
Natural Compounds and Glutathione: Beyond Mere Antioxidants

 ,
,  , , and
, , and 
Abstract
1. Introduction
2. Materials and Methods
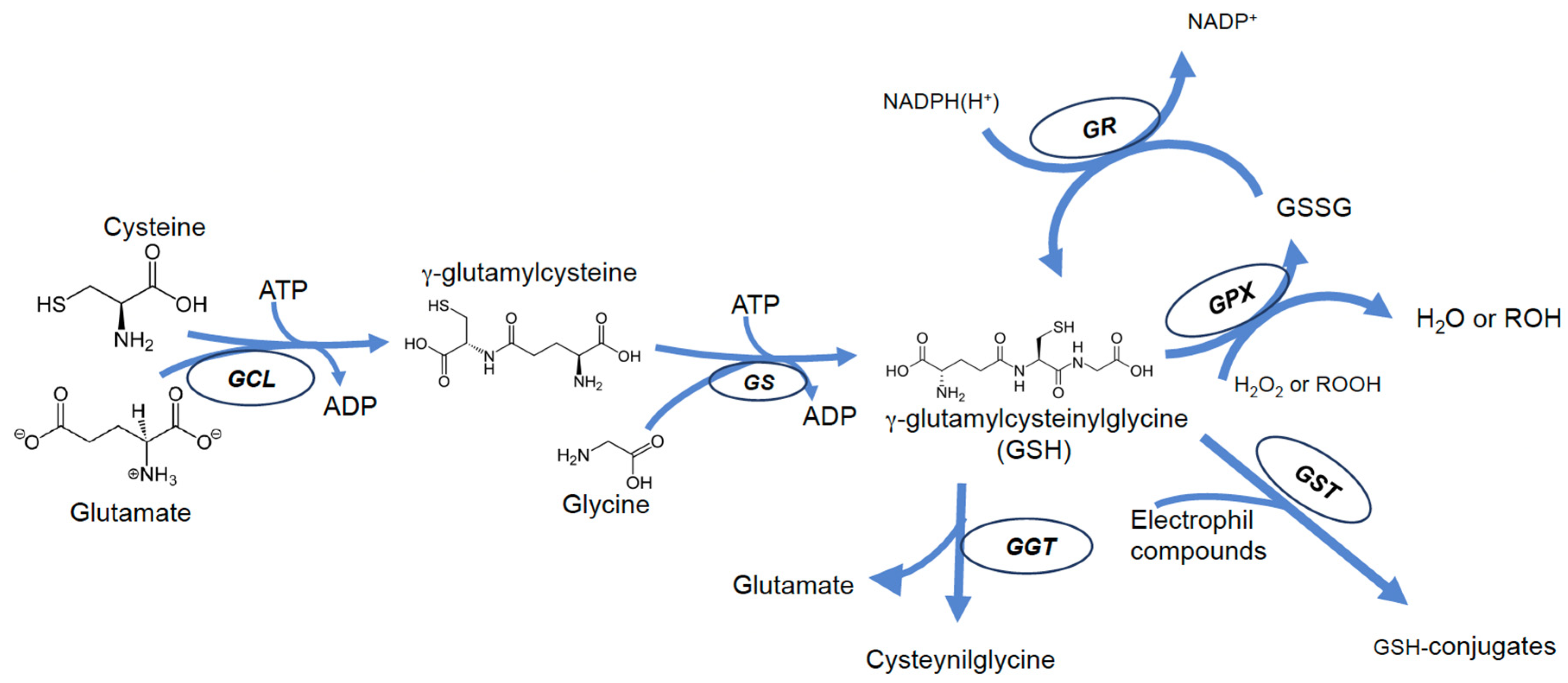
3. Glutathione Biosynthesis and Functions
4. GSH and Tumor Cells
5. Natural Compounds and GSH
5.1. Polyphenols and GSH
5.2. Terpenoids and GSH
5.3. Glucosinolates and GSH
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Meister, A. On the Discovery of Glutathione. Trends Biochem. Sci. 1988, 13, 185–188. [Google Scholar] [CrossRef] [PubMed]
- Hristov, B.D. The Role of Glutathione Metabolism in Chronic Illness Development and Its Potential Use as a Novel Therapeutic Target. Cureus 2022, 14, e29696. [Google Scholar] [CrossRef] [PubMed]
- Forman, H.J.; Zhang, H.; Rinna, A. Glutathione: Overview of Its Protective Roles, Measurement, and Biosynthesis. Mol. Asp. Med. 2009, 30, 1–12. [Google Scholar] [CrossRef]
- Averill-Bates, D.A. The Antioxidant Glutathione. Vitam. Horm. 2023, 121, 109–141. [Google Scholar] [CrossRef]
- Diaz-Vivancos, P.; De Simone, A.; Kiddle, G.; Foyer, C.H. Glutathione—Linking Cell Proliferation to Oxidative Stress. Free Radic. Biol. Med. 2015, 89, 1154–1164. [Google Scholar] [CrossRef] [PubMed]
- Tan, B.L.; Norhaizan, M.E.; Liew, W.P.P.; Rahman, H.S. Antioxidant and Oxidative Stress: A Mutual Interplay in Age-Related Diseases. Front. Pharmacol. 2018, 9, 1162. [Google Scholar] [CrossRef]
- Cheng, X.; Xu, H.D.; Ran, H.H.; Liang, G.; Wu, F.G. Glutathione-Depleting Nanomedicines for Synergistic Cancer Therapy. ACS Nano 2021, 15, 8039–8068. [Google Scholar] [CrossRef]
- Pallardó, F.V.; Markovic, J.; García, J.L.; Viña, J. Role of Nuclear Glutathione as a Key Regulator of Cell Proliferation. Mol. Asp. Med. 2009, 30, 77–85. [Google Scholar] [CrossRef]
- Bertrand, R.L. Iron Accumulation, Glutathione Depletion, and Lipid Peroxidation Must Occur Simultaneously during Ferroptosis and Are Mutually Amplifying Events. Med. Hypotheses 2017, 101, 69–74. [Google Scholar] [CrossRef]
- Traverso, N.; Ricciarelli, R.; Nitti, M.; Marengo, B.; Furfaro, A.L.; Pronzato, M.A.; Marinari, U.M.; Domenicotti, C. Role of Glutathione in Cancer Progression and Chemoresistance. Oxid. Med. Cell. Longev. 2023, 2013, 972913. [Google Scholar] [CrossRef]
- Jaganjac, M.; Milkovic, L.; Sunjic, S.B.; Zarkovic, N. The NRF2, Thioredoxin, and Glutathione System in Tumorigenesis and Anticancer Therapies. Antioxidants 2020, 9, 1151. [Google Scholar] [CrossRef] [PubMed]
- Teskey, G.; Cao, R.; Islamoglu, H.; Medina, A.; Prasad, C.; Prasad, R.; Sathananthan, A.; Fraix, M.; Subbian, S.; Zhong, L.; et al. The Synergistic Effects of the Glutathione Precursor, NAC and First-Line Antibiotics in the Granulomatous Response against Mycobacterium Tuberculosis. Front. Immunol. 2018, 9, 2069. [Google Scholar] [CrossRef] [PubMed]
- Scharf, G.; Prustomersky, S.; Knasmüller, S.; Schulte-Hermann, R.; Huber, W.W. Enhancement of Glutathione and γ-Glutamylcysteine Synthetase, the Rate Limiting Enzyme of Glutathione Synthesis, by Chemoprotective Plant-Derived Food and Beverage Components in the Human Hepatoma Cell Line HepG2. Nutr. Cancer 2003, 45, 74–83. [Google Scholar] [CrossRef]
- Luceri, C.; Caderni, G.; Sanna, A.; Dolara, P. Nutrition and Cancer-Research Communication Red Wine and Black Tea Polyphenols Modulate the Expression of Cycloxygenase-2, Inducible Nitric Oxide Synthase and Glutathione-Related Enzymes in Azoxymethane-Induced F344 Rat Colon Tumors. J. Nutr. 2002, 132, 1376–1379. [Google Scholar] [CrossRef]
- Havsteen, B.H. The Biochemistry and Medical Significance of the Flavonoids. Pharmacol. Therapeut. 2002, 96, 67–202. [Google Scholar] [CrossRef]
- Hodnick, W.F.; Ahmad, S.; Pardini, R.S. Induction of oxidative stress by redox active flavonoids in vitro production of reactive oxygen species. In Flavonoids in the Living System; Manthey, J.A., Buslig, B.S., Eds.; Plenum Press: New York, NY, USA, 1998; p. 131. [Google Scholar]
- van Zanden, J.J.; Geraets, L.; Wortelboer, H.M.; van Bladeren, P.J.; Rietjens, I.M.; Cnubben, N.H. Structural requirements for the flavonoid-mediated modulation of glutathione S-transferase P1-1 and GS-X pump activity in MCF7 breast cancer cells. Biochem. Pharmacol. 2004, 67, 1607–1617. [Google Scholar] [CrossRef] [PubMed]
- Engwa, G.A. Free Radicals and the Role of Plant Phytochemicals as Antioxidants Against Oxidative Stress-Related Diseases. In Phytochemicals—Source of Antioxidants and Role in Disease Prevention; InTechOpen: London, UK, 2018. [Google Scholar]
- Zhou, D.-D.; Luo, M.; Huang, S.-Y.; Saimaiti, A.; Shang, A.; Gan, R.-Y.; Li, H.-B. Effects and Mechanisms of Resveratrol on Aging and Age-Related Diseases. Oxid. Med. Cell. Longev. 2021, 2021, 9932218. [Google Scholar] [CrossRef] [PubMed]
- Valenti, G.E.; Tasso, B.; Traverso, N.; Domenicotti, C.; Marengo, B. Glutathione in Cancer Progression and Chemoresistance: An Update. Redox Exp. Med. 2023, 2023, e220023. [Google Scholar] [CrossRef]
- Lushchak, V.I. Glutathione Homeostasis and Functions: Potential Targets for Medical Interventions. J. Amino Acids 2012, 2012, 736837. [Google Scholar] [CrossRef]
- Lu, S.C. Glutathione Synthesis. Biochim. Biophys. Acta Gen. Subj. 2013, 1830, 3143–3153. [Google Scholar] [CrossRef]
- Meister, A.; Anderson, M.E. Glutathione. Annu. Rev. Biochem. 1983, 52, 711–760. [Google Scholar] [CrossRef] [PubMed]
- Halliwell, B.; Gutteridge, J.M.C. Antioxidant Defences Synthesized in Vivo. In Free Radicals in Biology and Medicine; Clarendon Press, Ed.; Oxford University Press: Oxford, UK, 2015; pp. 77–152. [Google Scholar]
- Oda, T.; Sadakata, N.; Komatsu, N.; Muramatsu, T. Specific Efflux of Glutathione from the Basolateral Membrance Domain in Polarized MDCK Cells during Ricin-Induced Apoptosis. J. Biochem. 1999, 126, 715–721. [Google Scholar] [CrossRef] [PubMed]
- Mattson, D.M.; Ahmad, I.M.; Dayal, D.; Parsons, A.D.; Aykin-Burns, N.; Li, L.; Orcutt, K.P.; Spitz, D.R.; Dornfeld, K.J.; Simons, A.L. Cisplatin Combined with Zidovudine Enhances Cytotoxicity and Oxidative Stress in Human Head and Neck Cancer Cells via a Thiol-Dependent Mechanism. Free Radic. Biol. Med. 2009, 46, 232–237. [Google Scholar] [CrossRef]
- Franco, R.; Cidlowski, J.A. Glutathione Efflux and Cell Death. Antioxid. Redox. Signal. 2012, 17, 1694–1713. [Google Scholar] [CrossRef]
- Franco, R.; Cidlowski, J.A. Apoptosis and Glutathione: Beyond an Antioxidant. Cell. Death Differ. 2009, 16, 1303–1314. [Google Scholar] [CrossRef]
- Trompier, D.; Chang, X.-B.; Barattin, R.; d’Hardemare, A.d.M.; Di Pietro, A.; Baubichon-Cortay, H. Verapamil and Its Derivative Trigger Apoptosis through Glutathione Extrusion by Multidrug Resistance Protein MRP1. Cancer Res. 2004, 64, 4950–4956. [Google Scholar] [CrossRef] [PubMed]
- Guha, P.; Dey, A.; Sen, R.; Chatterjee, M.; Chattopadhyay, S.; Bandyopadhyay, S.K. Intracellular GSH Depletion Triggered Mitochondrial Bax Translocation to Accomplish Resveratrol-Induced Apoptosis in the U937 Cell Line. J. Pharmacol. Exp. Ther. 2011, 336, 206–214. [Google Scholar] [CrossRef]
- Lim, C.H.; Dedon, P.C.; Deen, W.M. Kinetic Analysis of Intracellular Concentrations of Reactive Nitrogen Species. Chem. Res. Toxicol. 2008, 21, 2134–2147. [Google Scholar] [CrossRef]
- Sagone, A.J.; Husney, R.; O’Dorisio, M.; Metz, E. Mechanisms for the Oxidation of Reduced Gluthathione by Stimulated Granulocytes. Blood 1984, 63, 96–104. [Google Scholar] [CrossRef]
- Abedinzadeh, Z.; Gardes-Albert, M.; Ferradini, C. Kinetic Study of the Oxidation Mechanism of Glutathione by Hydrogen Peroxide in Neutral Aqueous Medium. Can. J. Chem. 1989, 67, 1247–1255. [Google Scholar] [CrossRef]
- Zinatullina, K.M.; Kasaikina, O.T.; Kuz’min, V.A.; Khrameeva, N.P. Interaction of Glutathione with Hydrogen Peroxide: A Kinetic Model. Kinet. Catal. 2019, 60, 266–272. [Google Scholar] [CrossRef]
- Winterbourn, C.C. Radical Scavenging by Thiols and the Fate of Thiyl Radicals. In Oxidative Stress and Redox Regulation; Springer Netherlands: Dordrecht, The Netherlands, 2013; pp. 43–58. [Google Scholar]
- Kalyanaraman, B.; Karoui, H.; Jit Singh, R.; Felix, C.C. Detection of Thiyl Radical Adducts Formed during Hydroxyl Radical- and Peroxynitrite-Mediated Oxidation of Thiols--A High Resolution ESR Spin-Trapping Study at Q-Band (35 GHz). Anal. Biochem. 1996, 241, 75–81. [Google Scholar] [CrossRef] [PubMed]
- Lu, S.C. Regulation of Glutathione Synthesis. Mol. Asp. Med. 2009, 30, 42–59. [Google Scholar] [CrossRef] [PubMed]
- Di Monte, D.; Sandy, M.S.; Smith, M.T. Increased Efflux Rather than Oxidation Is the Mechanism of Glutathione Depletion by 1-Methyl-4-Phenyl-1,2,3,6-Tetrahydropyridine (MPTP). Biochem. Biophys. Res. Commun. 1987, 148, 153–160. [Google Scholar] [CrossRef] [PubMed]
- Brechbuhl, H.M.; Gould, N.; Kachadourian, R.; Riekhof, W.R.; Voelker, D.R.; Day, B.J. Glutathione Transport Is a Unique Function of the ATP-Binding Cassette Protein ABCG2. J. Biol. Chem. 2010, 285, 16582–16587. [Google Scholar] [CrossRef]
- Nasr, R.; Lorendeau, D.; Khonkarn, R.; Dury, L.; Pérès, B.; Boumendjel, A.; Cortay, J.-C.; Falson, P.; Chaptal, V.; Baubuchon-Cortay, H. Molecular Analysis of the Massive GSH Transport Mechanism Mediated by The Human Multidrug Resistant Protein 1/ABCC1. Sci. Rep. 2020, 10, 7616. [Google Scholar] [CrossRef]
- Dickinson, D.A.; Forman, H.J. Cellular Glutathione and Thiols Metabolism. Biochem. Pharmacol. 2002, 64, 1019–1026. [Google Scholar] [CrossRef]
- Ketterer, B. The Role of Nonenzymatic Reactions of Glutathione in Xenobiotic Metabolism. Drug Metab. Rev. 1982, 13, 161–187. [Google Scholar] [CrossRef]
- Sheng-Huang, C.; Chieh-Hsin, C.; Mu-Chun, Y.; Wen-Tung, H.; Chia-Ying, H.; Ya-Ting, H.; Wan-Ling, S.; Jiuan-Jen, S.; Chih-Yang, H.; Jer-Yuh, L. Effects of Estrogen on Glutathione and Catalase Levels in Human Erythrocyte during Menstrual Cycle. Biomed. Rep. 2015, 3, 266–268. [Google Scholar] [CrossRef]
- Suojanen, J.N.; Gay, R.J.; Hilf, R. Influence of Estrogen on Glutathione Levels and Glutathione-Metabolizing Enzymes in Uteri and R3230AC Mammary Tumors of Rats. Biochim. Biophys. Acta General. Subj. 1980, 630, 485–496. [Google Scholar] [CrossRef]
- Almeida, M.; Soares, M.; Fonseca-Moutinho, J.; Ramalhinho, A.C.; Breitenfeld, L. Influence of Estrogenic Metabolic Pathway Genes Polymorphisms on Postmenopausal Breast Cancer Risk. Pharmaceuticals 2021, 14, 94. [Google Scholar] [CrossRef] [PubMed]
- Kotsos, D.; Tziomalos, K. Microsomal Prostaglandin E Synthase-1 and -2: Emerging Targets in Non-Alcoholic Fatty Liver Disease. Int. J. Mol. Sci. 2023, 24, 3049. [Google Scholar] [CrossRef]
- Thorén, S.; Jakobsson, P.-J. Coordinate Up- and down-Regulation of Glutathione-Dependent Prostaglandin E Synthase and Cyclooxygenase-2 in A549 Cells. Eur. J. Biochem. 2000, 267, 6428–6434. [Google Scholar] [CrossRef]
- Maryon, E.B.; Molloy, S.A.; Kaplan, J.H. Cellular Glutathione Plays a Key Role in Copper Uptake Mediated by Human Copper Transporter 1. Am. J. Physiol. Cell Physiol. 2013, 304, C768–C779. [Google Scholar] [CrossRef] [PubMed]
- Freedman, J.H.; Ciriolo, M.R.; Peisach, J. The Role of Glutathione in Copper Metabolism and Toxicity. J. Biol. Chem. 1989, 264, 5598–5605. [Google Scholar] [CrossRef]
- Yilmaz, A.; Çomakli, V. Investigation of Effects of Some Metal Ions and Some Pesticides on Glutathione S-Transferase (GST) Enzyme Purified from Van Lake Fish (Chalcalburnus Tarichi) Kidney. Iğdır Üniversitesi Fen. Bilim. Enstitüsü Derg. 2023, 13, 1101–1109. [Google Scholar] [CrossRef]
- Park, M.K.; Choi, B.Y.; Kho, A.R.; Lee, S.H.; Hong, D.K.; Kang, B.S.; Lee, S.H.; Suh, S.W. The Protective Role of Glutathione on Zinc-Induced Neuron Death after Brain Injuries. Int. J. Mol. Sci. 2023, 24, 2950. [Google Scholar] [CrossRef] [PubMed]
- de Paula Arrifano, G.; Crespo-Lopez, M.E.; Lopes-Araújo, A.; Santos-Sacramento, L.; Barthelemy, J.L.; de Nazaré, C.G.L.; Freitas, L.G.R.; Augusto-Oliveira, M. Neurotoxicity and the Global Worst Pollutants: Astroglial Involvement in Arsenic, Lead, and Mercury Intoxication. Neurochem. Res. 2023, 48, 1047–1065. [Google Scholar] [CrossRef]
- Hider, R.C.; Kong, X.L. Glutathione: A Key Component of the Cytoplasmic Labile Iron Pool. BioMetals 2011, 24, 1179–1187. [Google Scholar] [CrossRef]
- Hider, R.; Aviles, M.V.; Chen, Y.-L.; Latunde-Dada, G.O. The Role of GSH in Intracellular Iron Trafficking. Int. J. Mol. Sci. 2021, 22, 1278. [Google Scholar] [CrossRef]
- Kumar, C.; Igbaria, A.; D’Autreaux, B.; Planson, A.-G.; Junot, C.; Godat, E.; Bachhawat, A.K.; Delaunay-Moisan, A.; Toledano, M.B. Glutathione Revisited: A Vital Function in Iron Metabolism and Ancillary Role in Thiol-Redox Control. EMBO J. 2011, 30, 2044–2056. [Google Scholar] [CrossRef] [PubMed]
- Daniel, T.; Faruq, H.M.; Laura Magdalena, J.; Manuela, G.; Christopher Horst, L. Role of GSH and Iron-Sulfur Glutaredoxins in Iron Metabolism—Review. Molecules 2020, 25, 3860. [Google Scholar] [CrossRef]
- Wang, Y.; Yen, F.S.; Zhu, X.G.; Timson, R.C.; Weber, R.; Xing, C.; Liu, Y.; Allwein, B.; Luo, H.; Yeh, H.-W.; et al. SLC25A39 Is Necessary for Mitochondrial Glutathione Import in Mammalian Cells. Nature 2021, 599, 136–140. [Google Scholar] [CrossRef]
- Niu, B.; Liao, K.; Zhou, Y.; Wen, T.; Quan, G.; Pan, X.; Wu, C. Application of Glutathione Depletion in Cancer Therapy: Enhanced ROS-Based Therapy, Ferroptosis, and Chemotherapy. Biomaterials 2021, 277, 121110. [Google Scholar] [CrossRef] [PubMed]
- Chiang, S.-K.; Chen, S.-E.; Chang, L.-C. A Dual Role of Heme Oxygenase-1 in Cancer Cells. Int. J. Mol. Sci. 2018, 20, 39. [Google Scholar] [CrossRef] [PubMed]
- Appenzeller-Herzog, C. Glutathione- and Non-Glutathione-Based Oxidant Control in the Endoplasmic Reticulum. J. Cell. Sci. 2011, 124, 847–855. [Google Scholar] [CrossRef]
- Holmgren, A.; Sengupta, R. The Use of Thiols by Ribonucleotide Reductase. Free. Radic. Biol. Med. 2010, 49, 1617–1628. [Google Scholar] [CrossRef]
- Marí, M.; De Gregorio, E.; De Dios, C.; Roca-Agujetas, V.; Cucarull, B.; Tutusaus, A.; Morales, A.; Colell, A. Mitochondrial Glutathione: Recent Insights and Role in Disease. Antioxidants. 2020, 9, 909. [Google Scholar] [CrossRef]
- Oestreicher, J.; Morgan, B. Glutathione: Subcellular Distribution and Membrane Transport. Biochem. Cell Biol. 2019, 97, 270–289. [Google Scholar] [CrossRef]
- Bansal, A.; Simon, M.C. Glutathione Metabolism in Cancer Progression and Treatment Resistance. J. Cell Biol. 2018, 217, 2291–2298. [Google Scholar] [CrossRef]
- Kennedy, L.; Sandhu, J.K.; Harper, M.E.; Cuperlovic-culf, M. Role of Glutathione in Cancer: From Mechanisms to Therapies. Biomolecules 2020, 10, 1429. [Google Scholar] [CrossRef]
- Pal, D.; Rai, A.; Checker, R.; Patwardhan, R.S.; Singh, B.; Sharma, D.; Sandur, S.K. Role of Protein S-Glutathionylation in Cancer Progression and Development of Resistance to Anti-Cancer Drugs. Arch. Biochem. Biophys. 2021, 704, 108890. [Google Scholar] [CrossRef]
- Kansanen, E.; Kuosmanen, S.M.; Leinonen, H.; Levonen, A.-L. The Keap1-NRF2 Pathway: Mechanisms of Activation and Dysregulation in Cancer. Redox. Biol. 2013, 1, 45–49. [Google Scholar] [CrossRef]
- Gupta, R.K.; Patel, A.K.; Shah, N.; Choudhary, A.K.; Jha, U.K.; Yadav, U.C.; Gupta, P.K.; Pakuwal, U. Oxidative Stress and Antioxidants in Disease and Cancer: A Review. Asian Pac. J. Cancer Prev. 2014, 15, 4405–4409. [Google Scholar] [CrossRef]
- Kitano, Y.; Baba, Y.; Nakagawa, S.; Miyake, K.; Iwatsuki, M.; Ishimoto, T.; Yamashita, Y.; Yoshida, N.; Watanabe, M.; Nakao, M.; et al. NRF2 Promotes Oesophageal Cancer Cell Proliferation via Metabolic Reprogramming and Detoxification of Reactive Oxygen Species. J. Pathol. 2018, 244, 346–357. [Google Scholar] [CrossRef]
- Gamcsik, M.P.; Kasibhatla, M.S.; Teeter, S.D.; Colvin, O.M. Glutathione Levels in Human Tumors. Biomarkers 2012, 17, 671–691. [Google Scholar] [CrossRef]
- Potęga, A. Glutathione-Mediated Conjugation of Anticancer Drugs: An Overview of Reaction Mechanisms and Biological Significance for Drug Detoxification and Bioactivation. Molecules 2022, 27, 5252. [Google Scholar] [CrossRef]
- Alaswad, H.A.; Mahbub, A.A.; Le Maitre, C.L.; Jordan-Mahy, N. Molecular Action of Polyphenols in Leukaemia and Their Therapeutic Potential. Int. J. Mol. Sci. 2021, 22, 3085. [Google Scholar] [CrossRef]
- Jeon, S.E.; Choi-Kwon, S.; Park, K.A.; Lee, H.J.; Park, M.S.; Lee, J.H.; Kwon, S.B.; Park, K.C. Dietary Supplementation of (+)-Catechin Protects against UVB-Induced Skin Damage by Modulating Antioxidant Enzyme Activities. Photodermatol. Photoimmunol. Photomed. 2003, 19, 235–241. [Google Scholar] [CrossRef]
- Cao, Z.; Li, Y. Potent Induction of Cellular Antioxidants and Phase 2 Enzymes by Resveratrol in Cardiomyocytes: Protection against Oxidative and Electrophilic Injury. Eur. J. Pharmacol. 2004, 489, 39–48. [Google Scholar] [CrossRef]
- Acquaviva, R.; Di Giacomo, C.; Sorrenti, V.; Galvano, F.; Santangelo, R.; Cardile, V.; Gangia, S.; D’Orazio, N.; Abraham, N.G.; Vanella, L. Antiproliferative Effect of Oleuropein in Prostate Cell Lines. Int. J. Oncol. 2012, 41, 31–38. [Google Scholar] [CrossRef]
- Myhrstad, M.C.W.; Carlsen, H.; Nordström, O.; Blomhoff, R.; Moskaug, J.Ø. Flavonoids Increase the Intracellular Glutathione Level by Transactivation of the γ-Glutamylcysteine Synthetase Catalytical Subunit Promoter. Free Radic. Biol. Med. 2002, 32, 386–393. [Google Scholar] [CrossRef]
- Cao, G.; Sofic, E.; Prior, R.L. Antioxidant and Prooxidant Behavior of Flavonoids: Structure-Activity Relationships. Free Radic. Biol. Med. 1997, 22, 749–760. [Google Scholar] [CrossRef]
- Talalay, P.; Dinkova-Kostova, A.T.; Holtzclaw, W.D. Importance of Phase 2 Gene Regulation in Protection against Electrophile and Reactive Oxygen Toxicity and Carcinogenesis. Adv. Enzym. Regul. 2003, 43, 121–134. [Google Scholar] [CrossRef]
- Li, F.; Li, S.; Li, H.-B.; Deng, G.-F.; Ling, W.-H.; Wu, S.; Xu, X.-R.; Chen, F. Antiproliferative Activity of Peels, Pulps and Seeds of 61 Fruits. J. Funct. Foods 2013, 5, 1298–1309. [Google Scholar] [CrossRef]
- Li, F.; Li, S.; Li, H.-B.; Deng, G.-F.; Ling, W.-H.; Xu, X.-R. Antiproliferative Activities of Tea and Herbal Infusions. Food Funct. 2013, 4, 530–538. [Google Scholar] [CrossRef]
- Zhou, Y.; Zheng, J.; Li, Y.; Xu, D.-P.; Li, S.; Chen, Y.-M.; Li, H.-B. Natural Polyphenols for Prevention and Treatment of Cancer. Nutrients 2016, 8, 515. [Google Scholar] [CrossRef]
- Bohmont, C.; Aaronson, L.M.; Mann, K.; Pardini, R.S. Inhibition of Mitochondrial NADH Oxidase, Succinoxidase, and ATPase by Naturally Occurring Flavonoids. J. Nat. Prod. 1987, 50, 427–433. [Google Scholar] [CrossRef]
- Leslie, E.M.; Deeley, R.G.; Cole, S.P.C. Bioflavonoid Stimulation of Glutathione Transport by the 190-KDa Multidrug Resistance Protein 1 (MRP1). Drug Metab.Dispos. 2003, 31, 11–15. [Google Scholar] [CrossRef]
- Zhang, K.; Mack, P.; Wong, K.P. Glutathione-Related Mechanisms in Cellular Resistance to Anticancer Drugs. Int. J. Oncol. 1998, 12, 871–953. [Google Scholar] [CrossRef]
- Nguyen, H.; Zhang, S.; Morris, M.E. Effect of Flavonoids on MRP1-Mediated Transport in Panc-1 Cells. J. Pharm. Sci. 2003, 92, 250–257. [Google Scholar] [CrossRef]
- Ballatori, N.; Hammond, C.L.; Cunningham, J.B.; Krance, S.M.; Marchan, R. Molecular Mechanisms of Reduced Glutathione Transport: Role of the MRP/CFTR/ABCC and OATP/SLC21A Families of Membrane Proteins. Toxicol. Appl. Pharmacol. 2005, 204, 238–255. [Google Scholar] [CrossRef]
- Kachadourian, R.; Day, B.J. Flavonoid-Induced Glutathione Depletion: Potential Implications for Cancer Treatment. Free Radic. Biol. Med. 2006, 41, 65–76. [Google Scholar] [CrossRef]
- Sabzevari, O.; Galati, G.; Moridani, M.Y.; Siraki, A.; O’Brien, P.J. Molecular Cytotoxic Mechanisms of Anticancer Hydroxychalcones. Chem. Biol. Interact. 2004, 148, 57–67. [Google Scholar] [CrossRef]
- Mahbub, A.; Le Maitre, C.; Haywood-Small, S.; Cross, N.; Jordan-Mahy, N. Polyphenols Act Synergistically with Doxorubicin and Etoposide in Leukaemia Cell Lines. Cell Death Discov. 2015, 1, 15043. [Google Scholar] [CrossRef]
- Mahbub, A.; Le Maitre, C.; Haywood-Small, S.; Cross, N.; Jordan-Mahy, N. Dietary Polyphenols Influence Antimetabolite Agents: Methotrexate, 6-Mercaptopurine and 5-Fluorouracil in Leukemia Cell Lines. Oncotarget 2017, 8, 104877–104893. [Google Scholar] [CrossRef]
- Mahbub, A.A.; Le Maitre, C.L.; Haywood-Small, S.L.; Cross, N.A.; Jordan-Mahy, N. Glutathione Is Key to the Synergistic Enhancement of Doxorubicin and Etoposide by Polyphenols in Leukaemia Cell Lines. Cell. Death Dis. 2015, 6, e2028. [Google Scholar] [CrossRef]
- Li, S.; Qiao, S.; Zhang, J.; Li, K. Quercetin Increase the Chemosensitivity of Breast Cancer Cells to Doxorubicin Via PTEN/Akt Pathway. Anti-Cancer Agent Med. Chem. 2015, 15, 1185–1189. [Google Scholar] [CrossRef]
- Acquaviva, R.; Malfa, G.A.; Loizzo, M.R.; Xiao, J.; Bianchi, S.; Tundis, R. Advances on Natural Abietane, Labdane and Clerodane Diterpenes as Anti-Cancer Agents: Sources and Mechanisms of Action. Molecules 2022, 27, 4791. [Google Scholar] [CrossRef]
- Bailly, C. Atractylenolides, Essential Components of Atractylodes-Based Traditional Herbal Medicines: Antioxidant, Anti-Inflammatory and Anticancer Properties. Eur. J. Pharmacol. 2021, 891, 173735. [Google Scholar] [CrossRef]
- Kohandel, Z.; Farkhondeh, T.; Aschner, M.; Pourbagher-Shahri, A.M.; Samarghandian, S. Anti-Inflammatory Action of Astaxanthin and Its Use in the Treatment of Various Diseases. Biomed. Pharmacother. 2022, 145, 112179. [Google Scholar] [CrossRef]
- Sharifi-Rad, M.; Varoni, E.M.; Iriti, M.; Martorell, M.; Setzer, W.N.; del Mar Contreras, M.; Salehi, B.; Soltani-Nejad, A.; Rajabi, S.; Tajbakhsh, M.; et al. Carvacrol and Human Health: A Comprehensive Review. Phytother. Res. 2018, 32, 1675–1687. [Google Scholar] [CrossRef]
- Soethoudt, M.; Peskin, A.V.; Dickerhof, N.; Paton, L.N.; Pace, P.E.; Winterbourn, C.C. Interaction of Adenanthin with Glutathione and Thiol Enzymes: Selectivity for Thioredoxin Reductase and Inhibition of Peroxiredoxin Recycling. Free Radic. Biol. Med. 2014, 77, 331–339. [Google Scholar] [CrossRef]
- Merrill, J.C.; Kim, H.L.; Safe, S.; Murray, C.A.; Hayes, M.A. Role of Glutathione in the Toxicity of the Sesquiterpene Lactones Hymenoxon and Helenalin. J. Toxicol. Environ. Health 1988, 23, 159–169. [Google Scholar] [CrossRef]
- Kamran, S.; Sinniah, A.; Abdulghani, M.A.M.; Alshawsh, M.A. Therapeutic Potential of Certain Terpenoids as Anticancer Agents: A Scoping Review. Cancers 2022, 14, 1100. [Google Scholar] [CrossRef]
- Stockwell, B.R.; Jiang, X.; Gu, W. Emerging Mechanisms and Disease Relevance of Ferroptosis. Trends Cell. Biol. 2020, 30, 478–490. [Google Scholar] [CrossRef]
- Malfa, G.A.; Tomasello, B.; Acquaviva, R.; Genovese, C.; La Mantia, A.; Cammarata, F.P.; Ragusa, M.; Renis, M.; Di Giacomo, C. Betula Etnensis Raf. (Betulaceae) Extract Induced HO-1 Expression and Ferroptosis Cell Death in Human Colon Cancer Cells. Int. J. Mol. Sci. 2019, 20, 2723. [Google Scholar] [CrossRef]
- Jiang, X.; Stockwell, B.R.; Conrad, M. Ferroptosis: Mechanisms, Biology and Role in Disease. Nat. Rev. Mol. Cell. Biol. 2021, 22, 266–282. [Google Scholar] [CrossRef]
- Candido, S.; Tomasello, B.; Lavoro, A.; Falzone, L.; Gattuso, G.; Russo, A.; Paratore, S.; McCubrey, J.A.; Libra, M. Bioinformatic Analysis of the LCN2–SLC22A17–MMP9 Network in Cancer: The Role of DNA Methylation in the Modulation of Tumor Microenvironment. Front. Cell. Dev. Biol. 2022, 10, 945586. [Google Scholar] [CrossRef]
- Zheng, K.; Dong, Y.; Yang, R.; Liang, Y.; Wu, H.; He, Z. Regulation of Ferroptosis by Bioactive Phytochemicals: Implications for Medical Nutritional Therapy. Pharmacol. Res. 2021, 168, 105580. [Google Scholar] [CrossRef]
- Zhang, J.; Wang, N.; Zhou, Y.; Wang, K.; Sun, Y.; Yan, H.; Han, W.; Wang, X.; Wei, B.; Ke, Y.; et al. Oridonin Induces Ferroptosis by Inhibiting Gamma-glutamyl Cycle in TE1 Cells. Phytother. Res. 2021, 35, 494–503. [Google Scholar] [CrossRef] [PubMed]
- Fujii, J.; Homma, T.; Kobayashi, S. Ferroptosis Caused by Cysteine Insufficiency and Oxidative Insult. Free Radic. Res. 2020, 54, 969–980. [Google Scholar] [CrossRef] [PubMed]
- Wen, Y.; Chen, H.; Zhang, L.; Wu, M.; Zhang, F.; Yang, D.; Shen, J.; Chen, J. Glycyrrhetinic Acid Induces Oxidative/Nitrative Stress and Drives Ferroptosis through Activating NADPH Oxidases and INOS, and Depriving Glutathione in Triple-Negative Breast Cancer Cells. Free Radic. Biol. Med. 2021, 173, 41–51. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Liu, Z.; Sui, X.; Wu, Q.; Wang, J.; Xu, C. Elemene Injection as Adjunctive Treatment to Platinum-Based Chemotherapy in Patients with Stage III/IV Non-Small Cell Lung Cancer: A Meta-Analysis Following the PRISMA Guidelines. Phytomedicine 2019, 59, 152787. [Google Scholar] [CrossRef] [PubMed]
- Wu, P.; Dong, X.-M.; Song, G.-Q.; Wei, M.-M.; Fang, C.; Zheng, F.-B.; Zhao, Y.-J.; Lu, H.-Q.; Cheng, L.-H.; Zhou, J.-L.; et al. Bioactivity-Guided Discovery of Quality Control Markers in Rhizomes of Curcuma Wenyujin Based on Spectrum-Effect Relationship against Human Lung Cancer Cells. Phytomedicine 2021, 86, 153559. [Google Scholar] [CrossRef] [PubMed]
- Song, G.-Q.; Wu, P.; Dong, X.-M.; Cheng, L.-H.; Lu, H.-Q.; Lin, Y.-Y.; Tang, W.-Y.; Xie, T.; Zhou, J.-L. Elemene Induces Cell Apoptosis via Inhibiting Glutathione Synthesis in Lung Adenocarcinoma. J. Ethnopharmacol. 2023, 311, 116409. [Google Scholar] [CrossRef]
- Zhang, J.; Zhang, M.; Zhang, W.-H.; Zhu, Q.-M.; Ning, J.; Huo, X.-K.; Xiao, H.-T.; Sun, C.-P. Total Terpenoids of Inula Japonica Activated the NRF2 Receptor to Alleviate the Inflammation and Oxidative Stress in LPS-Induced Acute Lung Injury. Phytomedicine 2022, 107, 154377. [Google Scholar] [CrossRef]
- Suntres, Z.E.; Coccimiglio, J.; Alipour, M. The Bioactivity and Toxicological Actions of Carvacrol. Crit. Rev. Food Sci. Nutr. 2015, 55, 304–318. [Google Scholar] [CrossRef]
- Anwar, S.; Khan, A.; Irshad, N. Pharmacological Evaluation of Carvacrol Anti-Migraine Potential. Naunyn Schmiedebergs Arch. Pharmacol. 2023, 396, 1309–1324. [Google Scholar] [CrossRef]
- Dong, H.; Zhang, J.; Rong, H.; Zhang, X.; Dong, M. Paeoniflorin and Plycyrrhetinic Acid Synergistically Alleviate MPP+/MPTP-Induced Oxidative Stress through NRF2-Dependent Glutathione Biosynthesis Mechanisms. ACS Chem. Neurosci. 2021, 12, 1100–1111. [Google Scholar] [CrossRef]
- Castellano, J.M.; Guinda, A.; Delgado, T.; Rada, M.; Cayuela, J.A. Biochemical Basis of the Antidiabetic Activity of Oleanolic Acid and Related Pentacyclic Triterpenes. Diabetes 2013, 62, 1791–1799. [Google Scholar] [CrossRef]
- Kumar, S.; Sharma, S.; Vasudeva, N.; Ranga, V. In Vivo Anti-Hyperglycemic and Antioxidant Potentials of Ethanolic Extract from Tecomella Undulata. Diabetol. Metab. Syndr. 2012, 4, 33. [Google Scholar] [CrossRef]
- Bacanlı, M.; Anlar, H.G.; Aydın, S.; Çal, T.; Arı, N.; Ündeğer Bucurgat, Ü.; Başaran, A.A.; Başaran, N. D-Limonene Ameliorates Diabetes and Its Complications in Streptozotocin-Induced Diabetic Rats. Food Chemical. Toxicol. 2017, 110, 434–442. [Google Scholar] [CrossRef] [PubMed]
- Malfa, G.A.; Tomasello, B.; Acquaviva, R.; Mantia, A.L.; Pappalardo, F.; Ragusa, M.; Renis, M.; Di Giacomo, C. The Antioxidant Activities of Betula Etnensis Rafin. Ethanolic Extract Exert Protective and Anti-Diabetic Effects on Streptozotocin-Induced Diabetes in Rats. Antioxidants 2020, 9, 847. [Google Scholar] [CrossRef] [PubMed]
- Tan, M.; Yin, Y.; Ma, X.; Zhang, J.; Pan, W.; Tan, M.; Zhao, Y.; Yang, T.; Jiang, T.; Li, H. Glutathione System Enhancement for Cardiac Protection: Pharmacological Options against Oxidative Stress and Ferroptosis. Cell. Death Dis. 2023, 14, 131. [Google Scholar] [CrossRef]
- Lu, H.; Xiao, H.; Dai, M.; Xue, Y.; Zhao, R. Britanin Relieves Ferroptosis-Mediated Myocardial Ischaemia/Reperfusion Damage by Upregulating GPX4 through Activation of AMPK/GSK3β/NRF2 Signalling. Pharm. Biol. 2022, 60, 38–45. [Google Scholar] [CrossRef]
- Guo, Z.; Yan, M.; Chen, L.; Fang, P.; Li, Z.; Wan, Z.; Cao, S.; Hou, Z.; Wei, S.; Li, W.; et al. NRF2-dependent Antioxidant Response Mediated the Protective Effect of Tanshinone IIA on Doxorubicin-induced Cardiotoxicity. Exp. Ther. Med. 2018, 16, 3333–3344. [Google Scholar] [CrossRef]
- Lang, X.; Zhang, X.; Wang, D.; Zhou, W. In Vitro and In Vivo Metabolic Activation of Obacunone, A Bioactive and Potentially Hepatotoxic Constituent of Dictamni Cortex. Planta Med. 2020, 86, 686–695. [Google Scholar] [CrossRef] [PubMed]
- Prieto, M.A.; López, C.J.; Simal-Gandara, J. Glucosinolates: Molecular Structure, Breakdown, Genetic, Bioavailability, Properties and Healthy and Adverse Effects. Adv. Food Nutr. Res. 2019, 90, 305–350. [Google Scholar] [PubMed]
- Shirakawa, M.; Tanida, M.; Ito, T. The Cell Differentiation of Idioblast Myrosin Cells: Similarities with Vascular and Guard Cells. Front. Plant Sci. 2022, 12, 829541. [Google Scholar] [CrossRef] [PubMed]
- Kamal, R.M.; Abdull Razis, A.F.; Mohd Sukri, N.S.; Perimal, E.K.; Ahmad, H.; Patrick, R.; Djedaini-Pilard, F.; Mazzon, E.; Rigaud, S. Beneficial Health Effects of Glucosinolates-Derived Isothiocyanates on Cardiovascular and Neurodegenerative Diseases. Molecules 2022, 27, 624. [Google Scholar] [CrossRef]
- Vanduchova, A.; Anzenbacher, P.; Anzenbacherova, E. Isothiocyanate from Broccoli, Sulforaphane, and Its Properties. J. Med. Food 2019, 22, 121–126. [Google Scholar] [CrossRef]
- Figueiredo, S.; Filho, S.; Nogueira-Machado, J.; Caligiorne, R. The Anti-Oxidant Properties of Isothiocyanates: A Review. Recent. Pat. Endocr. Metab. Immune Drug Discov. 2013, 7, 213–225. [Google Scholar] [CrossRef] [PubMed]
- Abdull Razis, A.F.; Konsue, N.; Ioannides, C. Isothiocyanates and Xenobiotic Detoxification. Mol. Nutr. Food Res. 2018, 62, 1700916. [Google Scholar] [CrossRef] [PubMed]
- Costa-Pérez, A.; Núñez-Gómez, V.; Baenas, N.; Di Pede, G.; Achour, M.; Manach, C.; Mena, P.; Del Rio, D.; García-Viguera, C.; Moreno, D.A.; et al. Systematic Review on the Metabolic Interest of Glucosinolates and Their Bioactive Derivatives for Human Health. Nutrients 2023, 15, 1424. [Google Scholar] [CrossRef] [PubMed]
- Tomasello, B.; Di Mauro, M.D.; Malfa, G.A.; Acquaviva, R.; Sinatra, F.; Spampinato, G.; Laudani, S.; Villaggio, G.; Bielak-Zmijewska, A.; Grabowska, W.; et al. Rapha Myr®, a Blend of Sulforaphane and Myrosinase, Exerts Antitumor and Anoikis-Sensitizing Effects on Human Astrocytoma Cells Modulating Sirtuins and DNA Methylation. Int. J. Mol. Sci. 2020, 21, 5328. [Google Scholar] [CrossRef]
- Zhang, Y.; Wu, Q.; Liu, J.; Zhang, Z.; Ma, X.; Zhang, Y.; Zhu, J.; Thring, R.W.; Wu, M.; Gao, Y.; et al. Sulforaphane Alleviates High Fat Diet-Induced Insulin Resistance via AMPK/NRF2/GPx4 Axis. Biomed. Pharmacother. 2022, 152, 113273. [Google Scholar] [CrossRef]
- Kim, M.; Lee, J.Y. Beneficial Effects of Sulforaphane on Diabetes and Its Complications via Both NRF2-Dependent and Independent Mechanisms. Food Suppl. Biomater. Health 2023, 3, e6. [Google Scholar] [CrossRef]
- Na, G.; He, C.; Zhang, S.; Tian, S.; Bao, Y.; Shan, Y. Dietary Isothiocyanates: Novel Insights into the Potential for Cancer Prevention and Therapy. Int. J. Mol. Sci. 2023, 24, 1962. [Google Scholar] [CrossRef]
- Shoaib, S.; Khan, F.B.; Alsharif, M.A.; Malik, M.S.; Ahmed, S.A.; Jamous, Y.F.; Uddin, S.; Tan, C.S.; Ardianto, C.; Tufail, S.; et al. Reviewing the Prospective Pharmacological Potential of Isothiocyanates in Fight against Female-Specific Cancers. Cancers 2023, 15, 2390. [Google Scholar] [CrossRef]
- Patil, P.B.; Patel, J.K. Chemopreventive Aspects, Investigational Anticancer Applications and Current Perspectives on Allyl Isothiocyanate (AITC): A Review. Mol. Cell. Biochem. 2023. [Google Scholar] [CrossRef] [PubMed]
- Gasmi, A.; Gasmi Benahmed, A.; Shanaida, M.; Chirumbolo, S.; Menzel, A.; Anzar, W.; Arshad, M.; Cruz-Martins, N.; Lysiuk, R.; Beley, N.; et al. Anticancer Activity of Broccoli, Its Organosulfur and Polyphenolic Compounds. Crit. Rev. Food Sci. Nutr. 2023, 1–19. [Google Scholar] [CrossRef]
- Espinosa-Diez, C.; Fierro-Fernandez, M.; Sanchez-Gomez, F.; Rodriguez-Pascual, F.; Alique, M.; Ruiz-Ortega, M.; Beraza, N.; Martinez-Chantar, M.L.; Fernandez-Hernando, C.; Lamas, S. Targeting of gamma-glutamyl-cysteine ligase by miR-433 reduces glutathione biosynthesis and promotes TGF-beta-dependent fibrogenesis. Antioxid. Redox Signal. 2015, 23, 1092–1105. [Google Scholar] [CrossRef]
- Li, R.; Chung, A.C.; Dong, Y.; Yang, W.; Zhong, X.; Lan, H.Y. The microRNA miR-433 promotes renal fibrosis by amplifying the TGF-beta/Smad3-Azin1 pathway. Kidney Int. 2013, 84, 1129–1144. [Google Scholar] [CrossRef]
- Kinoshita, C.; Aoyama, K.; Matsumura, N.; Kikuchi-Utsumi, K.; Watabe, M.; Nakaki, T. Rhythmic oscillations of the microRNA miR-96-5p play a neuroprotective role by indirectly regulating glutathione levels. Nat. Commun. 2014, 5, 3823. [Google Scholar] [CrossRef] [PubMed]
- Maes, O.C.; An, J.; Sarojini, H.; Wang, E. Murine microRNAs implicated in liver functions and aging process. Mechan. Ageing Dev. 2008, 129, 534–541. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.; Jiao, J.W.; Sun, K.X.; Zong, Z.H.; Zhao, Y. microRNA-133b targets glutathione S-transferase pi expression to increase ovarian cancer cell sensitivity to chemotherapy drugs. Drug Des. Dev. Ther. 2015, 9, 5225–5235. [Google Scholar]
- Uchida, Y.; Chiyomaru, T.; Enokida, H.; Kawakami, K.; Tatarano, S.; Kawahara, K.; Nishiyama, K.; Seki, N.; Nakagawa, M. miR-133a induces apoptosis through direct regulation of GSTP1 in bladder cancer cell lines. Urol. Oncol. Semin. Orig. Investig. 2013, 31, 115–123. [Google Scholar] [CrossRef] [PubMed]
- Marengo, B.; Pulliero, A.; Izzotti, A.; Domenicotti, C. miRNA Regulation of Glutathione Homeostasis in Cancer Initiation, Progression and Therapy Resistance. MicroRNA 2020, 9, 187–197. [Google Scholar] [CrossRef]
- Matoušková, P.; Hanousková, B.; Skálová, L. MicroRNAs as Potential Regulators of Glutathione Peroxidases Expression and Their Role in Obesity and Related Pathologies. Int. J. Mol. Sci. 2018, 19, 1199. [Google Scholar] [CrossRef]
- Kinoshita, C.; Kubota, N.; Aoyama, K. Glutathione Depletion and MicroRNA Dysregulation in Multiple System Atrophy: A Review. Int. J. Mol. Sci. 2022, 23, 15076. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
| Polyphenols | Effects on GSH Metabolism | Model | References |
|---|---|---|---|
| Resveratrol, Quercetin, EGCG | Increase GSH levels | U937 cells | [30,72] |
| Catechin | Increase expression of GPX, GR, and GLS enzymes | In vivo | [73] |
| Resveratrol | Increase GSH levels, GR, and GLS activities | H9C2 cells | [74] |
| Epicatechin EGCG | Prevents the drop in GSH levels | PC12 cells | [73] |
| Oleuropein, Protocatechuic acid, Isoflavones | Increase GR and GPx activities | J774A.1, LNCap, PC3 cells | [75] |
| Quercetin | Increase γGLCL activity | COS-1, HepG2 cells | [76] |
| Myricetin | Inactive γGLCL activity | In vitro | [77] |
| Flavonoids | Modulation of the mitochondrial respiratory chain of complexes I and II | In vitro | [82] |
| Apigenin, Naringenin, Genistein, Quercetin | Stimulate GSH transport by MRP1 | In vitro | [83] |
| Chrysin, Apigenin | Decrease GSH levels | A549, PC-3, HL-60 cells | [87] |
| Hydroxychalcones | Modulation of the mitochondrial respiratory chain of complexes; MRP-mediated GSH depletion | HepG2 cells | [88] |
| Quercetin, Apigenin, Rhein, Resveratrol | Cell cycle arrest and apoptosis | U937, MCF7cells | [72,89,90,91,92] |
| Terpenoids | Effects on GSH Metabolism | Model | References |
|---|---|---|---|
| Oridonin | Inhibition of GGT1 activity, GSH synthesis, and GPX4 expression; Induction of ferropototic death | TE1 cells | [105,106] |
| 18-β-glycyrrhetinic acid | Downregulation of SLC7A11 expression; Reduction in GSH content and GPx activity; Increase in oxidative stress; Ferroptosis activation | MDA-MB-231 cells | [107] |
| Betulinic acid (Betula etnensis Raf. Extract) | Depletion of intracellular GSH; Lipoperoxidation; Upregulation of HO-1 expression; Ferroptosis induction Increase GSH in plasma and tissue; LOOH reduction | Caco-2, cells, In vivo | [101,118] |
| Elemene | Decrease in the GSH/GSSG ratio; Downregulation of SLC7A11, GS and glutaminase; Upregulation of GCLM | A549 and PC9 cells, In vivo | [110] |
| Total terpenoids of Inula japonica Thunb | Stimulation of GSH synthesis; Upregulation of GCLC and GCLM | In vivo | [111] |
| Carvacrol | Increase in GSH and GST levels | In vivo | [113] |
| Glycyrrhetinic acid and paeoniflorin | Upregulation of GCLC and GCLM | SH-SY5Y cells, In vivo | [114] |
| D-limonene | Increase in GSH levels and GPX enzyme activity; GR activity reduction | In vivo | [117] |
| Britanin | Enhancement of GSH levels and GPX activity | In vivo | [120] |
| Tanshinone IIA | Increase in GSH content and GCLC activity | H9c2 cells, In vivo | [121] |
| Hymenoxon and helenalin | Hepatic glutathione depletion | In vivo | [98] |
| Obacunone | GSH bioactivation | In vivo, Human liver microsomes | [122] |
| Glucosinolates | Effects on GSH Metabolism | Model | References |
|---|---|---|---|
| ITCs | Increase Phase II detoxification enzymes | In vitro and in vivo | [127,128] |
| ITCs | Decrease GSH levels | In vitro | [13,128,129] |
| ITCs | Increase γGLCL and GS activity | HepG2 cells | [13,130,131] |
| SFN | Modulation of Nrf2/GPX4 pathway | In vitro and in vivo | [130,131,132] |
| ITCS | Hormetic effects on cell proliferation and tumor growth through GSH levels modulation | HepG2, MDA-MB-231, MCF-7, HT1376 cells and in vivo | [133,134,135] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Di Giacomo, C.; Malfa, G.A.; Tomasello, B.; Bianchi, S.; Acquaviva, R. Natural Compounds and Glutathione: Beyond Mere Antioxidants. Antioxidants 2023, 12, 1445. https://doi.org/10.3390/antiox12071445
Di Giacomo C, Malfa GA, Tomasello B, Bianchi S, Acquaviva R. Natural Compounds and Glutathione: Beyond Mere Antioxidants. Antioxidants. 2023; 12(7):1445. https://doi.org/10.3390/antiox12071445
Chicago/Turabian StyleDi Giacomo, Claudia, Giuseppe Antonio Malfa, Barbara Tomasello, Simone Bianchi, and Rosaria Acquaviva. 2023. "Natural Compounds and Glutathione: Beyond Mere Antioxidants" Antioxidants 12, no. 7: 1445. https://doi.org/10.3390/antiox12071445
APA StyleDi Giacomo, C., Malfa, G. A., Tomasello, B., Bianchi, S., & Acquaviva, R. (2023). Natural Compounds and Glutathione: Beyond Mere Antioxidants. Antioxidants, 12(7), 1445. https://doi.org/10.3390/antiox12071445