


The Interplay of Lifestyle and Adipokines in the Non-Obese Stroke-Prone Spontaneously Hypertensive Rats


Abstract
1. Introduction
2. Materials and Methods
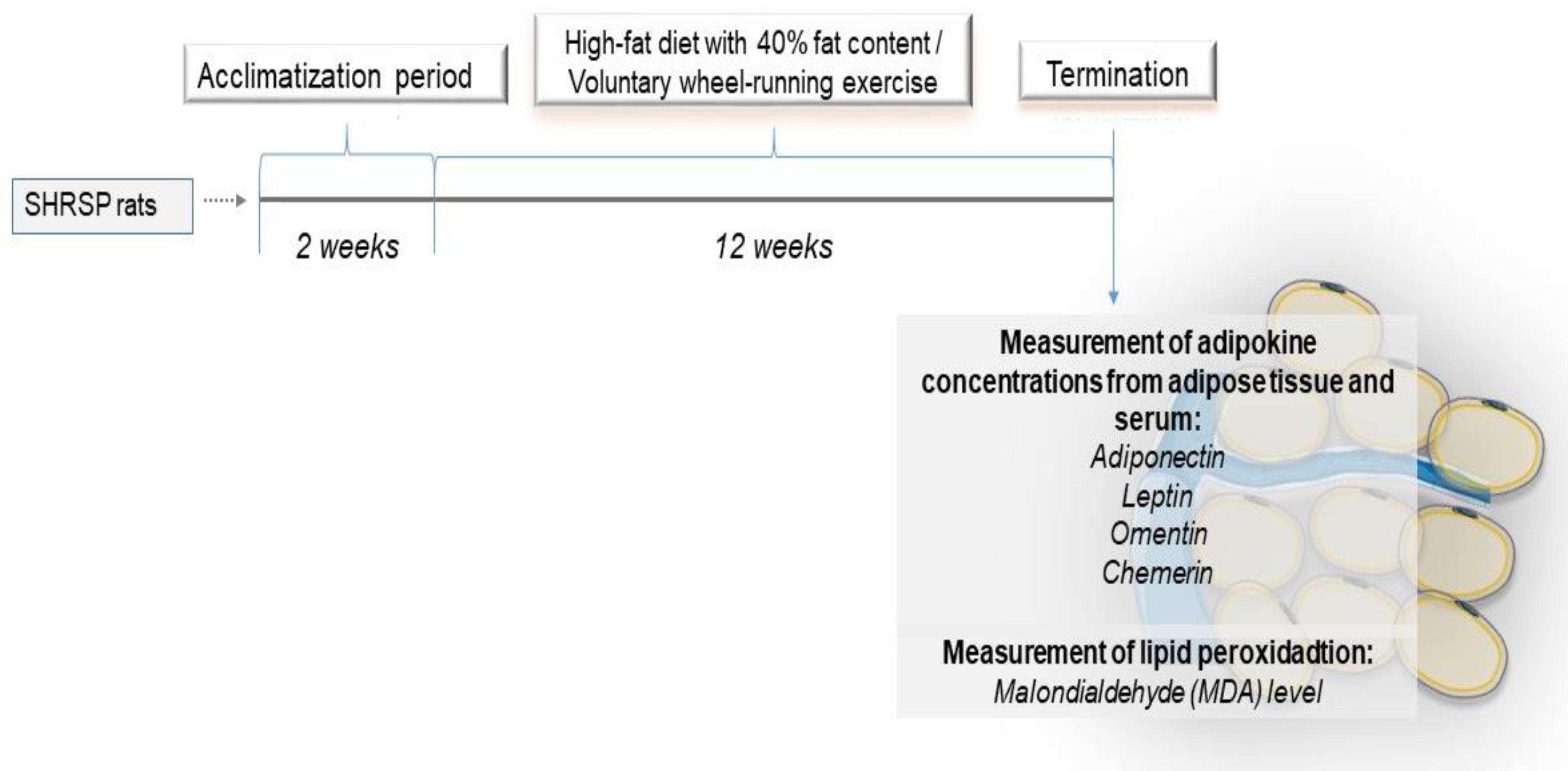
2.1. Experimental Protocol and Sample Collection
2.2. Ethical Approval
2.3. Determination of Adipokine Profile with ELISA
2.4. Determination of Lipid Peroxidation (MDA)
2.5. Statistical Analyses
3. Results
3.1. Body Weight Changes
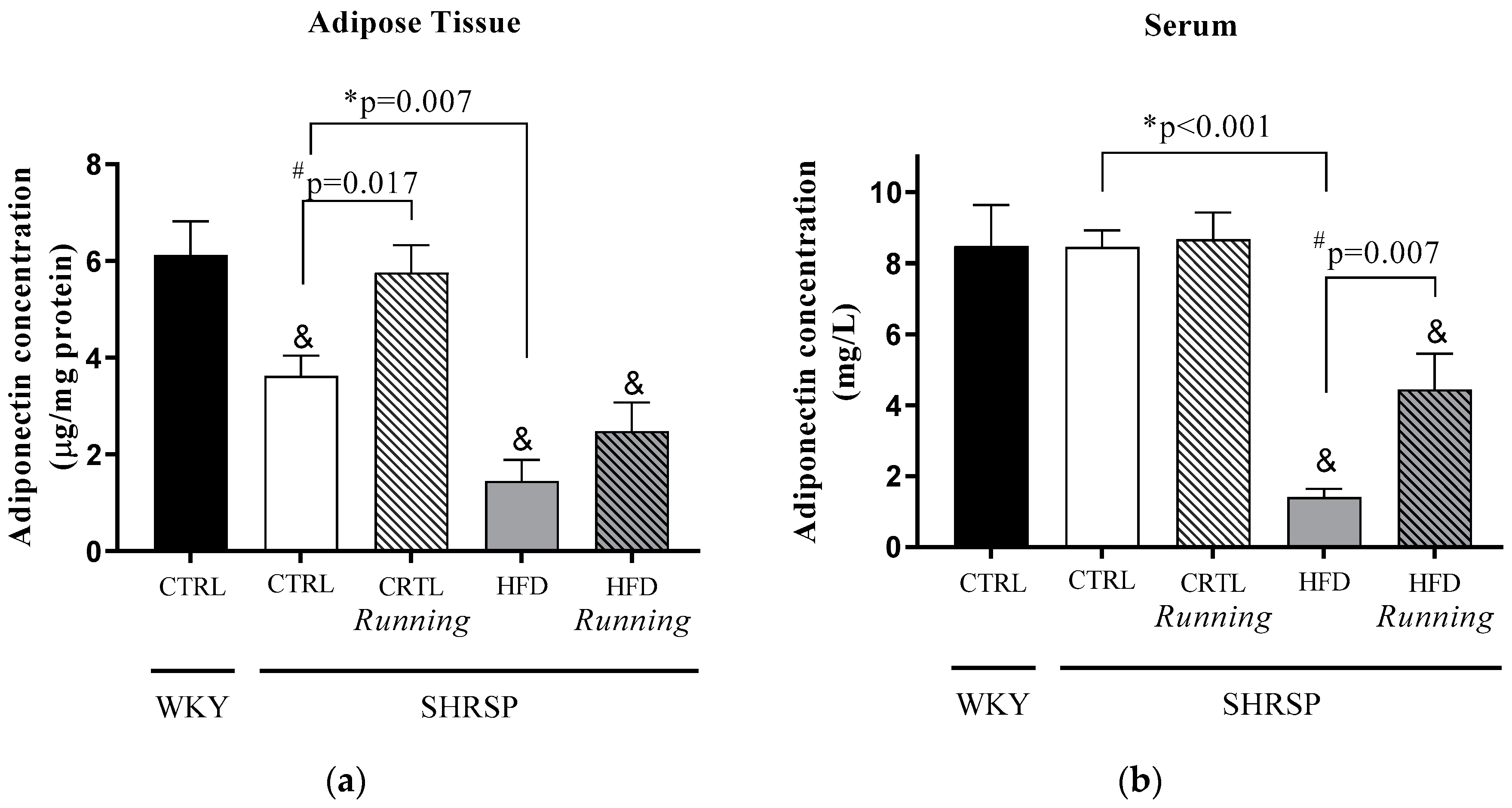
3.2. Adiponectin Concentrations

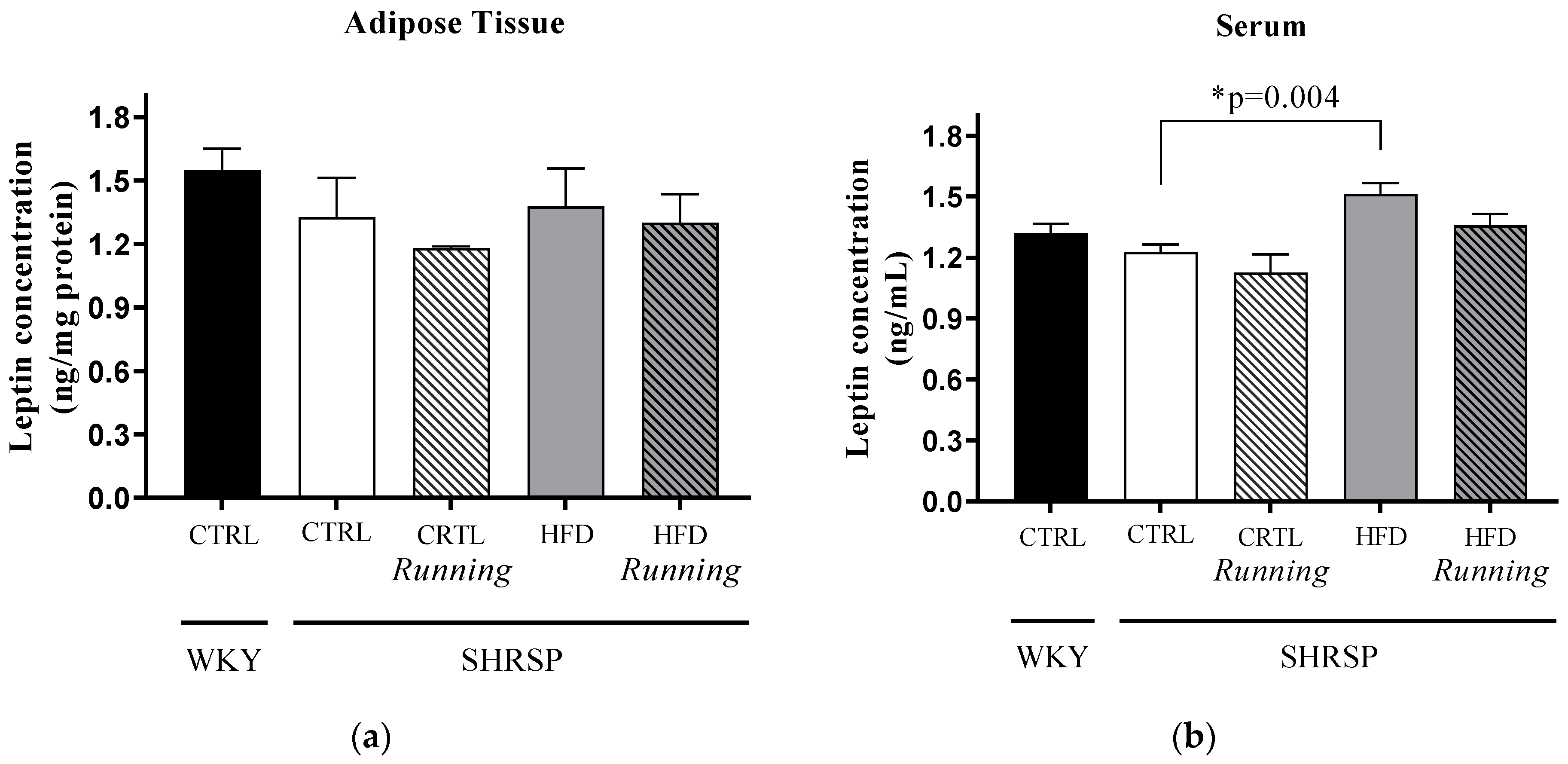
3.3. Leptin Concentrations
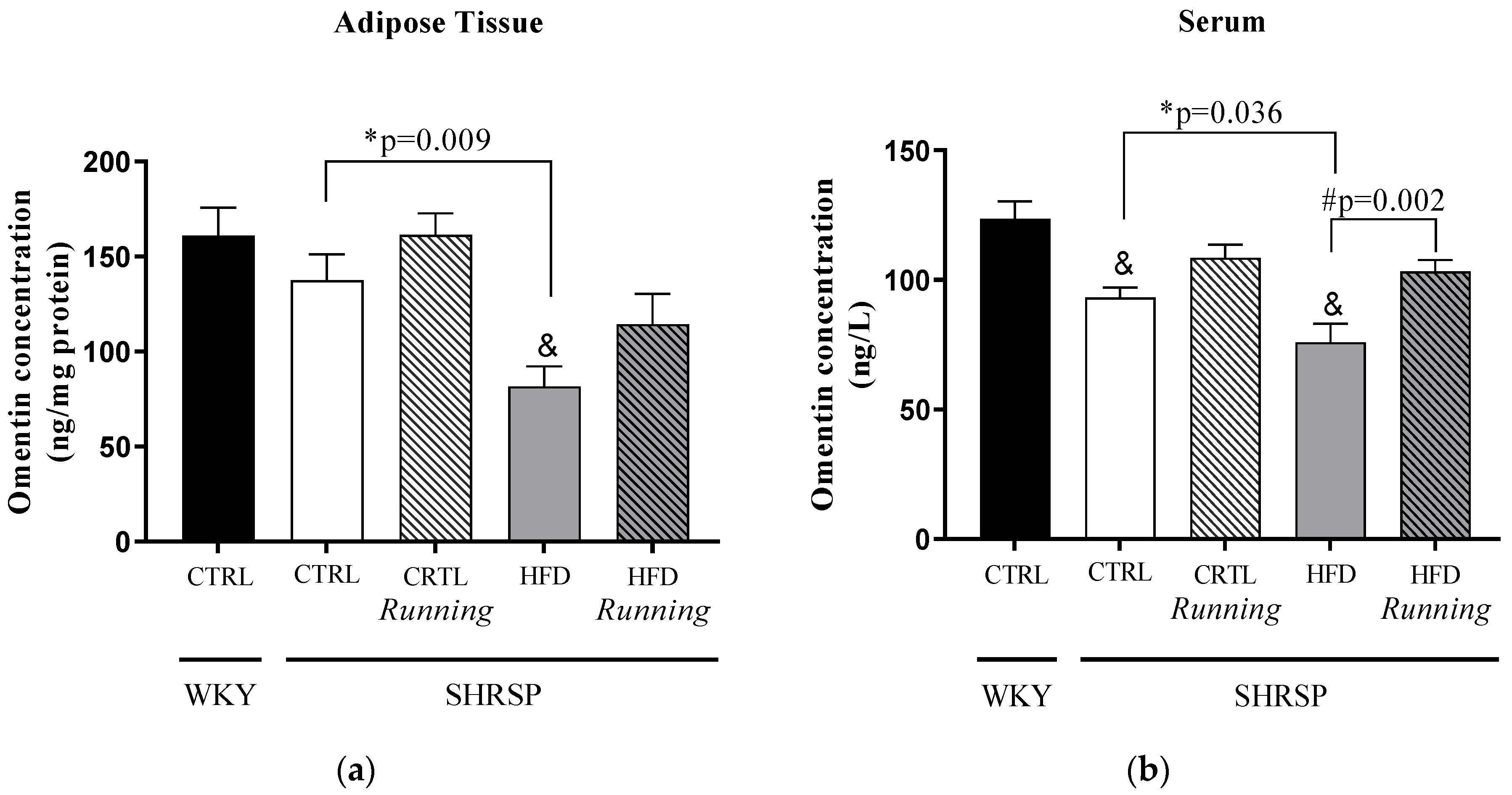
3.4. Omentin Concentrations
3.5. Chemerin Concentrations
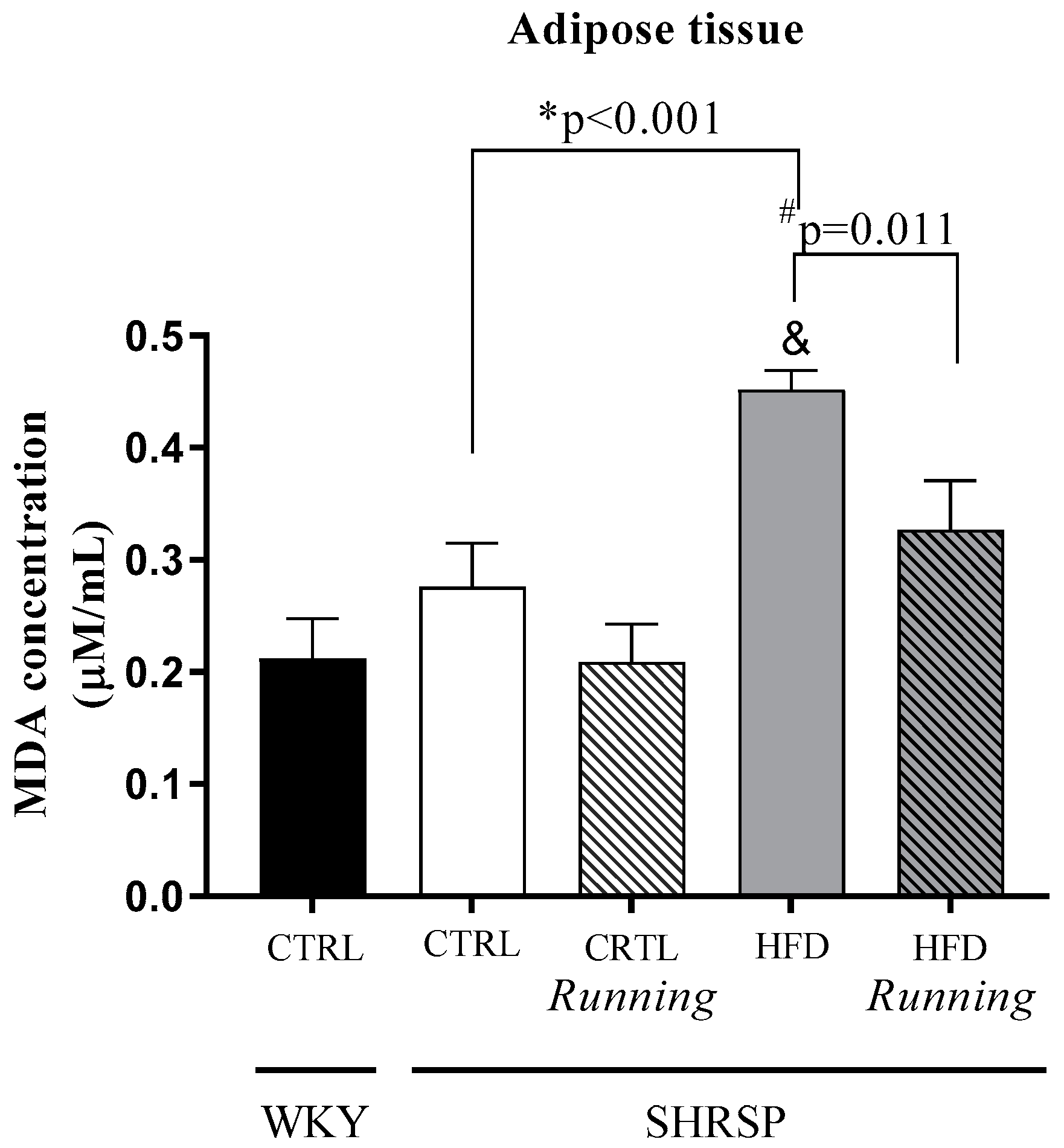
3.6. Determination of Lipid Peroxidation
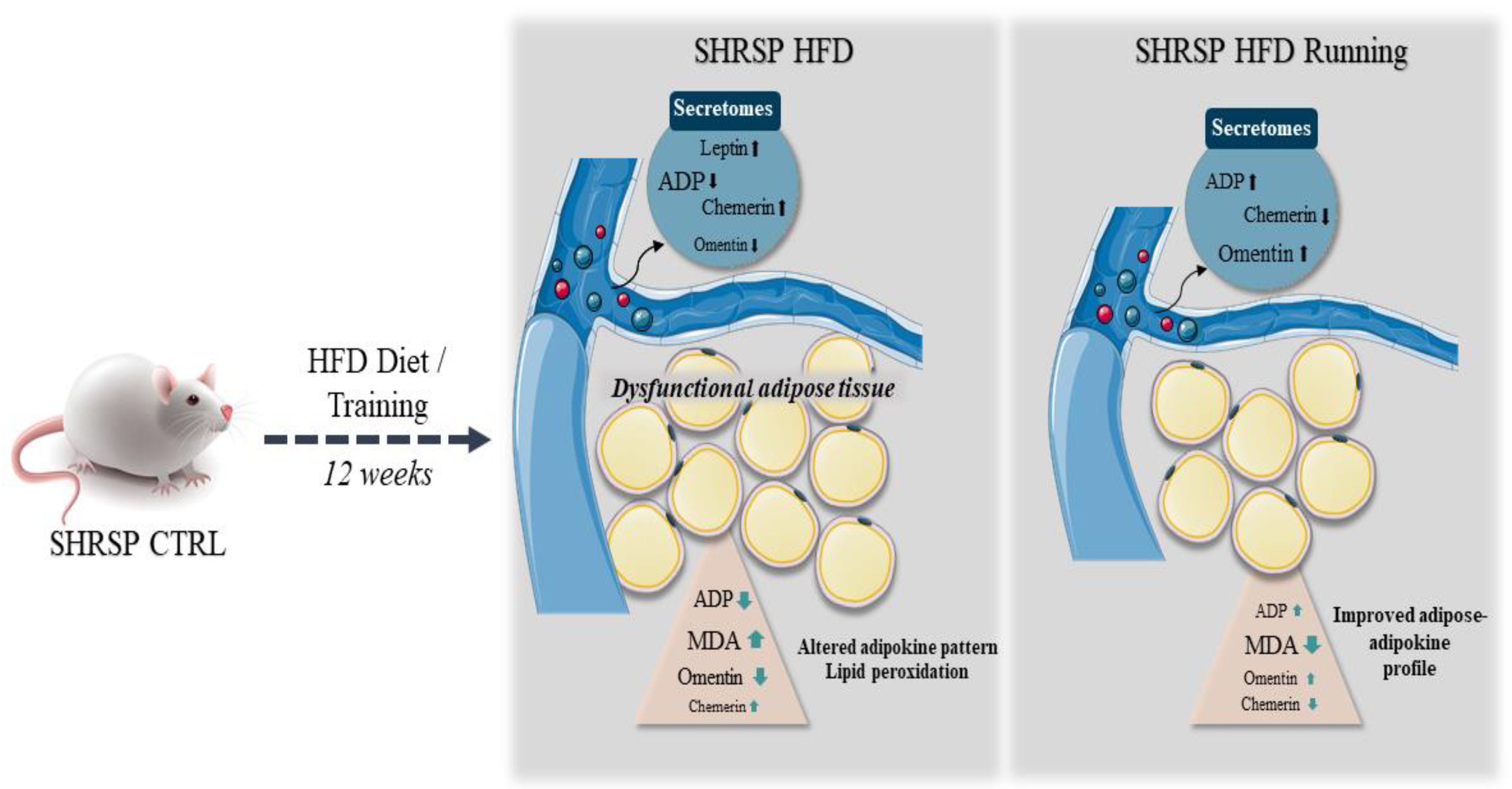
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Rutkowski, J.M.; Stern, J.H.; Scherer, P.E. The cell biology of fat expansion. J. Cell Biol. 2015, 208, 501–512. [Google Scholar] [CrossRef] [PubMed]
- Longo, M.; Zatterale, F.; Naderi, J.; Parrillo, L.; Formisano, P.; Raciti, G.A.; Beguinot, F.; Miele, C. Adipose Tissue Dysfunction as Determinant of Obesity-Associated Metabolic Complications. Int. J. Mol. Sci. 2019, 20, 2358. [Google Scholar] [CrossRef]
- Frühbeck, G.; Catalán, V.; Rodríguez, A.; Ramírez, B.; Becerril, S.; Salvador, J.; Portincasa, P.; Colina, I.; Gómez-Ambrosi, J. Involvement of the leptin-adiponectin axis in inflammation and oxidative stress in the metabolic syndrome. Sci. Rep. 2017, 7, 6619. [Google Scholar] [CrossRef] [PubMed]
- Scotece, M.; Conde, J.; López, V.; Lago, F.; Pino, J.; Gómez-Reino, J.J.; Gualillo, O. Adiponectin and leptin: New targets in inflammation. Basic Clin. Pharmacol. Toxicol. 2014, 114, 97–102. [Google Scholar] [CrossRef] [PubMed]
- Jiang, S.; Liu, H.; Li, C. Dietary Regulation of Oxidative Stress in Chronic Metabolic Diseases. Foods 2021, 10, 1854. [Google Scholar] [CrossRef]
- Zhou, Y.; Li, H.; Xia, N. The Interplay Between Adipose Tissue and Vasculature: Role of Oxidative Stress in Obesity. Front. Cardiovasc. Med. 2021, 8, 650214. [Google Scholar] [CrossRef]
- Yamori, Y.; Sagara, M.; Mori, H.; Mori, M. Stroke-Prone SHR as Experimental Models for Cardiovascular Disease Risk Reduction in Humans. Biomedicines 2022, 10, 2974. [Google Scholar] [CrossRef]
- Pagan, L.U.; Damatto, R.L.; Gomes, M.J.; Lima, A.R.R.; Cezar, M.D.M.; Damatto, F.C.; Reyes, D.R.A.; Caldonazo, T.M.M.; Polegato, B.F.; Okoshi, M.P.; et al. Low-intensity aerobic exercise improves cardiac remodelling of adult spontaneously hypertensive rats. J. Cell. Mol. Med. 2019, 23, 6504–6507. [Google Scholar] [CrossRef]
- Szabó, R.; Börzsei, D.; Hoffmann, A.; Lesi, Z.N.; Gesztelyi, R.; Juhász, B.; Szebeni, G.J.; Osman, J.; Sebestyén, J.; Nagy, A.; et al. Lifestyle-Induced Redox-Sensitive Alterations: Cross-Talk among the RAAS, Antioxidant/Inflammatory Status, and Hypertension. Oxidative Med. Cell. Longev. 2021, 2021, 3080863. [Google Scholar] [CrossRef]
- Smekal, A.; Vaclavik, J. Adipokines and cardiovascular disease: A comprehensive review. Biomed. Pap. Med. Fac. Palacky Univ. Olomouc 2017, 161, 31–40. [Google Scholar] [CrossRef]
- Yokoi, N.; Hoshino, M.; Hidaka, S.; Yoshida, E.; Beppu, M.; Hoshikawa, R.; Sudo, K.; Kawada, A.; Takagi, S.; Seino, S. A Novel Rat Model of Type 2 Diabetes: The Zucker Fatty Diabetes Mellitus ZFDM Rat. J. Diabetes Res. 2013, 2013, 103731. [Google Scholar] [CrossRef] [PubMed]
- Gao, M.; Ma, Y.; Liu, D. High-fat diet-induced adiposity, adipose inflammation, hepatic steatosis and hyperinsulinemia in outbred CD-1 mice. PLoS ONE 2015, 10, e0119784. [Google Scholar] [CrossRef] [PubMed]
- Würfel, M.; Breitfeld, J.; Gebhard, C.; Scholz, M.; Baber, R.; Riedel-Heller, S.G.; Blüher, M.; Stumvoll, M.; Kovacs, P.; Tönjes, A. Interplay between adipose tissue secreted proteins, eating behavior and obesity. Eur. J. Nutr. 2022, 61, 885–899. [Google Scholar] [CrossRef] [PubMed]
- Zhao, S.; Kusminski, C.M.; Scherer, P.E. Adiponectin, Leptin and Cardiovascular Disorders. Circ. Res. 2021, 128, 136–149. [Google Scholar] [CrossRef]
- Wang, Q.A.; Tao, C.; Gupta, R.K.; Scherer, P.E. Tracking adipogenesis during white adipose tissue development, expansion and regeneration. Nat. Med. 2013, 19, 1338–1344. [Google Scholar] [CrossRef]
- Martins, S.V.; Lopes, P.A.; Alfaia, C.M.; Rodrigues, P.O.; Alves, S.P.; Pinto, R.M.A.; Castro, M.F.; Bessa, R.J.B.; Prates, J.A.M. Serum adipokine profile and fatty acid composition of adipose tissues are affected by conjugated linoleic acid and saturated fat diets in obese Zucker rats. Br. J. Nutr. 2010, 103, 869–878. [Google Scholar] [CrossRef]
- Nabika, T.; Ohara, H.; Kato, N.; Isomura, M. The stroke-prone spontaneously hypertensive rat: Still a useful model for post-GWAS genetic studies? Hypertens. Res. 2012, 35, 477–484. [Google Scholar] [CrossRef]
- Jiang, S.Z.; Lu, W.; Zong, X.F.; Ruan, H.Y.; Liu, Y. Obesity and hypertension. Exp. Ther. Med. 2016, 12, 2395–2399. [Google Scholar] [CrossRef]
- El Meouchy, P.; Wahoud, M.; Allam, S.; Chedid, R.; Karam, W.; Karam, S. Hypertension Related to Obesity: Pathogenesis, Characteristics and Factors for Control. Int. J. Mol. Sci. 2022, 23, 12305. [Google Scholar] [CrossRef]
- Schütten, M.T.J.; Houben, A.J.H.M.; de Leeuw, P.W.; Stehouwer, C.D.A. The Link Between Adipose Tissue Renin-Angiotensin-Aldosterone System Signaling and Obesity-Associated Hypertension. Physiology 2017, 32, 197–209. [Google Scholar] [CrossRef]
- Park, H.-K.; Ahima, R.S. Physiology of leptin: Energy homeostasis, neuroendocrine function and metabolism. Metabolism 2015, 64, 24–34. [Google Scholar] [CrossRef] [PubMed]
- Borghi, F.; Silva, C.; da Silva, P.C.; Ferrucci, D.L.; Morais, C.L.; Conceição-Vertamatti, A.G.; Carvalho, H.F.; Fonseca, M.D.C.; Vieira, A.S.; Grassi-Kassisse, D.M. The influence of hypertensive environment on adipose tissue remodeling measured by fluorescence lifetime imaging in spontaneously hypertensive rats. Mol. Cell. Endocrinol. 2020, 506, 110758. [Google Scholar] [CrossRef] [PubMed]
- Cao, J.; Sodhi, K.; Puri, N.; Monu, S.R.; Rezzani, R.; Abraham, N.G. High fat diet enhances cardiac abnormalities in SHR rats: Protective role of heme oxygenase-adiponectin axis. Diabetol. Metab. Syndr. 2011, 3, 37. [Google Scholar] [CrossRef] [PubMed]
- Frühbeck, G.; Catalán, V.; Rodríguez, A.; Gómez-Ambrosi, J. Adiponectin-leptin ratio: A promising index to estimate adipose tissue dysfunction. Relation with obesity-associated cardiometabolic risk. Adipocyte 2017, 7, 57–62. [Google Scholar] [CrossRef] [PubMed]
- Doumatey, A.P.; Bentley, A.R.; Zhou, J.; Huang, H.; Adeyemo, A.; Rotimi, C.N. Paradoxical Hyperadiponectinemia is Associated With the Metabolically Healthy Obese (MHO) Phenotype in African Americans. J. Endocrinol. Metab. 2012, 2, 51–65. [Google Scholar] [CrossRef]
- Aprahamian, T.R.; Sam, F. Adiponectin in cardiovascular inflammation and obesity. Int. J. Inflamm. 2011, 2011, 376909. [Google Scholar] [CrossRef][Green Version]
- Xu, F.; Li, F.-X.; Lin, X.; Zhong, J.-Y.; Wu, F.; Shan, S.-K.; Tan, C.-M.; Yuan, L.-Q.; Liao, X.-B. Adipose tissue-derived omentin-1 attenuates arterial calcification via AMPK/Akt signaling pathway. Aging 2019, 11, 8760–8776. [Google Scholar] [CrossRef]
- Nosrati-Oskouie, M.; Asghari, G.; Yuzbashian, E.; Aghili-Moghaddam, N.S.; Zarkesh, M.; Safarian, M.; Mirmiran, P. Does Dietary Intake Impact Omentin Gene Expression and Plasma Concentration? A Systematic Review. Lifestyle Genom. 2021, 14, 49–61. [Google Scholar] [CrossRef]
- Ferland, D.J.; Mullick, E.A.; Watts, S.W. Chemerin as a Driver of Hypertension: A Consideration. Am. J. Hypertens. 2020, 33, 975–986. [Google Scholar] [CrossRef]
- Hojna, S.; Jordan, M.D.; Kollias, H.; Pausova, Z. High-fat diet induces emergence of brown-like adipocytes in white adipose tissue of spontaneously hypertensive rats. Hypertens. Res. 2012, 35, 279–286. [Google Scholar] [CrossRef]
- Poole, D.C.; Copp, S.W.; Colburn, T.D.; Craig, J.C.; Allen, D.L.; Sturek, M.; O’leary, D.S.; Zucker, I.H.; Musch, T.I. Guidelines for animal exercise and training protocols for cardiovascular studies. Am. J. Physiol. Circ. Physiol. 2020, 318, H1100–H1138. [Google Scholar] [CrossRef] [PubMed]
- Yan, P.; Liu, D.; Long, M.; Ren, Y.; Pang, J.; Li, R. Changes of serum omentin levels and relationship between omentin and adiponectin concentrations in type 2 diabetes mellitus. Exp. Clin. Endocrinol. Diabetes 2011, 119, 257–263. [Google Scholar] [CrossRef] [PubMed]
- Feng, W.-H.; Yuan, X.-W.; Tong, G.-Y.; Wang, W.-M.; Hu, Y.; Shen, S.-M.; Li, P.; Bi, Y.; Hu, J.; Shao, L.-L.; et al. Correlated increase of omentin-1 and adiponectin by exenatide, avandamet and dietary change in diet-induced obese rats. Folia Biol. 2013, 59, 217–224. [Google Scholar]
- De Castro, C.A.; da Silva, K.A.; Rocha, M.C.; Sene-Fiorese, M.; Nonaka, K.O.; Malavazi, I.; Anibal, F.d.F.; Duarte, A.C.G.d.O. Exercise and Omentin: Their Role in the Crosstalk Between Muscle and Adipose Tissues in Type 2 Diabetes Mellitus Rat Models. Front. Physiol. 2018, 9, 1881. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| WKY CTRL | SHRSP CTRL | SHRSP CTRL + R | SHRSP HFD | SHRSP HFD + R | ||
|---|---|---|---|---|---|---|
| (a) | Body weight at the start of the experiment | 157.25 ± 2.66 | 146.17 ± 4.5 | 156.50 ± 5.15 | 143.25 ± 5.94 | 192.25 ± 6.39 |
| Body weight at the end of the experiment | 294.5 ± 1.93 | 258.83 ± 2.33 | 294.0 ± 2.98 | 303.63 ± 2.62 | 295.38 ± 4.14 | |
| (b) | Changes in the body weights | 137.500 ± 44.863 | 112.667 ± 44.088 | 160.375 ± 5.322 # | 103.125 ± 3.461 | 137.250 ± 4.109 # |
| Source of Variation | p Value | |
|---|---|---|
| (a) | (b) | |
| Interaction between Running × Diet | 0.305 | 0.066 |
| Source of Variation | p Value | |
|---|---|---|
| (a) | (b) | |
| Interaction between Running × Diet | 0.836 | 0.680 |
| Source of Variation | p Value | |
|---|---|---|
| (a) | (b) | |
| Interaction between Running × Diet | 0.751 | 0.290 |
| Source of Variation | p Value | |
|---|---|---|
| (a) | (b) | |
| Interaction between Running × Diet | 0.251 | 0.127 |
| Source of Variation | p Value |
|---|---|
| Interaction between Running × Diet | 0.390 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Szabó, R.; Börzsei, D.; Hoffmann, A.; Kiss, V.; Nagy, A.; Török, S.; Veszelka, M.; Almási, N.; Varga, C. The Interplay of Lifestyle and Adipokines in the Non-Obese Stroke-Prone Spontaneously Hypertensive Rats. Antioxidants 2023, 12, 1450. https://doi.org/10.3390/antiox12071450
Szabó R, Börzsei D, Hoffmann A, Kiss V, Nagy A, Török S, Veszelka M, Almási N, Varga C. The Interplay of Lifestyle and Adipokines in the Non-Obese Stroke-Prone Spontaneously Hypertensive Rats. Antioxidants. 2023; 12(7):1450. https://doi.org/10.3390/antiox12071450
Chicago/Turabian StyleSzabó, Renáta, Denise Börzsei, Alexandra Hoffmann, Viktória Kiss, András Nagy, Szilvia Török, Médea Veszelka, Nikoletta Almási, and Csaba Varga. 2023. "The Interplay of Lifestyle and Adipokines in the Non-Obese Stroke-Prone Spontaneously Hypertensive Rats" Antioxidants 12, no. 7: 1450. https://doi.org/10.3390/antiox12071450
APA StyleSzabó, R., Börzsei, D., Hoffmann, A., Kiss, V., Nagy, A., Török, S., Veszelka, M., Almási, N., & Varga, C. (2023). The Interplay of Lifestyle and Adipokines in the Non-Obese Stroke-Prone Spontaneously Hypertensive Rats. Antioxidants, 12(7), 1450. https://doi.org/10.3390/antiox12071450