
Pepper Fruit Extracts Show Anti-Proliferative Activity against Tumor Cells Altering Their NADPH-Generating Dehydrogenase and Catalase Profiles

 ,
,  ,
,  and
and 
Abstract
1. Introduction
2. Materials and Methods
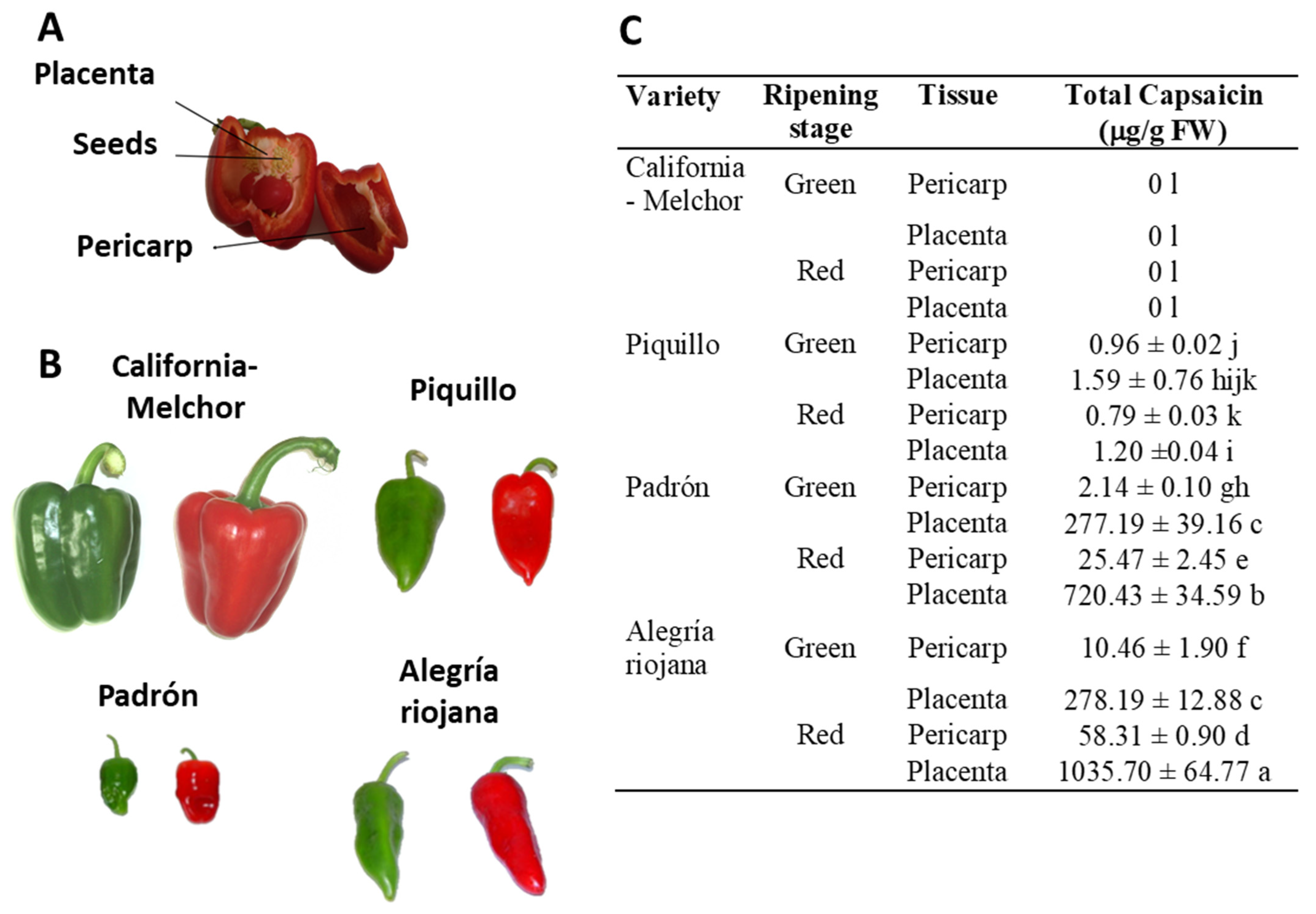
2.1. Plant Material and Preparation of Crude Pepper Fruit Extracts
2.2. Determination of Capsaicin
2.3. Tumor Cell Lines, Anti-Proliferative Activity Assays, and Preparation of Crude Extracts
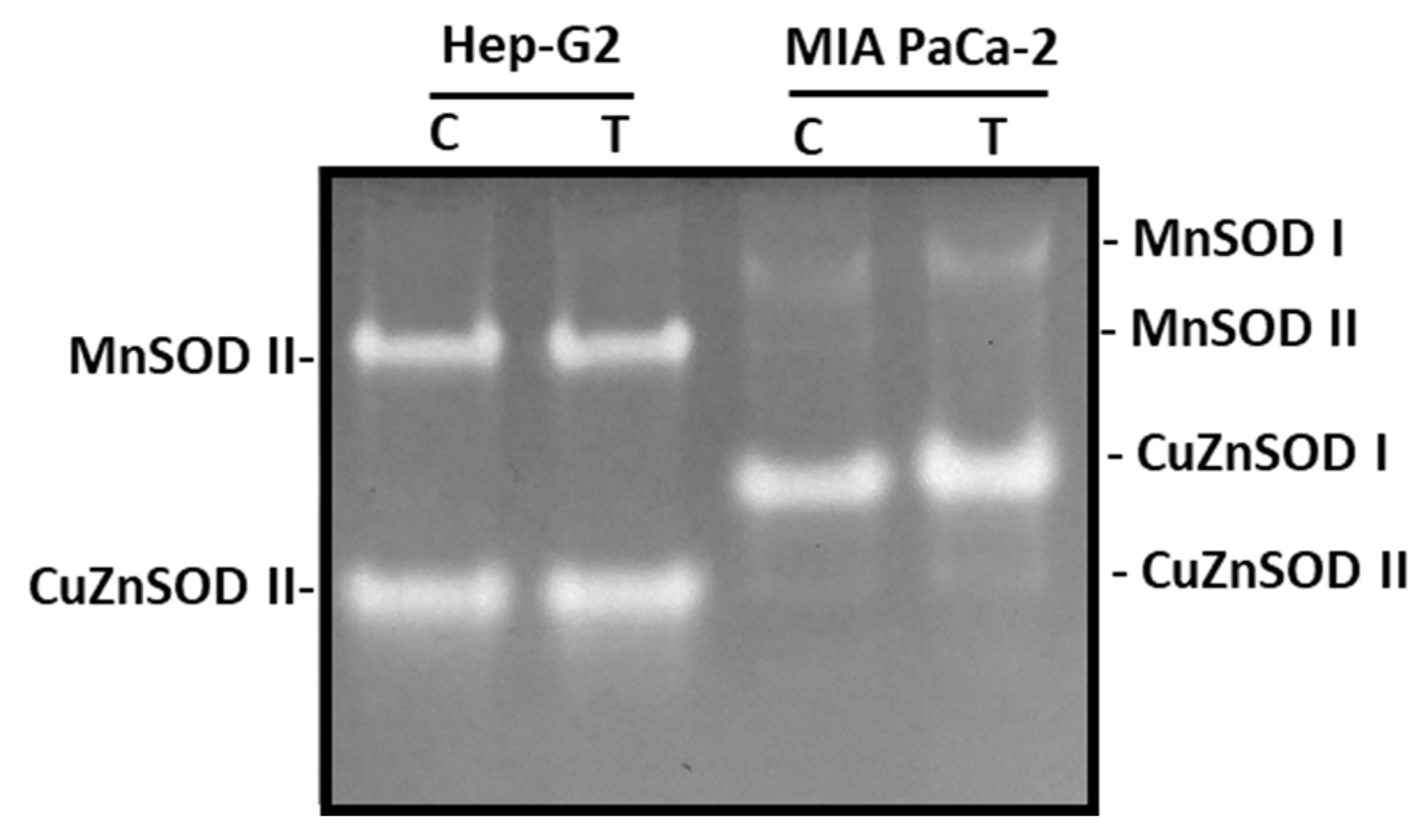
2.4. Enzyme Activities
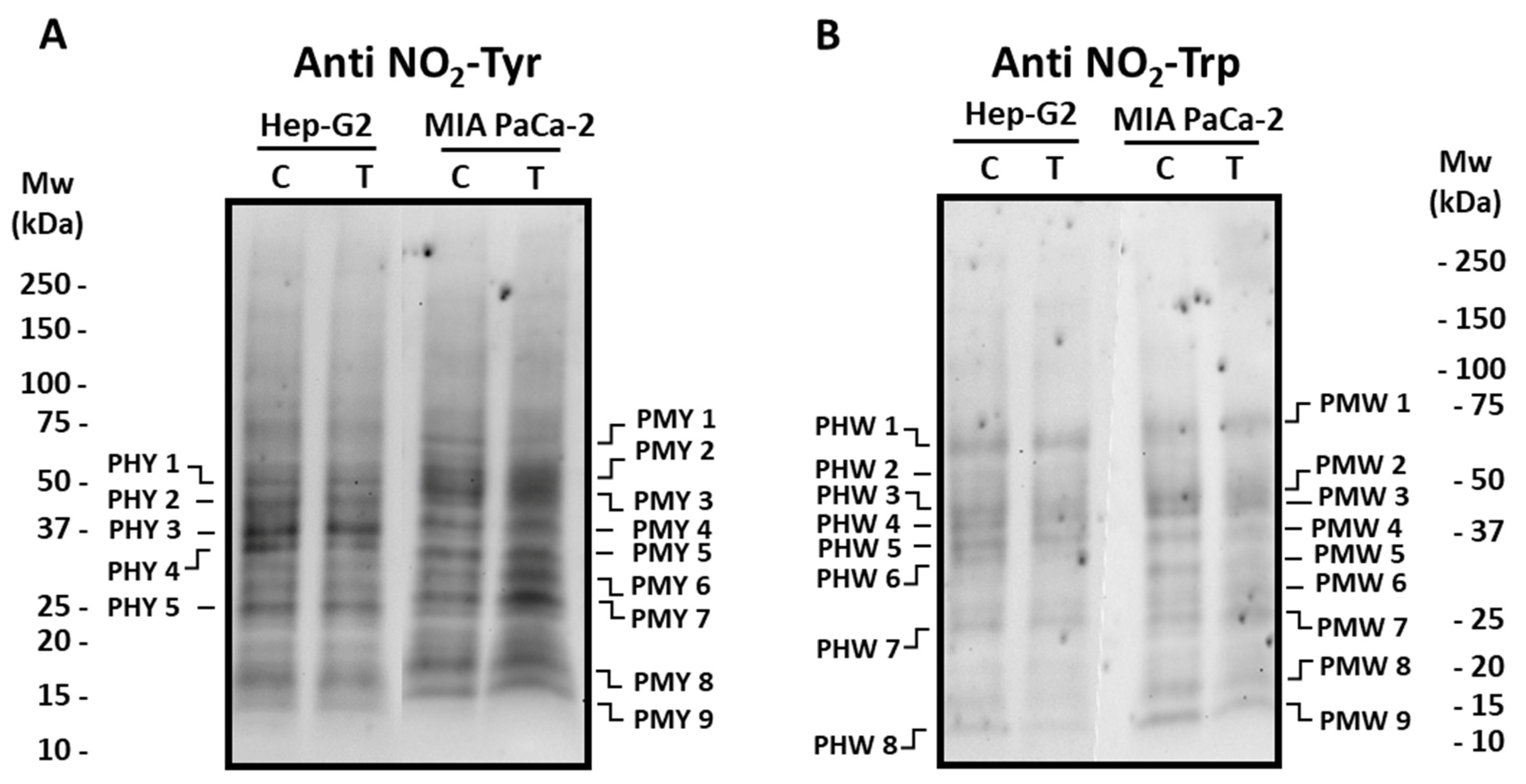
2.5. SDS-PAGE and Immunoblot Analyses
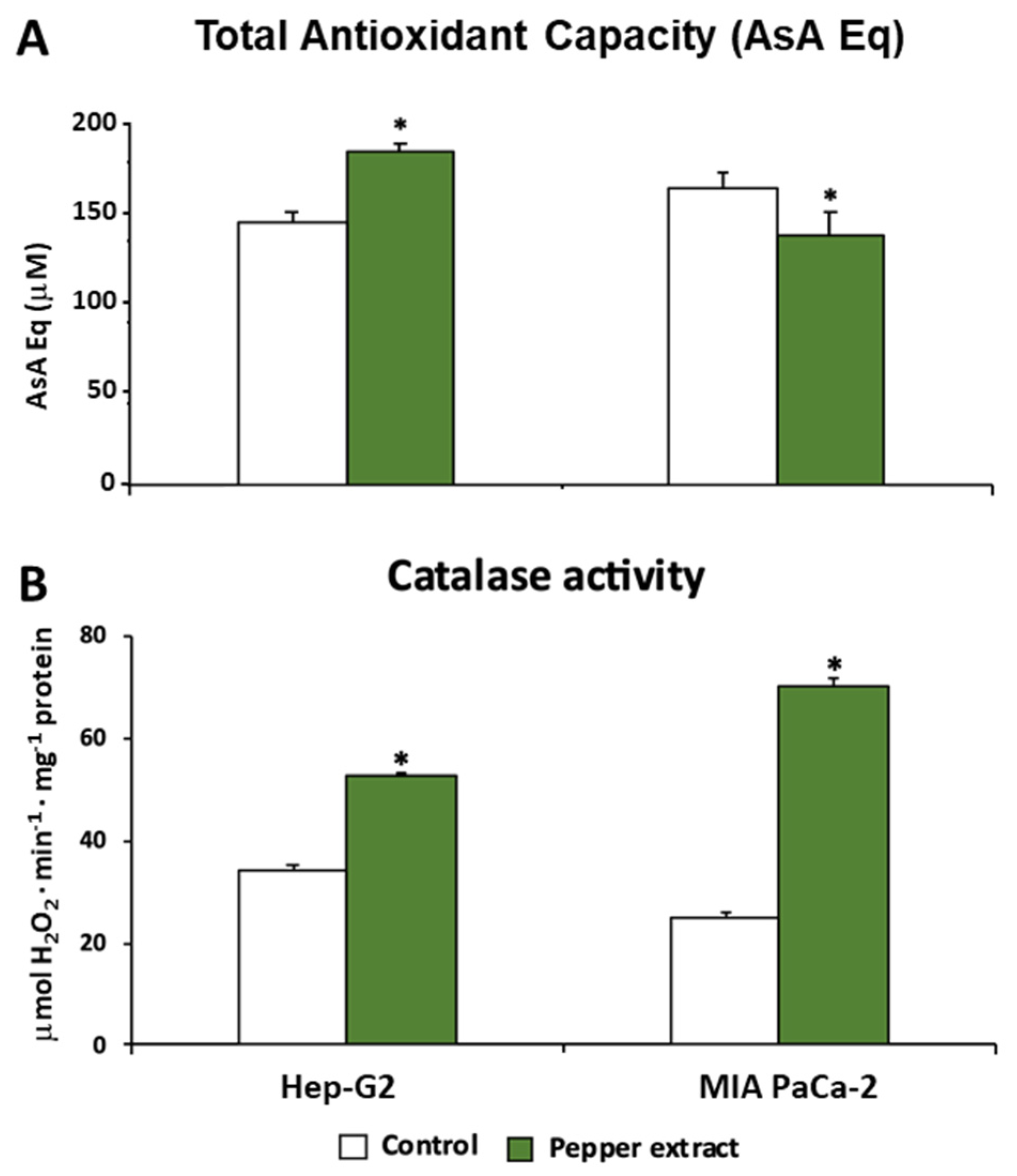
2.6. Antioxidant Capacity Assay
2.7. Statistical Analysis
3. Results
3.1. Crude Extracts from Pepper Fruit Show Anti-Proliferative Activity against Tumor Cell Lines
3.2. Pepper Fruit Extracts Slightly Alter the Nitro-Oxidative Status of Tumor Cells
3.3. ROS Metabolism in Tumor Cells under Pepper Fruit Treatment
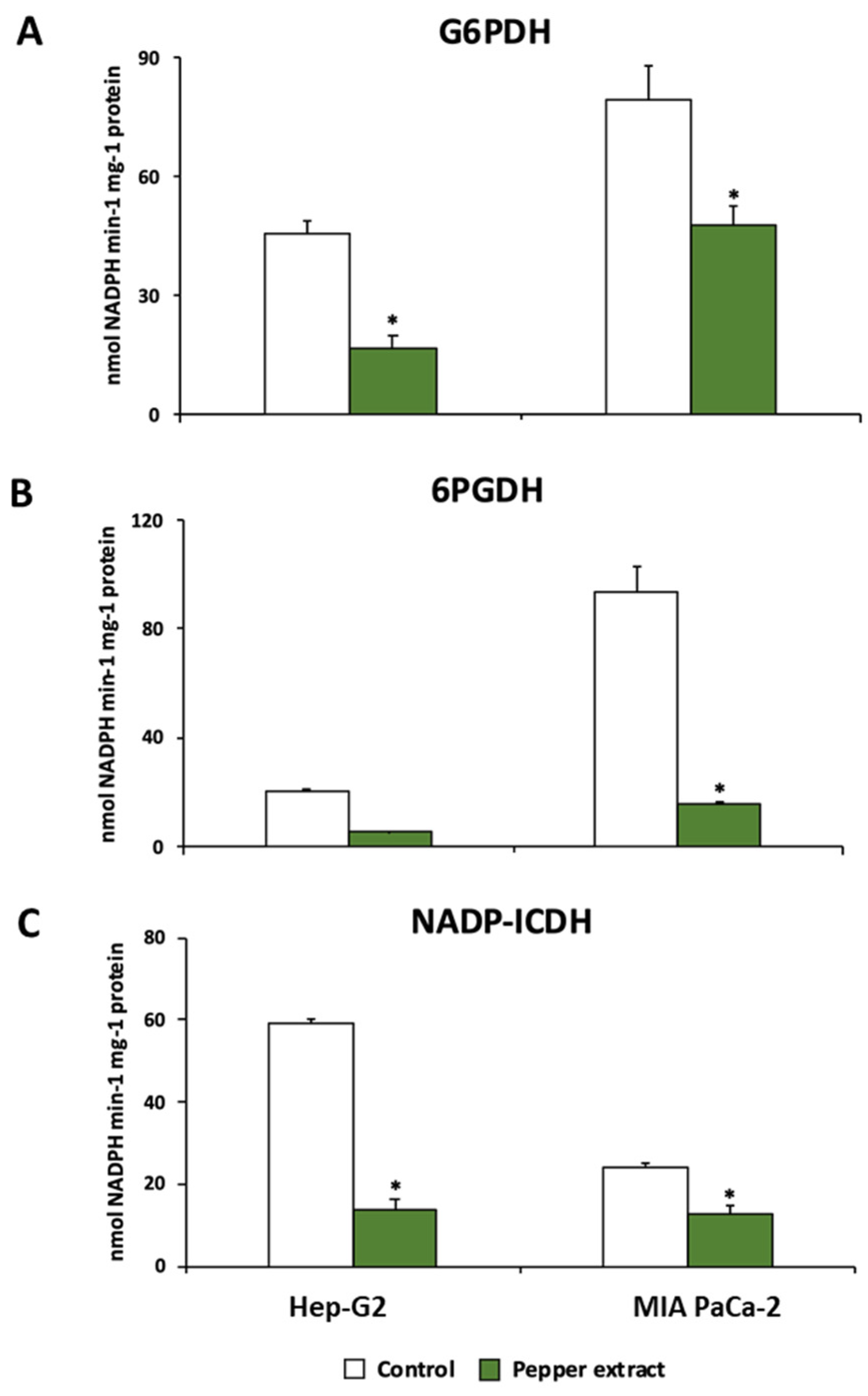
3.4. Pepper Fruit Treatment Disturbs the NADPH-Generating Systems in Tumor Cells
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Klaunik, J.E. Oxidative stress and cancer. Curr. Pharm. Des. 2018, 24, 4771–4778. [Google Scholar] [CrossRef] [PubMed]
- Tas, F.; Hansel, H.; Belce, A.; Ilvan, S.; Argon, A.; Camlica, H. Topuz, Oxidative stress in breast cancer. Med. Oncol. 2005, 22, 11–15. [Google Scholar] [CrossRef] [PubMed]
- Shen, J.; Gammon, M.D.; Terry, M.B.; Wang, Q.; Bradshaw, P.; Teitelbaum, S.L.; Neugut, A.I.; Santella, R.M. Telomere length, oxidative damage, antioxidants and breast cancer risk. Int. J. Cancer 2009, 124, 1637–1643. [Google Scholar] [CrossRef] [PubMed]
- Lawless, M.W.; O’Byrne, K.J.; Gray, S.G. Targeting oxidative stress in cancer. Expert Opin. Ther. Targets 2010, 14, 1225–1245. [Google Scholar] [CrossRef]
- Tong, L.Y.; Chuang, C.C.; Wu, S.Y.; Zuo, L. Reactive oxygen species in redox cancer therapy. Cancer Lett. 2015, 367, 18–25. [Google Scholar] [CrossRef] [PubMed]
- Forouzandeh, H.; Kalantari, H.; Saki, N.; Foruozandeh, Z.; Arefian, E.; Farahani, A.; Hassani, G.; Bazrafshan, M.R. Rasouli, Role of oxidative stress in liver cancer. Clin. Cancer Investig. J. 2017, 6, 1–9. [Google Scholar] [CrossRef]
- Jelic, M.D.; Mandic, A.D.; Maricic, S.M.; Srdjenovic, B.U. Oxidative stress and its role in cancer. J. Cancer Res. Ther. 2021, 17, 22–28. [Google Scholar] [CrossRef]
- Cecerska-Heryc, E.; Surowska, O.; Heryc, R.L.; Serwin, N.; Napiontek-Balinska, S. Dolegowska, Are antioxidant enzymes essential markers in the diagnosis and monitoring of cancer patients?—A review. Clin. Biochem. 2021, 93, 1–8. [Google Scholar] [CrossRef]
- Oberley, T.D.; Oberley, L.W. Antioxidant enzyme levels in cancer. Histol. Histopathol. 1997, 12, 525–535. [Google Scholar]
- Cockfield, J.A.; Schafer, Z.T. Antioxidant defenses: A context-specific vulnerability of cancer cells. Cancers 2019, 11, 1208. [Google Scholar] [CrossRef]
- George, S.; Abrahamse, H. Redox Potential of antioxidants in cancer progression and prevention. Antioxidants 2020, 9, 1156. [Google Scholar] [CrossRef]
- Taucher, E.; Mykoliuk, I.; Fediuk, M.; Smolle-Juettner, F.M. Autophagy, oxidative stress and cancer development. Cancers 2022, 14, 1637. [Google Scholar] [CrossRef] [PubMed]
- Banerjee, S.; Li, Y.W.; Wang, Z.W.; Sarkar, F.H. Multi-targeted therapy of cancer by genistein. Cancer Lett. 2008, 269, 226–242. [Google Scholar] [CrossRef] [PubMed]
- Greenlee, H.; Kwan, M.L.; Kushi, L.H.; Song, J.; Castillo, A.; Weltzien, E.; Quesenberry, C.P., Jr.; Caan, B.J. Antioxidant supplement use after breast cancer diagnosis and mortality in the Life After Cancer Epidemiology LACE cohort. Cancer 2012, 118, 2048–2058. [Google Scholar] [CrossRef]
- Moradi-Joo, M.; Heidari, S.; Seyed-Nezhad, M.; Akbari, M.E.; Moosavi, A.; Davoodi, S.H. Antioxidant supplements and breast cancer: A systematic review and meta-analysis. Int. J. Cancer Manag. 2018, 11, 10082. [Google Scholar] [CrossRef]
- Jayaprakash, V.; Marshall, J.R. Selenium and other antioxidants for chemoprevention of gastrointestinal cancers. Best Pract. Res. Clin. Gastroenterol. 2011, 25, 507–518. [Google Scholar] [CrossRef]
- Han, X.S.; Li, J.J.; Brasky, T.M.; Xun, P.C.; Stevens, J.; White, E.; Gammon, M.D.; He, K. Antioxidant intake and pancreatic cancer risk: The VITamins And Lifestyle VITAL study. Cancer 2013, 119, 1314–1320. [Google Scholar] [CrossRef]
- Saha, S.K.; Lee, S.B.; Won, J.; Choi, H.Y.; Kim, K.; Yang, G.M.; Dayem, A.A.; Cho, S.G. Correlation between oxidative stress, nutrition, and cancer initiation. Int. J. Mol. Sci. 2017, 18, 1544. [Google Scholar] [CrossRef]
- Serafini, M.; Jakszyn, P.; Lujan-Barroso, L.; Agudo, A.; Bueno-de-Mesquita, H.B.; van Duijnhoven, F.J.B.; Jenab, M.; Navarro, C.; Palli, D.; Boeing, H. Dietary total antioxidant capacity and gastric cancer risk in the European prospective investigation into cancer and nutrition study. Int. J. Cancer. 2012, 131, E544–E554. [Google Scholar] [CrossRef]
- Roy, K.; Wu, Y.Z.; Meitzler, J.L.; Juhasz, A.; Liu, H.; Jiang, G.J.; Lu, J.M.; Antony, S.; Doroshow, J.H. NADPH oxidases and cancer. Clin. Sci. London 2015, 128, 863–875. [Google Scholar] [CrossRef]
- Rather, G.M.; Pramono, A.A.; Szekely, Z.; Bertino, J.R.; Tedeschi, P.M. In cancer, all roads lead to NADPH. Pharmacol. Therap. 2021, 226, 107864. [Google Scholar] [CrossRef] [PubMed]
- Ju, H.Q.; Lin, J.F.; Tian, T.; Xie, D.; Xu, R.H. NADPH homeostasis in cancer: Functions, mechanisms and therapeutic implications. Signal Trans. Targ. Ther. 2020, 5, 231. [Google Scholar] [CrossRef] [PubMed]
- Yang, H.C.; Wu, Y.H.; Yen, W.C.; Liu, H.Y.; Hwang, T.L.; Stern, A.; Chiu, D.T.Y. The Redox role of G6PD in cell growth, cell death, and cancer. Cells 2019, 8, 1055. [Google Scholar] [CrossRef] [PubMed]
- Yang, H.C.; Stern, A.; Chiu, D.T.Y. G6PD: A hub for metabolic reprogramming and redox signaling in cancer. Biomed. J. 2021, 44, 285–292. [Google Scholar] [CrossRef]
- Alfarouk, K.O.; Ahmed, S.B.M.; Elliott, R.L.; Benoit, A.; Alqahtani, S.S.; Ibrahim, M.E.; Bashir, A.H.H.; Alhoufie, S.T.S.; Elhassan, G.O.; Wales, C.C.; et al. The pentose phosphate pathway dynamics in cancer and its dependency on intracellular pH. Metabolites 2020, 10, 285. [Google Scholar] [CrossRef]
- Ge, T.X.; Yang, J.W.; Zhou, S.H.; Wang, Y.C.; Li, Y.K.; Tong, X.M. The role of the pentose phosphate pathway in diabetes and cancer. Front. Endocrinol. 2020, 11, 365. [Google Scholar] [CrossRef]
- Li, R.; Wan, W.; Yang, Y.; Gu, C.Y. Exploring the role of glucose-6-phosphate dehydrogenase in cancer. Oncol. Rep. 2020, 44, 2325–2336. [Google Scholar] [CrossRef]
- Liu, S.; Cadoux-Hudson, T.; Schofield, C.J. Isocitrate dehydrogenase variants in cancer—Cellular consequences and therapeutic opportunities. Curr. Op. Chem. Biol. 2020, 57, 122–134. [Google Scholar] [CrossRef]
- Simmen, F.A.; Alhallak, I.; Simmen, R.M.C. Malic enzyme 1 ME1 in the biology of cancer: It is not just intermediary metabolism. J. Mol. Endocrinol. 2020, 65, R77–R90. [Google Scholar] [CrossRef]
- Palma, J.M.; Seiquer, I. To be or not be…an antioxidant? That is the question. Antioxidants 2020, 9, 1234. [Google Scholar] [CrossRef]
- Li, W.; Li, J.; Zhao, J.; He, C. Evolutionary developmental genetics of fruit morphological variation within the Solanaceae. Front. Plant Sci. 2015, 6, 248. [Google Scholar]
- Gebhardt, C. The historical role of species from the Solanaceae plant family in genetic research. Theor. Appl. Genet. 2016, 129, 2281–2294. [Google Scholar] [CrossRef] [PubMed]
- Baenas, N.; Belović, N.; Ilicb, N.; Moreno, D.A.; García-Viguera, C. Industrial use of pepper Capsicum annum L. derived products: Technological benefits and biological advantages. Food Chem. 2019, 274, 872–885. [Google Scholar] [CrossRef] [PubMed]
- Corpas, F.J.; Freschi, L.; Palma, J.M. ROS metabolism and ripening of fleshy fruits. Adv. Bot. Res. 2023, 105, 205–238. [Google Scholar]
- Wahyuni, Y.; Ballester, A.R.; Sudarmonowati, E.; Bino, R.J.; Bovy, A.G. Secondary metabolites of Capsicum species and their importance in the human diet. J. Nat. Prod. 2013, 76, 783–793. [Google Scholar] [CrossRef]
- Li, Y.; Yao, J.; Han, C.; Yang, J.; Chaudhry, M.T.; Wang, S.; Liu, H.; Yin, Y. Quercetin, inflammation and immunity. Nutrients 2016, 8, 167. [Google Scholar] [CrossRef]
- Reyes-Farias, M.; Carrasco-Pozo, C. The anti-cancer effect of quercetin: Molecular implications in cancer metabolism. Int. J. Mol. Sci. 2019, 20, 3177. [Google Scholar] [CrossRef]
- Metsämuuronen, S.; Sirén, H. Bioactive phenolic compounds, metabolism and properties: A review on valuable chemical compounds in Scots pine and Norway spruce. Phytochem. Rev. 2019, 18, 623–664. [Google Scholar] [CrossRef]
- Batiha, G.E.S.; Alqahtani, A.; Oluwafemi, A.O.; Shaheen, H.M.; Wasef, L.; Elzeiny, M.; Ismail, M.; Shalaby, M.; Murata, T.; Zaragoza-Bastida, A.; et al. Hetta, Biological properties, bioactive constituents, and pharmacokinetics of some Capsicum spp. and capsaicinoids. Int. J. Mol. Sci. 2020, 21, 5179. [Google Scholar] [CrossRef]
- Dessalegn, E.; Bultosa, G.; Haki, G.D.; Chen, F.; Rupasinghe, H.P.V. Antioxidant and cytotoxicity to liver cancer HepG2 cells in vitro of Korarima Aframomum corrorima Braun PCM Jansen seed extracts. Int. J. Food Prop. 2022, 25, 1–10. [Google Scholar] [CrossRef]
- Guevara, L.; Domínguez-Anaya, M.A.; Ortigosa, A.; González-Gordo, S.; Díaz, C.; Vicente, F.; Corpas, F.J.; del Palacio, J.P.; Palma, J.M. Identification of compounds with potential therapeutic uses from sweet pepper Capsicum annuum L. fruits and their modulation by nitric oxide NO. Int. J. Mol. Sci. 2021, 22, 4476. [Google Scholar] [CrossRef] [PubMed]
- Cao, E.; Cordero-Morales, J.F.; Liu, B.Y.; Qin, F.; Julis, D. TRPV1 Channels are intrinsically heat sensitive and negatively regulated by phosphoinositide lipids. Neuron 2013, 77, 667–679. [Google Scholar] [CrossRef] [PubMed]
- Chapa-Oliver, A.M.; Mejía-Teniente, L. Capsaicin: From plants to a cancer-suppressing agent. Molecules 2016, 21, 931. [Google Scholar] [CrossRef] [PubMed]
- Clark, R.; Lee, S.H. Anticancer properties of capsaicin against human cancer. Anticaner Res. 2016, 36, 837–844. [Google Scholar]
- Georgescu, S.R.; Sârbu, M.I.; Matei, C.; Ilie, M.A.; Caruntu, C.; Constantin, C.; Tampa, M.N.M. Capsaicin: Friend or foe in skin cancer and other related malignancies? Nutrients 2017, 9, 1365. [Google Scholar] [CrossRef] [PubMed]
- Tabrizi, M.A.; Baraldi, P.G.; Baraldi, S.; Gessi, S.; Merighi, S.; Borea, P.A. Medicinal chemistry; pharmacology, and clinical implications of TRPV1 receptor antagonists. Med. Res. Rev. 2017, 37, 936–983. [Google Scholar] [CrossRef]
- Yang, F.; Zheng, J. Understand spiciness: Mechanism of TRPV1 channel activation by capsaicin. Protein Cell 2017, 8, 169–177. [Google Scholar] [CrossRef]
- Zhang, K.H.; Julis, D.; Cheng, Y.F. Structural snapshots of TRPV1 reveal mechanism of polymodal functionality. Cell 2021, 184, 5138–5150. [Google Scholar] [CrossRef]
- Karasawa, M.M.G.; Mohan, C. Fruits as prospective reserves of bioactive compounds: A review. Nat. Prod. Bioprospect. 2018, 8, 335–346. [Google Scholar] [CrossRef]
- Yoshida, Y.; Koyama, N.; Tamura, H. Color and anthocyanin composition of strawberry fruit: Changes during fruit development and differences among cultivars, with special reference to the occurrence of pelargonidin 3-malonylglucoside. J. Jpn. Soc. Hortic. Sci. 2020, 7, 355–361. [Google Scholar] [CrossRef]
- Palma, J.M.; Terán, F.; Contreras-Ruiz, A.; Rodríguez-Ruiz, M.; Corpas, F.J. Antioxidant profile of pepper Capsicum annuum L. fruits containing diverse levels of capsaicinoids. Antioxidants 2020, 9, 878. [Google Scholar] [CrossRef] [PubMed]
- Mao, Z.; Liu, Z.; Chen, L.; Yang, J.; Zhao, B.; Jung, Y.M.; Wang, X.; Zhao, C. Predictive value of the surface-enhanced resonance raman scattering-based MTT assay: A rapid and ultrasensitive method for cell viability in situ. Anal. Chem. 2013, 85, 7361–7368. [Google Scholar] [CrossRef] [PubMed]
- Smith, P.K.; Krohn, R.I.; Hermanson, G.T.; Mallia, A.K.; Gartner, F.H.; Provenzano, M.D.; Fujimoto, E.K.; Goeke, N.M.; Olson, B.J.; Klenk, D.C. Measurement of protein using bicinchoninic acid. Anal. Biochem. 1985, 150, 76–85. [Google Scholar] [CrossRef] [PubMed]
- Aebi, H. Catalase in vitro. Methods Enzymol. 1984, 105, 121–126. [Google Scholar]
- Corpas, F.J.; Barroso, J.B.; Sandalio, L.M.; Distefano, S.; Palma, J.M.; Lupiáñez, J.A.; del Río, L.A. A dehydrogenase-mediated recycling system of NADPH peroxisomes. Biochem. J. 1998, 330, 777–784. [Google Scholar] [CrossRef]
- Corpas, F.J.; Barroso, J.B.; Sandalio, L.M.; Palma, J.M.; Lupiáñez, J.A.; del Río, L.A. Peroxisomal NADP-dependent isocitrate dehydrogenase. characterization and activity regulation during natural senescence. Plant Physiol. 1999, 121, 921–928. [Google Scholar] [CrossRef]
- Beauchamp, C.M.; Fridovich, I. Superoxide dismutase. Improved assay and an assay applicable to acrylamide gels. Anal. Biochem. 1971, 44, 276–287. [Google Scholar] [CrossRef]
- Lin, C.L.; Chen, H.J.; Hou, W.C. Activity staining of glutathione peroxidase after electrophoresis on native and sodium dodecyl sulfate polyacrylamide gels. Electrophoresis 2002, 23, 513–516. [Google Scholar] [CrossRef]
- Roskoski, R., Jr. ERK1/2 MAP kinases: Structure; function; regulation. Pharmacol. Res. 2012, 66, 105–143. [Google Scholar] [CrossRef]
- Raskin, I.; Ribnicky, D.M.; Komarnytsky, S.; Ilic, N.; Poulev, A.; Borisjuk, N.; Brinker, A.; Moreno, D.A.; Ripoll, C.; Yakoby, N.; et al. Plants and human health in the twenty-first century. Trends Biotechnol. 2020, 20, 522–531. [Google Scholar] [CrossRef]
- Salmerón-Manzano, E.; Garrido-Cardenas, J.A.; Manzano-Agugliaro, F. Worldwide research trends on medicinal plants. Int. J. Environ. Res. Public Health 2020, 17, 3376. [Google Scholar] [CrossRef] [PubMed]
- Pattanayak, S. Plants in healthcare: Past, present and future. Explor. Anim. Med. Res. 2022, 11, 140–144. [Google Scholar] [CrossRef]
- Palma, J.M.; del Palacio, J.P.; Rodríguez-Ruiz, M.; González-Gordo, S.; Díaz, C.; Ramos, C.; Cautain, B.; Vicente, F.; Corpas, F.J. Pepper Fruit as a Nutraceutical Food with Antiproliferative Activity against Tumor Cells Potentiated by Nitric Oxide NO. In Nitric Oxide in Health and Disease: Therapeutic Applications in Cancer and Inflammatory Disorders; Morbidelli, L., Bonavida, B., Muntané, J., Eds.; Academic Press: Cambridge, MA, USA, 2023; pp. 193–210. [Google Scholar]
- Materska, M.; Perucka, I. Antioxidant activity of the main phenolic compounds isolated from hot pepper fruit Capsicum annuum L. J. Agric. Food Chem. 2005, 53, 1750–1756. [Google Scholar] [CrossRef] [PubMed]
- Roskoski, R., Jr. Targeting ERK1/2 protein-serine/threonine kinases in human cancers. Pharmacol. Res. 2019, 142, 151–168. [Google Scholar] [CrossRef]
- Leon, L.; Jeannin, J.F.; Bettaieb, A. Post-translational modifications induced by nitric oxide (NO): Implication in cancer cells apoptosis. Nitric Oxide Biol. Chem. 2008, 19, 77–83. [Google Scholar] [CrossRef]
- Moldogazieva, N.T.; Lutsenko, S.V.; Terentiev, A.A. Reactive oxygen and nitrogen species-induced protein modifications: Implication in carcinogenesis and anticancer therapy. Cancer Res. 2018, 78, 6040–6047. [Google Scholar] [CrossRef] [PubMed]
- Aguilar-Melero, P.; Ferrin, G.; Muntané, J. Effects of nitric oxide synthase-3 overexpres sion on post-translational modifications and cell survival in HepG2 cells. J. Proteomics 2012, 75, 740–755. [Google Scholar] [CrossRef] [PubMed]
- Corpas, F.J.; González-Gordo, S.; Palma, J.M. Nitric oxide: A radical molecule with potential biotechnological applications in fruit ripening. J. Biotechnol. 2020, 20, 211–219. [Google Scholar] [CrossRef]
- Musaogullari, A.; Chai, Y.C. Redox regulation by protein s-glutathionylation: From molecular mechanisms to implications in health and disease. Int. J. Mol. Sci. 2020, 21, 8113. [Google Scholar] [CrossRef]
- Pal, D.; Rai, A.; Checker, R.; Patwardhan, R.S.; Singh, B.; Sharma, D.; Sandur, S.K. Role of protein S-Glutathionylation in cancer progression and development of resistance to anti-cancer drugs. Arch. Biochem. Biophys. 2021, 704, 108890. [Google Scholar] [CrossRef]
- Skrzycki, M.; Majewska, M.; Czeczot, H. Superoxide dismutase mRNA and protein level in human colorectal cáncer. Cent. Eur. J. Biol. 2010, 5, 590–599. [Google Scholar] [CrossRef]
- Shah, M.H.; Liu, G.S.; Thompson, E.W.; Dusting, G.J.; Peshavariya, H.M. Differential effects of superoxide dismutase and superoxide dismutase/catalase mimetics on human breast cancer cells. Breast Cancer Res. Treat. 2015, 150, 523–534. [Google Scholar] [CrossRef] [PubMed]
- Liu, M.; Sun, X.; Chen, B.; Dai, R.; Xi, Z.; Xu, H. Insights into manganese superoxide dismutase and human diseases. Int. J. Mol. Sci. 2022, 23, 24. [Google Scholar] [CrossRef] [PubMed]
- Ogle, M.M., Jr.; Trevino, R., Jr.; Schell, J.; Varmazyad, M.; Horikoshi, N.; Gius, D. Manganese superoxide dismutase acetylation and regulation of protein structure in breast cancer biology and therapy. Antioxidants 2022, 11, 4. [Google Scholar] [CrossRef] [PubMed]
- Di Ilio, C.; Sacchetta, P.; Angelucci, S.; Zezza, A.; Tenaglia, R.; Aceto, A. Glutathione-peroxidase and glutathione-reductase activities in cancerous and non-cancerous human kidney tissues. Arch. Toxicol. 1995, 91, 19–23. [Google Scholar]
- Gromadzinska, J.; Wasowicz, W.; Andrijewski, M.; Skłodowska, M.; Quispe, O.Z.; Wołkanin, P.; Ołborski, B.; Pluzanska, A. Glutathione and glutathione metabolizing enzymes in tissues and blood of breast cancer patients. Neoplasma 1997, 44, 45–51. [Google Scholar]
- Saydam, N.; Kirb, A.; Demir, O.; Hazan, E.; Oto, O.; Saydam, O.; Guner, G. Determination of glutathione, glutathione reductase, glutathione peroxidase and glutathione S-transferase levels in human lung cancer tissues. Cancer Lett. 1997, 119, 13–19. [Google Scholar] [CrossRef]
- Brigelius-Flohé, R.; Maiorino, M. Glutathione peroxidases. Biochim. Biophys. Acta 2013, 1830, 3289–3303. [Google Scholar] [CrossRef]
- Mates, J.M.; Campos-Sandoval, J.A.; de los Santos-Jiménez, J.; Márquez, J. Glutaminases regulate glutathione and oxidative stress in cancer. Arch. Toxicol. 2020, 94, 2603–2623. [Google Scholar] [CrossRef]
- Zhao, Y.J.; Wang, H.; Zhou, J.D.; Shao, Q.X. Glutathione peroxidase GPX1 and its dichotomous roles in cancer. Cancers 2022, 14, 2560. [Google Scholar] [CrossRef]
- Palma, J.M.; Mateos, R.M.; López-Jaramillo, J.; Rodríguez-Ruiz, M.; González-Gordo, S.; Lechuga-Sancho, A.M.; Corpas, F.J. Plant catalases as NO and H2S targets. Redox Biol. 2020, 34, 101525. [Google Scholar] [CrossRef] [PubMed]
- Glorieux, C.; Dejeans, N.; Sid, B.; Beck, R.; Calderon, P.B.; Verrax, J. Catalase overexpression in mammary cancer cells leads to a less aggressive phenotype and an altered response to chemotherapy. Biochem. Pharmacol. 2011, 82, 1384–1390. [Google Scholar] [CrossRef] [PubMed]
- Glorieux, C.; Calderon, P.B. Catalase, a remarkable enzyme: Targeting the oldest antioxidant enzyme to find a new cancer treatment approach. Biol. Chem. 2017, 398, 1095–1108. [Google Scholar] [CrossRef] [PubMed]
- Galasso, M.; Gambino, S.; Romanelli, M.G.; Donadelli, M.; Scupoli, M.T. Browsing the oldest antioxidant enzyme: Catalase and its multiple regulation in cancer. Free Radic. Biol. Med. 2021, 172, 264–272. [Google Scholar] [CrossRef] [PubMed]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cell Lines | IC50 (μM Capsaicin) | Capsaicin in the Assay (μg/well) |
|---|---|---|
| A549 | 40.70 | 1.243 |
| A2058 | 53.00 | 1.618 |
| Hep-G2 | 28.00 | 0.855 |
| HT-29 | 51.50 | 1.573 |
| MCF-7 | 33.15 | 1.012 |
| MIA PaCa-2 | 61.25 | 1.870 |
| PC-3 | 72.20 | 2.199 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rodríguez-Ruiz, M.; Ramos, M.C.; Campos, M.J.; Díaz-Sánchez, I.; Cautain, B.; Mackenzie, T.A.; Vicente, F.; Corpas, F.J.; Palma, J.M. Pepper Fruit Extracts Show Anti-Proliferative Activity against Tumor Cells Altering Their NADPH-Generating Dehydrogenase and Catalase Profiles. Antioxidants 2023, 12, 1461. https://doi.org/10.3390/antiox12071461
Rodríguez-Ruiz M, Ramos MC, Campos MJ, Díaz-Sánchez I, Cautain B, Mackenzie TA, Vicente F, Corpas FJ, Palma JM. Pepper Fruit Extracts Show Anti-Proliferative Activity against Tumor Cells Altering Their NADPH-Generating Dehydrogenase and Catalase Profiles. Antioxidants. 2023; 12(7):1461. https://doi.org/10.3390/antiox12071461
Chicago/Turabian StyleRodríguez-Ruiz, Marta, María C. Ramos, María J. Campos, Inmaculada Díaz-Sánchez, Bastien Cautain, Thomas A. Mackenzie, Francisca Vicente, Francisco J. Corpas, and José M. Palma. 2023. "Pepper Fruit Extracts Show Anti-Proliferative Activity against Tumor Cells Altering Their NADPH-Generating Dehydrogenase and Catalase Profiles" Antioxidants 12, no. 7: 1461. https://doi.org/10.3390/antiox12071461
APA StyleRodríguez-Ruiz, M., Ramos, M. C., Campos, M. J., Díaz-Sánchez, I., Cautain, B., Mackenzie, T. A., Vicente, F., Corpas, F. J., & Palma, J. M. (2023). Pepper Fruit Extracts Show Anti-Proliferative Activity against Tumor Cells Altering Their NADPH-Generating Dehydrogenase and Catalase Profiles. Antioxidants, 12(7), 1461. https://doi.org/10.3390/antiox12071461