
Health Benefits of Electrolyzed Hydrogen Water: Antioxidant and Anti-Inflammatory Effects in Living Organisms



Abstract
1. Introduction
2. Properties of Hydrogen
3. Antioxidant Effects of Hydrogen
4. Anti-Inflammatory Effects of EHW
5. Analgesic Effects of EHW
6. Anti-Stress Effects of EHW
7. Other Biological Effects of EHW
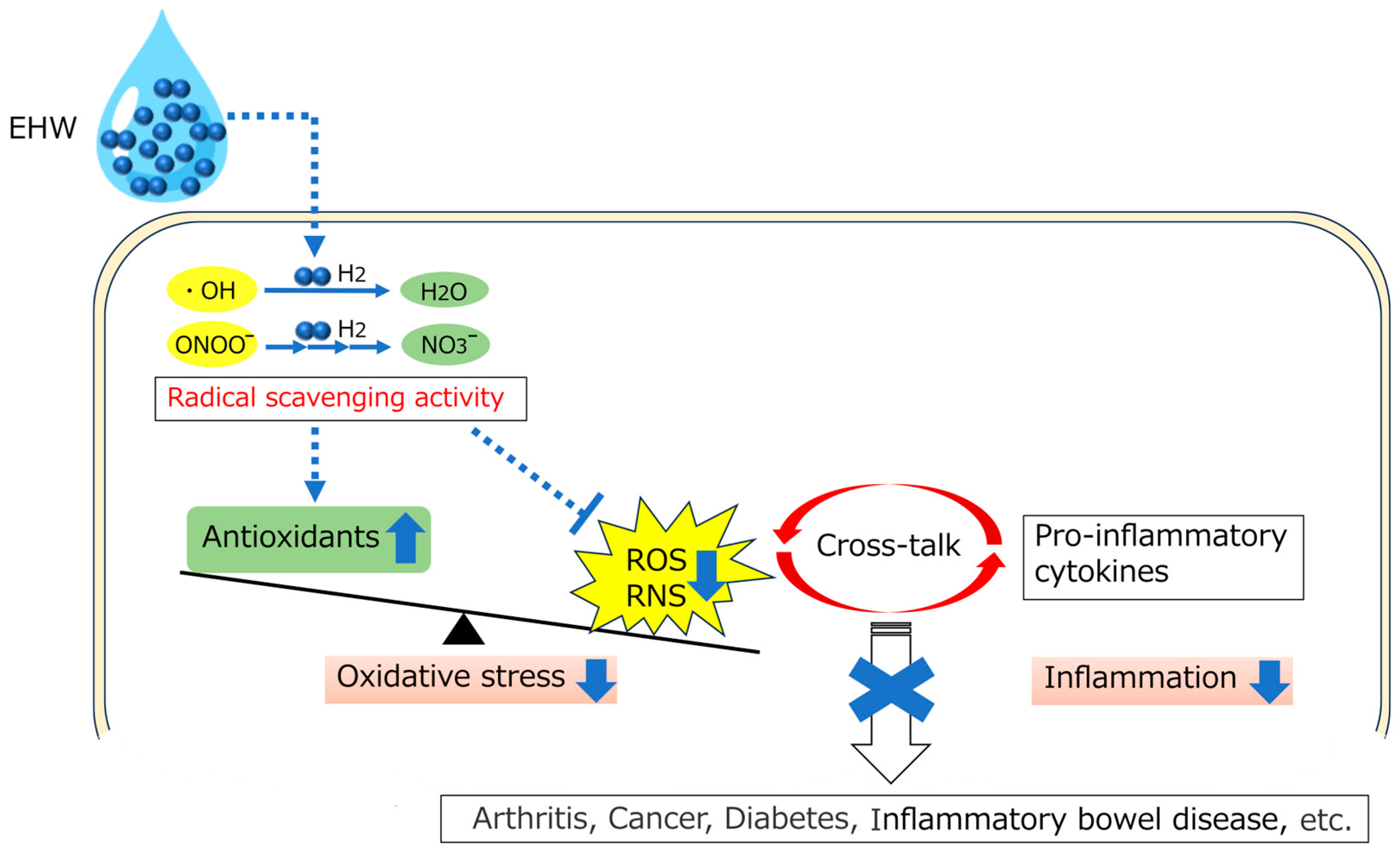
8. Conclusions and Perspectives
Author Contributions
Funding
Conflicts of Interest
References
- Weller, M.; Overton, T.; Rourke, J.; Armstrong, F. Inorganic Chemistry, 6th ed.; Oxford University Press: Oxford, UK, 2016. [Google Scholar]
- Lee, J.D. Concise Inorganic Chemistry for JEE (Main & Advanced), 5ed ed.; Wiley India Pvt Ltd.: New Delhi, India, 2023. [Google Scholar]
- Kayar, S.R.; Axley, M.J.; Homer, L.D.; Harabin, A.L. Hydrogen gas is not oxidized by mammalian tissues under hyperbaric conditions. Undersea Hyperb. Med. 1994, 21, 265–275. [Google Scholar] [PubMed]
- Ohsawa, I.; Ishikawa, M.; Takahashi, K.; Watanabe, M.; Nishimaki, K.; Yamagata, K.; Katsura, K.; Katayama, Y.; Asoh, S.; Ohta, S. Hydrogen acts as a therapeutic antioxidant by selectively reducing cytotoxic oxygen radicals. Nat. Med. 2007, 13, 688–694. [Google Scholar] [CrossRef] [PubMed]
- Coral-Pérez, S.; Martínez-Martel, I.; Martínez-Serrat, M.; Batallé, G.; Bai, X.; Leite-Panissi, C.; Pol, O. Treatment with Hydrogen-Rich Water Improves the Nociceptive and Anxio-Depressive-like Behaviors Associated with Chronic Inflammatory Pain in Mice. Antioxidants 2022, 11, 2153. [Google Scholar] [CrossRef] [PubMed]
- Hu, D.; Huang, T.L.; Shigeta, M.; Ochi, Y.; Kabayama, S.; Watanabe, Y.; Cui, Y.L. Electrolyzed Hydrogen Water Alleviates Abdominal Pain through Suppression of Colonic Tissue Inflammation in a Rat Model of Inflammatory Bowel Disease. Nutrients 2022, 14, 4451. [Google Scholar] [CrossRef] [PubMed]
- Naito, Y.; Takagi, T.; Uchiyama, K.; Tomatsuri, N.; Matsuyama, K.; Fujii, T.; Yagi, N.; Yoshida, N.; Yoshikawa, T. Chronic administration with electrolyzed alkaline water inhibits aspirin-induced gastric mucosal injury in rats through the inhibition of tumor necrosis factor-alpha expression. J. Clin. Biochem. Nutr. 2002, 32, 69–81. [Google Scholar] [CrossRef]
- Martínez-Serrat, M.; Martínez-Martel, I.; Coral-Pérez, S.; Bai, X.; Batallé, G.; Pol, O. Hydrogen-Rich Water as a Novel Therapeutic Strategy for the Affective Disorders Linked with Chronic Neuropathic Pain in Mice. Antioxidants 2022, 11, 1826. [Google Scholar] [CrossRef] [PubMed]
- Itoh, T.; Fujita, Y.; Ito, M.; Masuda, A.; Ohno, K.; Ichihara, M.; Kojima, T.; Nozawa, Y.; Ito, M. Molecular hydrogen suppresses FcepsilonRI-mediated signal transduction and prevents degranulation of mast cells. Biochem. Biophys. Res. Commun. 2009, 389, 651–656. [Google Scholar] [CrossRef]
- Ishibashi, T.; Sato, B.; Rikitake, M.; Seo, T.; Kurokawa, R.; Hara, Y.; Naritomi, Y.; Hara, H.; Nagao, T. Consumption of water containing a high concentration of molecular hydrogen reduces oxidative stress and disease activity in patients with rheumatoid arthritis: An open-label pilot study. Med. Gas. Res. 2012, 2, 27. [Google Scholar] [CrossRef]
- Ogawa, S.; Ohsaki, Y.; Shimizu, M.; Nako, K.; Okamura, M.; Kabayama, S.; Tabata, K.; Tanaka, Y.; Ito, S. Electrolyzed hydrogen-rich water for oxidative stress suppression and improvement of insulin resistance: A multicenter prospective double-blind randomized control trial. Diabetol. Int. 2022, 13, 209–219. [Google Scholar] [CrossRef]
- Mizuno, K.; Sasaki, A.; Ebisu, K.; Tajima, K.; Kajimoto, O.; Nojima, J.; Kuratsune, H.; Hori, H.; Watanabe, Y. Hydrogen-rich water for improvements of mood, anxiety, and autonomic nerve function in daily life. Med. Gas. Res. 2017, 7, 247. [Google Scholar] [CrossRef]
- Nakayama, M.; Itami, N.; Suzuki, H.; Hamada, H.; Yamamoto, R.; Tsunoda, K.; Osaka, N.; Nakano, H.; Maruyama, Y.; Kabayama, S.; et al. Novel haemodialysis (HD) treatment employing molecular hydrogen (H2)-enriched dialysis solution improves prognosis of chronic dialysis patients: A prospective observational study. Sci. Rep. 2018, 8, 254. [Google Scholar] [CrossRef]
- Song, L.; Zhang, Y.; Zhu, C.; Ding, X.; Yang, L.; Yan, H. Hydrogen-rich water partially alleviate inflammation, oxidative stress and intestinal flora dysbiosis in DSS-induced chronic ulcerative colitis mice. Adv. Med. Sci. 2022, 67, 29–38. [Google Scholar] [CrossRef]
- Kajisa, T.; Yamaguchi, T.; Hu, A.; Suetake, N.; Kobayashi, H. Hydrogen water ameliorates the severity of atopic dermatitis-like lesions and decreases interleukin-1β, interleukin-33, and mast cell infiltration in NC/Nga mice. Saudi Med. J. 2017, 38, 928–933. [Google Scholar] [CrossRef] [PubMed]
- Peng, J.; He, Q.; Li, S.; Liu, T.; Zhang, J. Hydrogen-Rich Water Mitigates LPS-Induced Chronic Intestinal Inflammatory Response in Rats via Nrf-2 and NF-κB Signaling Pathways. Vet. Sci. 2022, 9, 621. [Google Scholar] [CrossRef]
- Hu, D.; Li, D.; Shigeta, M.; Ochi, Y.; Okauchi, T.; Neyama, H.; Kabayama, S.; Watanabe, Y.; Cui, Y. Alleviation of the chronic stress response attributed to the antioxidant and anti-inflammatory effects of electrolyzed hydrogen water. Biochem. Biophys. Res. Commun. 2021, 535, 1–5. [Google Scholar] [CrossRef] [PubMed]
- Gu, Y.; Huang, C.S.; Inoue, T.; Yamashita, T.; Ishida, T.; Kang, K.M.; Nakao, A. Drinking Hydrogen Water Ameliorated Cognitive Impairment in Senescence-Accelerated Mice. J. Clin. Biochem. Nutr. 2010, 46, 269–276. [Google Scholar] [CrossRef] [PubMed]
- Hou, Z.; Luo, W.; Sun, X.; Hao, S.; Zhang, Y.; Xu, F.; Wang, Z.; Liu, B. Hydrogen-rich saline protects against oxidative damage and cognitive deficits after mild traumatic brain injury. Brain Res. Bull. 2012, 88, 560–565. [Google Scholar] [CrossRef] [PubMed]
- Smith, G.N.; Emerson, R.J.; Temple, L.A.; Galbraith, T.W. The oxidation of molecular tritium in mammals. Arch. Biochem. Biophys. 1953, 46, 22–30. [Google Scholar] [CrossRef] [PubMed]
- Ostojic, S.M. Targeting molecular hydrogen to mitochondria: Barriers and gateways. Pharmacol. Res. 2015, 94, 51–53. [Google Scholar] [CrossRef] [PubMed]
- Ohta, S. Molecular hydrogen as a novel antioxidant: Overview of the advantages of hydrogen for medical applications. Methods Enzymol. 2015, 555, 289–317. [Google Scholar] [CrossRef]
- Nagata, K.; Nakashima-Kamimura, N.; Mikami, T.; Ohsawa, I.; Ohta, S. Consumption of Molecular Hydrogen Prevents the Stress-Induced Impairments in Hippocampus-Dependent Learning Tasks during Chronic Physical Restraint in Mice. Neuropsychopharmacology 2009, 34, 501–508. [Google Scholar] [CrossRef] [PubMed]
- Grinter, R.; Kropp, A.; Venugopal, H.; Senger, M.; Badley, J.; Cabotaje, P.R.; Jia, R.; Duan, Z.; Huang, P.; Stripp, S.T.; et al. Structural basis for bacterial energy extraction from atmospheric hydrogen. Nature 2023, 615, 541–547. [Google Scholar] [CrossRef] [PubMed]
- Dole, M.; Wilson, F.R.; Fife, W.P. Hyperbaric hydrogen therapy: A possible treatment for cancer. Science 1975, 190, 152–154. [Google Scholar] [CrossRef] [PubMed]
- Gharib, B.; Hanna, S.; Abdallahi, O.M.; Lepidi, H.; Gardette, B.; Reggi, M.D. Anti-inflammatory properties of molecular hydrogen: Investigation on parasite-induced liver inflammation. C R. Acad. Sci. III 2001, 324, 719–724. [Google Scholar] [CrossRef]
- Ichihara, M.; Sobue, S.; Ito, M.; Ito, M.; Hirayama, M.; Ohno, K. Beneficial biological effects and the underlying mechanisms of molecular hydrogen—Comprehensive review of 321 original articles. Med. Gas. Res. 2015, 5, 12. [Google Scholar] [CrossRef] [PubMed]
- Itoh, T.; Hamada, N.; Terazawa, R.; Ito, M.; Ohno, K.; Ichihara, M.; Tanaka, Y.; Ito, M. Molecular hydrogen inhibits lipopolysaccharide/interferon gamma-induced nitric oxide production through modulation of signal transduction in macrophages. Biochem. Biophys. Res. Commun. 2011, 411, 143–149. [Google Scholar] [CrossRef]
- Terasaki, Y.; Ohsawa, I.; Terasaki, M.; Takahashi, M.; Kunugi, S.; Dedong, K.; Urushiyama, H.; Amenomori, S.; Kaneko-Togashi, M.; Kuwahara, N.; et al. Hydrogen therapy attenuates irradiation-induced lung damage by reducing oxidative stress. Am. J. Physiol. Lung Cell Mol. Physiol. 2011, 301, L415–L426. [Google Scholar] [CrossRef]
- Kamimura, N.; Nishimaki, K.; Ohsawa, I.; Ohta, S. Molecular Hydrogen Improves Obesity and Diabetes by Inducing Hepatic FGF21 and Stimulating Energy Metabolism in db/db Mice. Obesity 2011, 19, 1396–1403. [Google Scholar] [CrossRef]
- Katakura, M.; Hashimoto, M.; Tanabe, Y.; Shido, O. Hydrogen-rich water inhibits glucose and α,β -dicarbonyl compound-induced reactive oxygen species production in the SHR.Cg-Leprcp/NDmcr rat kidney. Med. Gas. Res. 2012, 2, 18. [Google Scholar] [CrossRef] [PubMed]
- Hamauzu, Y.; Ishikawa, K.; Morisawa, S. Effects of Deoxidized Nutrient Solution on Growth of Komatsuna (Brassica rapa var. perviridis) Plants. Environ. Control. Biol. 2014, 52, 107–111. [Google Scholar] [CrossRef]
- Itamura, H.; Nomura, K.; Kabayama, S.; Amano, M.; Hamauzu, Y.; Nakai, K.; Matsushita, I.; Sawada, K.; Esumi, T.; Nakatsuka, A. Effect of cultivation conditions and electrolyzed hydrogen water treatment on black staining in ‘Taishu’ persimmon fruit. Acta Hortic. 2022, 1338, 105–111. [Google Scholar] [CrossRef]
- Abraini, J.H.; Gardette-Chauffour, M.C.; Martinez, E.; Rostain, J.C.; Lemaire, C. Psychophysiological reactions in humans during an open sea dive to 500 m with a hydrogen-helium-oxygen mixture. J. Appl. Physiol. 1994, 76, 1113–1118. [Google Scholar] [CrossRef] [PubMed]
- Fontanari, P.; Badier, M.; Guillot, C.; Tomei, C.; Burnet, H.; Gardette, B.; Jammes, Y. Changes in maximal performance of inspiratory and skeletal muscles during and after the 7.1-MPa Hydra 10 record human dive. Eur. J. Appl. Physiol. 2000, 81, 325–328. [Google Scholar] [CrossRef] [PubMed]
- Shimouchi, A.; Nose, K.; Shirai, M.; Kondo, T. Estimation of molecular hydrogen consumption in the human whole body after the ingestion of hydrogen-rich water. Adv. Exp. Med. Biol. 2012, 737, 245–250. [Google Scholar] [CrossRef] [PubMed]
- Nakashima-Kamimura, N.; Mori, T.; Ohsawa, I.; Asoh, S.; Ohta, S. Molecular hydrogen alleviates nephrotoxicity induced by an anticancer drug cisplatin without compromising anti-tumor activity in mice. Cancer Chemother. Pharmacol. 2009, 64, 753–761. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; Kurokawa, R.; Fujino, M.; Hirano, S.; Sato, B.; Li, X. Corrigendum: Estimation of the hydrogen concentration in rat tissue using an airtight tube following the administration of hydrogen via various routes. Sci. Rep. 2015, 5, 9629. [Google Scholar] [CrossRef] [PubMed]
- Lin, C.P.; Chuang, W.C.; Lu, F.J.; Chen, C.Y. Anti-oxidant and anti-inflammatory effects of hydrogen-rich water alleviate ethanol-induced fatty liver in mice. World J. Gastroenterol. 2017, 23, 4920–4934. [Google Scholar] [CrossRef]
- Ara, J.; Fadriquela, A.; Ahmed, M.F.; Bajgai, J.; Sajo, M.E.J.; Lee, S.P.; Kim, T.S.; Jung, J.Y.; Kim, C.S.; Kim, S.K.; et al. Hydrogen Water Drinking Exerts Antifatigue Effects in Chronic Forced Swimming Mice via Antioxidative and Anti-Inflammatory Activities. Biomed. Res. Int. 2018, 2018, 2571269. [Google Scholar] [CrossRef]
- Dohi, K.; Kraemer, B.C.; Erickson, M.A.; McMillan, P.J.; Kovac, A.; Flachbartova, Z.; Hansen, K.M.; Shah, G.N.; Sheibani, N.; Salameh, T.; et al. Molecular Hydrogen in Drinking Water Protects against Neurodegenerative Changes Induced by Traumatic Brain Injury. PLoS ONE 2014, 9, e108034. [Google Scholar] [CrossRef]
- Shirahata, S.; Kabayama, S.; Nakano, M.; Miura, T.; Kusumoto, K.; Gotoh, M.; Hayashi, H.; Otsubo, K.; Morisawa, S.; Katakura, Y. Electrolyzed-reduced water scavenges active oxygen species and protects DNA from oxidative damage. Biochem. Biophys. Res. Commun. 1997, 234, 269–274. [Google Scholar] [CrossRef]
- Lee, M.Y.; Kim, Y.K.; Ryoo, K.K.; Lee, Y.B.; Park, E.J. Electrolyzed-reduced water protects against oxidative damage to DNA, RNA, and protein. Appl. Biochem. Biotechnol. 2006, 135, 133–144. [Google Scholar] [CrossRef]
- Zhang, J.Y.; Wu, Q.F.; Wan, Y.; Song, S.D.; Xu, J.; Xu, X.S.; Chang, H.L.; Tai, M.H.; Dong, Y.F.; Liu, C. Protective role of hydrogen-rich water on aspirin-induced gastric mucosal damage in rats. World J. Gastroenterol. 2014, 20, 1614–1622. [Google Scholar] [CrossRef]
- Jin, D.; Ryu, S.H.; Kim, H.W.; Yang, E.J.; Lim, S.J.; Ryang, Y.S.; Chung, C.H.; Park, S.K.; Lee, K.J. Anti-Diabetic Effect of Alkaline-Reduced Water on OLETF Rats. Biosci. Biotechnol. Biochem. 2006, 70, 31–37. [Google Scholar] [CrossRef]
- Saitoh, Y.; Okayasu, H.; Xiao, L.; Harata, Y.; Miwa, N. Neutral pH hydrogen-enriched electrolyzed water achieves tumor-preferential clonal growth inhibition over normal cells and tumor invasion inhibition concurrently with intracellular oxidant repression. Oncol. Res. 2008, 17, 247–255. [Google Scholar] [CrossRef]
- Nishikawa, R.; Teruya, K.; Katakura, Y.; Osada, K.; Hamasaki, T.; Kashiwagi, T.; Komatsu, T.; Li, Y.; Ye, J.; Ichikawa, A.; et al. Electrolyzed Reduced Water Supplemented with Platinum Nanoparticles Suppresses Promotion of Two-stage Cell Transformation. Cytotechnology 2005, 47, 97–105. [Google Scholar] [CrossRef]
- Hamasaki, T.; Harada, G.; Nakamichi, N.; Kabayama, S.; Teruya, K.; Fugetsu, B.; Gong, W.; Sakata, I.; Shirahata, S. Electrochemically reduced water exerts superior reactive oxygen species scavenging activity in HT1080 cells than the equivalent level of hydrogen-dissolved water. PLoS ONE 2017, 12, e0171192. [Google Scholar] [CrossRef]
- Wang, Y.; Fugetsu, B.; Sakata, I.; Fujisue, C.; Kabayama, S.; Tahara, N.; Morisawa, S. Monolayered Platinum Nanoparticles as Efficient Electrocatalysts for the Mass Production of Electrolyzed Hydrogen Water. Sci. Rep. 2020, 10, 10126. [Google Scholar] [CrossRef] [PubMed]
- Xue, J.; Shang, G.; Tanaka, Y.; Saihara, Y.; Hou, L.; Velasquez, N.; Liu, W.; Lu, Y. Dose-dependent inhibition of gastric injury by hydrogen in alkaline electrolyzed drinking water. BMC Complement. Altern. Med. 2014, 14, 81. [Google Scholar] [CrossRef] [PubMed]
- Spulber, S.; Edoff, K.; Hong, L.; Morisawa, S.; Shirahata, S.; Ceccatelli, S. Molecular Hydrogen Reduces LPS-Induced Neuroinflammation and Promotes Recovery from Sickness Behaviour in Mice. PLoS ONE 2012, 7, e42078. [Google Scholar] [CrossRef] [PubMed]
- Kashiwagi, T.; Yan, H.; Hamasaki, T.; Kinjo, T.; Nakamichi, N.; Teruya, K.; Kabayama, S.; Shirahata, S. Electrochemically Reduced Water Protects Neural Cells from Oxidative Damage. Oxid. Med. Cell Longev. 2014, 2014, 869121. [Google Scholar] [CrossRef] [PubMed]
- Yano, S.; Wang, J.; Kabayama, S.; Hara, T. Electrolyzed Hydrogen Water Protects against Ethanol-Induced Cytotoxicity by Regulating Aldehyde Metabolism-Associated Enzymes in the Hepatic Cell Line HepG2. Antioxidants 2021, 10, 801. [Google Scholar] [CrossRef]
- Huang, K.C.; Yang, C.C.; Lee, K.T.; Chien, C.T. Reduced hemodialysis-induced oxidative stress in end-stage renal disease patients by electrolyzed reduced water. Kidney Int. 2003, 64, 704–714. [Google Scholar] [CrossRef] [PubMed]
- Morgan, M.J.; Liu, Z. Crosstalk of reactive oxygen species and NF-kappaB signaling. Cell Res. 2011, 21, 103–115. [Google Scholar] [CrossRef] [PubMed]
- Bourgonje, A.R.; Feelisch, M.; Faber, K.N.; Pasch, A.; Dijkstra, G.; van Goor, H. Oxidative Stress and Redox-Modulating Therapeutics in Inflammatory Bowel Disease. Trends Mol. Med. 2020, 26, 1034–1046. [Google Scholar] [CrossRef] [PubMed]
- Lingappan, K. NF-κB in oxidative stress. Curr. Opin. Toxicol. 2018, 7, 81–86. [Google Scholar] [CrossRef] [PubMed]
- Ranneh, Y.; Ali, F.; Akim, A.M.; Hamid, H.A.; Khazaai, H.; Fadel, A. Crosstalk between reactive oxygen species and pro-inflammatory markers in developing various chronic diseases: A review. Appl. Biol. Chem. 2017, 60, 327–338. [Google Scholar] [CrossRef]
- Canton, M.; Sánchez-Rodríguez, R.; Spera, I.; Venegas, F.C.; Favia, M.; Viola, A.; Castegna, A. Reactive Oxygen Species in Macrophages: Sources and Targets. Front. Immunol. 2021, 12, 734229. [Google Scholar] [CrossRef] [PubMed]
- Simon, A.R.; Rai, U.; Fanburg, B.L.; Cochran, B.H. Activation of the JAK-STAT pathway by reactive oxygen species. Am. J. Physiol. 1998, 275, C1640–C1652. [Google Scholar] [CrossRef] [PubMed]
- Fan, G.; Li, Y.; Liu, Y.; Suo, X.; Jia, Y.; Yang, X. Gondoic acid alleviates LPS-induced Kupffer cells inflammation by inhibiting ROS production and PKCθ/ERK/STAT3 signaling pathway. Int. Immunopharmacol. 2022, 111, 109171. [Google Scholar] [CrossRef]
- Yu, W.; Tu, Y.; Long, Z.; Liu, J.; Kong, D.; Peng, J.; Wu, H.; Zheng, G.; Zhao, J.; Chen, Y.; et al. Reactive Oxygen Species Bridge the Gap between Chronic Inflammation and Tumor Development. Oxid. Med. Cell Longev. 2022, 2022, 2606928. [Google Scholar] [CrossRef]
- Hotamisligil, G.S. Endoplasmic Reticulum Stress and the Inflammatory Basis of Metabolic Disease. Cell 2010, 140, 900–917. [Google Scholar] [CrossRef] [PubMed]
- Kaser, A.; Lee, A.; Franke, A.; Glickman, J.N.; Zeissig, S.; Tilg, H.; Nieuwenhuis, E.E.S.; Higgins, D.E.; Schreiber, S.; Glimcher, L.H.; et al. XBP1 Links ER Stress to Intestinal Inflammation and Confers Genetic Risk for Human Inflammatory Bowel Disease. Cell 2008, 134, 743–756. [Google Scholar] [CrossRef] [PubMed]
- Chevet, E.; Hetz, C.; Samali, A. Endoplasmic Reticulum Stress–Activated Cell Reprogramming in Oncogenesis. Cancer Discov. 2015, 5, 586–597. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Sun, Y.; Li, Y.; Liu, W.; Xiang, H.; Fan, L.; Sun, Q.; Xu, X.; Cai, J.; Ruan, C.; et al. Hydrogen-rich saline ameliorates the severity of l-arginine-induced acute pancreatitis in rats. Biochem. Biophys. Res. Commun. 2010, 393, 308–313. [Google Scholar] [CrossRef] [PubMed]
- Meng, J.; Yu, P.; Jiang, H.; Yuan, T.; Liu, N.; Tong, J.; Chen, H.; Bao, N.; Zhao, J. Molecular hydrogen decelerates rheumatoid arthritis progression through inhibition of oxidative stress. Am. J. Transl. Res. 2016, 8, 4472–4477. [Google Scholar]
- Hu, A.; Yamaguchi, T.; Tabuchi, M.; Ikarashi, Y.; Mizushima, A.; Kobayashi, H. A pilot study to evaluate the potential therapeutic effect of hydrogen-water bathing on atopic dermatitis in humans. Adv. Integr. Med. 2023. [CrossRef]
- Kawaguchi, M.; Satoh, Y.; Otsubo, Y.; Kazama, T. Molecular Hydrogen Attenuates Neuropathic Pain in Mice. PLoS ONE 2014, 9, e100352. [Google Scholar] [CrossRef]
- Morris, G.P.; Beck, P.L.; Herridge, M.S.; Depew, W.T.; Szewczuk, M.R.; Wallace, J.L. Hapten-induced model of chronic inflammation and ulceration in the rat colon. Gastroenterology 1989, 96, 795–803. [Google Scholar] [CrossRef] [PubMed]
- Antoniou, E.; Margonis, G.A.; Angelou, A.; Pikouli, A.; Argiri, P.; Karavokyros, I.; Papalois, A.; Pikoulis, E. The TNBS-induced colitis animal model: An overview. Ann. Med. Surg. 2016, 11, 9–15. [Google Scholar] [CrossRef]
- Huang, T.; Okauchi, T.; Hu, D.; Shigeta, M.; Wu, Y.; Wada, Y.; Hayashinaka, E.; Wang, S.; Kogure, Y.; Noguchi, K.; et al. Pain matrix shift in the rat brain following persistent colonic inflammation revealed by voxel-based statistical analysis. Mol. Pain. 2019, 15, 1744806919891327. [Google Scholar] [CrossRef]
- Rizo-Téllez, S.A.; Sekheri, M.; Filep, J.G. Myeloperoxidase: Regulation of Neutrophil Function and Target for Therapy. Antioxidants 2022, 11, 2302. [Google Scholar] [CrossRef] [PubMed]
- Bielefeldt, K.; Davis, B.; Binion, D.G. Pain and inflammatory bowel disease. Inflamm. Bowel Dis. 2009, 15, 778–788. [Google Scholar] [CrossRef] [PubMed]
- Srinath, A.; Young, E.; Szigethy, E. Pain management in patients with inflammatory bowel disease: Translational approaches from bench to bedside. Inflamm. Bowel Dis. 2014, 20, 2433–2449. [Google Scholar] [CrossRef] [PubMed]
- Liu, Q.; Liang, X.; Wang, Q.; Wilson, E.N.; Lam, R.; Wang, J.; Kong, W.; Tsai, C.; Pan, T.; Larkin, P.B.; et al. PGE2 signaling via the neuronal EP2 receptor increases injury in a model of cerebral ischemia. Proc. Natl. Acad. Sci. USA 2019, 116, 10019–10024. [Google Scholar] [CrossRef] [PubMed]
- Dejani, N.N.; Orlando, A.B.; Nino, V.E.; Penteado, L.A.; Verdan, F.F.; Bazzano, J.M.R.; Codo, A.C.; Salina, A.C.G.; Saraiva, A.C.; Avelar, M.R.; et al. Intestinal host defense outcome is dictated by PGE2 production during efferocytosis of infected cells. Proc. Natl. Acad. Sci. USA 2018, 115, E8469–E8478. [Google Scholar] [CrossRef] [PubMed]
- Crittenden, S.; Goepp, M.; Pollock, J.; Robb, C.T.; Smyth, D.J.; Zhou, Y.; Andrews, R.; Tyrrell, V.; Gkikas, K.; Adima, A.; et al. Prostaglandin E2 promotes intestinal inflammation via inhibiting microbiota-dependent regulatory T cells. Sci. Adv. 2021, 7, eabd7954. [Google Scholar] [CrossRef]
- Jones, R.C.; Xu, L.; Gebhart, G.F. The mechanosensitivity of mouse colon afferent fibers and their sensitization by inflammatory mediators require transient receptor potential vanilloid 1 and acid-sensing ion channel 3. J. Neurosci. 2005, 25, 10981–10989. [Google Scholar] [CrossRef]
- Karai, L.; Brown, D.C.; Mannes, A.J.; Connelly, S.T.; Brown, J.; Gandal, M.; Wellisch, O.M.; Neubert, J.K.; Olah, Z.; Iadarola, M.J. Deletion of vanilloid receptor 1-expressing primary afferent neurons for pain control. J. Clin. Investig. 2004, 113, 1344–1352. [Google Scholar] [CrossRef]
- Delafoy, L.; Gelot, A.; Ardid, D.; Eschalier, A.; Bertrand, C.; Doherty, A.M.; Diop, L. Interactive involvement of brain derived neurotrophic factor, nerve growth factor, and calcitonin gene related peptide in colonic hypersensitivity in the rat. Gut 2006, 55, 940–945. [Google Scholar] [CrossRef][Green Version]
- Ellis, A.; Bennett, D.L.H. Neuroinflammation and the generation of neuropathic pain. Br. J. Anaesth. 2013, 111, 26–37. [Google Scholar] [CrossRef]
- Menghini, P.; Corridoni, D.; Buttó, L.F.; Osme, A.; Shivaswamy, S.; Lam, M.; Bamias, G.; Pizarro, T.T.; Rodriguez-Palacios, A.; Dinarello, C.A.; et al. Neutralization of IL-1α ameliorates Crohn’s disease-like ileitis by functional alterations of the gut microbiome. Proc. Natl. Acad. Sci. USA 2019, 116, 26717–26726. [Google Scholar] [CrossRef]
- Ey, B.; Eyking, A.; Klepak, M.; Salzman, N.H.; Göthert, J.R.; Rünzi, M.; Schmid, K.W.; Gerken, G.; Podolsky, D.K.; Cario, E. Loss of TLR2 Worsens Spontaneous Colitis in MDR1A Deficiency through Commensally Induced Pyroptosis. J. Immunol. 2013, 190, 5676–5688. [Google Scholar] [CrossRef]
- Rosen, J.M.; Yaggie, R.E.; Woida, P.J.; Miller, R.J.; Schaefer, A.J.; Klumpp, D.J. TRPV1 and the MCP-1/CCR2 Axis Modulate Post-UTI Chronic Pain. Sci. Rep. 2018, 8, 7188. [Google Scholar] [CrossRef]
- Abbadie, C.; Lindia, J.A.; Cumiskey, A.M.; Peterson, L.B.; Mudgett, J.S.; Bayne, E.K.; DeMartino, J.A.; MacIntyre, D.E.; Forrest, M.J. Impaired neuropathic pain responses in mice lacking the chemokine receptor CCR2. Proc. Natl. Acad. Sci. USA 2003, 100, 7947–7952. [Google Scholar] [CrossRef]
- Cui, M.; Honore, P.; Zhong, C.; Gauvin, D.; Mikusa, J.; Hernandez, G.; Chandran, P.; Gomtsyan, A.; Brown, B.; Bayburt, E.K.; et al. TRPV1 receptors in the CNS play a key role in broad-spectrum analgesia of TRPV1 antagonists. J. Neurosci. 2006, 26, 9385–9393. [Google Scholar] [CrossRef] [PubMed]
- Ma, F.; Zhang, L.; Westlund, K.N. Reactive oxygen species mediate TNFR1 increase after TRPV1 activation in mouse DRG neurons. Mol. Pain. 2009, 5, 31. [Google Scholar] [CrossRef]
- Westlund, K.N.; Kochukov, M.Y.; Lu, Y.; McNearney, T.A. Impact of central and peripheral TRPV1 and ROS levels on proinflammatory mediators and nociceptive behavior. Mol. Pain. 2010, 6, 46. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Zhou, F.; Chen, Q.; Kang, A.; Lu, M.; Liu, W.; Zang, X.; Wang, G.; Zhang, J. Chronic inflammation up-regulates P-gp in peripheral mononuclear blood cells via the STAT3/Nf-kappab pathway in 2,4,6-trinitrobenzene sulfonic acid-induced colitis mice. Sci. Rep. 2015, 5, 13558. [Google Scholar] [CrossRef]
- Hofma, B.R.; Wardill, H.R.; Mavrangelos, C.; Campaniello, M.A.; Dimasi, D.; Bowen, J.M.; Smid, S.D.; Bonder, C.S.; Beckett, E.A.; Hughes, P.A. Colonic migrating motor complexes are inhibited in acute tri-nitro benzene sulphonic acid colitis. PLoS ONE 2018, 13, e0199394. [Google Scholar] [CrossRef]
- Park, J.; Min, J.; Kim, B.; Chae, U.; Yun, J.W.; Choi, M.; Kong, I.; Chang, K.; Lee, D. Mitochondrial ROS govern the LPS-induced pro-inflammatory response in microglia cells by regulating MAPK and NF-κB pathways. Neurosci. Lett. 2015, 584, 191–196. [Google Scholar] [CrossRef] [PubMed]
- Esworthy, R.S.; Kim, B.W.; Chow, J.; Shen, B.; Doroshow, J.H.; Chu, F. Nox1 causes ileocolitis in mice deficient in glutathione peroxidase-1 and -2. Free Radic. Biol. Med. 2014, 68, 315–325. [Google Scholar] [CrossRef]
- Spiers, J.G.; Chen, H.C.; Sernia, C.; Lavidis, N.A. Activation of the hypothalamic-pituitary-adrenal stress axis induces cellular oxidative stress. Front. Neurosci. 2015, 8, 456. [Google Scholar] [CrossRef]
- McEwen, B.S.; Gianaros, P.J. Central role of the brain in stress and adaptation: Links to socioeconomic status, health, and disease. Ann. N. Y. Acad. Sci. 2010, 1186, 190–222. [Google Scholar] [CrossRef] [PubMed]
- Sato, H.; Takahashi, T.; Sumitani, K.; Takatsu, H.; Urano, S. Glucocorticoid Generates ROS to Induce Oxidative Injury in the Hippocampus, Leading to Impairment of Cognitive Function of Rats. J. Clin. Biochem. Nutr. 2010, 47, 224–232. [Google Scholar] [CrossRef] [PubMed]
- Asaba, K.; Iwasaki, Y.; Yoshida, M.; Asai, M.; Oiso, Y.; Murohara, T.; Hashimoto, K. Attenuation by Reactive Oxygen Species of Glucocorticoid Suppression on Proopiomelanocortin Gene Expression in Pituitary Corticotroph Cells. Endocrinology 2004, 145, 39–42. [Google Scholar] [CrossRef]
- Zhao, X.; Zhao, Z.; Yang, D.; Cao, L.; Zhang, L.; Ji, J.; Gu, J.; Huang, J.; Sun, X. Activation of ATP-sensitive potassium channel by iptakalim normalizes stress-induced HPA axis disorder and depressive behaviour by alleviating inflammation and oxidative stress in mouse hypothalamus. Brain Res. Bull. 2017, 130, 146–155. [Google Scholar] [CrossRef]
- Mikulska, J.; Juszczyk, G.; Gawrońska-Grzywacz, M.; Herbet, M. HPA Axis in the Pathomechanism of Depression and Schizophrenia: New Therapeutic Strategies Based on Its Participation. Brain Sci. 2021, 11, 1298. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Su, W.; Chen, Y.; Wu, T.; Gong, H.; Shen, X.; Wang, Y.; Sun, X.; Jiang, C. Effects of hydrogen-rich water on depressive-like behavior in mice. Sci. Rep. 2016, 6, 23742. [Google Scholar] [CrossRef]
- Herman, J.P.; McKlveen, J.M.; Ghosal, S.; Kopp, B.; Wulsin, A.; Makinson, R.; Scheimann, J.; Myers, B. Regulation of the Hypothalamic-Pituitary-Adrenocortical Stress Response. Compr. Physiol. 2016, 6, 603–621. [Google Scholar] [CrossRef]
- Shintani, F.; Nakaki, T.; Kanba, S.; Sato, K.; Yagi, G.; Shiozawa, M.; Aiso, S.; Kato, R.; Asai, M. Involvement of interleukin-1 in immobilization stress-induced increase in plasma adrenocorticotropic hormone and in release of hypothalamic monoamines in the rat. J. Neurosci. 1995, 15, 1961–1970. [Google Scholar] [CrossRef]
- Schmidt, E.D.; Janszen, A.W.; Wouterlood, F.G.; Tilders, F.J. Interleukin-1-induced long-lasting changes in hypothalamic corticotropin-releasing hormone (CRH)--neurons and hyperresponsiveness of the hypothalamus-pituitary-adrenal axis. J. Neurosci. 1995, 15, 7417–7426. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Kelly, A.; Lynch, A.; Vereker, E.; Nolan, Y.; Queenan, P.; Whittaker, E.; O’Neill, L.A.; Lynch, M.A. The anti-inflammatory cytokine, interleukin (IL)-10, blocks the inhibitory effect of IL-1 beta on long term potentiation. J. Biol. Chem. 2001, 276, 45564–45572. [Google Scholar] [CrossRef] [PubMed]
- Kil, I.S.; Lee, S.K.; Ryu, K.W.; Woo, H.A.; Hu, M.; Bae, S.H.; Rhee, S.G. Feedback control of adrenal steroidogenesis via H2O2 dependent reversible inactivation of peroxiredoxin iii in mitochondria. Mol. Cell 2012, 46, 584–594. [Google Scholar] [CrossRef] [PubMed]
- Heim, C.; Ehlert, U.; Hellhammer, D.H. The potential role of hypocortisolism in the pathophysiology of stress-related bodily disorders. Psychoneuroendocrinology 2000, 25, 1–35. [Google Scholar] [CrossRef] [PubMed]
- Ji, X.; Tian, Y.; Xie, K.; Liu, W.; Qu, Y.; Fei, Z. Protective effects of hydrogen-rich saline in a rat model of traumatic brain injury via reducing oxidative stress. J. Surg. Res. 2012, 178, e9–e16. [Google Scholar] [CrossRef]
- Tian, R.; Hou, Z.; Hao, S.; Wu, W.; Mao, X.; Tao, X.; Lu, T.; Liu, B. Hydrogen-rich water attenuates brain damage and inflammation after traumatic brain injury in rats. Brain Res. 2016, 1637, 1–13. [Google Scholar] [CrossRef]
- Li, J.; Wang, C.; Zhang, J.H.; Cai, J.; Cao, Y.; Sun, X. Hydrogen-rich saline improves memory function in a rat model of amyloid-beta-induced Alzheimer’s disease by reduction of oxidative stress. Brain Res. 2010, 1328, 152–161. [Google Scholar] [CrossRef]
- Sato, Y.; Kajiyama, S.; Amano, A.; Kondo, Y.; Sasaki, T.; Handa, S.; Takahashi, R.; Fukui, M.; Hasegawa, G.; Nakamura, N.; et al. Hydrogen-rich pure water prevents superoxide formation in brain slices of vitamin C-depleted SMP30/GNL knockout mice. Biochem. Biophys. Res. Commun. 2008, 375, 346–350. [Google Scholar] [CrossRef]
- Xiao, H.; Li, Y.; Luo, D.; Dong, J.; Zhou, L.; Zhao, S.; Zheng, Q.; Wang, H.; Cui, M.; Fan, S. Hydrogen-water ameliorates radiation-induced gastrointestinal toxicity via MyD88’s effects on the gut microbiota. Exp. Mol. Med. 2018, 50, e433. [Google Scholar] [CrossRef]
- Zhou, P.; Lin, B.; Wang, P.; Pan, T.; Wang, S.; Chen, W.; Cheng, S.; Liu, S. The healing effect of hydrogen-rich water on acute radiation-induced skin injury in rats. J. Radiat. Res. 2019, 60, 17–22. [Google Scholar] [CrossRef] [PubMed]
- Kawasaki, H.; Guan, J.; Tamama, K. Hydrogen gas treatment prolongs replicative lifespan of bone marrow multipotential stromal cells in vitro while preserving differentiation and paracrine potentials. Biochem. Biophys. Res. Commun. 2010, 397, 608–613. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
| Benefits | Subject | Outline of Effects | References |
|---|---|---|---|
| Antioxidant Effects | IBD model rats | ROS↓, antioxidant defense↑ | [6] |
| Stress model rats | ROS↓, antioxidant defense↑ | [17] | |
| In vitro | ROS↓ | [42,43,46,47,48,52] | |
| Gastric injury model rats | Lipid peroxidation↓ | [50] | |
| In vitro | Ethanol-induced cytotoxicity↓ | [53] | |
| End-stage renal disease patients | Antioxidant defense↑ | [54] | |
| Neuropathic pain model mouse | Antioxidant defense↑ | [8] | |
| Inflammatory pain model mouse | Antioxidant defense↑ | [5] | |
| Anti-inflammatory Effects | IBD model rats | Inflammatory cell infiltration↓, | [6] |
| Proinflammatory cytokines↓ | |||
| Atopic dermatitis model mouse | Chemokine↓, cytokines↓ | [15] | |
| Infiltration of mast cells↓ | |||
| Stress model rats | Proinflammatory cytokines↓ | [17] | |
| Gastric injury model rats | Proinflammatory cytokines↓ | [7,50] | |
| Neuroinflammation model mouse/In vitro | Proinflammatory cytokines↓ | [51] | |
| Anti-inflammatory cytokines↑ | |||
| Microglial activation↓ | |||
| End-stage renal disease patients | Inflammatory markers↓ | [54] | |
| Inflammatory pain model mouse | Inflammatory mediator↓ | [5] | |
| Analgesic Effects | IBD model rats | Abdominal pain↓ | [6] |
| Neuropathic pain model mouse | Allodynia and hyperalgesia↓ | [8] | |
| Inflammatory pain model mouse | Mechanical allodynia↓, Thermal hyperalgesia↓ | [5] | |
| Other Effects | |||
| Anti-fatigue effect | Neuroinflammation model | Sickness behavior↓, recovery↑ | [51] |
| Anti-apoptotic effect | Gastric injury model rats | Epithelial cell apoptosis↓ | [7] |
| In vitro | Neuronal cell apoptosis↓ | [52] | |
| Neuropathic pain model mouse | Cellular death marker↓ | [8] | |
| Anti-diabetic effect | Type 2 diabetes patients | Lactate↓ | [11] |
| Anti-tumor effect | In vitro | Tumor growth and invasion↓ | [46] |
| Tumor cell transformation↓ | [47] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hu, D.; Kabayama, S.; Watanabe, Y.; Cui, Y. Health Benefits of Electrolyzed Hydrogen Water: Antioxidant and Anti-Inflammatory Effects in Living Organisms. Antioxidants 2024, 13, 313. https://doi.org/10.3390/antiox13030313
Hu D, Kabayama S, Watanabe Y, Cui Y. Health Benefits of Electrolyzed Hydrogen Water: Antioxidant and Anti-Inflammatory Effects in Living Organisms. Antioxidants. 2024; 13(3):313. https://doi.org/10.3390/antiox13030313
Chicago/Turabian StyleHu, Di, Shigeru Kabayama, Yasuyoshi Watanabe, and Yilong Cui. 2024. "Health Benefits of Electrolyzed Hydrogen Water: Antioxidant and Anti-Inflammatory Effects in Living Organisms" Antioxidants 13, no. 3: 313. https://doi.org/10.3390/antiox13030313
APA StyleHu, D., Kabayama, S., Watanabe, Y., & Cui, Y. (2024). Health Benefits of Electrolyzed Hydrogen Water: Antioxidant and Anti-Inflammatory Effects in Living Organisms. Antioxidants, 13(3), 313. https://doi.org/10.3390/antiox13030313