

Acute Antioxidant Response to Two Types of Exercises: 2000 M Run vs. Burpee Test

 ,
,  , ,
, ,  , ,
, ,  ,
,  and
and 
Abstract
1. Introduction
2. Materials and Methods
2.1. Participants and Intervention Protocol
2.2. Blood Cell Purification
2.3. Determination of Antioxidant Enzymatic Activities and Oxidative Stress Markers
2.4. Statistical Analysis
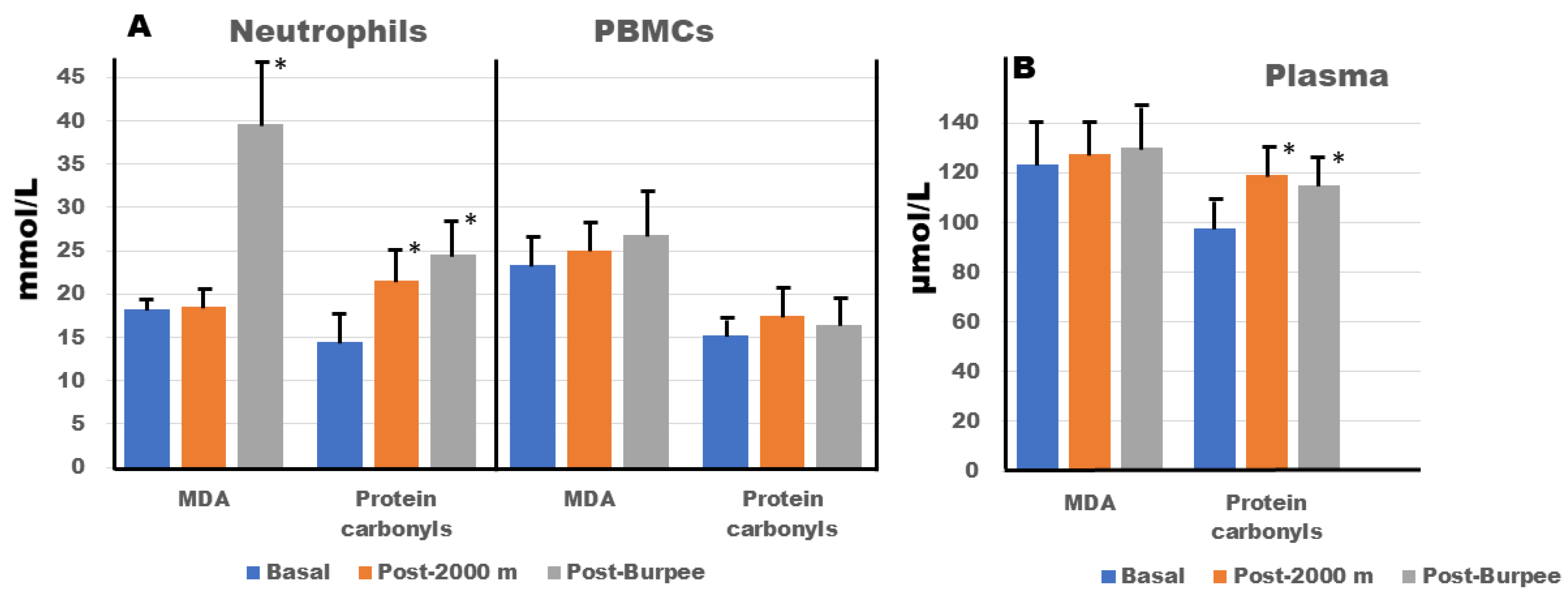
3. Results
4. Discussion
4.1. Presence of Oxidative Stress Post-exercise
4.2. Post-Exercise Antioxidant Response
4.3. Novelty and Limitations of the Study
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Powers, S.K.; Radak, Z.; Ji, L.L. Exercise-induced oxidative stress: Past, present and future. J. Physiol. 2016, 594, 5081–5092. [Google Scholar] [CrossRef] [PubMed]
- Thirupathi, A.; Pinho, R.A. Effects of reactive oxygen species and interplay of antioxidants during physical exercise in skeletal muscles. J. Physiol. Biochem. 2018, 74, 359–367. [Google Scholar] [CrossRef] [PubMed]
- Gao, L.; Wang, H.J.; Tian, C.; Zucker, I.H. Skeletal Muscle Nrf2 Contributes to Exercise-Evoked Systemic Antioxidant Defense Via Extracellular Vesicular Communication. Exerc. Sport Sci. Rev. 2021, 49, 213–222. [Google Scholar] [CrossRef] [PubMed]
- Matés, J.M.; Pérez-Gómez, C.; Núñez de Castro, I. Antioxidant enzymes and human diseases. Clin. Biochem. 1999, 32, 595–603. [Google Scholar] [CrossRef] [PubMed]
- He, F.; Li, J.; Liu, Z.; Chuang, C.C.; Yang, W.; Zuo, L. Redox Mechanism of Reactive Oxygen Species in Exercise. Front. Physiol. 2016, 7, 486. [Google Scholar] [CrossRef] [PubMed]
- Di Meo, S.; Napolitano, G.; Venditti, P. Mediators of Physical Activity Protection against ROS-Linked Skeletal Muscle Damage. Int. J. Mol. Sci. 2019, 20, 3024. [Google Scholar] [CrossRef] [PubMed]
- Ji, L.L.; Gomez-Cabrera, M.C.; Vina, J. Exercise and hormesis: Activation of cellular antioxidant signaling pathway. Ann. N. Y. Acad. Sci. 2006, 1067, 425–435. [Google Scholar] [CrossRef]
- Ji, L.L.; Kang, C.; Zhang, Y. Exercise-induced hormesis and skeletal muscle health. Free Radic. Biol. Med. 2016, 98, 113–122. [Google Scholar] [CrossRef]
- Powers, S.K.; Deminice, R.; Ozdemir, M.; Yoshihara, T.; Bomkamp, M.P.; Hyatt, H. Exercise-induced oxidative stress: Friend or foe? J. Sport Health Sci. 2020, 9, 415–425. [Google Scholar] [CrossRef]
- Marzatico, F.; Pansarasa, O.; Bertorelli, L.; Somenzini, L.; Della Valle, G. Blood free radical antioxidant enzymes and lipid peroxides following long-distance and lactacidemic performances in highly trained aerobic and sprint athletes. J. Sports Med. Phys. Fit. 1997, 37, 235–239. [Google Scholar]
- Margaritelis, N.V.; Paschalis, V.; Theodorou, A.A.; Kyparos, A.; Nikolaidis, M.G. Antioxidants in Personalized Nutrition and Exercise. Adv. Nutr. 2018, 9, 813–823. [Google Scholar] [CrossRef] [PubMed]
- Carrera-Quintanar, L.; Funes, L.; Herranz-López, M.; Martínez-Peinado, P.; Pascual-García, S.; Sempere, J.M.; Boix-Castejón, M.; Córdova, A.; Pons, A.; Micol, V.; et al. Antioxidant supplementation modulates neutrophil inflammatory response to exercise-induced stress. Antioxidants 2020, 9, 1242. [Google Scholar] [CrossRef] [PubMed]
- Thirupathi, A.; Wang, M.; Lin, J.K.; Fekete, G.; István, B.; Baker, J.S.; Gu, Y. Effect of Different Exercise Modalities on Oxidative Stress: A Systematic Review. Biomed. Res. Int. 2021, 2021, 1947928. [Google Scholar] [CrossRef] [PubMed]
- Shi, M.; Wang, X.; Yamanaka, T.; Ogita, F.; Nakatani, K.; Takeuchi, T. Effects of anaerobic exercise and aerobic exercise on biomarkers of oxidative stress. Environ. Health Prev. Med. 2007, 12, 202–208. [Google Scholar] [CrossRef] [PubMed]
- Ammar, A.; Trabelsi, K.; Boukhris, O.; Glenn, J.M.; Bott, N.; Masmoudi, L.; Hakim, A.; Chtourou, H.; Driss, T.; Hoekelmann, A.; et al. Effects of Aerobic-, Anaerobic- and Combined-Based Exercises on Plasma Oxidative Stress Biomarkers in Healthy Untrained Young Adults. Int. J. Environ. Res. Public Health 2020, 17, 2601. [Google Scholar] [CrossRef] [PubMed]
- Sureda, A.; Tauler, P.; Aguiló, A.; Cases, N.; Fuentespina, E.; Córdova, A.; Tur, J.A.; Pons, A. Relation between oxidative stress markers and antioxidant endogenous defences during exhaustive exercise. Free Radic. Res. 2005, 39, 1317–1324. [Google Scholar] [CrossRef] [PubMed]
- Radak, Z.; Taylor, A.W. Exercise and hormesis. In The Science of Hormesis in Health and Longevity; Rattan, S., Kyriazi, M., Eds.; Elsevier: Oxford, UK, 2018; pp. 63–73. [Google Scholar]
- Bloomer, R.J.; Goldfarb, A.H.; Wideman, L.; McKenzie, M.J.; Consitt, L.A. Effects of acute aerobic and anaerobic exercise on blood markers of oxidative stress. J. Strength Cond. Res. 2005, 19, 276–285. [Google Scholar] [CrossRef]
- Bloomer, R.J.; Fry, A.C.; Falvo, M.J.; Moore, C.A. Protein carbonyls are acutely elevated following single set anaerobic exercise in resistance trained men. J. Sci. Med. Sport 2007, 10, 411–417. [Google Scholar] [CrossRef]
- Ismaeel, A.; Holmes, M.; Papoutsi, E.; Panton, L.; Koutakis, P. Resistance Training, Antioxidant Status, and Antioxidant Supplementation. Int. J. Sport Nutr. Exerc. Metab. 2019, 29, 539–547. [Google Scholar] [CrossRef]
- Bogdanis, G.C.; Stavrinou, P.; Fatouros, I.G.; Philippou, A.; Chatzinikolaou, A.; Draganidis, D.; Ermidis, G.; Maridaki, M. Short-term high-intensity interval exercise training attenuates oxidative stress responses and improves antioxidant status in healthy humans. Food Chem. Toxicol. 2013, 61, 171–177. [Google Scholar] [CrossRef]
- Viña, J.; Gomez-Cabrera, M.C.; Lloret, A.; Marquez, R.; Miñana, J.B.; Pallardó, F.V.; Sastre, J. Free radicals in exhaustive physical exercise: Mechanism of production, and protection by antioxidants. IUBMB Life 2000, 50, 271–277. [Google Scholar] [CrossRef] [PubMed]
- Lu, Y.; Wiltshire, H.D.; Baker, J.S.; Wang, Q. Effects of High Intensity Exercise on Oxidative Stress and Antioxidant Status in Untrained Humans: A Systematic Review. Biology 2021, 10, 1272. [Google Scholar] [CrossRef] [PubMed]
- Schneider, C.D.; Barp, J.; Ribeiro, J.L.; Belló-Klein, A.; Oliveira, A.R. Oxidative stress after three different intensities of running. Can. J. Appl. Physiol. 2005, 30, 723–734. [Google Scholar] [CrossRef] [PubMed]
- Bloomer, R.J.; Goldfarb, A.H. Anaerobic exercise and oxidative stress: A review. Can. J. Appl. Physiol. 2004, 29, 245–263. [Google Scholar] [CrossRef] [PubMed]
- Córdova-Martínez, A.; Caballero-García, A.; Bello, H.J.; Perez-Valdecantos, D.; Roche, E. Effects of Eccentric vs. Concentric Sports on Blood Muscular Damage Markers in Male Professional Players. Biology 2022, 11, 343. [Google Scholar] [CrossRef] [PubMed]
- González-Bartholin, R.; Mackay, K.; Valladares, D.; Zbinden-Foncea, H.; Nosaka, K.; Peñailillo, L. Changes in oxidative stress, inflammation and muscle damage markers following eccentric versus concentric cycling in older adults. Eur. J. Appl. Physiol. 2019, 119, 2301–2312. [Google Scholar] [CrossRef]
- Marfell-Jones, M.; Olds, T.; Steward, A.; Carter, L. International Standards for Anthropometric Assessment; International Society for the Advancement of Kinanthropometry: Potchefstroom, South Africa, 2006. [Google Scholar]
- Young, D.S.; Friedman, R.B. Effects of Disease on Clinical Laboratory Tests; AACC Press: Washington, DC, USA, 2001. [Google Scholar]
- Ainsworth, B.E.; Haskell, W.L.; Whitt, M.C.; Irwin, M.L.; Swartz, A.M.; Strath, S.J.; O’Brien, W.L.; Bassett, D.R., Jr.; Schmitz, K.H.; Emplaincourt, P.O.; et al. Compendium of physical activities: An update of activity codes and MET intensities. Med. Sci. Sports Exerc. 2000, 32, S498–S504. [Google Scholar] [CrossRef]
- Funes, L.; Carrera-Quintanar, L.; Cerdán-Calero, M.; Ferrer, M.D.; Drobnic, F.; Pons, A.; Roche, E.; Micol, V. Effect of lemon verbena supplementation on muscular damage markers, proinflammatory cytokines release and neutrophils’ oxidative stress in chronic exercise. Eur. J. Appl. Physiol. 2011, 111, 695–705. [Google Scholar] [CrossRef]
- Carrera-Quintanar, L.; Funes, L.; Sánchez-Martos, M.; Martinez-Peinado, P.; Sempere, J.M.; Pons, A.; Micol, V.; Roche, E. Effect of a 2000-m running test on antioxidant and cytokine response in plasma and circulating cells. J. Physiol. Biochem. 2017, 73, 523–530. [Google Scholar] [CrossRef]
- Zhao, H.; Zhang, R.; Yan, X.; Fan, K. Superoxide dismutase nanozymes: An emerging star for anti-oxidation. J. Mater. Chem. B 2021, 9, 6939–6957. [Google Scholar] [CrossRef]
- Lubos, E.; Loscalzo, J.; Handy, D.E. Glutathione peroxidase-1 in health and disease: From molecular mechanisms to therapeutic opportunities. Antioxid. Redox Signal. 2011, 15, 1957–1997. [Google Scholar] [CrossRef] [PubMed]
- Sepasi Tehrani, H.; Moosavi-Movahedi, A.A. Catalase and its mysteries. Prog. Biophys. Mol. Biol. 2018, 140, 5–12. [Google Scholar] [CrossRef] [PubMed]
- Couto, N.; Wood, J.; Barber, J. The role of glutathione reductase and related enzymes on cellular redox homoeostasis network. Free Radic. Biol. Med. 2016, 95, 27–42. [Google Scholar] [CrossRef] [PubMed]
- Owens, D.J.; Twist, C.; Cobley, J.N.; Howatson, G.; Close, G.L. Exercise-induced muscle damage: What is it, what causes it and what are the nutritional solutions? Eur. J. Sport Sci. 2019, 19, 71–85. [Google Scholar] [CrossRef] [PubMed]
- Mrakic-Sposta, S.; Gussoni, M.; Montorsi, M.; Porcelli, S.; Vezzoli, A. A quantitative method to monitor reactive oxygen species production by electron paramagnetic resonance in physiological and pathological conditions. Oxid. Med. Cell Longev. 2014, 2014, 306179. [Google Scholar] [CrossRef]
- Madian, A.G.; Regnier, F.E. Proteomic identification of carbonylated proteins and their oxidation sites. J. Proteome Res. 2010, 9, 3766–3780. [Google Scholar] [CrossRef] [PubMed]
- Gaschler, M.M.; Stockwell, B.R. Lipid peroxidation in cell death. Biochem. Biophys. Res. Commun. 2017, 482, 419–425. [Google Scholar] [CrossRef]
- Carrera-Quintanar, L.; Funes, L.; Viudes, E.; Tur, J.; Micol, V.; Roche, E.; Pons, A. Antioxidant effect of lemon verbena extracts in lymphocytes of university students performing aerobic training program. Scand. J. Med. Sci. Sports 2012, 22, 454–461. [Google Scholar] [CrossRef]
- Sies, H. Hydrogen peroxide as a central redox signaling molecule in physiological oxidative stress: Oxidative eustress. Redox Biol. 2017, 11, 613–619. [Google Scholar] [CrossRef]
- Spolarics, Z. Endotoxemia, pentose cycle, and the oxidant/antioxidant balance in the hepatic sinusoid. J. Leukoc. Biol. 1998, 63, 534–541. [Google Scholar] [CrossRef]
- Bentley, D.J.; Ackerman, J.; Clifford, T.; Slattery, K.S. Acute and Chronic Effects of Antioxidant Supplementation on Exercise Performance. In Antioxidants in Sport Nutrition; Lamprecht, M., Ed.; CRC Press/Taylor & Francis: Boca Raton, FL, USA, 2015; Chapter 9. [Google Scholar]
- Korbecki, J.; Bobiński, R.; Dutka, M. Self-regulation of the inflammatory response by peroxisome proliferator-activated receptors. Inflamm. Res. 2019, 68, 443–458. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
| Parameter (Units) | Reference Values in Adult Males | Day 1 |
|---|---|---|
| Hemogram | ||
| Erythrocytes (106 cells/μL) | 4.4–5.9 | 5.3 ± 0.2 |
| Hemoglobin (g/g serum protein) | 20–26 | 25.4 ± 0.8 |
| Platelets (103 cells/μL) | 150–450 | 228.1 ± 37.8 |
| Neutrophils (103 cells/μL) | 1.8–7.7 | 3.3 ± 0.3 |
| Lymphocytes (103 cells/μL) | 1.0–4.8 | 2.2 ± 0.4 |
| Circulating metabolites | ||
| Glucose (mg/dL) | <100 | 81.7 ± 5.2 |
| Triglycerides (mg/dL) | <160 | 78.3 ± 8.6 |
| Total cholesterol (mg/dL) | <200 | 151.6 ± 6.1 |
| LDL (mg/dL) | <130 | 96 ± 22.2 |
| HDL (mg/dL) | >55 | 77 ± 13.6 |
| Creatinine (mg/dL) | 0.8–1.4 | 0.9 ± 0.1 |
| Uric acid (mg/dL) | 2.5–8.0 | 4.5 ± 0.7 |
| Urea (mg/dL) | 23–44 | 35.3 ± 2.1 |
| Circulating tissue markers | ||
| ALT (U/L) | <40 | 20.9 ± 1.7 |
| AST (U/L) | <40 | 22.8 ± 2.0 |
| GGT (U/L) | <55 | 19.3 ± 1.8 |
| CK (U/L) | 39–308 | 162.5 ± 32.5 |
| Parameter (Units) | Basal | Post-2000 m Test | Post-Burpee Test | d |
|---|---|---|---|---|
| Circulating lactate (mg/dL) | 8.3 ± 1.9 | 26.1 ± 2.3 * | 23.6 ± 3.6 † | 0.8 |
| Circulating CK (U/L) | 162.5 ± 32.5 | 181.2 ± 19.7 * | 208.9 ± 22.6 † | 1.3 |
| Heart rate (bpm) | 58.2 ± 8.7 | 168.4 ± 10.2 * | 171.7 ± 17.4 † | 0.2 |
| Enzymatic Activity (Units) | Basal | Post-2000 m Run | Post-Burpee Test | d |
|---|---|---|---|---|
| Neutrophils | ||||
| SOD (pkat/109 cells) | 22.9 ± 2.7 | 32.8 ± 4.6 * | 28.3 ± 5.2 † | 0.9 |
| CAT (k/109 cells) | 57.9 ± 5.1 | 67.5 ± 9.8 | 106.3 ± 14.3 † | 3.1 |
| GPX (nkat/109 cells) | 121.1 ± 18.8 | 174.7 ± 21.2 * | 147.5 ± 9.3 † | 1.7 |
| GRD (nkat/109 cells) | 546.2 ± 41.3 | 575.4 ± 51.3 | 572.8 ± 26.2 | 0.1 |
| MPO (µkat/109 cells) | 32.2 ± 6.2 | 33.3 ± 7.5 | 36.8 ± 4.9 | 0.6 |
| PBMCs | ||||
| SOD (pkat/109 cells) | 51.4 ± 3.5 | 60.8 ± 4.2 | 56.1 ± 2.9 | 1.3 |
| CAT (k/109 cells) | 89.7 ± 13.6 | 127.3 ± 13.6 * | 113.8 ± 9.0 † | 1.2 |
| GPX (nkat/109 cells) | 85.5 ± 9.5 | 97.5 ± 11.4 | 108.9 ± 24.0 | 0.6 |
| GRD (nkat/109 cells) | 451.1 ± 48.6 | 463.4 ± 26.8 | 477.6 ± 25.5 | 0.5 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Carrera-Quintanar, L.; Funes, L.; Herranz-López, M.; Vicente-Salar, N.; Mielgo-Ayuso, J.; Moya-Ramón, M.; Pons, A.; Micol, V.; Roche, E. Acute Antioxidant Response to Two Types of Exercises: 2000 M Run vs. Burpee Test. Antioxidants 2024, 13, 144. https://doi.org/10.3390/antiox13020144
Carrera-Quintanar L, Funes L, Herranz-López M, Vicente-Salar N, Mielgo-Ayuso J, Moya-Ramón M, Pons A, Micol V, Roche E. Acute Antioxidant Response to Two Types of Exercises: 2000 M Run vs. Burpee Test. Antioxidants. 2024; 13(2):144. https://doi.org/10.3390/antiox13020144
Chicago/Turabian StyleCarrera-Quintanar, Lucrecia, Lorena Funes, María Herranz-López, Néstor Vicente-Salar, Juan Mielgo-Ayuso, Manuel Moya-Ramón, Antoni Pons, Vicente Micol, and Enrique Roche. 2024. "Acute Antioxidant Response to Two Types of Exercises: 2000 M Run vs. Burpee Test" Antioxidants 13, no. 2: 144. https://doi.org/10.3390/antiox13020144
APA StyleCarrera-Quintanar, L., Funes, L., Herranz-López, M., Vicente-Salar, N., Mielgo-Ayuso, J., Moya-Ramón, M., Pons, A., Micol, V., & Roche, E. (2024). Acute Antioxidant Response to Two Types of Exercises: 2000 M Run vs. Burpee Test. Antioxidants, 13(2), 144. https://doi.org/10.3390/antiox13020144