubMed]
- Tossetta, G.; Fantone, S.; Piani, F.; Crescimanno, C.; Ciavattini, A.; Giannubilo, S.R.; Marzioni, D. Modulation of NRF2/KEAP1 Signaling in Preeclampsia. Cells 2023, 12, 1545. [Google Scholar] [CrossRef]
- Auvinen, P.; Vehviläinen, J.; Marjonen, H.; Modhukur, V.; Sokka, J.; Wallén, E.; Rämö, K.; Ahola, L.; Salumets, A.; Otonkoski, T.; et al. Chromatin modifier developmental pluripotency associated factor 4 (DPPA4) is a candidate gene for alcohol-induced developmental disorders. BMC Med. 2022, 20, 495. [Google Scholar] [CrossRef]
- Giorgi, C.; Marchi, S.; Simoes, I.C.M.; Ren, Z.; Morciano, G.; Perrone, M.; Patalas-Krawczyk, P.; Borchard, S.; Jędrak, P.; Pierzynowska, K.; et al. Mitochondria and Reactive Oxygen Species in Aging and Age-Related Diseases. Int. Rev. Cell Mol. Biol. 2018, 340, 209–344. [Google Scholar] [CrossRef] [PubMed]
- Wells, P.G.; Bhatia, S.; Drake, D.M.; Miller-Pinsler, L. Fetal oxidative stress mechanisms of neurodevelopmental deficits and exacerbation by ethanol and methamphetamine. Birth Defects Res. Part. C—Embryo Today Rev. 2016, 108, 108–130. [Google Scholar] [CrossRef] [PubMed]
- Wallace, D.C. A mitochondrial paradigm of metabolic and degenerative diseases, aging, and cancer: A dawn for evolutionary medicine. Annu. Rev. Genet. 2005, 39, 359–407. [Google Scholar] [CrossRef] [PubMed]
- Weinberg, F.; Ramnath, N.; Nagrath, D. Reactive oxygen species in the tumor microenvironment: An overview. Cancers 2019, 11, 1191. [Google Scholar] [CrossRef] [PubMed]
- Seitz, H.K.; Mueller, S. Alcohol and cancer: An overview with special emphasis on the role of acetaldehyde and cytochrome P450 2E1. Adv. Exp. Med. Biol. 2015, 815, 59–70. [Google Scholar] [CrossRef] [PubMed]
- Matilainen, O.; Quirós, P.M.; Auwerx, J. Mitochondria and Epigenetics—Crosstalk in Homeostasis and Stress. Trends Cell Biol. 2017, 27, 453–463. [Google Scholar] [CrossRef] [PubMed]
- Bhatia, S.; Drake, D.M.; Miller, L.; Wells, P.G. Oxidative stress and DNA damage in the mechanism of fetal alcohol spectrum disorders. Birth Defects Res. 2019, 111, 714–748. [Google Scholar] [CrossRef] [PubMed]
- Starkman, B.G.; Sakharkar, A.J.; Pandey, S.C. Epigenetics-beyond the genome in alcoholism. Alcohol. Res. 2012, 34, 293–305. [Google Scholar]
- Sarkar, D.K.; Gangisetty, O.; Wozniak, J.R.; Eckerle, J.K.; Georgieff, M.K.; Foroud, T.M.; Wetherill, L.; Wertelecki, W.; Chambers, C.D.; Riley, E.; et al. Persistent Changes in Stress-Regulatory Genes in Pregnant Women or Children Exposed Prenatally to Alcohol. Alcohol. Clin. Exp. Res. 2019, 43, 1887–1897. [Google Scholar] [CrossRef]
- Romero-Márquez, J.M.; Navarro-Hortal, M.D.; Jiménez-Trigo, V.; Muñoz-Ollero, P.; Forbes-Hernández, T.Y.; Esteban-Muñoz, A.; Giampieri, F.; Noya, I.D.; Bullón, P.; Vera-Ramírez, L.; et al. An Olive-Derived Extract 20% Rich in Hydroxytyrosol Prevents β-Amyloid Aggregation and Oxidative Stress, Two Features of Alzheimer Disease, via SKN-1/NRF2 and HSP-16.2 in Caenorhabditis elegans. Antioxidants 2022, 11, 629. [Google Scholar] [CrossRef]
- Chu, J.; Tong, M.; Monte, S.M. Chronic ethanol exposure causes mitochondrial dysfunction and oxidative stress in immature central nervous system neurons. Acta Neuropathol. 2007, 113, 659–673. [Google Scholar] [CrossRef] [PubMed]
- Higuchi, S.; Matsushita, S.; Murayama, M.; Takagi, S.; Hayashida, M. Alcohol and aldehyde dehydrogenase polymorphisms and the risk for alcoholism. Am. J. Psychiatry 1995, 152, 1219–1221. [Google Scholar] [CrossRef] [PubMed]
- Darbinian, N.; Darbinyan, A.; Merabova, N.; Kassem, M.; Tatevosian, G.; Amini, S.; Goetzl, L.; Selzer, M.E. In utero ethanol exposure induces mitochondrial DNA damage and inhibits mtDNA repair in developing brain. Front. Neurosci. 2023, 17, 958. [Google Scholar] [CrossRef] [PubMed]
- Grzeszczak, K.; Łanocha-Arendarczyk, N.; Malinowski, W.; Ziętek, P.; Kosik-Bogacka, D. Oxidative Stress in Pregnancy. Biomolecules 2023, 13, 1768. [Google Scholar] [CrossRef] [PubMed]
- Singh, S.; Verma, S.K.; Kumar, S.; Ahmad, M.K.; Nischal, A.; Singh, S.K.; Dixit, R.K. Evaluation of Oxidative Stress and Antioxidant Status in Chronic Obstructive Pulmonary Disease. Scand. J. Immunol. 2017, 85, 130–137. [Google Scholar] [CrossRef] [PubMed]
- Gutala, R.; Wang, J.; Kadapakkam, S.; Hwang, Y.; Ticku, M.; Li, M.D. Microarray analysis of ethanol-treated cortical neurons reveals disruption of genes related to the ubiquitin-proteasome pathway and protein synthesis. Alcohol. Clin. Exp. Res. 2004, 28, 1779–1788. [Google Scholar] [CrossRef] [PubMed]
- Carabulea, A.L.; Janeski, J.D.; Naik, V.D.; Chen, K.; Mor, G.; Ramadoss, J. A multi-organ analysis of the role of mTOR in fetal alcohol spectrum disorders. FASEB J. 2023, 37, e22897. [Google Scholar] [CrossRef] [PubMed]
- Sokoloff, L. Energetics of functional activation in neural tissues. Neurochem. Res. 1999, 24, 321–329. [Google Scholar] [CrossRef]
- Gerlach, M.; Ben-Shachar, D.; Riederer, P.; Youdim, M.B.H. Altered brain metabolism of iron as a cause of neurodegenerative diseases? J. Neurochem. 1994, 63, 793–807. [Google Scholar] [CrossRef]
- Floyd, R.A.; Carney, J.M. Free radical damage to protein and DNA: Mechanisms involved and relevant observations on brain undergoing oxidative stress. Ann. Neurol. 1992, 32, S22–S27. [Google Scholar] [CrossRef]
- Bergamini, C.; Gambetti, S.; Dondi, A.; Cervellati, C. Oxygen, Reactive Oxygen Species and Tissue Damage. Curr. Pharm. Des. 2005, 10, 1611–1626. [Google Scholar] [CrossRef] [PubMed]
- Bakoyiannis, I.; Gkioka, E.; Pergialiotis, V.; Mastroleon, I.; Prodromidou, A.; Vlachos, G.D.; Perrea, D. Fetal alcohol spectrum disorders and cognitive functions of young children. Rev. Neurosci. 2014, 25, 631–639. [Google Scholar] [CrossRef] [PubMed]
- Rompala, G.R.; Finegersh, A.; Homanics, G.E. Paternal preconception ethanol exposure blunts hypothalamic-pituitary-adrenal axis responsivity and stress-induced excessive fluid intake in male mice. Alcohol 2016, 53, 19–25. [Google Scholar] [CrossRef] [PubMed]
- Pearson, J.; Tarabulsy, G.M.; Bussières, E.-L. Foetal programming and cortisol secretion in early childhood: A meta-analysis of different programming variables. Infant. Behav. Dev. 2015, 40, 204–215. [Google Scholar] [CrossRef] [PubMed]
- Racine, E.; Bell, E.; Di Pietro, N.C.; Wade, L.; Illes, J. Evidence-Based Neuroethics for Neurodevelopmental Disorders. Semin. Pediatr. Neurol. 2011, 18, 21–25. [Google Scholar] [CrossRef] [PubMed]
- Olson, H.C.; Oti, R.; Gelo, J.; Beck, S. “Family matters:” Fetal alcohol spectrum disorders and the family. Dev. Disabil. Res. Rev. 2009, 15, 235–249. [Google Scholar] [CrossRef] [PubMed]
- Roozen, S.; Black, D.; Peters, G.J.Y.; Kok, G.; Townend, D.; Nijhuis, J.G.; Koek, G.H.; Curfs, L.M.G. Fetal Alcohol Spectrum Disorders (FASD): An Approach to Effective Prevention. Curr. Dev. Disord. Rep. 2016, 3, 229–234. [Google Scholar] [CrossRef] [PubMed]
- Pei, J.; Flannigan, K. Interventions for Fetal Alcohol Spectrum Disorder: Meeting Needs Across the Lifespan. Int. J. Neurorehabilitation 2016, 3, 1–9. [Google Scholar] [CrossRef]
- Ordenewitz, L.K.; Weinmann, T.; Schlüter, J.A.; Moder, J.E.; Jung, J.; Kerber, K.; Greif-Kohistani, N.; Heinen, F.; Landgraf, M.N. Evidence-based interventions for children and adolescents with fetal alcohol spectrum disorders—A systematic review. Eur. J. Paediatr. Neurol. 2021, 33, 50–60. [Google Scholar] [CrossRef]
- Christine Christensen Løhaugen, G.; Cecilie Tveiten, A.; Skranes, J. Interventions for children and adolescents with Fetal Alcohol Spectrum Disorders (FASD). In Handbook of Substance Misuse and Addictions; Patel, V.B., Preedy, V.R., Eds.; Springer International Publishing: Cham, Switzerland, 2022; pp. 1–28. ISBN 978-3-030-67928-6. [Google Scholar]
- Nazarova, V.A.; Sokolov, A.V.; Chubarev, V.N.; Tarasov, V.V.; Schiöth, H.B. Treatment of ADHD: Drugs, psychological therapies, devices, complementary and alternative methods as well as the trends in clinical trials. Front. Pharmacol. 2022, 13, 1066988. [Google Scholar] [CrossRef]
- Ritfeld, G.J.; Kable, J.A.; Holton, J.E.; Coles, C.D. Psychopharmacological Treatments in Children with Fetal Alcohol Spectrum Disorders: A Review. Child. Psychiatry Hum. Dev. 2022, 53, 268–277. [Google Scholar] [CrossRef] [PubMed]
- Mela, M.; Okpalauwaekwe, U.; Anderson, T.; Eng, J.; Nomani, S.; Ahmed, A.; Barr, A.M. The utility of psychotropic drugs on patients with Fetal Alcohol Spectrum Disorder (FASD): A systematic review. Psychiatry Clin. Psychopharmacol. 2018, 28, 436–445. [Google Scholar] [CrossRef]
- Paley, B.; O’Connor, M.J. Behavioral interventions for children and adolescents with fetal alcohol spectrum disorders. Alcohol. Res. Health 2011, 34, 64–75. [Google Scholar]
- Riley, E.P.; McGee, C.L. Fetal alcohol spectrum disorders: An overview with emphasis on changes in brain and behavior. Exp. Biol. Med. 2005, 230, 357–365. [Google Scholar] [CrossRef] [PubMed]
- Kerns, K.A.; MacSween, J.; Vander Wekken, S.; Gruppuso, V. Investigating the efficacy of an attention training programme in children with foetal alcohol spectrum disorder. Dev. Neurorehabil. 2010, 13, 413–422. [Google Scholar] [CrossRef] [PubMed]
- Kable, J.A.; Taddeo, E.; Strickland, D.; Coles, C.D. Community translation of the Math Interactive Learning Experience Program for children with FASD. Res. Dev. Disabil. 2015, 39, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Adnams, C.M.; Sorour, P.; Kalberg, W.O.; Kodituwakku, P.; Perold, M.D.; Kotze, A.; September, S.; Castle, B.; Gossage, J.; May, P.A. Language and literacy outcomes from a pilot intervention study for children with fetal alcohol spectrum disorders in South Africa. Alcohol 2007, 41, 403–414. [Google Scholar] [CrossRef]
- Frankel, F.; Myatt, R. Children’s Friendship Training; Taylor & Francis Ltd.: London, UK, 2013; pp. 1–198. [Google Scholar] [CrossRef]
- Laugeson, E.A.; Paley, B.; Schonfeld, A.M.; Carpenter, E.M.; Frankel, F.; O’Connor, M.J. Adaptation of the children’s friendship training program for children with Fetal Alcohol Spectrum Disorders. Child. Fam. Behav. Ther. 2007, 29, 57–69. [Google Scholar] [CrossRef]
- Leenaars, L.S.; Denys, K.; Henneveld, D.; Rasmussen, C. The impact of fetal alcohol spectrum disorders on families: Evaluation of a family intervention program. Community Ment. Health J. 2012, 48, 431–435. [Google Scholar] [CrossRef]
- Kable, J.A.; Coles, C.D.; Taddeo, E. Socio-cognitive habilitation using the math interactive learning experience program for alcohol-affected children. Alcohol. Clin. Exp. Res. 2007, 31, 1425–1434. [Google Scholar] [CrossRef]
- Sambo, D.; Goldman, D. Genetic influences on Fetal Alcohol Spectrum Disorder. Genes 2023, 14, 195. [Google Scholar] [CrossRef] [PubMed]
- Freeman, J.; Condon, C.; Hamilton, S.; Mutch, R.C.; Bower, C.; Watkins, R.E. Challenges in Accurately Assessing Prenatal Alcohol Exposure in a Study of Fetal Alcohol Spectrum Disorder in a Youth Detention Center. Alcohol. Clin. Exp. Res. 2019, 43, 309–316. [Google Scholar] [CrossRef] [PubMed]
- Cohen-Kerem, R.; Koren, G. Antioxidants and fetal protection against ethanol teratogenicity. I. Review of the experimental data and implications to humans. Neurotoxicol. Teratol. 2003, 25, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Micangeli, G.; Menghi, M.; Profeta, G.; Tarani, F.; Mariani, A.; Petrella, C.; Barbato, C.; Ferraguti, G.; Ceccanti, M.; Tarani, L.; et al. The Impact of Oxidative Stress on Pediatrics Syndromes. Antioxidants 2022, 11, 1983. [Google Scholar] [CrossRef]
- Petrella, C.; Carito, V.; Carere, C.; Ferraguti, G.; Ciafrè, S.; Natella, F.; Bello, C.; Greco, A.; Ralli, M.; Mancinelli, R.; et al. Oxidative stress inhibition by resveratrol in alcohol-dependent mice. Nutrition 2020, 79–80, 110783. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Wang, H.; Li, Y.; Peng, Y. A review of interventions against fetal alcohol spectrum disorder targeting oxidative stress. Int. J. Dev. Neurosci. 2018, 71, 140–145. [Google Scholar] [CrossRef] [PubMed]
- Zheng, D.; Li, Y.; He, L.; Tang, Y.; Li, X.; Shen, Q.; Yin, D.; Peng, Y. The protective effect of astaxanthin on fetal alcohol spectrum disorder in mice. Neuropharmacology 2014, 84, 13–18. [Google Scholar] [CrossRef] [PubMed]
- Davis, K.; Desrocher, M.; Moore, T. Fetal Alcohol Spectrum Disorder: A Review of Neurodevelopmental Findings and Interventions. J. Dev. Phys. Disabil. 2011, 23, 143–167. [Google Scholar] [CrossRef]
- Carito, V.; Ceccanti, M.; Cestari, V.; Natella, F.; Bello, C.; Coccurello, R.; Mancinelli, R.; Fiore, M. Olive polyphenol effects in a mouse model of chronic ethanol addiction. Nutrition 2017, 33, 65–69. [Google Scholar] [CrossRef]
- Ferraguti, G.; Terracina, S.; Micangeli, G.; Lucarelli, M.; Tarani, L.; Ceccanti, M.; Spaziani, M.; D’Orazi, V.; Petrella, C.; Fiore, M. NGF and BDNF in pediatrics syndromes. Neurosci. Biobehav. Rev. 2023, 145, 105015. [Google Scholar] [CrossRef]
- Jawaid, S.; Strainic, J.P.; Kim, J.; Ford, M.R.; Thrane, L.; Karunamuni, G.H.; Sheehan, M.M.; Chowdhury, A.; Gillespie, C.A.; Rollins, A.M.; et al. Glutathione Protects the Developing Heart from Defects and Global DNA Hypomethylation Induced by Prenatal Alcohol Exposure. Alcohol. Clin. Exp. Res. 2021, 45, 69–78. [Google Scholar] [CrossRef] [PubMed]
- Ciafrè, S.; Carito, V.; Ferraguti, G.; Greco, A.; Chaldakov, G.N.; Fiore, M.; Ceccanti, M. How alcohol drinking affects our genes: An epigenetic point of view. Biochem. Cell Biol. 2019, 97, 345–356. [Google Scholar] [CrossRef] [PubMed]
- Denny, L.A.; Coles, S.; Blitz, R. Fetal Alcohol Syndrome and Fetal Alcohol Spectrum Disorders. Am. Fam. Physician 2017, 96, 515–522. [Google Scholar] [CrossRef] [PubMed]
- Ojeda, L.; Nogales, F.; Murillo, L.; Carreras, O. The role of folic acid and selenium against oxidative damage from ethanol in early life programming: A review. Biochem. Cell Biol. 2018, 96, 178–188. [Google Scholar] [CrossRef] [PubMed]
- Karunamuni, G.; Sheehan, M.M.; Doughman, Y.Q.; Gu, S.; Sun, J.; Li, Y.; Strainic, J.P.; Rollins, A.M.; Jenkins, M.W.; Watanabe, M. Supplementation with the Methyl Donor Betaine Prevents Congenital Defects Induced by Prenatal Alcohol Exposure. Alcohol. Clin. Exp. Res. 2017, 41, 1917–1927. [Google Scholar] [CrossRef] [PubMed]
- Hoyme, H.E.; Kalberg, W.O.; Elliott, A.J.; Blankenship, J.; Buckley, D.; Marais, A.S.; Manning, M.A.; Robinson, L.K.; Adam, M.P.; Abdul-Rahman, O.; et al. Updated clinical guidelines for diagnosing fetal alcohol spectrum disorders. Pediatrics 2016, 138, e20154256. [Google Scholar] [CrossRef]
- Fainsod, A.; Bendelac-Kapon, L.; Shabtai, Y. Fetal Alcohol Spectrum Disorder: Embryogenesis Under Reduced Retinoic Acid Signaling Conditions. Subcell. Biochem. 2020, 95, 197–225. [Google Scholar] [CrossRef] [PubMed]
- Ceci, F.M.; Ferraguti, G.; Petrella, C.; Greco, A.; Ralli, M.; Iannitelli, A.; Carito, V.; Tirassa, P.; Chaldakov, G.N.; Messina, M.P.; et al. Nerve Growth Factor in Alcohol Use Disorders. Curr. Neuropharmacol. 2020, 19, 45–60. [Google Scholar] [CrossRef] [PubMed]
- Thanh, N.X.; Jonsson, E. Life expectancy of people with fetal alcohol syndrome. J. Popul. Ther. Clin. Pharmacol. 2016, 23, e53–e59. [Google Scholar]
- May, P.A.; Gossage, J.P. Maternal risk factors for fetal alcohol spectrum disorders: Not as simple as it might seem. Alcohol. Res. Health 2011, 34, 15–26. [Google Scholar]
- Thanh, N.X.; Jonsson, E.; Salmon, A.; Sebastianski, M. Incidence and prevalence of fetal alcohol spectrum disorder by sex and age group in Alberta, Canada. J. Popul. Ther. Clin. Pharmacol. 2014, 21, e395–e404. [Google Scholar] [PubMed]
- Popova, S.; Lange, S.; Probst, C.; Gmel, G.; Rehm, J. Estimation of national, regional, and global prevalence of alcohol use during pregnancy and fetal alcohol syndrome: A systematic review and meta-analysis. Lancet Glob. Health 2017, 5, e290–e299. [Google Scholar] [CrossRef] [PubMed]
- Ferraguti, G.; Ciolli, P.; Carito, V.; Battagliese, G.; Mancinelli, R.; Ciafrè, S.; Tirassa, P.; Ciccarelli, R.; Cipriani, A.; Messina, M.P.; et al. Ethylglucuronide in the urine as a marker of alcohol consumption during pregnancy: Comparison with four alcohol screening questionnaires. Toxicol. Lett. 2017, 275, 49–56. [Google Scholar] [CrossRef] [PubMed]
- Steane, S.E.; Cuffe, J.S.M.; Moritz, K.M. The role of maternal choline, folate and one-carbon metabolism in mediating the impact of prenatal alcohol exposure on placental and fetal development. J. Physiol. 2023, 601, 1061–1075. [Google Scholar] [CrossRef] [PubMed]
- May, P.A.; Marais, A.-S.; Kalberg, W.O.; de Vries, M.M.; Buckley, D.; Hasken, J.M.; Snell, C.L.; Barnard Röhrs, R.; Hedrick, D.M.; Bezuidenhout, H.; et al. Multifaceted case management during pregnancy is associated with better child outcomes and less fetal alcohol syndrome. Ann. Med. 2023, 55, 926–945. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Population | Intervention | Outcome | Reference |
|---|---|---|---|
| FAS patients and animal models | Prenatal antioxidant administration food supplements, folic acid, choline, neuroactive peptides, neurotrophic growth factors, and lifestyle interventions. | The treatment options for FAS have recently started to be explored, although none are currently approved clinically. Furthermore, avoiding comorbidities and addressing the family system can significantly improve the quality of life. | [15] |
| Animal models | Responsiveness to various stimuli including perinatal care, diet, and physical activity. | Environments richer in prenatal care, or richer in stimuli, or giving the possibility of practice a specific skill (e.g., motor abilities), or providing a diet richer in antioxidants, tend to minimize the noxious effects of alcohol exposure, suggesting the plasticity of the central nervous system when there are favorable contextual factors and timely therapeutic interventions. | [72] |
| Cell cultures, animal models | Vitamin E, β-carotene, flavonoids, and folic acid | Antioxidants have neuroprotective effects and prevent ethanol teratogenicity. | [106] |
| Animal models and FASD patients | Polyphenols, carotenoids, thioredoxins, vitamin E | Although further studies are needed to better understand the relationship between oxidative stress and pediatric diseases, evidence encourages future therapeutic strategies. | [108] |
| Animal models | Resveratrol | Potential use as a dietary supplement to prevent damage due to oxidative stress associated with chronic alcohol abuse. | [109] |
| Cell cultures, animal models | Astaxanthin, ascorbic acid (Vitamin C), Vitamin E, β-carotene, (-)-epigallocatechin-3-gallate (EGCG), omega-3 fatty acids, folic acid, neurotrophic factor-9 | We have many interventions effective against oxidative stress associated with FASD, but most evidence comes from animal models; more clinical trials are needed to show whether or not antioxidants may act against FASD damage. | [110] |
| Animal models | Astaxanthin | Protective effect on FASD, acting against oxidative stress- and TLR4 signaling-associated inflammatory reaction. | [111] |
| Animal models | Polyphenols | Polyphenol supplementation partially counteracts the pro-oxidant effects of alcohol. | [113] |
| Animal embryo models | Glutathione | Glutathione supplementation protects from heart defects and global DNA hypomethylation induced by prenatal alcohol exposure. | [115] |
| Cell cultures, animal models | NA | Targeting the alcohol-mediated epigenetics effects may offer new avenues for therapeutic interventions in FASD. | [116] |
| Animal models | Folic acid, selenium | These two antioxidants may play a major role in FASD management by acting on the methionine metabolic cycle. | [118] |
| Animal embryo models | Methyl donor betaine | Supplementation with the methyl donor betaine prevents congenital defects induced by prenatal alcohol exposure. | [119] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Terracina, S.; Tarani, L.; Ceccanti, M.; Vitali, M.; Francati, S.; Lucarelli, M.; Venditti, S.; Verdone, L.; Ferraguti, G.; Fiore, M. The Impact of Oxidative Stress on the Epigenetics of Fetal Alcohol Spectrum Disorders. Antioxidants 2024, 13, 410. https://doi.org/10.3390/antiox13040410
Terracina S, Tarani L, Ceccanti M, Vitali M, Francati S, Lucarelli M, Venditti S, Verdone L, Ferraguti G, Fiore M. The Impact of Oxidative Stress on the Epigenetics of Fetal Alcohol Spectrum Disorders. Antioxidants. 2024; 13(4):410. https://doi.org/10.3390/antiox13040410
Chicago/Turabian StyleTerracina, Sergio, Luigi Tarani, Mauro Ceccanti, Mario Vitali, Silvia Francati, Marco Lucarelli, Sabrina Venditti, Loredana Verdone, Giampiero Ferraguti, and Marco Fiore. 2024. "The Impact of Oxidative Stress on the Epigenetics of Fetal Alcohol Spectrum Disorders" Antioxidants 13, no. 4: 410. https://doi.org/10.3390/antiox13040410
APA StyleTerracina, S., Tarani, L., Ceccanti, M., Vitali, M., Francati, S., Lucarelli, M., Venditti, S., Verdone, L., Ferraguti, G., & Fiore, M. (2024). The Impact of Oxidative Stress on the Epigenetics of Fetal Alcohol Spectrum Disorders. Antioxidants, 13(4), 410. https://doi.org/10.3390/antiox13040410