
Selenomethionine Supplementation Mitigates Liver Dysfunction, Oxidative Injury and Apoptosis through Enhancing Antioxidant Capacity and Inhibiting JNK MAPK Pathway in Piglets Fed Deoxynivalenol-Contaminated Diets


Abstract
1. Introduction
2. Materials and Methods
2.1. DON and Se-Met Information
2.2. Animals and Experimental Design
2.3. Sample Collection
2.4. Laboratory Analysis
2.4.1. Hepatic Histopathological Analysis
2.4.2. Serum Biochemical Parameters Related to Hepatic Functions
2.4.3. ROS Level in The Liver
2.4.4. Hepatic Antioxidant Parameters
2.4.5. RT-qPCR Analysis
2.4.6. Western Blot Analysis
2.5. Statistical Analysis
3. Results
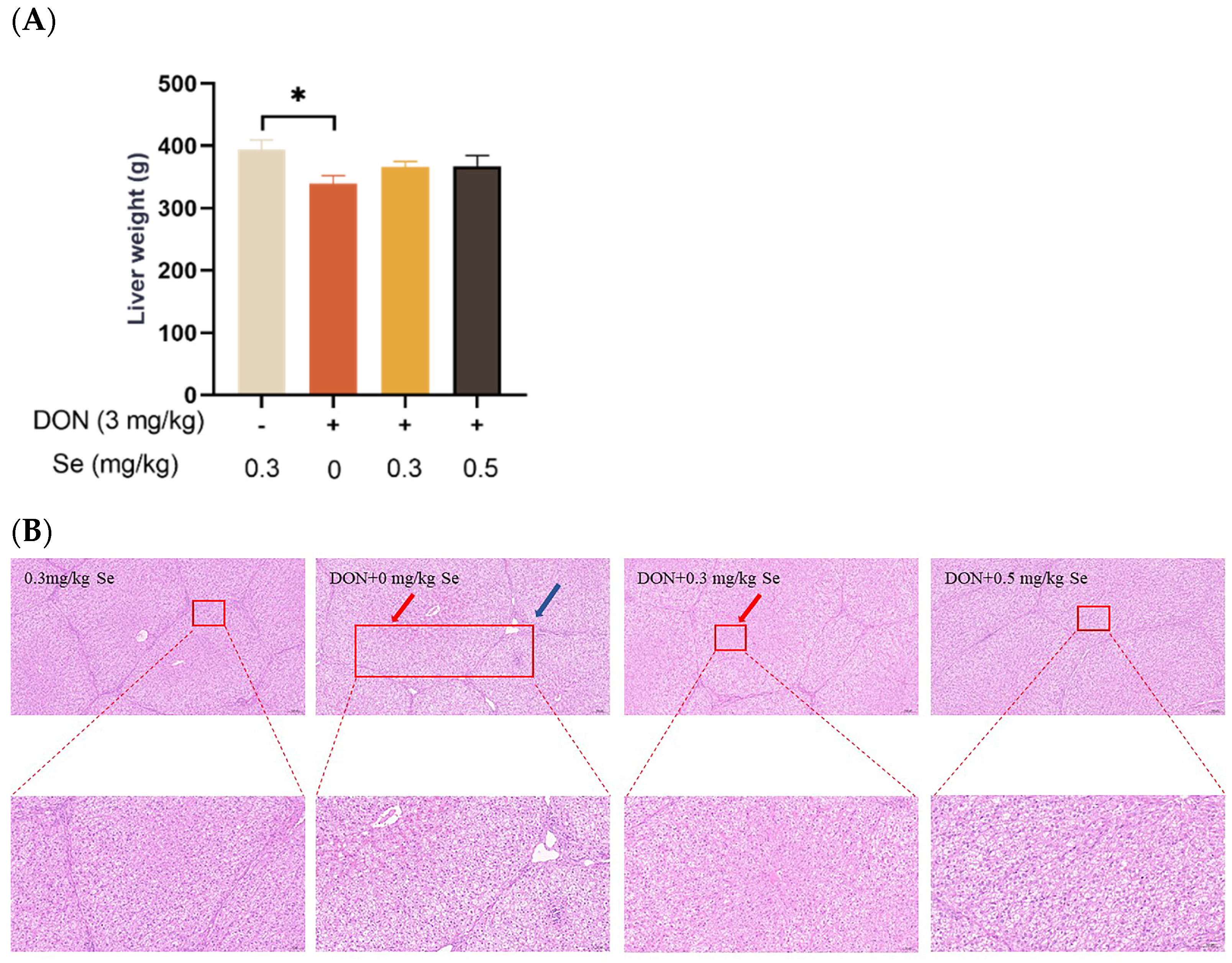
3.1. Liver Weight and Histopathology
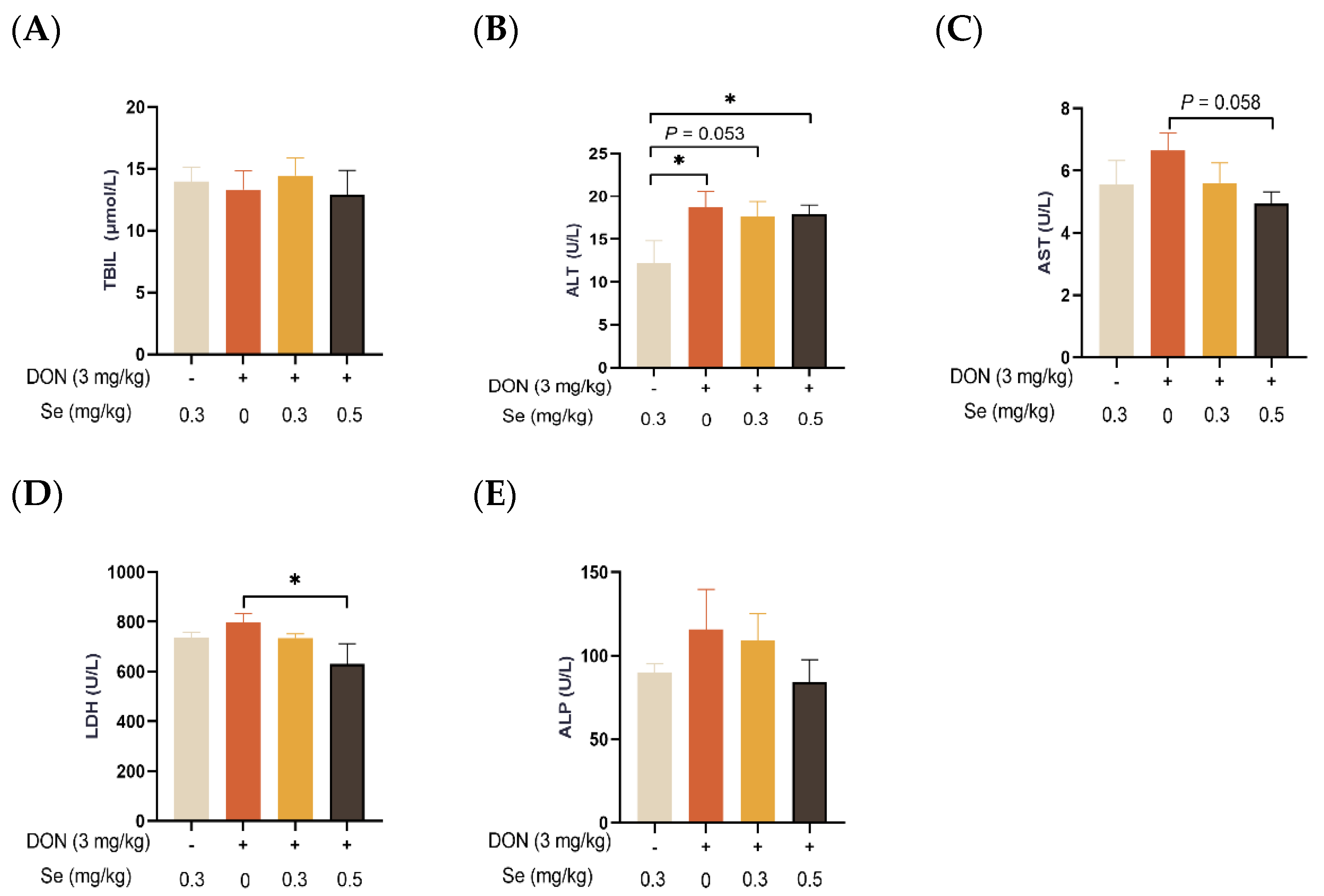
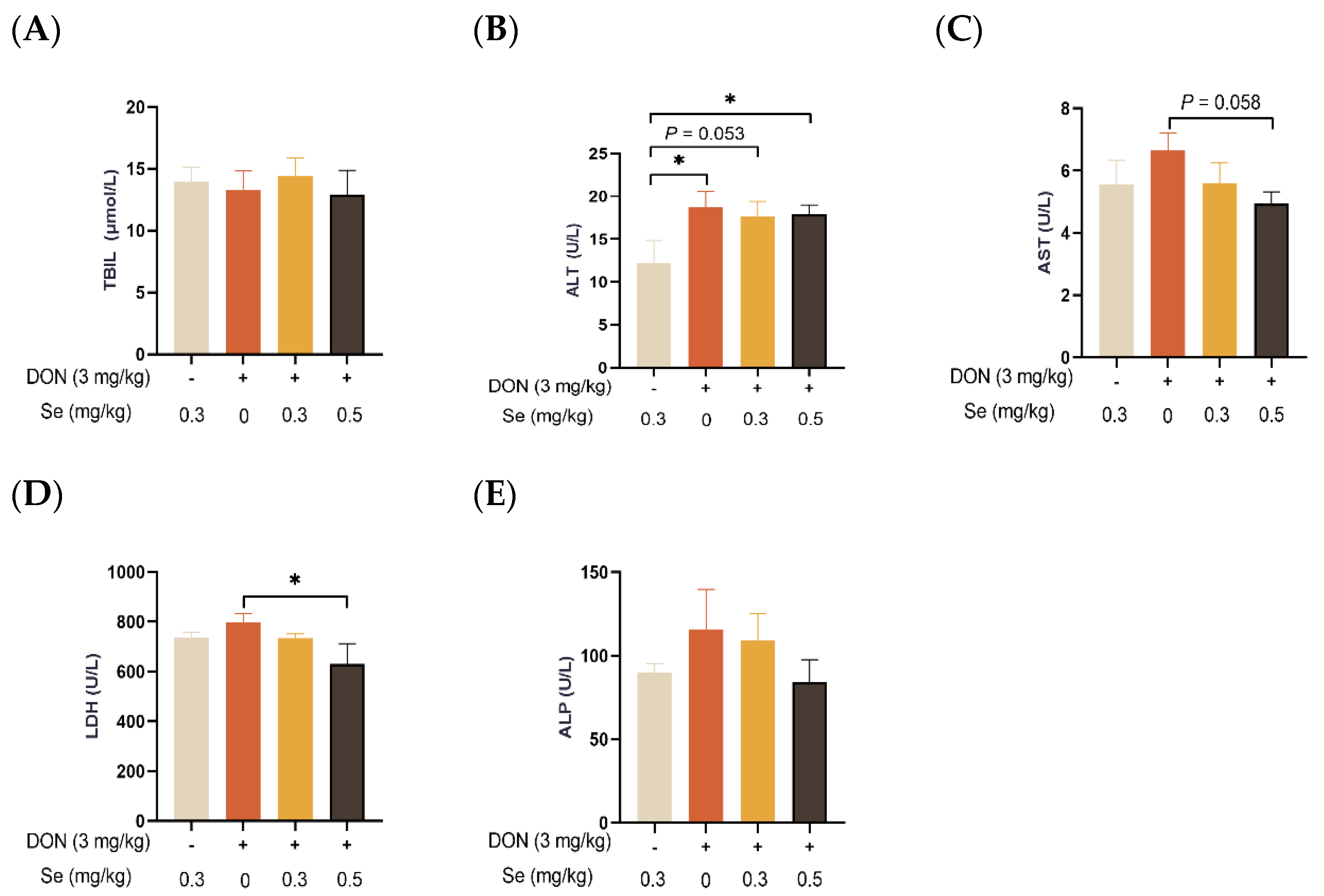
3.2. Serum Biochemical Parameters Related to Hepatic Functions
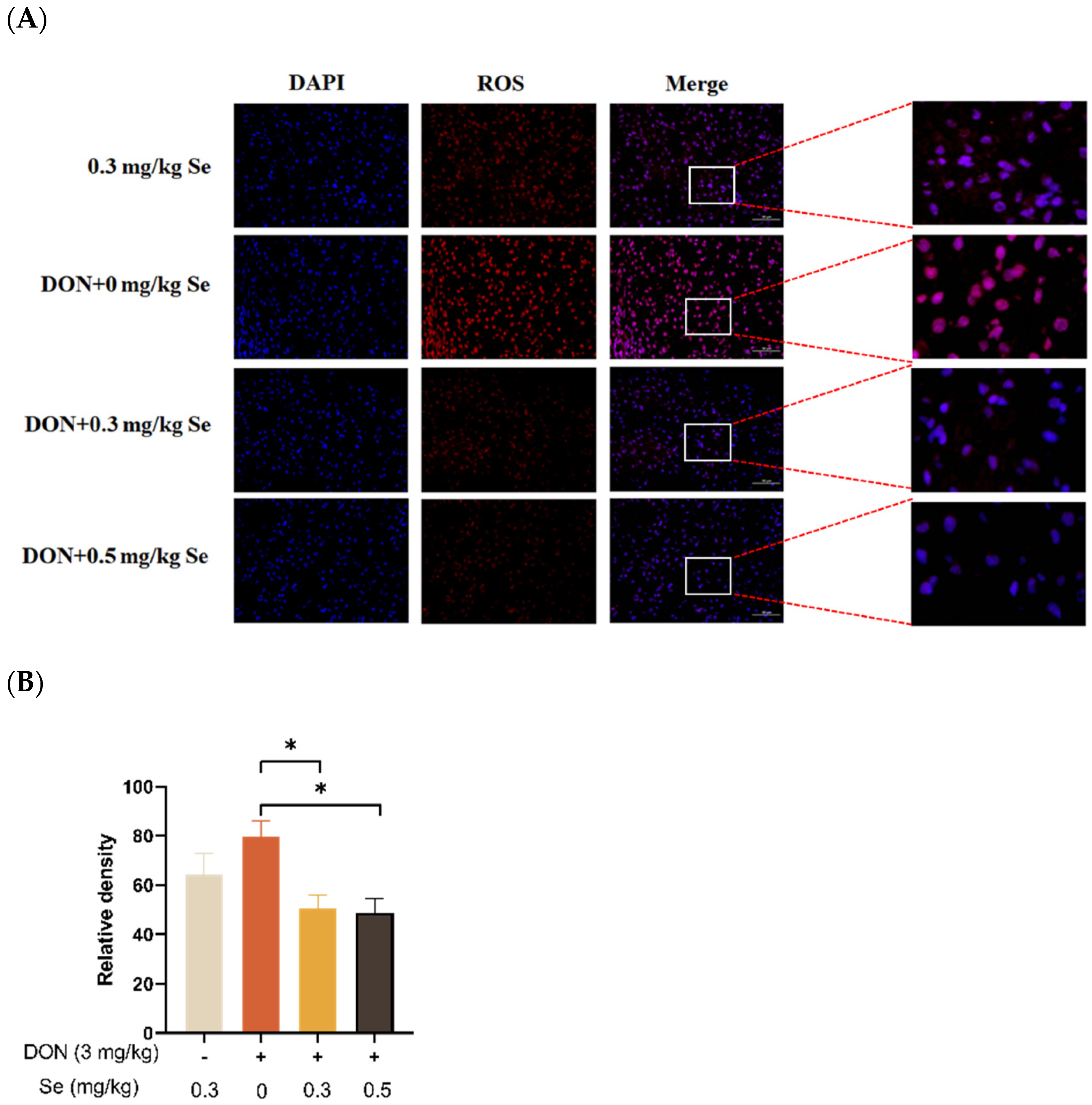
3.3. Hepatic ROS Level
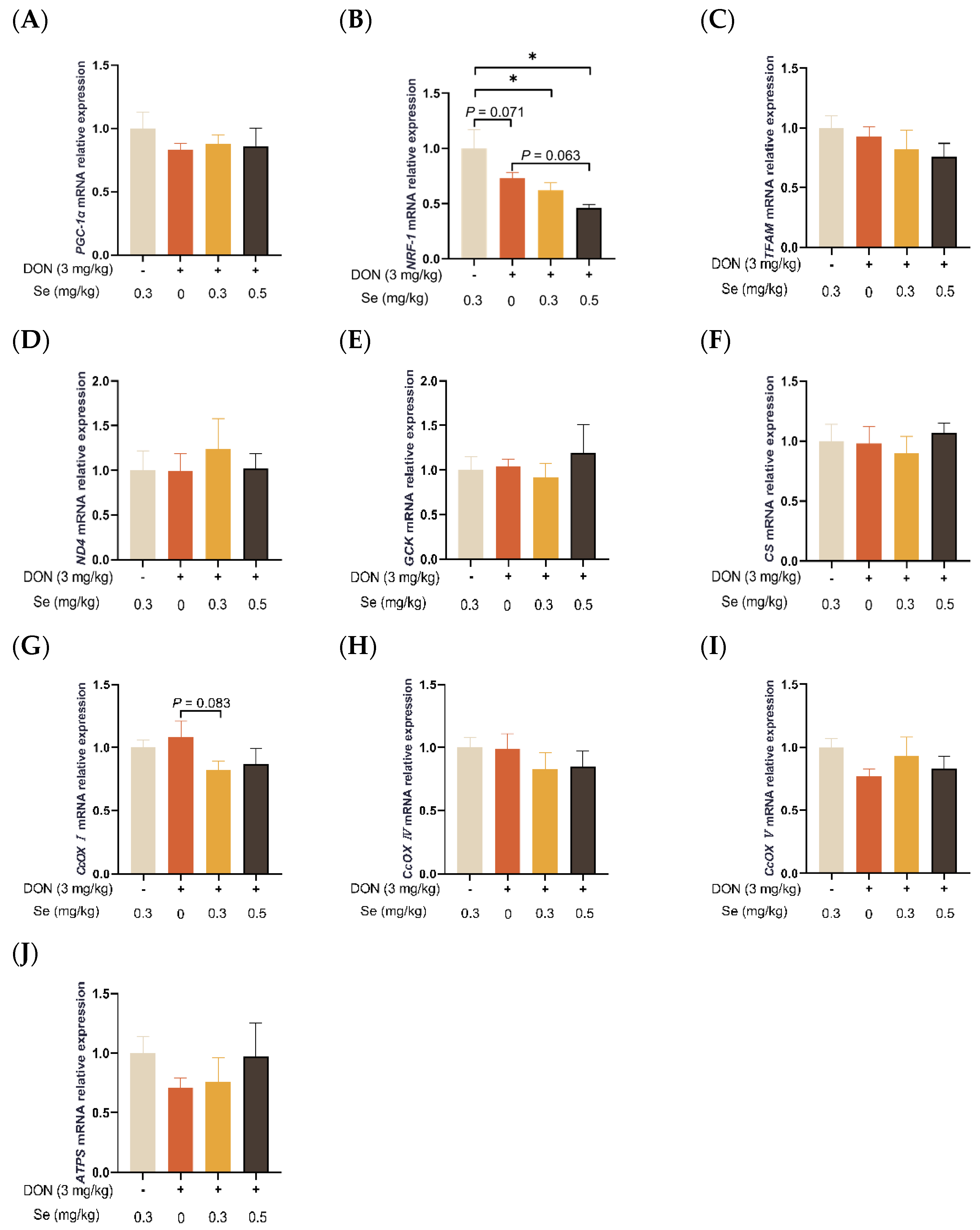
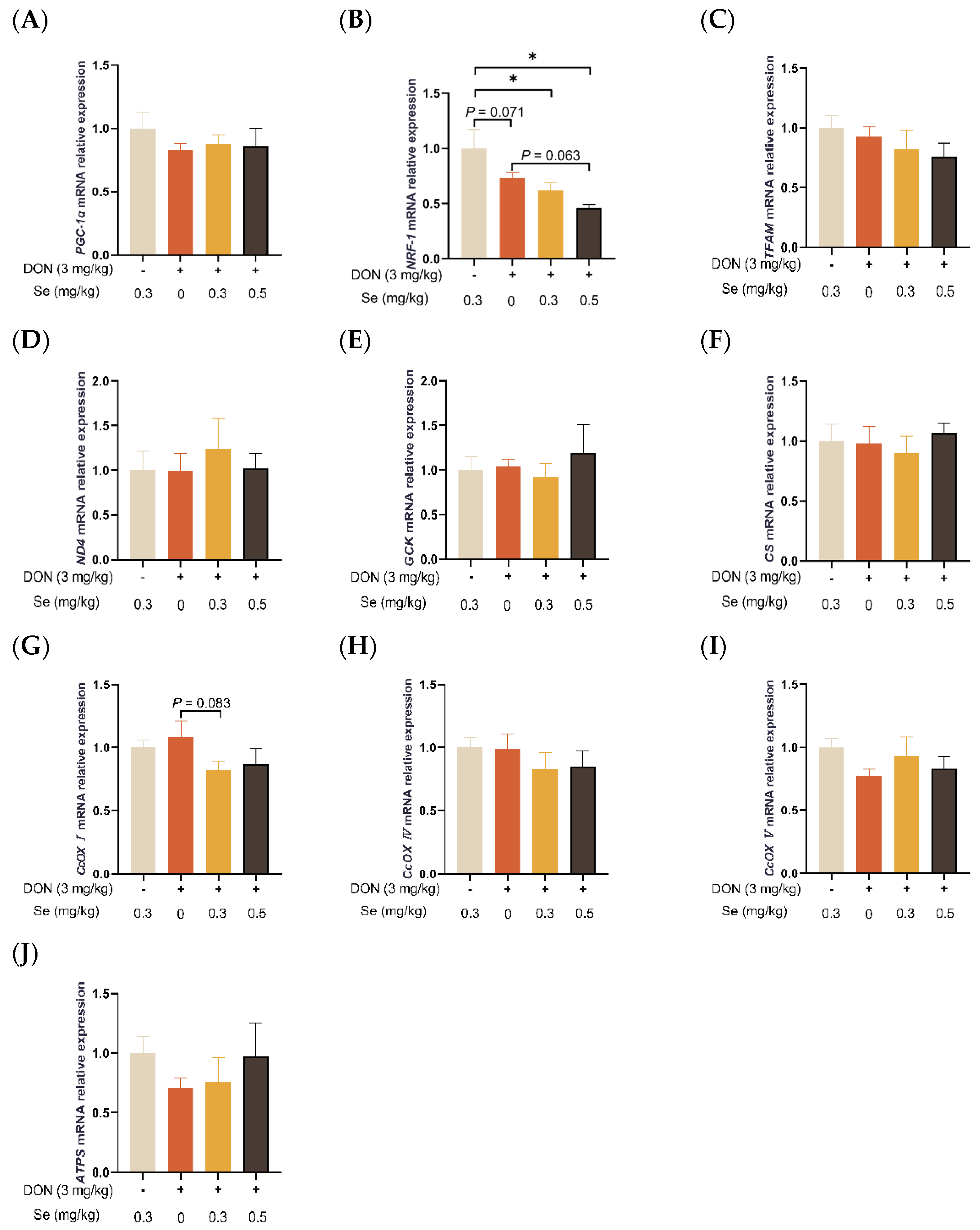
3.4. Mitochondrial Function-Related Genes Expression in Liver
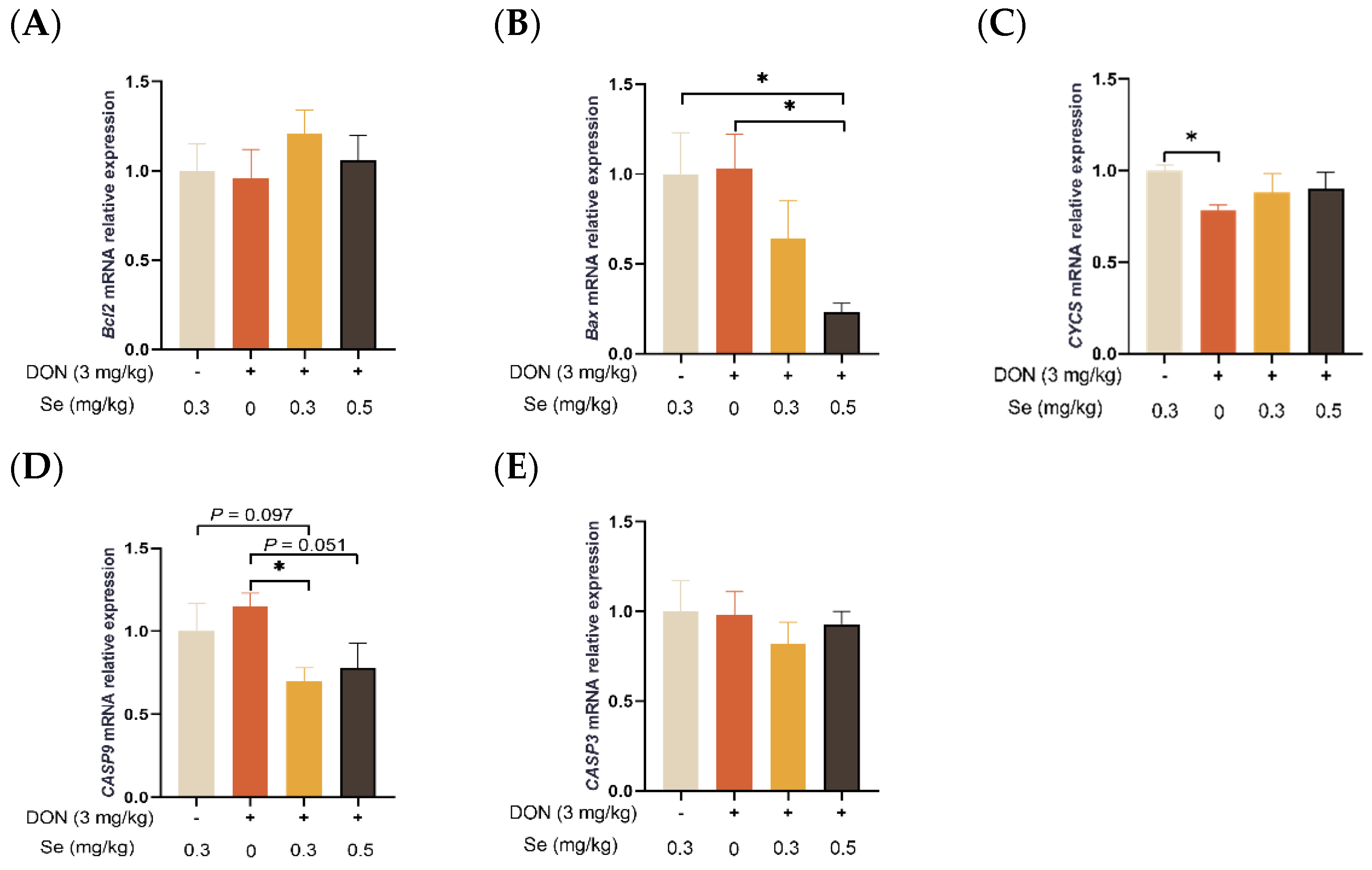
3.5. Apoptosis-Related Genes Expression in Liver
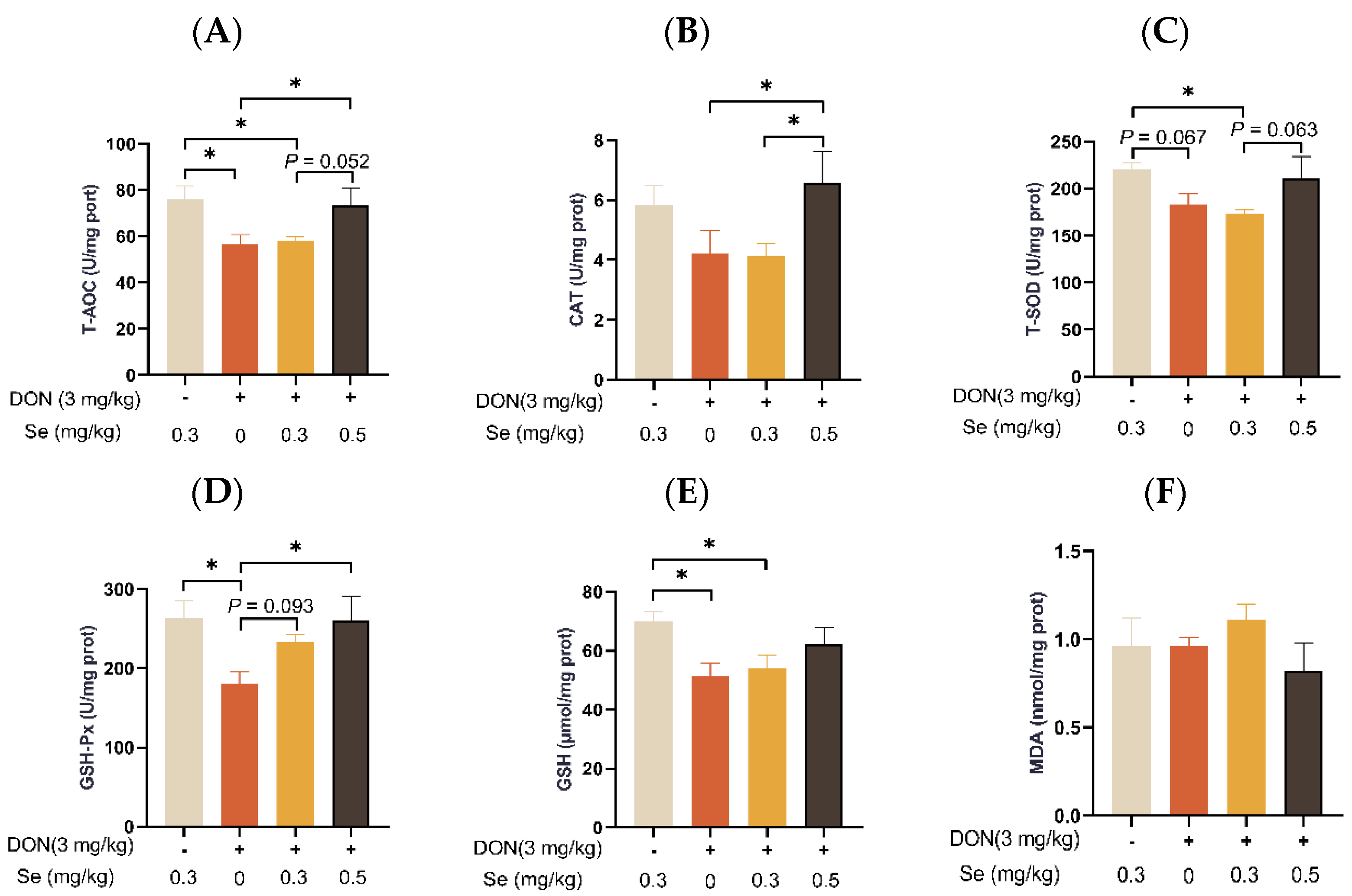
3.6. Hepatic Antioxidant Capacity
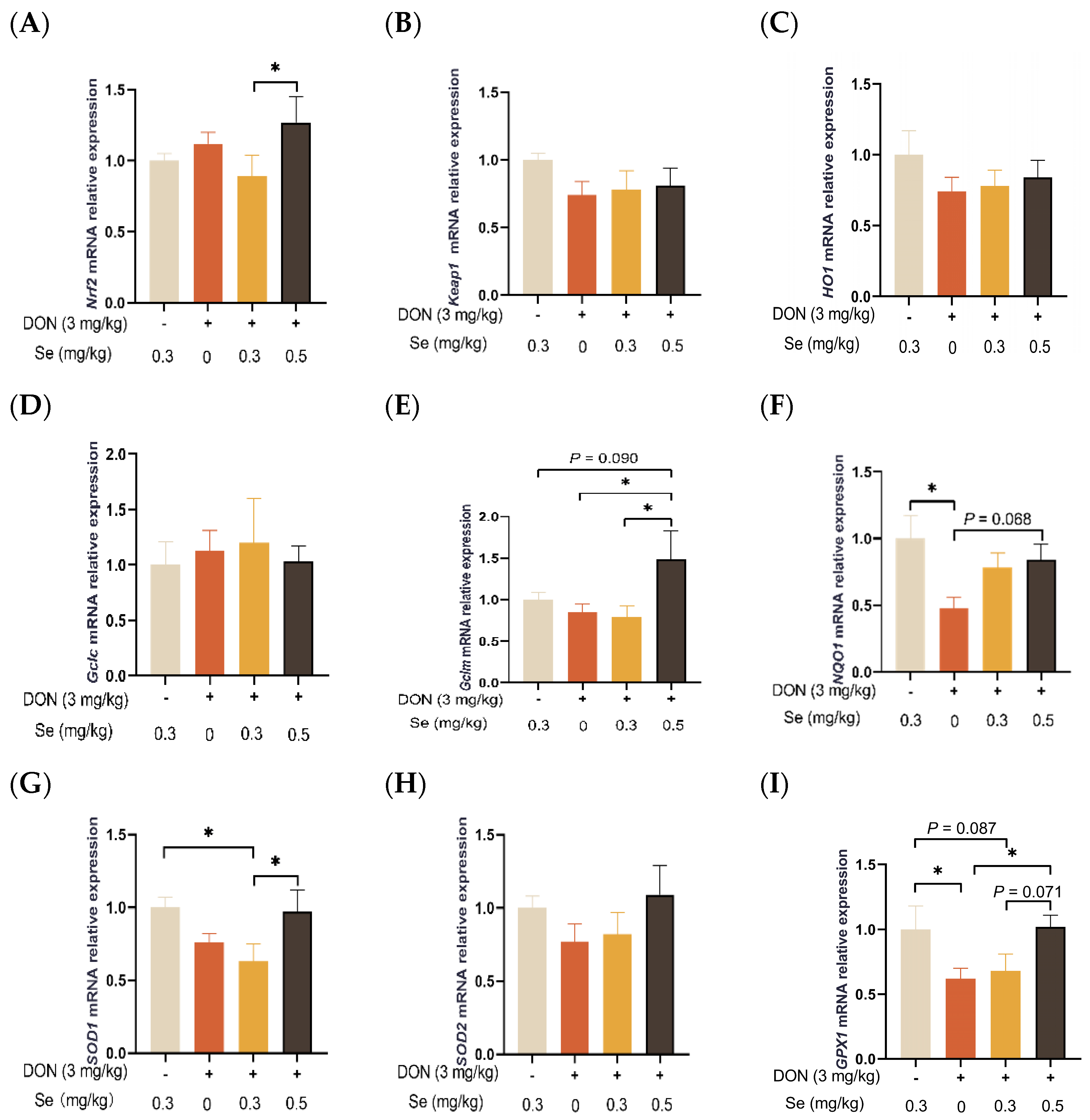
3.7. Antioxidant-Related Genes Expression in the Liver
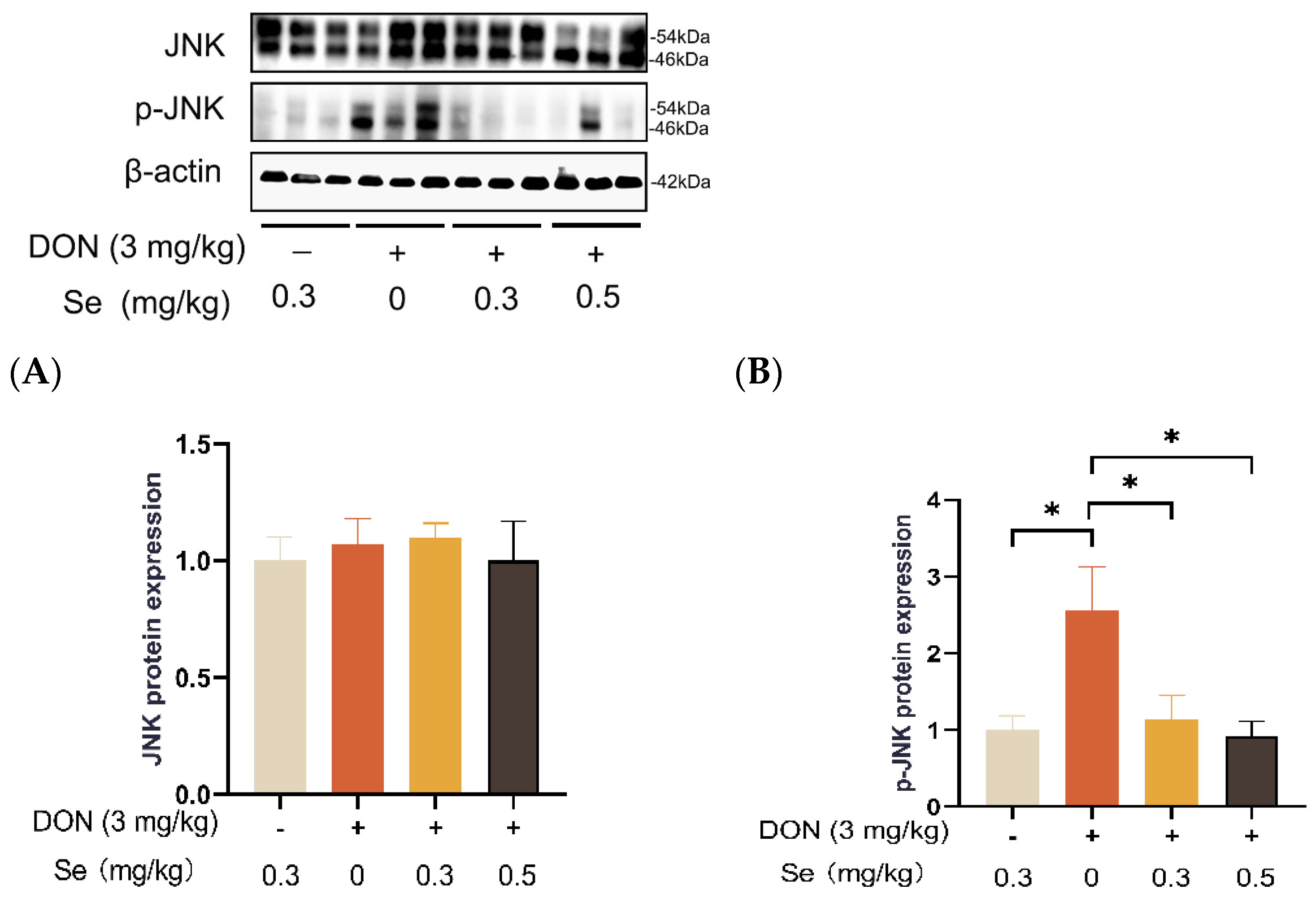
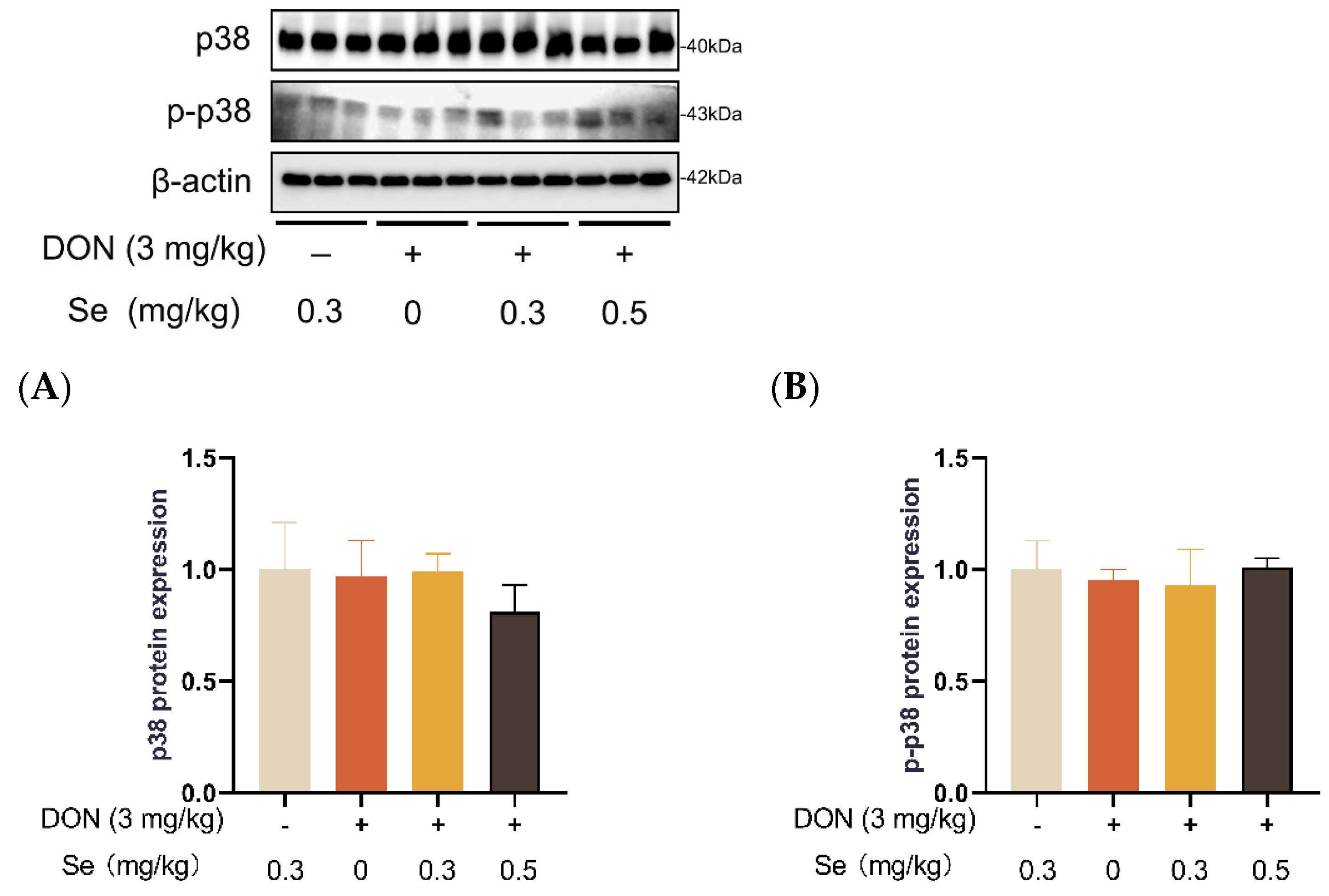
3.8. Protein Expression of Key Proteins in MAPK Pathways
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Zha, A.; Yuan, D.; Cui, Z.; Qi, M.; Liao, S.; Liao, P.; Tan, B. The evaluation of the antioxidant and intestinal protective effects of baicalin-copper in deoxynivalenol-challenged piglets. Oxid. Med. Cell Longev. 2020, 2020, 5363546. [Google Scholar] [CrossRef]
- Chen, J.; Huang, Z.; Cao, X.; Chen, X.; Zou, T.; You, J. Plant-derived polyphenols as Nrf2 activators to counteract oxidative stress and intestinal toxicity induced by deoxynivalenol in swine: An emerging research direction. Antioxidants 2022, 11, 2379. [Google Scholar] [CrossRef] [PubMed]
- Zhao, L.; Li, X.; Ji, C.; Rong, X.; Liu, S.; Zhang, J.; Ma, Q. Protective effect of Devosia sp. ANSB714 on growth performance, serum chemistry, immunity function and residues in kidneys of mice exposed to deoxynivalenol. Food Chem. Toxicol. 2016, 92, 143–149. [Google Scholar] [CrossRef] [PubMed]
- Ma, K.; Bai, Y.; Li, J.; Ren, Z.; Li, J.; Zhang, J.; Shan, A. Lactobacillus rhamnosus GG ameliorates deoxynivalenol-induced kidney oxidative damage and mitochondrial injury in weaned piglets. Food Funct. 2022, 13, 3905–3916. [Google Scholar] [CrossRef]
- Rotter, B.A. Invited review: Toxicology of deoxynivalenol (vomitoxin). J. Toxicol. Environ. Health 1996, 48, 1–34. [Google Scholar] [CrossRef]
- Lunney, J.K.; Van Goor, A.; Walker, K.E.; Hailstock, T.; Franklin, J.; Dai, C. Importance of the pig as a human biomedical model. Sci. Transl. Med. 2021, 13, eabd5758. [Google Scholar] [CrossRef]
- Ruan, H.; Lu, Q.; Wu, J.; Qin, J.; Sui, M.; Sun, X.; Shi, Y.; Luo, J.; Yang, M. Hepatotoxicity of food-borne mycotoxins: Molecular mechanism, anti-hepatotoxic medicines and target prediction. Crit. Rev. Food Sci. Nutr. 2022, 62, 2281–2308. [Google Scholar] [CrossRef]
- Peng, Z.; Liao, Y.; Wang, X.; Chen, L.; Wang, L.; Qin, C.; Wang, Z.; Cai, M.; Hu, J.; Li, D.; et al. Heme oxygenase-1 regulates autophagy through carbon–oxygen to alleviate deoxynivalenol-induced hepatic damage. Arch. Toxicol. 2020, 94, 573–588. [Google Scholar] [CrossRef]
- Ji, X.; Tang, Z.; Zhang, F.; Zhou, F.; Wu, Y.; Wu, D. Dietary taurine supplementation counteracts deoxynivalenol-induced liver injury via alleviating oxidative stress, mitochondrial dysfunction, apoptosis, and inflammation in piglets. Ecotoxicol. Environ. Saf. 2023, 253, 114705. [Google Scholar] [CrossRef]
- Hou, S.; Ma, J.; Cheng, Y.; Wang, H.; Sun, J.; Yan, Y. The toxicity mechanisms of DON to humans and animals and potential biological treatment strategies. Crit. Rev. Food Sci. Nutr. 2023, 63, 790–812. [Google Scholar] [CrossRef]
- Peng, Z.; Chen, L.; Nüssler, A.K.; Liu, L.; Yang, W. Current sights for mechanisms of deoxynivalenol-induced hepatotoxicity and prospective views for future scientific research: A mini review. J. Appl. Toxicol. 2017, 37, 518–529. [Google Scholar] [CrossRef] [PubMed]
- Zeng, H.; Combs, G.F. Selenium as an anticancer nutrient: Roles in cell proliferation and tumor cell invasion. J. Nutr. Biochem. 2008, 19, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Wang, H.; Zhang, M.; Wang, J.; Zhang, Z.; Wang, Y.; Sun, Y.; Zhang, Z. Protective effect of selenomethionine on T-2 toxin-induced liver injury in New Zealand rabbits. BMC Vet. Res. 2021, 17, 153. [Google Scholar] [CrossRef] [PubMed]
- Díaz Huerta, V.; Fernández Sánchez, M.L.; Sanz-Medel, A. An attempt to differentiate HPLC-ICP-MS selenium speciation in natural and selenised agaricus mushrooms using different species extraction procedures. Anal. Bioanal. Chem. 2006, 384, 902–907. [Google Scholar] [CrossRef] [PubMed]
- Papp, L.V.; Lu, J.; Holmgren, A.; Khanna, K.K. From selenium to selenoproteins: Synthesis, identity, and their role in human health. Antioxid. Redox Signal. 2007, 9, 775–806. [Google Scholar] [CrossRef] [PubMed]
- Zhu, C.; Liang, S.; Zan, G.; Wang, X.; Gao, C.; Yan, H.; Wang, X.; Zhou, J. Selenomethionine alleviates DON-induced oxidative stress via modulating Keap1/Nrf2 signaling in the small intestinal epithelium. J. Agric. Food Chem. 2023, 71, 895–904. [Google Scholar] [CrossRef] [PubMed]
- Xie, L.; Xu, Y.; Ding, X.; Li, K.; Liang, S.; Li, D.; Wang, Y.; Fu, A.; Yu, W.; Zhan, X. Selenomethionine attenuated H2O2-induced oxidative stress and apoptosis by Nrf2 in chicken liver cells. Antioxidants 2023, 12, 1685. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Wang, Y.; Xu, A.; Zhao, B.; Xia, Y.; He, Y.; Xue, H.; Li, S. Dietary selenomethionine reduced oxidative stress by resisting METTL3-mediated m6A methylation level of Nrf2 to ameliorate LPS-induced liver necroptosis in laying hens. J. Nutr. Biochem. 2024, 125, 109563. [Google Scholar] [CrossRef]
- Dong, B.; Jiang, Y.; Shi, B.; Zhang, Z.; Zhang, Z. Selenomethionine alleviates decabromodiphenyl ether-induced oxidative stress and ferroptosis via the NRF2/GPX4 pathway in the chicken brain. J. Hazard. Mater. 2024, 465, 133307. [Google Scholar] [CrossRef]
- Chen, J.; Tian, M.; Guan, W.; Wen, T.; Yang, F.; Chen, F.; Zhang, S.; Song, J.; Ren, C.; Zhang, Y.; et al. Increasing selenium supplementation to a momderately-reduced energy and protein diet improves antioxidant status and meat quality without affecting growth performance in finishing pigs. J. Trace Elem. Med. Biol. 2019, 56, 38–45. [Google Scholar] [CrossRef]
- Bracarense, A.-P.F.L.; Lucioli, J.; Grenier, B.; Drociunas Pacheco, G.; Moll, W.-D.; Schatzmayr, G.; Oswald, I.P. Chronic ingestion of deoxynivalenol and fumonisin, alone or in interaction, induces morphological and immunological changes in the intestine of piglets. Br. J. Nutr. 2012, 107, 1776–1786. [Google Scholar] [CrossRef]
- Chen, J.; Xia, Y.; Hu, Y.; Zhao, X.; You, J.; Zou, T. A blend of formic acid, benzoic acid, and tributyrin alleviates ETEC K88-induced intestinal barrier dysfunction by regulating intestinal inflammation and gut microbiota in a murine model. Int. Immunopharmacol. 2023, 114, 109538. [Google Scholar] [CrossRef]
- Panisson, J.C.; Wellington, M.O.; Bosompem, M.A.; Nagl, V.; Schwartz-Zimmermann, H.E.; Columbus, D.A. Urinary and serum concentration of deoxynivalenol (DON) and DON metabolites as an indicator of DON contamination in swine diets. Toxins 2023, 15, 120. [Google Scholar] [CrossRef]
- Nasim, M.J.; Zuraik, M.M.; Abdin, A.Y.; Ney, Y.; Jacob, C. Selenomethionine: A pink trojan redox horse with implications in aging and various age-related diseases. Antioxidants 2021, 10, 882. [Google Scholar] [CrossRef]
- Awad, W.; Böhm, J.; Razzazi-Fazeli, E.; Zentek, J. Effects of feeding deoxynivalenol contaminated wheat on growth performance, organ weights and histological parameters of the intestine of broiler chickens. J. Anim. Physiol. Anim. Nutr. 2006, 90, 32–37. [Google Scholar] [CrossRef]
- Lucke, A.; Doupovec, B.; Paulsen, P.; Zebeli, Q.; Böhm, J. Effects of low to moderate levels of deoxynivalenol on feed and water intake, weight gain, and slaughtering traits of broiler chickens. Mycotoxin Res. 2017, 33, 261–271. [Google Scholar] [CrossRef] [PubMed]
- Tardivel, C.; Airault, C.; Djelloul, M.; Guillebaud, F.; Barbouche, R.; Troadec, J.-D.; Gaigé, S.; Dallaporta, M. The food born mycotoxin deoxynivalenol induces low-grade inflammation in mice in the absence of observed-adverse effects. Toxicol. Lett. 2015, 232, 601–611. [Google Scholar] [CrossRef]
- Bai, Y.; Ma, K.; Li, J.; Li, J.; Bi, C.; Shan, A. Deoxynivalenol exposure induces liver damage in mice: Inflammation and immune responses, oxidative stress, and protective effects of Lactobacillus rhamnosus GG. Food Chem. Toxicol. 2021, 156, 112514. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Wang, X.; Chen, S.; Wang, S.; Tu, Z.; Zhang, G.; Zhu, H.; Li, X.; Xiong, J.; Liu, Y. Medium-chain triglycerides attenuate liver injury in lipopolysaccharide-challenged pigs by inhibiting necroptotic and inflammatory signaling pathways. Int. J. Mol. Sci. 2018, 19, 3697. [Google Scholar] [CrossRef] [PubMed]
- Atmaca, M.; Bilgin, H.M.; Obay, B.D.; Diken, H.; Kelle, M.; Kale, E. The hepatoprotective effect of coumarin and coumarin derivates on carbon tetrachloride-induced hepatic injury by antioxidative activities in rats. J. Physiol. Biochem. 2011, 67, 569–576. [Google Scholar] [CrossRef] [PubMed]
- Schieber, M.; Chandel, N.S. ROS function in redox signaling and oxidative stress. Curr. Biol. 2014, 24, R453–R462. [Google Scholar] [CrossRef]
- Li, S.; Tan, H.-Y.; Wang, N.; Zhang, Z.-J.; Lao, L.; Wong, C.-W.; Feng, Y. The role of oxidative stress and antioxidants in liver diseases. Int. J. Mol. Sci. 2015, 16, 26087–26124. [Google Scholar] [CrossRef]
- Yu, M.; Peng, Z.; Liao, Y.; Wang, L.; Li, D.; Qin, C.; Hu, J.; Wang, Z.; Cai, M.; Cai, Q.; et al. Deoxynivalenol-induced oxidative stress and Nrf2 translocation in maternal liver on gestation day 12.5 d and 18.5 d. Toxicon 2019, 161, 17–22. [Google Scholar] [CrossRef] [PubMed]
- Flynn, J.M.; Melov, S. SOD2 in mitochondrial dysfunction and neurodegeneration. Free Radic. Biol. Med. 2013, 62, 4–12. [Google Scholar] [CrossRef] [PubMed]
- Li, P.A.; Hou, X.; Hao, S. Mitochondrial biogenesis in neurodegeneration. J. Neurosci. Res. 2017, 95, 2025–2029. [Google Scholar] [CrossRef]
- Cao, S.; Wu, H.; Wang, C.; Zhang, Q.; Jiao, L.; Lin, F.; Hu, C.H. Diquat-induced oxidative stress increases intestinal permeability, impairs mitochondrial function, and triggers mitophagy in piglets. J. Anim. Sci. 2018, 96, 1795–1805. [Google Scholar] [CrossRef] [PubMed]
- Kang, R.; Li, R.; Dai, P.; Li, Z.; Li, Y.; Li, C. Deoxynivalenol induced apoptosis and inflammation of IPEC-J2 cells by promoting ROS production. Environ. Pollut. 2019, 251, 689–698. [Google Scholar] [CrossRef]
- Ji, X.; Zheng, W.; Yao, W. Protective role of hydrogen gas on oxidative damage and apoptosis in intestinal porcine epithelial cells (IPEC-J2) induced by deoxynivalenol: A preliminary study. Toxins 2019, 12, 5. [Google Scholar] [CrossRef]
- Kulsoom, B.; Shamsi, T.S.; Afsar, N.A.; Memon, Z.; Ahmed, N.; Hasnain, S.N. Bax, Bcl-2, and Bax/Bcl-2 as prognostic markers in acute myeloid leukemia: Are we ready for Bcl-2-directed therapy? Cancer Manag. Res. 2018, 10, 403–416. [Google Scholar] [CrossRef]
- Ni, X.; Lu, Y.; Li, M.; Liu, Y.; Zhang, M.; Sun, F.; Dong, S.; Zhao, L. Application of Se-Met to CdTe QDs significantly reduces toxicity by modulating redox balance and inhibiting apoptosis. Ecotoxicol. Environ. Saf. 2023, 267, 115614. [Google Scholar] [CrossRef]
- Chen, J.; Zhang, Y.; Lv, Y.; Tian, M.; You, J.; Chen, F.; Zhang, S.; Guan, W. Effects of selenomethionine on cell viability, selenoprotein expression and antioxidant function in porcine mammary epithelial cells. Front. Nutr. 2021, 8, 665855. [Google Scholar] [CrossRef]
- Li, L.; Chen, Y.; Jiao, D.; Yang, S.; Li, L.; Li, P. Protective effect of astaxanthin on ochratoxin a-induced kidney injury to mice by regulating oxidative stress-related NRF2/KEAP1 pathway. Molecules 2020, 25, 1386. [Google Scholar] [CrossRef] [PubMed]
- Ma, Q. Role of Nrf2 in oxidative stress and toxicity. Annu. Rev. Pharmacol. Toxicol. 2013, 53, 401–426. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Liu, C.; Zhao, Y.; Wang, J.; Li, J.; Zheng, M. Selenium regulates Nrf2 signaling to prevent hepatotoxicity induced by hexavalent chromium in broilers. Poult. Sci. 2023, 102, 102335. [Google Scholar] [CrossRef] [PubMed]
- Bao, S.; Lin, J.; Xie, M.; Wang, C.; Nie, X. Simvastatin affects Nrf2/MAPK signaling pathway and hepatic histological structure change in Gambusia Affinis. Chemosphere 2021, 269, 128725. [Google Scholar] [CrossRef]
- Li, E.; Horn, N.; Ajuwon, K.M. Mechanisms of deoxynivalenol-induced endocytosis and degradation of tight junction proteins in jejunal IPEC-J2 cells involve selective activation of the MAPK pathways. Arch. Toxicol. 2021, 95, 2065–2079. [Google Scholar] [CrossRef]
- Pestka, J.J.; Amuzie, C.J. Tissue distribution and proinflammatory cytokine gene expression following acute oral exposure to deoxynivalenol: Comparison of weanling and adult mice. Food Chem. Toxicol. 2008, 46, 2826–2831. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Item | Ingredient, % | Item | Nutrient Levels c |
|---|---|---|---|
| Corn | 36.50 | Digestible energy, kcal/kg | 3467.00 |
| Extruded corn | 20.00 | Crude protein, % | 19.07 |
| Extruded soybean | 10.00 | Lysine, % | 1.39 |
| Soybean meal | 6.50 | Methionine, % | 0.48 |
| Fish meal | 5.00 | Threonine, % | 0.79 |
| Whey powder | 5.00 | Calcium, % | 0.68 |
| Soy protein concentrate | 4.00 | Total phosphorus, % | 0.55 |
| Wheat flour | 5.00 | Available phosphorus, % | 0.33 |
| Wheat bran | 4.00 | ||
| Limestone | 0.70 | ||
| Dicalcium phosphate | 0.50 | ||
| Salt | 0.30 | ||
| Chromium trioxide | 0.25 | ||
| Choline chloride (60%) | 0.10 | ||
| L-Lysine hydrochloride | 0.50 | ||
| DL-Methionine | 0.16 | ||
| L-Threonine | 0.10 | ||
| L-Tryptophan | 0.03 | ||
| L-Valine | 0.14 | ||
| Vitamin premix a | 0.16 | ||
| Mineral premix b | 0.10 | ||
| Zeolite powder | 0.96 | ||
| Total | 100.00 |
| Genes | Accession Number | Primer Sequence (5′-3′) |
|---|---|---|
| NQO1 | NM_001159613.1 | F: GCCCAGATATTGTGGCCGAA R: AACTCCCCTATGAGCACACG |
| SOD1 | NM_001190422.1 | F: AAGGCCGTGTGTGTGCTGAA R: GATCACCTTCAGCCAGTCCTTT |
| SOD2 | NM_214127.2 | F: GGCCTACGTGAACAACCTGA R: TGATTGATGTGGCCTCCACC |
| GPX1 | NM_214201.1 | F: TGAATGGCGCAAATGCTCAC R: ATTGCGACACACTGGAGACC |
| Keap1 | NM_001114671.1 | F: GCCTCATCGAGTTCGCTTAC R: ACGGACCACACTGTCAATCT |
| Nrf2 | XM_021075133.1 | F: CCCATTCACAAAAGACAAACATTC R: GCTTTTGCCCTTAGCTCATCTC |
| HO1 | NM_001004027.1 | F: TCAAGCAGAGGGTCCTCGAA R: CCTCTTGCGGATGTCGGATG |
| Gclm | XM_001926378.4 | F: GATGCCGCCCGATTTAACTG R: ACAATGACCGAGTACCGCAG |
| Gclc | XM_021098556.1 | F: GACGACGCCAATGAGTCTGA R: AGCACCACAAACACCACGTA |
| PGC-1α | NM_213963.2 | F: GATGACCCTCCTCACACCAA R: TTGGAGGTGCACTTGTCTCT |
| NRF-1 | XM_021078993.1 | F: GCTGTGGCAACAGGAAAGAA R: AAGACAGGGTTGGGTTTGGA |
| TFAM | NM_001130211.1 | F: GTCTGAAGAGTTGCTTGCGA R: ACCCGTAGACAAAGCACTGA |
| GCK | XM_013985832.2 | F: ACCTTCTCCTTTCCCGTGAG R: CCCACGATGTTGTTCCCTTC |
| CS | XM_021091147.1 | F: TCAGGAAGTGCTTGTTTGGC R: GGCAGGTGTTTCAGAGCAAA |
| ND4 | NM_001097468.2 | F: TTATTGGTGCCGGAGGTACTG R: CCCAGTTTATTCCAGGGTTCTG |
| CcOX I | AJ950517.1 | F: ATTATCCTGACGCATACACAGCA R: GCAGATACTTCTCGTTTTGATGC |
| CcOX IV | AK233334.1 | F: CCAAGTGGGACTACGACAAGAAC R: CCTGCTCGTTTATTAGCACTGG |
| CcOX V | NM_001007517.1 | F: ATCTGGAGGTGGTGTTCCTACTG R: GTTGGTGATGGAGGGGACTAAA |
| ATPS | AK230503 | F: TGTCCTCCTCCCTATCACACATT R: TAGTGGTTATGACGTTGGCTTGA |
| Bcl2 | XM_005666256.3 | F: CTTCTGCAAATCCCGGACTG R: CCGTAGGAATCCCAACCAGA |
| Bax | XM_013998624.2 | F: TCGCTCACCATCTGGAAGAA R: ATTGTCCTCCGAGACCACTC |
| CYCS | NM_001129970.1 | F: ACCTCCATGGTCTCTTTGGG R: TGCCTTTGTTCTTGTTGGCA |
| CASP9 | XM_003127618.4 | F: TGGAACTCAAGCCAGAGGAG R: CTGCATTCAGGACGTAAGCC |
| CASP3 | NM_214131.1 | F: CGGACAGTGGGACTGAAGAT R: GCTGCACAAAGTGACTGGAT |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhong, H.; Huang, Z.; Li, L.; Chen, X.; Zou, T.; Chen, J.; You, J. Selenomethionine Supplementation Mitigates Liver Dysfunction, Oxidative Injury and Apoptosis through Enhancing Antioxidant Capacity and Inhibiting JNK MAPK Pathway in Piglets Fed Deoxynivalenol-Contaminated Diets. Antioxidants 2024, 13, 295. https://doi.org/10.3390/antiox13030295
Zhong H, Huang Z, Li L, Chen X, Zou T, Chen J, You J. Selenomethionine Supplementation Mitigates Liver Dysfunction, Oxidative Injury and Apoptosis through Enhancing Antioxidant Capacity and Inhibiting JNK MAPK Pathway in Piglets Fed Deoxynivalenol-Contaminated Diets. Antioxidants. 2024; 13(3):295. https://doi.org/10.3390/antiox13030295
Chicago/Turabian StyleZhong, Haopeng, Zhouyin Huang, Lin Li, Xingping Chen, Tiande Zou, Jun Chen, and Jinming You. 2024. "Selenomethionine Supplementation Mitigates Liver Dysfunction, Oxidative Injury and Apoptosis through Enhancing Antioxidant Capacity and Inhibiting JNK MAPK Pathway in Piglets Fed Deoxynivalenol-Contaminated Diets" Antioxidants 13, no. 3: 295. https://doi.org/10.3390/antiox13030295
APA StyleZhong, H., Huang, Z., Li, L., Chen, X., Zou, T., Chen, J., & You, J. (2024). Selenomethionine Supplementation Mitigates Liver Dysfunction, Oxidative Injury and Apoptosis through Enhancing Antioxidant Capacity and Inhibiting JNK MAPK Pathway in Piglets Fed Deoxynivalenol-Contaminated Diets. Antioxidants, 13(3), 295. https://doi.org/10.3390/antiox13030295