

Sappanone A Alleviates the Severity of Carbon Tetrachloride-Induced Liver Fibrosis in Mice

Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material
2.2. Animals
2.3. Biochemical Assays
2.4. Staining of Liver Sections
2.5. Quantitative Real-Time Polymerase Chain Reaction (qRT-PCR)
2.6. Enzyme-Linked Immunosorbent Assay (ELISA)
2.7. Cell Culture and Treatments
2.8. Lactate Dehydrogenase (LDH) Assay
2.9. Cell Viability Assay
2.10. Statistical Analysis
3. Results
3.1. Treatment with SA Alleviates Chronic Liver Injury in Mice
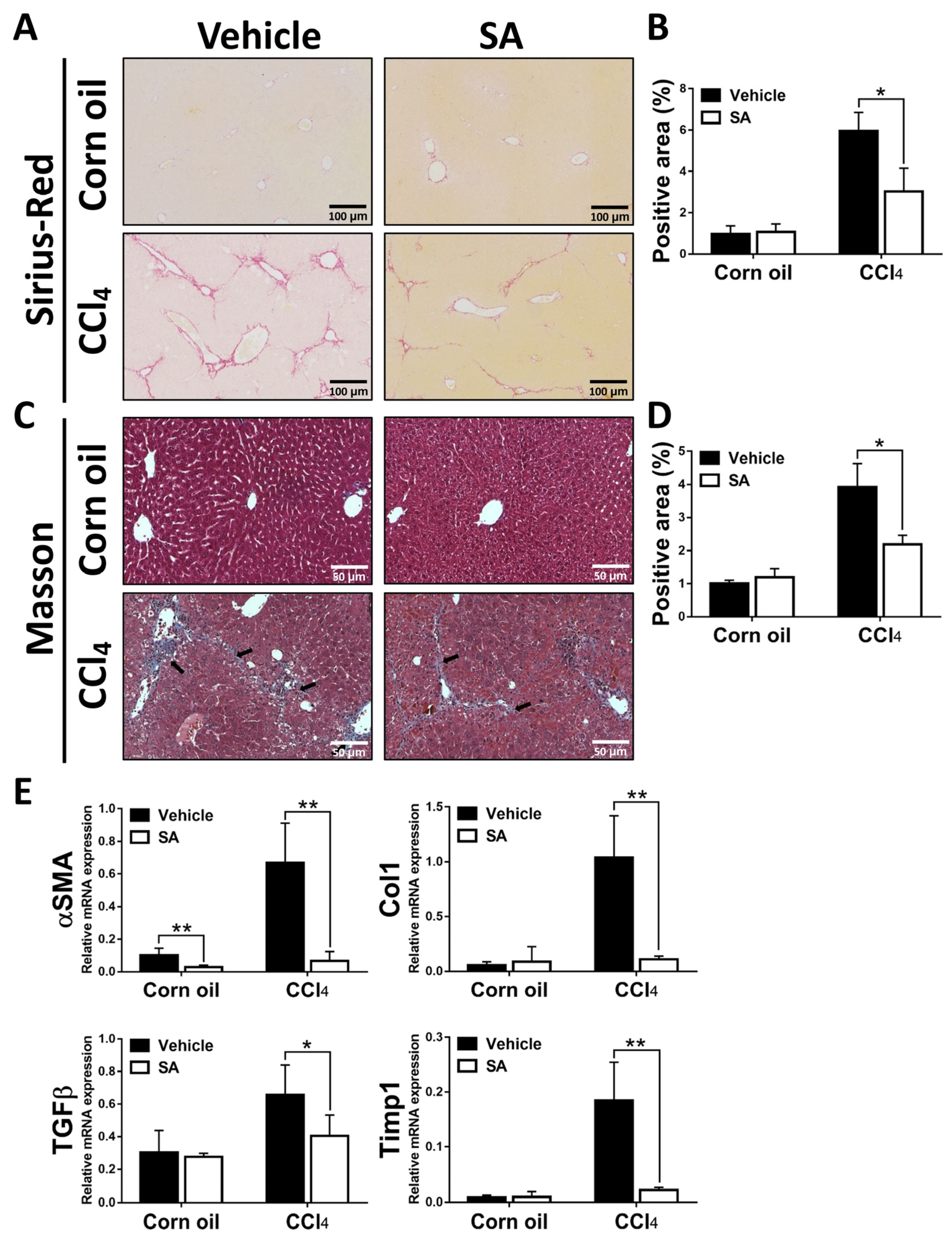
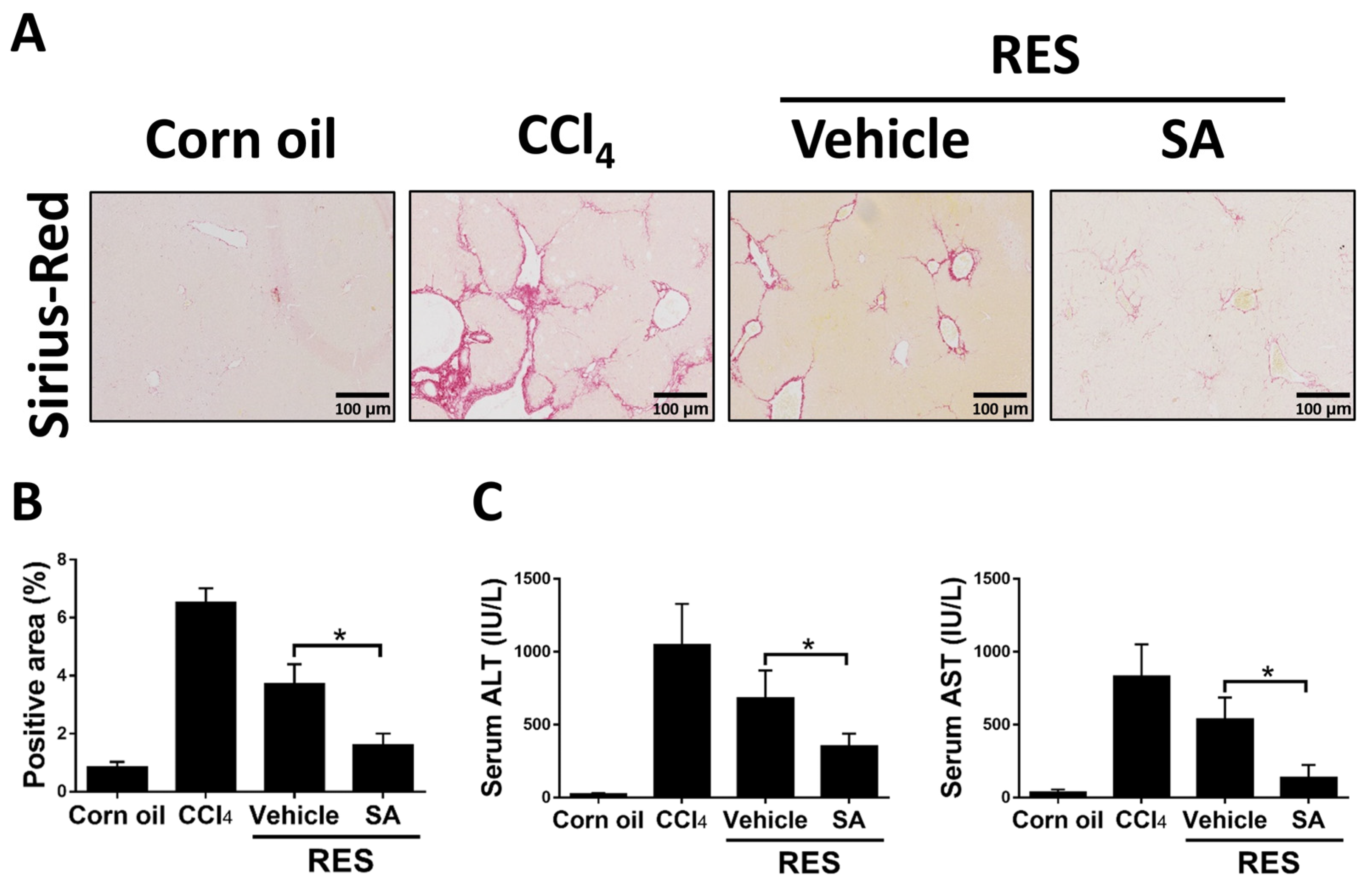
3.2. SA Treatment Ameliorates CCl4-Induced Liver Fibrosis in Mice
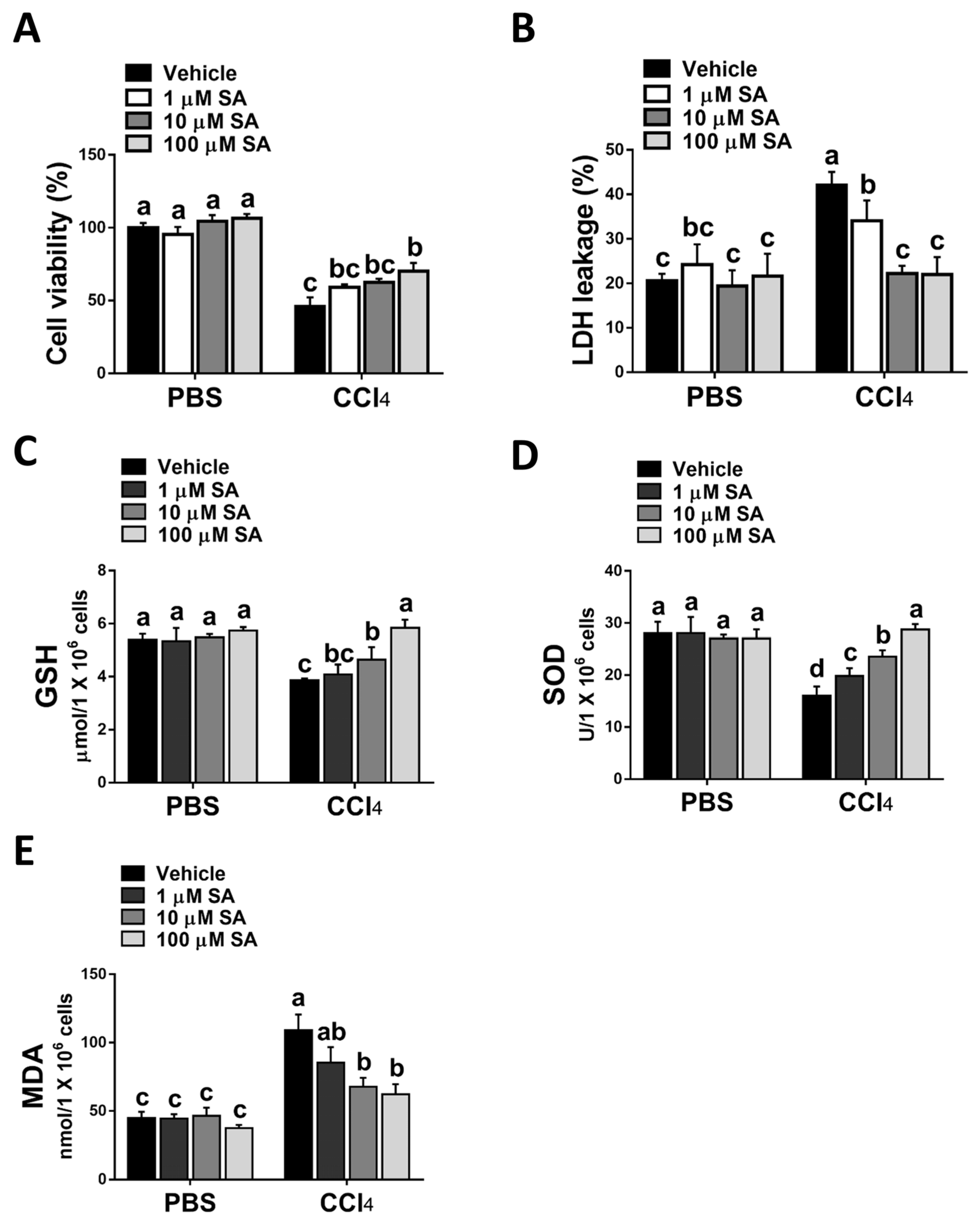
3.3. SA Protects Hepatocytes from Oxidative Stress Damage
3.4. SA Could Not Directly Affect HSC Activation
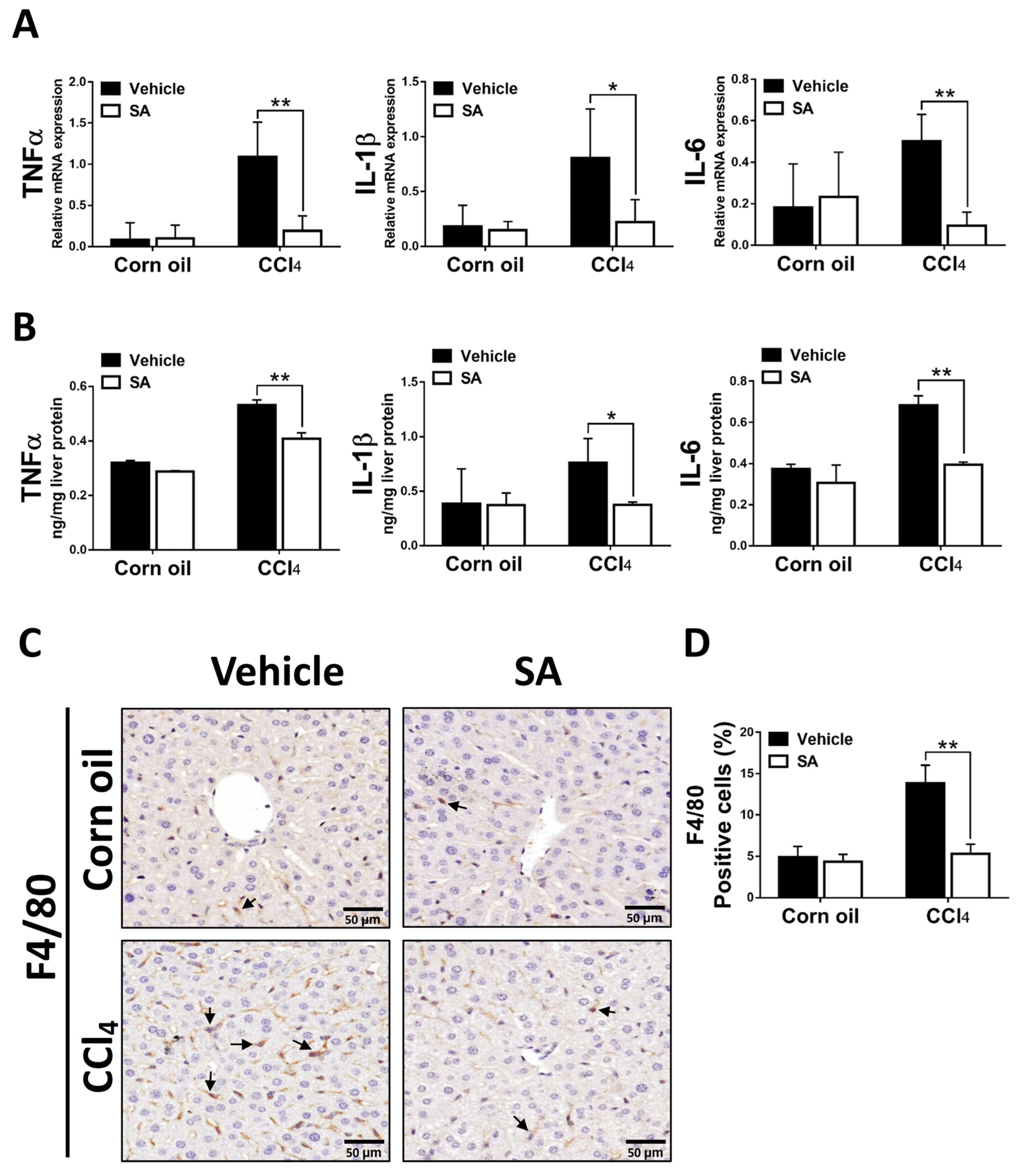
3.5. SA Decreases Hepatic Inflammatory Responses in Fibrotic Livers
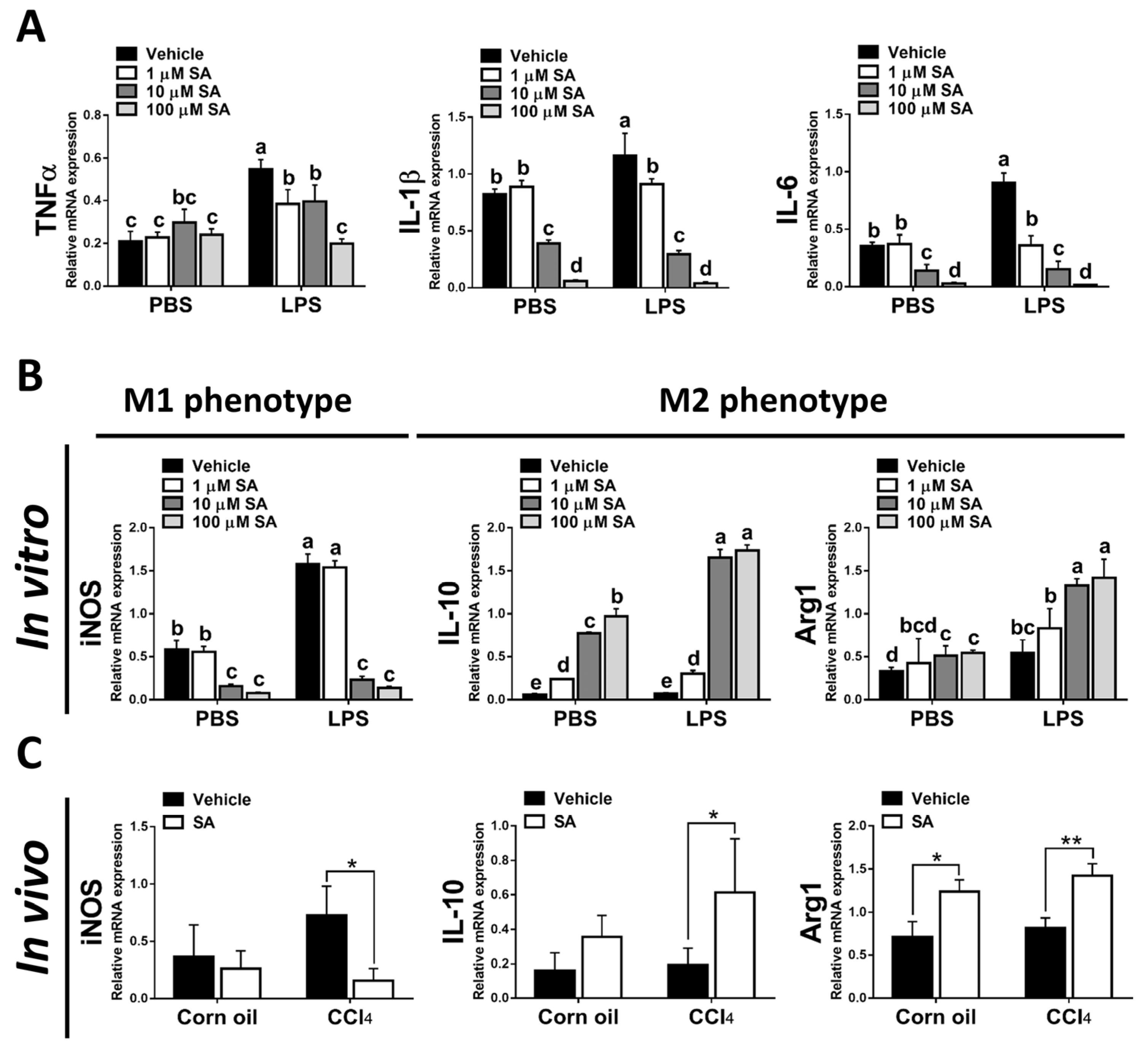
3.6. SA Could Inhibit Macrophage Activation and Promote Polarization of Macrophages towards the M2 Phenotype by Modulating PPARγ
3.7. Treatment with SA Promotes Fibrosis Regression
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Kisseleva, T.; Brenner, D. Molecular and cellular mechanisms of liver fibrosis and its regression. Nature reviews. Gastroenterol. Hepatol. 2021, 18, 151–166. [Google Scholar] [CrossRef]
- Bosch, F.X.; Ribes, J.; Borràs, J. Epidemiology of primary liver cancer. Semin. Liver Dis. 1999, 19, 271–285. [Google Scholar] [CrossRef]
- Boyer-Diaz, Z.; Aristu-Zabalza, P.; Andrés-Rozas, M.; Robert, C.; Ortega-Ribera, M.; Fernández-Iglesias, A.; Broqua, P.; Junien, J.L.; Wettstein, G.; Bosch, J.; et al. Pan-PPAR agonist lanifibranor improves portal hypertension and hepatic fibrosis in experimental advanced chronic liver disease. J. Hepatol. 2021, 74, 1188–1199. [Google Scholar] [CrossRef]
- Shvarts, Y.S.; Zubakhin, A.A.; Dushkin, M.I. Suppression of hemopoiesis during CCl4-induced hepatic fibrosis: Role of systemic endotoxemia. Bull. Exp. Biol. Med. 2000, 130, 759–762. [Google Scholar] [CrossRef]
- Sirica, A.E.; Gores, G.J.; Groopman, J.D.; Selaru, F.M.; Strazzabosco, M.; Wang, X.W.; Zhu, A.X. Intrahepatic Cholangiocarcinoma: Continuing Challenges and Translational Advances. Hepatology 2019, 69, 1803–1815. [Google Scholar] [CrossRef]
- Wagner, K.D.; Wagner, N. Peroxisome proliferator-activated receptor beta/delta (PPARbeta/delta) acts as regulator of metabolism linked to multiple cellular functions. Pharmacol. Ther. 2010, 125, 423–435. [Google Scholar] [CrossRef]
- Pascual, G.; Fong, A.L.; Ogawa, S.; Gamliel, A.; Li, A.C.; Perissi, V.; Rose, D.W.; Willson, T.M.; Rosenfeld, M.G.; Glass, C.K. A SUMOylation-dependent pathway mediates transrepression of inflammatory response genes by PPAR-gamma. Nature 2005, 437, 759–763. [Google Scholar] [CrossRef]
- Choi, S.; Jung, J.E.; Yang, Y.R.; Kim, E.S.; Jang, H.J.; Kim, E.K.; Kim, I.S.; Lee, J.Y.; Kim, J.K.; Seo, J.K.; et al. Novel phosphorylation of PPARγ ameliorates obesity-induced adipose tissue inflammation and improves insulin sensitivity. Cell. Signal. 2015, 27, 2488–2495. [Google Scholar] [CrossRef]
- Kim, J.W.; Zhou, Z.; Yun, H.; Park, S.; Choi, S.J.; Lee, S.H.; Lim, C.W.; Lee, K.; Kim, B. Cigarette smoking differentially regulates inflammatory responses in a mouse model of nonalcoholic steatohepatitis depending on exposure time point. Food Chem. Toxicol. 2020, 135, 110930. [Google Scholar] [CrossRef]
- Li, M.; Pascual, G.; Glass, C.K. Peroxisome proliferator-activated receptor gamma-dependent repression of the inducible nitric oxide synthase gene. Mol. Cell. Biol. 2000, 20, 4699–4707. [Google Scholar] [CrossRef]
- Odegaard, J.I.; Ricardo-Gonzalez, R.R.; Goforth, M.H.; Morel, C.R.; Subramanian, V.; Mukundan, L.; Red Eagle, A.; Vats, D.; Brombacher, F.; Ferrante, A.W.; et al. Macrophage-specific PPARgamma controls alternative activation and improves insulin resistance. Nature 2007, 447, 1116–1120. [Google Scholar] [CrossRef]
- Norris, A.W.; Chen, L.; Fisher, S.J.; Szanto, I.; Ristow, M.; Jozsi, A.C.; Hirshman, M.F.; Rosen, E.D.; Goodyear, L.J.; Gonzalez, F.J.; et al. Muscle-specific PPARgamma-deficient mice develop increased adiposity and insulin resistance but respond to thiazolidinediones. J. Clin. Investig. 2003, 112, 608–618. [Google Scholar] [CrossRef]
- Harada, K.; Isse, K.; Kamihira, T.; Shimoda, S.; Nakanuma, Y. Th1 cytokine-induced downregulation of PPARgamma in human biliary cells relates to cholangitis in primary biliary cirrhosis. Hepatology 2005, 41, 1329–1338. [Google Scholar] [CrossRef]
- Ahmadian, M.; Suh, J.M.; Hah, N.; Liddle, C.; Atkins, A.R.; Downes, M.; Evans, R.M. PPARγ signaling and metabolism: The good, the bad and the future. Nat. Med. 2013, 19, 557–566. [Google Scholar] [CrossRef]
- Nan, Y.M.; Han, F.; Kong, L.B.; Zhao, S.X.; Wang, R.Q.; Wu, W.J.; Yu, J. Adenovirus-mediated peroxisome proliferator activated receptor gamma overexpression prevents nutritional fibrotic steatohepatitis in mice. Scand. J. Gastroenterol. 2011, 46, 358–369. [Google Scholar] [CrossRef]
- Gupte, A.A.; Liu, J.Z.; Ren, Y.; Minze, L.J.; Wiles, J.R.; Collins, A.R.; Lyon, C.J.; Pratico, D.; Finegold, M.J.; Wong, S.T.; et al. Rosiglitazone attenuates age- and diet-associated nonalcoholic steatohepatitis in male low-density lipoprotein receptor knockout mice. Hepatology 2010, 52, 2001–2011. [Google Scholar] [CrossRef]
- Kim, J.W.; Yang, D.; Jeong, H.; Park, I.S.; Lee, M.H.; Lim, C.W.; Kim, B. Dietary zerumbone, a sesquiterpene, ameliorates hepatotoxin-mediated acute and chronic liver injury in mice. Phytother. Res. PTR 2019, 33, 1538–1550. [Google Scholar] [CrossRef]
- Roehlen, N.; Crouchet, E.; Baumert, T.F. Liver Fibrosis: Mechanistic Concepts and Therapeutic Perspectives. Cells 2020, 9, 875. [Google Scholar] [CrossRef]
- Wu, J.W.; Lin, L.C.; Hung, S.C.; Chi, C.W.; Tsai, T.H. Analysis of silibinin in rat plasma and bile for hepatobiliary excretion and oral bioavailability application. J. Pharm. Biomed. Anal. 2007, 45, 635–641. [Google Scholar] [CrossRef]
- Faghihzadeh, F.; Adibi, P.; Rafiei, R.; Hekmatdoost, A. Resveratrol supplementation improves inflammatory biomarkers in patients with nonalcoholic fatty liver disease. Nutr. Res. 2014, 34, 837–843. [Google Scholar] [CrossRef]
- Wang, M.; Chen, Z.; Yang, L.; Ding, L. Sappanone A Protects Against Inflammation, Oxidative Stress and Apoptosis in Cerebral Ischemia-Reperfusion Injury by Alleviating Endoplasmic Reticulum Stress. Inflammation 2021, 44, 934–945. [Google Scholar] [CrossRef] [PubMed]
- Shi, X.; Li, Y.; Wang, Y.; Ding, T.; Zhang, X.; Wu, N. Pharmacological postconditioning with sappanone A ameliorates myocardial ischemia reperfusion injury and mitochondrial dysfunction via AMPK-mediated mitochondrial quality control. Toxicol. Appl. Pharmacol. 2021, 427, 115668. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.; Choi, S.Y.; Choo, Y.Y.; Kim, O.; Tran, P.T.; Dao, C.T.; Min, B.S.; Lee, J.H. Sappanone A exhibits anti-inflammatory effects via modulation of Nrf2 and NF-κB. Int. Immunopharmacol. 2015, 28, 328–336. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Yu, D.; Wang, T. Sappanone A Attenuates Allergic Airway Inflammation in Ovalbumin-Induced Asthma. Int. Arch. Allergy Immunol. 2016, 170, 180–186. [Google Scholar] [CrossRef]
- Choo, Y.Y.; Tran, P.T.; Min, B.S.; Kim, O.; Nguyen, H.D.; Kwon, S.H.; Lee, J.H. Sappanone A inhibits RANKL-induced osteoclastogenesis in BMMs and prevents inflammation-mediated bone loss. Int. Immunopharmacol. 2017, 52, 230–237. [Google Scholar] [CrossRef]
- Zhou, Z.; Qi, J.; Zhao, J.; Lim, C.W.; Kim, J.W.; Kim, B. Dual TBK1/IKKε inhibitor amlexanox attenuates the severity of hepatotoxin-induced liver fibrosis and biliary fibrosis in mice. J. Cell. Mol. Med. 2020, 24, 1383–1398. [Google Scholar] [CrossRef]
- Xu, Q.; Liu, M.; Chao, X.; Zhang, C.; Yang, H.; Chen, J.; Zhou, B. Stevioside Improves Antioxidant Capacity and Intestinal Barrier Function while Attenuating Inflammation and Apoptosis by Regulating the NF-κB/MAPK Pathways in Diquat-Induced Oxidative Stress of IPEC-J2 Cells. Antioxidants 2023, 12, 1070. [Google Scholar] [CrossRef]
- Zhou, Z.; Qi, J.; Wu, Y.; Li, C.; Bao, W.; Lin, X.; Zhu, A. Nuciferine Effectively Protects Mice against Acetaminophen-Induced Liver Injury. Antioxidants 2023, 12, 949. [Google Scholar] [CrossRef]
- Huang, Y.; Wu, H.; Hu, Y.; Zhou, C.; Wu, J.; Wu, Y.; Wang, H.; Lenahan, C.; Huang, L.; Nie, S.; et al. Puerarin Attenuates Oxidative Stress and Ferroptosis via AMPK/PGC1α/Nrf2 Pathway after Subarachnoid Hemorrhage in Rats. Antioxidants 2022, 11, 1259. [Google Scholar] [CrossRef]
- Zhou, Z.; Wu, Y.; Hua, W.; Yan, X.; Li, L.; Zhu, A.; Qi, J. Sappanone A ameliorates acetaminophen-induced acute liver injury in mice. Toxicology 2022, 480, 153336. [Google Scholar] [CrossRef]
- Qi, J.; Kim, J.W.; Zhou, Z.; Lim, C.W.; Kim, B. Ferroptosis Affects the Progression of Nonalcoholic Steatohepatitis via the Modulation of Lipid Peroxidation-Mediated Cell Death in Mice. Am. J. Pathol. 2020, 190, 68–81. [Google Scholar] [CrossRef] [PubMed]
- Ruart, M.; Chavarria, L.; Campreciós, G.; Suárez-Herrera, N.; Montironi, C.; Guixé-Muntet, S.; Bosch, J.; Friedman, S.L.; Garcia-Pagán, J.C.; Hernández-Gea, V. Impaired endothelial autophagy promotes liver fibrosis by aggravating the oxidative stress response during acute liver injury. J. Hepatol. 2019, 70, 458–469. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Z.; Kim, J.W.; Qi, J.; Eo, S.K.; Lim, C.W.; Kim, B. Toll-Like Receptor 5 Signaling Ameliorates Liver Fibrosis by Inducing Interferon β-Modulated IL-1 Receptor Antagonist in Mice. Am. J. Pathol. 2020, 190, 614–629. [Google Scholar] [CrossRef] [PubMed]
- Cheng, D.; Chai, J.; Wang, H.; Fu, L.; Peng, S.; Ni, X. Hepatic macrophages: Key players in the development and progression of liver fibrosis. Liver Int. 2021, 41, 2279–2294. [Google Scholar] [CrossRef]
- Cheng, P.; Xie, J.; Liu, Z.; Wang, J. Aldose reductase deficiency inhibits LPS-induced M1 response in macrophages by activating autophagy. Cell Biosci. 2021, 11, 61. [Google Scholar] [CrossRef]
- Raghupathi, W.; Raghupathi, V. An Empirical Study of Chronic Diseases in the United States: A Visual Analytics Approach. Int. J. Environ. Res. Public Health 2018, 15, 431. [Google Scholar] [CrossRef]
- Rana, J.S.; Khan, S.S.; Lloyd-Jones, D.M.; Sidney, S. Changes in Mortality in Top 10 Causes of Death from 2011 to 2018. J. Gen. Intern. Med. 2021, 36, 2517–2518. [Google Scholar] [CrossRef]
- Wynn, T.A. Common and unique mechanisms regulate fibrosis in various fibroproliferative diseases. J. Clin. Investig. 2007, 117, 524–529. [Google Scholar] [CrossRef]
- Hillaire, S.; Cazals-Hatem, D.; Bruno, O.; de Miranda, S.; Grenet, D.; Poté, N.; Soubrane, O.; Erlinger, S.; Lacaille, F.; Mellot, F.; et al. Liver transplantation in adult cystic fibrosis: Clinical, imaging, and pathological evidence of obliterative portal venopathy. Liver Transplant. 2017, 23, 1342–1347. [Google Scholar] [CrossRef]
- Fang, C.; Jaffer, O.S.; Yusuf, G.T.; Konstantatou, E.; Quinlan, D.J.; Agarwal, K.; Quaglia, A.; Sidhu, P.S. Reducing the Number of Measurements in Liver Point Shear-Wave Elastography: Factors that Influence the Number and Reliability of Measurements in Assessment of Liver Fibrosis in Clinical Practice. Radiology 2018, 287, 844–852. [Google Scholar] [CrossRef]
- Seki, E.; Schwabe, R.F. Hepatic inflammation and fibrosis: Functional links and key pathways. Hepatology 2015, 61, 1066–1079. [Google Scholar] [CrossRef] [PubMed]
- Tewtrakul, S.; Tungcharoen, P.; Sudsai, T.; Karalai, C.; Ponglimanont, C.; Yodsaoue, O. Antiinflammatory and Wound Healing Effects of Caesalpinia sappan L. Phytother. Res. PTR 2015, 29, 850–856. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.Z.; Sun, S.Q.; Zhou, Y.B. Extract of the dried heartwood of Caesalpinia sappan L. attenuates collagen-induced arthritis. J. Ethnopharmacol. 2011, 136, 271–278. [Google Scholar] [CrossRef]
- Brempelis, K.J.; Crispe, I.N. Infiltrating monocytes in liver injury and repair. Clin. Transl. Immunol. 2016, 5, e113. [Google Scholar] [CrossRef] [PubMed]
- Krenkel, O.; Tacke, F. Liver macrophages in tissue homeostasis and disease. Nat. Rev. Immunol. 2017, 17, 306–321. [Google Scholar] [CrossRef]
- Zhang, J.; Liu, Y.; Chen, H.; Yuan, Q.; Wang, J.; Niu, M.; Hou, L.; Gu, J.; Zhang, J. MyD88 in hepatic stellate cells enhances liver fibrosis via promoting macrophage M1 polarization. Cell Death Dis. 2022, 13, 411. [Google Scholar] [CrossRef]
- Xie, C.; Wan, L.; Li, C.; Feng, Y.; Kang, Y.J. Selective suppression of M1 macrophages is involved in zinc inhibition of liver fibrosis in mice. J. Nutr. Biochem. 2021, 97, 108802. [Google Scholar] [CrossRef]
- Rao, J.; Wang, H.; Ni, M.; Wang, Z.; Wang, Z.; Wei, S.; Liu, M.; Wang, P.; Qiu, J.; Zhang, L.; et al. FSTL1 promotes liver fibrosis by reprogramming macrophage function through modulating the intracellular function of PKM2. Gut 2022, 71, 2539–2550. [Google Scholar] [CrossRef]
- Bai, L.; Liu, X.; Zheng, Q.; Kong, M.; Zhang, X.; Hu, R.; Lou, J.; Ren, F.; Chen, Y.; Zheng, S.; et al. M2-like macrophages in the fibrotic liver protect mice against lethal insults through conferring apoptosis resistance to hepatocytes. Sci. Rep. 2017, 7, 10518. [Google Scholar] [CrossRef]
- Ramachandran, P.; Pellicoro, A.; Vernon, M.A.; Boulter, L.; Aucott, R.L.; Ali, A.; Hartland, S.N.; Snowdon, V.K.; Cappon, A.; Gordon-Walker, T.T.; et al. Differential Ly-6C expression identifies the recruited macrophage phenotype, which orchestrates the regression of murine liver fibrosis. Proc. Natl. Acad. Sci. USA 2012, 109, E3186–E3195. [Google Scholar] [CrossRef]
- Wu, L.; Guo, C.; Wu, J. Therapeutic potential of PPARγ natural agonists in liver diseases. J. Cell. Mol. Med. 2020, 24, 2736–2748. [Google Scholar] [CrossRef]
- Ye, L.; Chen, T.; Cao, J.; Sun, L.; Li, W.; Zhang, C. Short hairpin RNA attenuates liver fibrosis by regulating the PPAR-γ and NF-κB pathways in HBV-induced liver fibrosis in mice. Int. J. Oncol. 2020, 57, 1116–1128. [Google Scholar] [CrossRef]
- Feng, X.; Weng, D.; Zhou, F.; Owen, Y.D.; Qin, H.; Zhao, J.; Yu, W.; Huang, Y.; Chen, J.; Fu, H.; et al. Activation of PPARγ by a Natural Flavonoid Modulator, Apigenin Ameliorates Obesity-Related Inflammation Via Regulation of Macrophage Polarization. EBioMedicine 2016, 9, 61–76. [Google Scholar] [CrossRef] [PubMed]
- Schofield, R. Assessment of cytotoxic injury to bone marrow. Br. J. Cancer Suppl. 1986, 7, 115–125. [Google Scholar] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Groups | Volume of Corn Oil (Body Weight) | Volume of CCl4 (CCl4:Corn Oil = 2:5, v/v) (Body Weight) | Dose of SA or Vehicle (Body Weight) | Animal Number |
|---|---|---|---|---|
| 1 | 2 mL/kg, i.p. | _ | PBS (200 μL, i.p.) | n = 8 |
| 2 | 2 mL/kg, i.p. | _ | 25 mg/kg SA (200 μL, i.p.) | n = 8 |
| 3 | 2 mL/kg, i.p. | _ | 50 mg/kg SA (200 μL, i.p.) | n = 8 |
| 4 | 2mL/kg, i.p. | _ | 100 mg/kg SA (200 μL, i.p.) | n = 8 |
| 5 | _ | 2 mL/kg, i.p. | PBS (200 μL, i.p.) | n = 10 |
| 6 | _ | 2 mL/kg, i.p. | 25 mg/kg SA (200 μL, i.p.) | n = 10 |
| 7 | _ | 2 mL/kg, i.p. | 50 mg/kg SA (200 μL, i.p.) | n = 10 |
| 8 | _ | 2 mL/kg, i.p. | 100 mg/kg SA (200 μL, i.p.) | n = 10 |
| Gene | Gene Accession Number | Forward (5′-3′) | Reverse (5′-3′) |
|---|---|---|---|
| αSMA | NM_007392 | 5′-TGCAGGTGATGCTGACAGAGG-3′ | 5′-GGGATGAGCTAGTGCTGATCTGG-3′ |
| Col1 | NM_007742 | 5′-CAGCCAATCAGCGTTCGGTA-3′ | 5′-CTTCATGGCGTAGTTGAATGATGTC-3′ |
| TGFβ | NM_001013025 | 5′-AGTTGGAGCAGCTGTATCAGTGG-3′ | 5′-TTTAGCAAAGGCAGTCAAATCTGG-3′ |
| Timp1 | NM_001044384 | 5′-AATGACTGTTGCCAGGTGGATG-3′ | 5′-GGTTGCACTTCCAAATGAGGCTA-3′ |
| Bax | NM_007527 | 5′-AAGCGCTTCGGGCCAG-3′ | 5′-TAGCCATGCAGGACCACGA-3′ |
| Bcl2 | NM_177410 | 5′-GGTCAGAAAGCCGTGGTTG-3′ | 5′-GACATGGCCTAACGTGCAG-3′ |
| TNFα | NM_013693.3 | 5′-CGCCTCTCCTTGCTGTCACA-3′ | 5′-CTTTGCCTTCTGCCTCAAGT-3′ |
| IL-1β | NM_008361.4 | 5′-GTCTACTCCCAGGTTTCTCTTCAAGG-3′ | 5′-GCAAATCGGCTGACGGTGTG-3′ |
| IL-6 | NM_001314054.1 | 5′-CTCGCAGCAGCACATCAACA-3′ | 5′-CCACGGGAAAGACACAGGTA-3′ |
| iNOS | NM_010927 | 5′-TGCACCCAAACCGAAGTC-3′ | 5′-GTCAGAAGCCAGCGTTCACC-3′ |
| Arg1 | NM_007482 | 5′-GCCAAGGGTTGACTTCAAGAACA-3′ | 5′-AGGCTCCTCCTTTCCAGGTCA-3′ |
| IL-10 | NM_010548 | 5′-AGCAGCAGGTGTCCCAAAGA-3′ | 5′-GTGCTGAAGACCTTAGGGCAGA-3′ |
| PPARγ | NM_133249 | 5′-ACGGCAAATTCAACGGCACAG-3′ | 5′-GAAGACTCCACGACATACTCAGCAC-3′ |
| hαSMA | NM_001613 | 5′-ATAGAACATGGCATCATCACCAAC-3′ | 5′-GGGCAACACGAAGCTCATTGTA-3′ |
| hCol1 | NM_000088 | 5′-GCTTGGTCCACTTGCTTGAAGA-3′ | 5′-GAGCATTGCCTTTGATTGCTG-3′ |
| GAPDH | NM_001411840.1 | 5′-ACGGCAAATTCAACGGCACAG-3′ | 5′-AGACTCCACGACATACTCAGCAC-3′ |
| hGAPDH | NM_001256799 | 5′-GCACCGTCAAGGCTGAGAAC-3′ | 5′-TGGTGAAGACGCCAGTGGA-3′ |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Qi, J.; Li, L.; Yan, X.; Hua, W.; Zhou, Z. Sappanone A Alleviates the Severity of Carbon Tetrachloride-Induced Liver Fibrosis in Mice. Antioxidants 2023, 12, 1718. https://doi.org/10.3390/antiox12091718
Qi J, Li L, Yan X, Hua W, Zhou Z. Sappanone A Alleviates the Severity of Carbon Tetrachloride-Induced Liver Fibrosis in Mice. Antioxidants. 2023; 12(9):1718. https://doi.org/10.3390/antiox12091718
Chicago/Turabian StyleQi, Jing, Lanqian Li, Xueqing Yan, Wenxi Hua, and Zixiong Zhou. 2023. "Sappanone A Alleviates the Severity of Carbon Tetrachloride-Induced Liver Fibrosis in Mice" Antioxidants 12, no. 9: 1718. https://doi.org/10.3390/antiox12091718
APA StyleQi, J., Li, L., Yan, X., Hua, W., & Zhou, Z. (2023). Sappanone A Alleviates the Severity of Carbon Tetrachloride-Induced Liver Fibrosis in Mice. Antioxidants, 12(9), 1718. https://doi.org/10.3390/antiox12091718