

Melatonin: A Potential Regulator of DNA Methylation

 ,
,  ,
,  , ,
, , 
Abstract
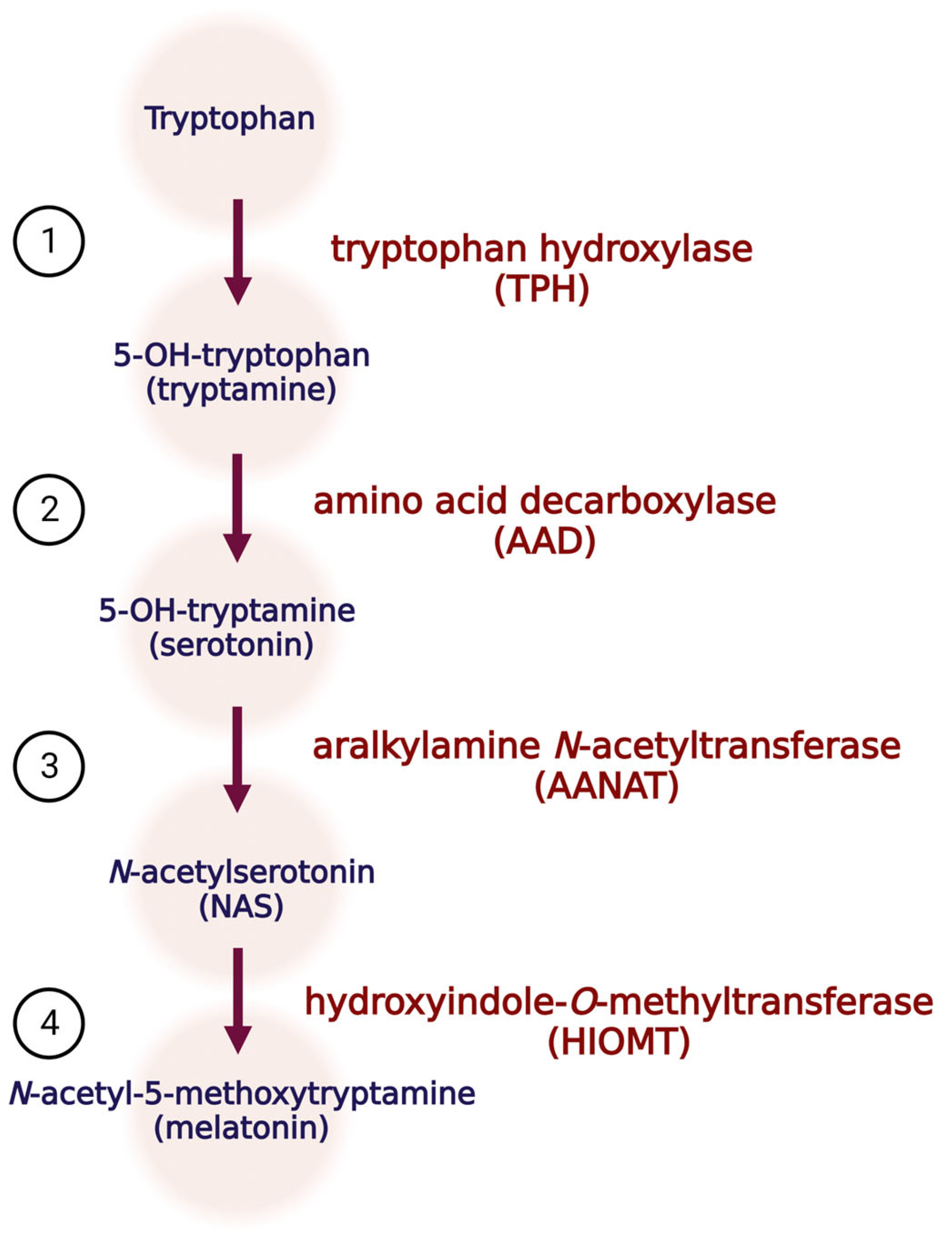
1. Melatonin Biosynthesis and Function
2. Alterations in DNA Methylation
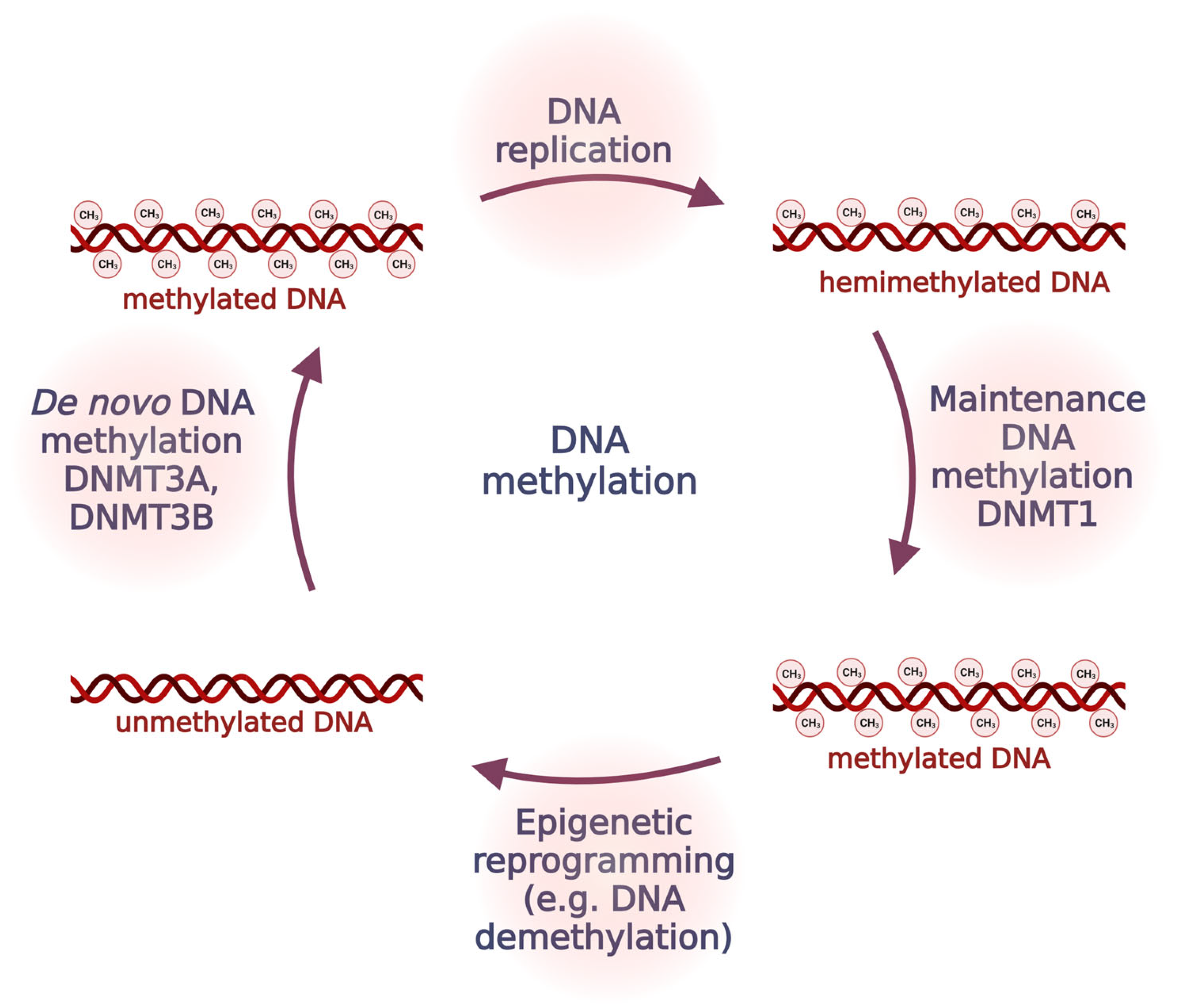
2.1. DNA Methylation
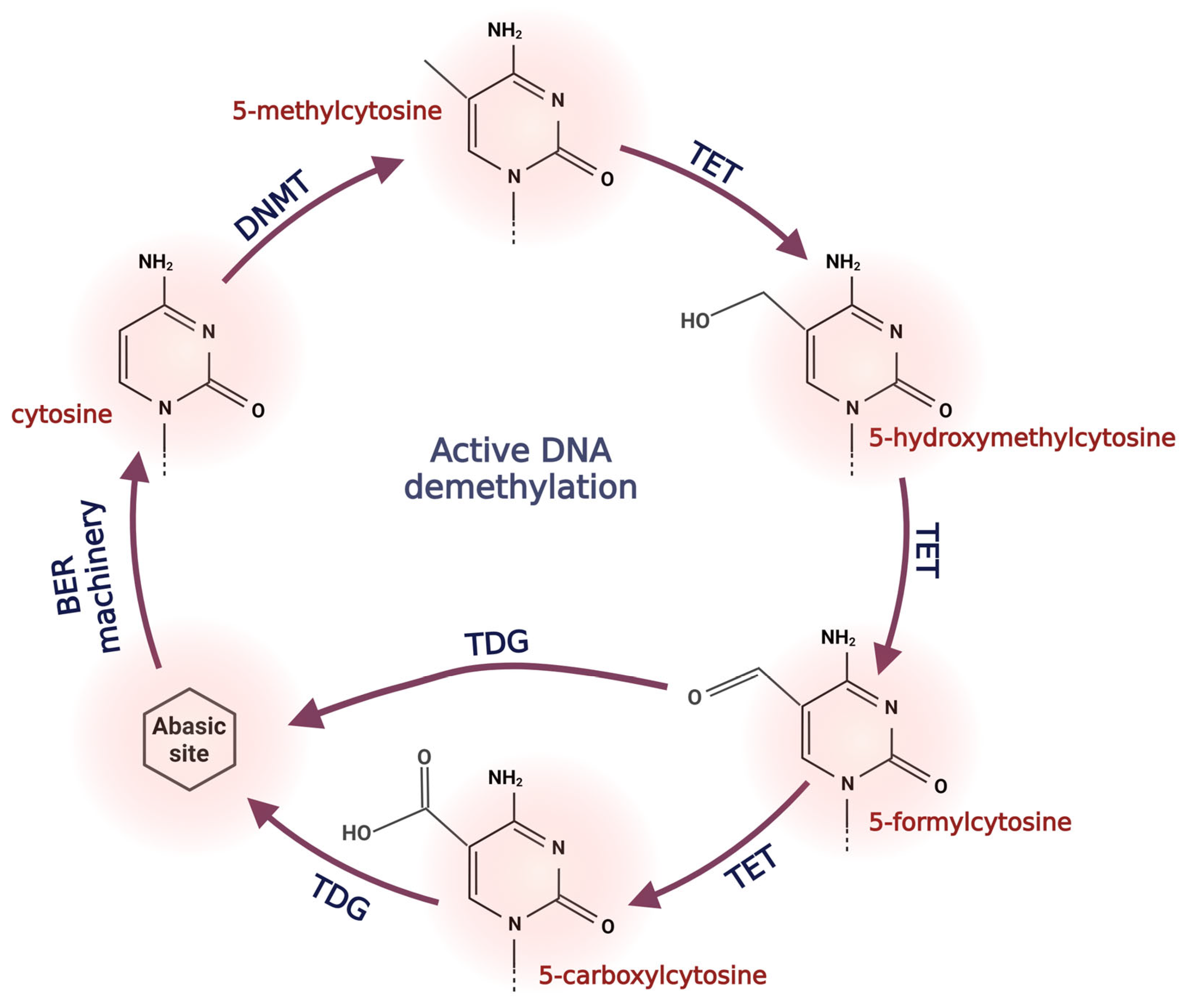
2.2. Active DNA Demethylation
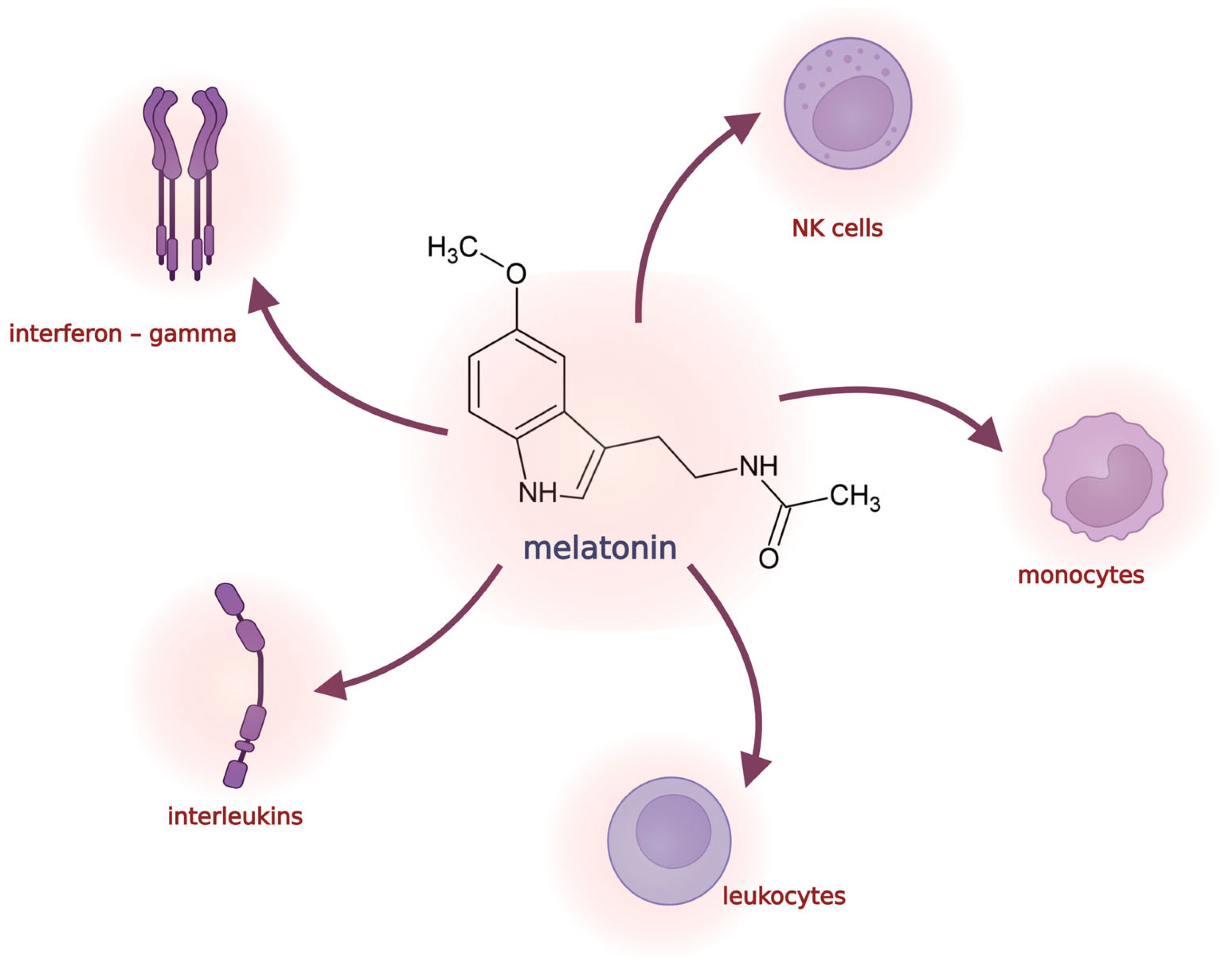
3. Melatonin and Its Role in DNA Methylation
3.1. Melatonin Regulates the Expression of DNMTs
3.2. Melatonin and Its Possible Impact on TET Proteins
3.3. Melatonin and DNA Methylation Changes under Artificial Light at Night
3.4. Melatonin and Circadian Clock Epigenetics
3.5. Melatonin and Methylation during Cell Development and Differentiation
4. Melatonin as a Part of Combined Epigenetic Therapy
4.1. Epigenetic Drugs Limitations
4.2. Melatonin’s Potential in Combined Therapy
4.3. Melatonin Plays Significant Role in Changing of DNA Methylation Pattern of Cancer-Related Genes

{kind=link}
{kind=link}
{kind=link}
{kind=link}
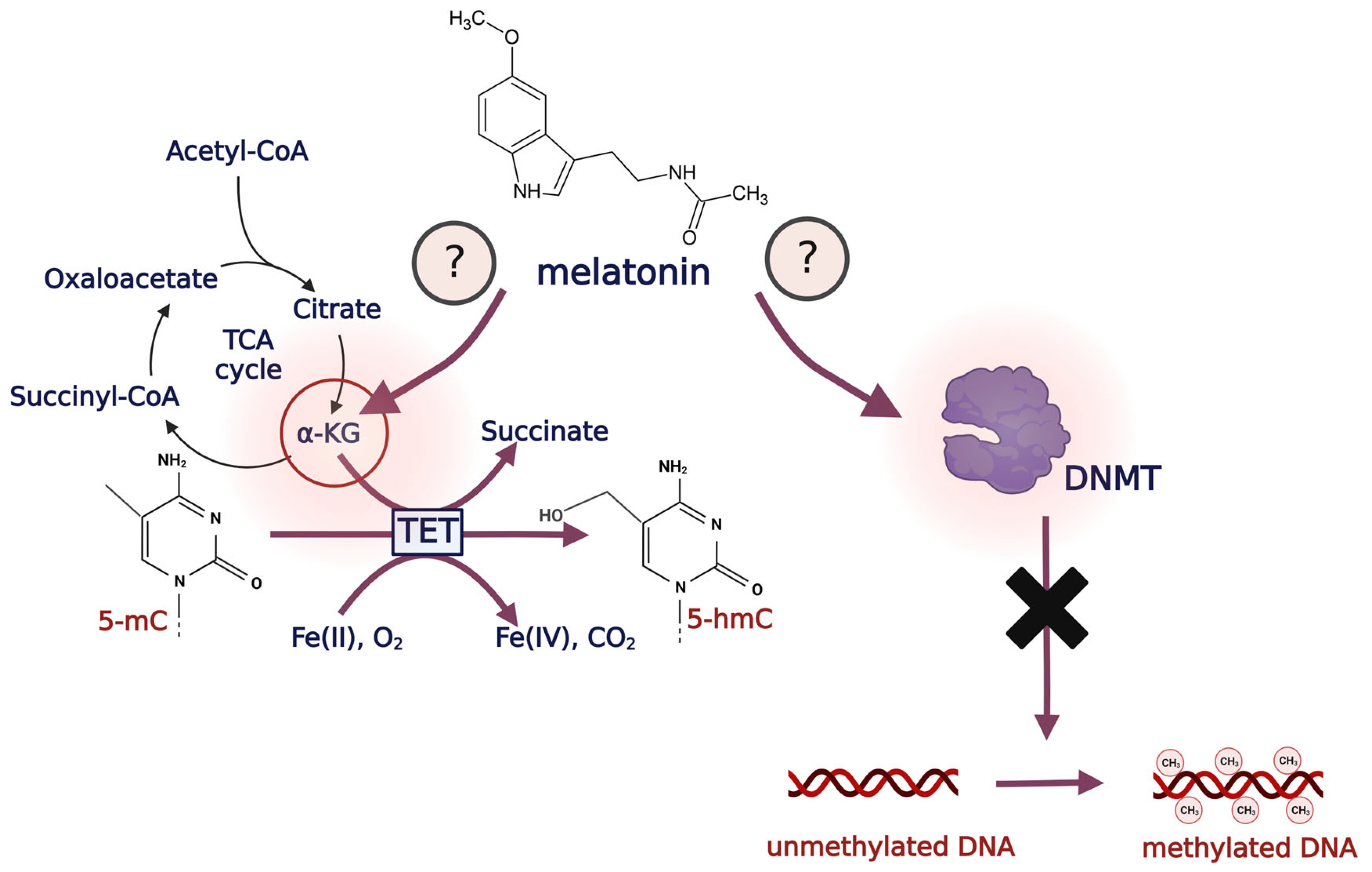{kind=link}
| Studied Cell Line/Mice Model | Main Finding | References |
|---|---|---|
| Oral cell carcinoma cell lines | Hypermethylation of CpG islands in MT1 gene restored after treatment with 5-AZA | [169] |
| Rat C6 glioma cells | Treatment with 5-AZA increases expression of MT1 gene | [168] |
| Porcine oocytes | Treatment with melatonin increased level of 5-mC and DNMT in prolonged culture | [179] |
| 4T1 mouse breast cancer model | Melatonin can mitigate the changes in DNMT activity and global DNA methylation level in mice exposed to artificial light at night | [102,103] |
| Brain tumor stem cell, A172 malignant glioma tumor cells | Melatonin treatment increases methylation of genes ABCG2/BCRP associated with multidrug resistance | [178] |
| MCF-7 mice breast cancer model | Melatonin treatment increases the favorable effect of paclitaxel by impact on IL-6, STAT3 and DNMT1 gene expression | [173] |
| MCF-7 breast cancer cells | Supplementation of melatonin increases expression of tumor suppressor gene and decreases expression of oncogenic genes possibly by impacting on methylation level | [177] |
5. Conclusions and Future Perspectives
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Reiter, R.J. Pineal Melatonin: Cell Biology of Its Synthesis and of Its Physiological Interactions. Endocr. Rev. 1991, 12, 151–180. [Google Scholar] [CrossRef]
- Roseboom, P.H.; Namboodiri, M.A.; Zimonjic, D.B.; Popescu, N.C.; Rodriguez, I.R.; Gastel, J.A.; Klein, D.C. Natural Melatonin “knockdown” in C57BL/6J Mice: Rare Mechanism Truncates Serotonin N-Acetyltransferase. Mol. Brain Res. Brain Res. 1998, 63, 189–197. [Google Scholar] [CrossRef]
- Naranjo, M.C.; Guerrero, J.M.; Rubio, A.; Lardone, P.J.; Carrillo-Vico, A.; Carrascosa-Salmoral, M.P.; Jiménez-Jorge, S.; Arellano, M.V.; Leal-Noval, S.R.; Leal, M.; et al. Melatonin Biosynthesis in the Thymus of Humans and Rats. Cell. Mol. Life Sci. 2007, 64, 781–790. [Google Scholar] [CrossRef]
- Janjetovic, Z.; Jarrett, S.G.; Lee, E.F.; Duprey, C.; Reiter, R.J.; Slominski, A.T. Melatonin and Its Metabolites Protect Human Melanocytes against UVB-Induced Damage: Involvement of NRF2-Mediated Pathways. Sci. Rep. 2017, 7, 1274. [Google Scholar] [CrossRef]
- Grant, S.G.; Melan, M.A.; Latimer, J.J.; Witt-Enderby, P.A. Melatonin and Breast Cancer: Cellular Mechanisms, Clinical Studies and Future Perspectives. Expert Rev. Mol. Med. 2009, 11, e5. [Google Scholar] [CrossRef]
- Tosini, G.; Fukuhara, C. Photic and Circadian Regulation of Retinal Melatonin in Mammals. J. Neuroendocrinol. 2003, 15, 364–369. [Google Scholar] [CrossRef]
- Konturek, S.J.; Konturek, P.C.; Brzozowska, I.; Pawlik, M.; Sliwowski, Z.; Cześnikiewicz-Guzik, M.; Kwiecień, S.; Brzozowski, T.; Bubenik, G.A.; Pawlik, W.W. Localization and Biological Activities of Melatonin in Intact and Diseased Gastrointestinal Tract (GIT). J. Physiol. Pharmacol. 2007, 58, 381–405. [Google Scholar]
- Tan, D.-X.; Manchester, L.C.; Terron, M.P.; Flores, L.J.; Reiter, R.J. One Molecule, Many Derivatives: A Never-Ending Interaction of Melatonin with Reactive Oxygen and Nitrogen Species? J. Pineal Res. 2007, 42, 28–42. [Google Scholar] [CrossRef]
- Wiechmann, A.F.; Summers, J.A. Circadian Rhythms in the Eye: The Physiological Significance of Melatonin Receptors in Ocular Tissues. Prog. Retin. Eye Res. 2008, 27, 137–160. [Google Scholar] [CrossRef]
- Reiter, R.J.; Tan, D.-X.; Fuentes-Broto, L. Melatonin: A Multitasking Molecule. Prog. Brain Res. 2010, 181, 127–151. [Google Scholar] [CrossRef]
- Hardeland, R.; Cardinali, D.P.; Srinivasan, V.; Spence, D.W.; Brown, G.M.; Pandi-Perumal, S.R. Melatonin—A Pleiotropic, Orchestrating Regulator Molecule. Prog. Neurobiol. 2011, 93, 350–384. [Google Scholar] [CrossRef]
- Lanoix, D.; Lacasse, A.-A.; Reiter, R.J.; Vaillancourt, C. Melatonin: The Smart Killer: The Human Trophoblast as a Model. Mol. Cell. Endocrinol. 2012, 348, 1–11. [Google Scholar] [CrossRef]
- Slominski, A.T.; Semak, I.; Fischer, T.W.; Kim, T.-K.; Kleszczyński, K.; Hardeland, R.; Reiter, R.J. Metabolism of Melatonin in the Skin: Why Is It Important? Exp. Dermatol. 2017, 26, 563–568. [Google Scholar] [CrossRef]
- Slominski, A.T.; Hardeland, R.; Zmijewski, M.A.; Slominski, R.M.; Reiter, R.J.; Paus, R. Melatonin: A Cutaneous Perspective on Its Production, Metabolism, and Functions. J. Investig. Dermatol. 2018, 138, 490–499. [Google Scholar] [CrossRef]
- Slominski, A.T.; Kim, T.-K.; Kleszczyński, K.; Semak, I.; Janjetovic, Z.; Sweatman, T.; Skobowiat, C.; Steketee, J.D.; Lin, Z.; Postlethwaite, A.; et al. Characterization of Serotonin and N-Acetylserotonin Systems in the Human Epidermis and Skin Cells. J. Pineal Res. 2020, 68, e12626. [Google Scholar] [CrossRef]
- Bocheva, G.; Slominski, R.M.; Janjetovic, Z.; Kim, T.-K.; Böhm, M.; Steinbrink, K.; Reiter, R.J.; Kleszczyński, K.; Slominski, A.T. Protective Role of Melatonin and Its Metabolites in Skin Aging. Int. J. Mol. Sci. 2022, 23, 1238. [Google Scholar] [CrossRef]
- Slominski, A.; Baker, J.; Rosano, T.G.; Guisti, L.W.; Ermak, G.; Grande, M.; Gaudet, S.J. Metabolism of Serotonin to N-Acetylserotonin, Melatonin, and 5-Methoxytryptamine in Hamster Skin Culture. J. Biol. Chem. 1996, 271, 12281–12286. [Google Scholar] [CrossRef]
- Slominski, A.; Pisarchik, A.; Semak, I.; Sweatman, T.; Szczesniewski, A.; Wortsman, J. Serotoninergic System in Hamster Skin. J. Investig. Dermatol. 2002, 119, 934–942. [Google Scholar] [CrossRef]
- Kleszczyński, K.; Bilska, B.; Stegemann, A.; Flis, D.J.; Ziolkowski, W.; Pyza, E.; Luger, T.A.; Reiter, R.J.; Böhm, M.; Slominski, A.T. Melatonin and Its Metabolites Ameliorate UVR-Induced Mitochondrial Oxidative Stress in Human MNT-1 Melanoma Cells. Int. J. Mol. Sci. 2018, 19, 3786. [Google Scholar] [CrossRef]
- Kleszczyński, K.; Zwicker, S.; Tukaj, S.; Kasperkiewicz, M.; Zillikens, D.; Wolf, R.; Fischer, T.W. Melatonin Compensates Silencing of Heat Shock Protein 70 and Suppresses Ultraviolet Radiation-Induced Inflammation in Human Skin Ex Vivo and Cultured Keratinocytes. J. Pineal Res. 2015, 58, 117–126. [Google Scholar] [CrossRef]
- Janjetovic, Z.; Nahmias, Z.P.; Hanna, S.; Jarrett, S.G.; Kim, T.-K.; Reiter, R.J.; Slominski, A.T. Melatonin and Its Metabolites Ameliorate Ultraviolet B-Induced Damage in Human Epidermal Keratinocytes. J. Pineal Res. 2014, 57, 90–102. [Google Scholar] [CrossRef] [PubMed]
- Skobowiat, C.; Brożyna, A.A.; Janjetovic, Z.; Jeayeng, S.; Oak, A.S.W.; Kim, T.-K.; Panich, U.; Reiter, R.J.; Slominski, A.T. Melatonin and Its Derivatives Counteract the Ultraviolet B Radiation-Induced Damage in Human and Porcine Skin Ex Vivo. J. Pineal Res. 2018, 65, e12501. [Google Scholar] [CrossRef] [PubMed]
- Pandi-Perumal, S.R.; Srinivasan, V.; Maestroni, G.J.M.; Cardinali, D.P.; Poeggeler, B.; Hardeland, R. Melatonin. Nature’s Most Versatile Biological Signal? FEBS J. 2006, 273, 2813–2838. [Google Scholar] [CrossRef] [PubMed]
- Reiter, R.J.; Tan, D.X.; Manchester, L.C.; Pilar Terron, M.; Flores, L.J.; Koppisepi, S. Medical Implications of Melatonin: Receptor-Mediated and Receptor-Independent Actions. Adv. Med. Sci. 2007, 52, 11–28. [Google Scholar]
- Slominski, A.T.; Zmijewski, M.A.; Jetten, A.M. RORα Is Not a Receptor for Melatonin. Bioessays 2016, 38, 1193–1194. [Google Scholar] [CrossRef]
- Hardeland, R. Melatonin and Retinoid Orphan Receptors: Demand for New Interpretations after Their Exclusion as Nuclear Melatonin Receptors. Melatonin Res. 2018, 1, 78–93. [Google Scholar] [CrossRef]
- Reiter, R.J.; Mayo, J.C.; Tan, D.-X.; Sainz, R.M.; Alatorre-Jimenez, M.; Qin, L. Melatonin as an Antioxidant: Under Promises but over Delivers. J. Pineal Res. 2016, 61, 253–278. [Google Scholar] [CrossRef]
- Torres, F.F.; Bernardo, V.S.; de Paula, C.P.; da Silva, J.P.M.d.O.; de Almeida, E.A.; da Cunha, A.F.; da Silva, D.G.H. Influence of Melatonin Treatment on Cellular Mechanisms of Redox Adaptation in K562 Erythroleukemic Cells. Genes 2022, 13, 2337. [Google Scholar] [CrossRef]
- Vriend, J.; Reiter, R.J. The Keap1-Nrf2-Antioxidant Response Element Pathway: A Review of Its Regulation by Melatonin and the Proteasome. Mol. Cell. Endocrinol. 2015, 401, 213–220. [Google Scholar] [CrossRef]
- Kleszczyński, K.; Zillikens, D.; Fischer, T.W. Melatonin Enhances Mitochondrial ATP Synthesis, Reduces Reactive Oxygen Species Formation, and Mediates Translocation of the Nuclear Erythroid 2-Related Factor 2 Resulting in Activation of Phase-2 Antioxidant Enzymes (γ-GCS, HO-1, NQO1) in Ultraviolet Radiation-Treated Normal Human Epidermal Keratinocytes (NHEK). J. Pineal Res. 2016, 61, 187–197. [Google Scholar] [CrossRef]
- Deng, Y.; Zhu, J.; Mi, C.; Xu, B.; Jiao, C.; Li, Y.; Xu, D.; Liu, W.; Xu, Z. Melatonin Antagonizes Mn-Induced Oxidative Injury through the Activation of Keap1-Nrf2-ARE Signaling Pathway in the Striatum of Mice. Neurotox. Res. 2015, 27, 156–171. [Google Scholar] [CrossRef] [PubMed]
- Ryšavá, A.; Vostálová, J.; Rajnochová Svobodová, A. Effect of Ultraviolet Radiation on the Nrf2 Signaling Pathway in Skin Cells. Int. J. Radiat. Biol. 2021, 97, 1383–1403. [Google Scholar] [CrossRef] [PubMed]
- Suwannakot, K.; Sritawan, N.; Prajit, R.; Aranarochana, A.; Sirichoat, A.; Pannangrong, W.; Wigmore, P.; Welbat, J.U. Melatonin Protects against the Side-Effects of 5-Fluorouracil on Hippocampal Neurogenesis and Ameliorates Antioxidant Activity in an Adult Rat Hippocampus and Prefrontal Cortex. Antioxidants 2021, 10, 615. [Google Scholar] [CrossRef] [PubMed]
- Sobhani, S.; Tehrani, A.-A.; Sobhani, G.; Fatima, S.; Ulloa, L.; Motaghinejad, M.; Atif, A. Melatonin Protects Against Titanium Oxide-Induced Neurotoxicity: Neurochemical, Neurobehavioral, and Histopathological Evidences. Biol. Trace Elem. Res. 2022. [Google Scholar] [CrossRef] [PubMed]
- Srinivasan, V.; Spence, D.W.; Trakht, I.; Pandi-Perumal, S.R.; Cardinali, D.P.; Maestroni, G.J. Immunomodulation by Melatonin: Its Significance for Seasonally Occurring Diseases. Neuroimmunomodulation 2008, 15, 93–101. [Google Scholar] [CrossRef]
- Cos, S.; Sánchez-Barceló, E.J. Melatonin and Mammary Pathological Growth. Front. Neuroendocrinol. 2000, 21, 133–170. [Google Scholar] [CrossRef]
- Reiter, R.J.; Rosales-Corral, S.A.; Tan, D.-X.; Acuna-Castroviejo, D.; Qin, L.; Yang, S.-F.; Xu, K. Melatonin, a Full Service Anti-Cancer Agent: Inhibition of Initiation, Progression and Metastasis. Int. J. Mol. Sci. 2017, 18, 843. [Google Scholar] [CrossRef]
- Bilska, B.; Schedel, F.; Piotrowska, A.; Stefan, J.; Zmijewski, M.; Pyza, E.; Reiter, R.J.; Steinbrink, K.; Slominski, A.T.; Tulic, M.K.; et al. Mitochondrial Function Is Controlled by Melatonin and Its Metabolites in Vitro in Human Melanoma Cells. J. Pineal Res. 2021, 70, e12728. [Google Scholar] [CrossRef]
- Nyamsambuu, A.; Khan, M.A.; Zhou, X.; Chen, H.-C. Molecular Mechanism of Inhibitory Effects of Melatonin on Prostate Cancer Cell Proliferation, Migration and Invasion. PLoS ONE 2022, 17, e0261341. [Google Scholar] [CrossRef]
- Park, S.-Y.; Jang, W.-J.; Yi, E.-Y.; Jang, J.-Y.; Jung, Y.; Jeong, J.-W.; Kim, Y.-J. Melatonin Suppresses Tumor Angiogenesis by Inhibiting HIF-1alpha Stabilization under Hypoxia. J. Pineal Res. 2010, 48, 178–184. [Google Scholar] [CrossRef]
- Alvarez-García, V.; González, A.; Alonso-González, C.; Martínez-Campa, C.; Cos, S. Regulation of Vascular Endothelial Growth Factor by Melatonin in Human Breast Cancer Cells. J. Pineal Res. 2013, 54, 373–380. [Google Scholar] [CrossRef]
- Maschio-Signorini, L.B.; Gelaleti, G.B.; Moschetta, M.G.; Borin, T.F.; Jardim-Perassi, B.V.; Lopes, J.R.; Lacerda, J.Z.; Roela, R.A.; Bordin, N.A.; Corrêa, L.A.; et al. Melatonin Regulates Angiogenic and Inflammatory Proteins in MDA-MB-231 Cell Line and in Co-Culture with Cancer-Associated Fibroblasts. Anticancer Agents Med. Chem. 2016, 16, 1474–1484. [Google Scholar] [CrossRef]
- Gonzalez-Haba, M.G.; Garcia-Mauriño, S.; Calvo, J.R.; Goberna, R.; Guerrero, J.M. High-Affinity Binding of Melatonin by Human Circulating T Lymphocytes (CD4+). FASEB J. 1995, 9, 1331–1335. [Google Scholar] [CrossRef]
- Plaimee, P.; Khamphio, M.; Weerapreeyakul, N.; Barusrux, S.; Johns, N.P. Immunomodulatory Effect of Melatonin in SK-LU-1 Human Lung Adenocarcinoma Cells Co-Cultured with Peripheral Blood Mononuclear Cells. Cell Prolif. 2014, 47, 406–415. [Google Scholar] [CrossRef]
- Kleszczyński, K.; Kim, T.-K.; Bilska, B.; Sarna, M.; Mokrzynski, K.; Stegemann, A.; Pyza, E.; Reiter, R.J.; Steinbrink, K.; Böhm, M.; et al. Melatonin Exerts Oncostatic Capacity and Decreases Melanogenesis in Human MNT-1 Melanoma Cells. J. Pineal Res. 2019, 67, e12610. [Google Scholar] [CrossRef]
- Reiter, R.J.; Sharma, R.; Ma, Q.; Rorsales-Corral, S.; de Almeida Chuffa, L.G. Melatonin Inhibits Warburg-Dependent Cancer by Redirecting Glucose Oxidation to the Mitochondria: A Mechanistic Hypothesis. Cell. Mol. Life Sci. 2020, 77, 2527–2542. [Google Scholar] [CrossRef]
- Reiter, R.J.; Sharma, R.; Ma, Q. Switching Diseased Cells from Cytosolic Aerobic Glycolysis to Mitochondrial Oxidative Phosphorylation: A Metabolic Rhythm Regulated by Melatonin? J. Pineal Res. 2021, 70, e12677. [Google Scholar] [CrossRef]
- Chok, K.C.; Koh, R.Y.; Ng, M.G.; Ng, P.Y.; Chye, S.M. Melatonin Induces Autophagy via Reactive Oxygen Species-Mediated Endoplasmic Reticulum Stress Pathway in Colorectal Cancer Cells. Molecules 2021, 26, 5038. [Google Scholar] [CrossRef]
- Kleszczyński, K.; Böhm, M. Can Melatonin and Its Metabolites Boost the Efficacy of Targeted Therapy in Patients with Advanced Melanoma? Exp. Dermatol. 2020, 29, 860–863. [Google Scholar] [CrossRef]
- Cheng, X. Structure and Function of DNA Methyltransferases. Annu. Rev. Biophys. Biomol. Struct. 1995, 24, 293–318. [Google Scholar] [CrossRef]
- Illingworth, R.; Kerr, A.; Desousa, D.; Jørgensen, H.; Ellis, P.; Stalker, J.; Jackson, D.; Clee, C.; Plumb, R.; Rogers, J.; et al. A Novel CpG Island Set Identifies Tissue-Specific Methylation at Developmental Gene Loci. PLoS Biol. 2008, 6, e22. [Google Scholar] [CrossRef]
- Hermann, A.; Gowher, H.; Jeltsch, A. Biochemistry and Biology of Mammalian DNA Methyltransferases. Cell. Mol. Life Sci. 2004, 61, 2571–2587. [Google Scholar] [CrossRef]
- Yoder, J.A.; Soman, N.S.; Verdine, G.L.; Bestor, T.H. DNA (Cytosine-5)-Methyltransferases in Mouse Cells and Tissues. Studies with a Mechanism-Based Probe. J. Mol. Biol. 1997, 270, 385–395. [Google Scholar] [CrossRef]
- Okano, M.; Bell, D.W.; Haber, D.A.; Li, E. DNA Methyltransferases Dnmt3a and Dnmt3b Are Essential for de Novo Methylation and Mammalian Development. Cell 1999, 99, 247–257. [Google Scholar] [CrossRef]
- Vinson, C.; Chatterjee, R. CG Methylation. Epigenomics 2012, 4, 655–663. [Google Scholar] [CrossRef]
- Wu, H.; Zhang, Y. Mechanisms and Functions of Tet Protein-Mediated 5-Methylcytosine Oxidation. Genes Dev. 2011, 25, 2436–2452. [Google Scholar] [CrossRef]
- Zhu, H.; Wang, G.; Qian, J. Transcription Factors as Readers and Effectors of DNA Methylation. Nat. Rev. Genet. 2016, 17, 551–565. [Google Scholar] [CrossRef]
- Hendrich, B.; Bird, A. Identification and Characterization of a Family of Mammalian Methyl-CpG Binding Proteins. Mol. Cell. Biol. 1998, 18, 6538–6547. [Google Scholar] [CrossRef]
- Ballestar, E.; Paz, M.F.; Valle, L.; Wei, S.; Fraga, M.F.; Espada, J.; Cruz Cigudosa, J.; Huang, T.H.-M.; Esteller, M. Methyl-CpG Binding Proteins Identify Novel Sites of Epigenetic Inactivation in Human Cancer. EMBO J. 2003, 22, 6335–6345. [Google Scholar] [CrossRef]
- Tahiliani, M.; Koh, K.P.; Shen, Y.; Pastor, W.A.; Bandukwala, H.; Brudno, Y.; Agarwal, S.; Iyer, L.M.; Liu, D.R.; Aravind, L.; et al. Conversion of 5-Methylcytosine to 5-Hydroxymethylcytosine in Mammalian DNA by MLL Partner TET1. Science 2009, 324, 930–935. [Google Scholar] [CrossRef]
- Ono, R.; Taki, T.; Taketani, T.; Taniwaki, M.; Kobayashi, H.; Hayashi, Y. LCX, Leukemia-Associated Protein with a CXXC Domain, Is Fused to MLL in Acute Myeloid Leukemia with Trilineage Dysplasia Having t(10;11)(Q22;Q23). Cancer Res. 2002, 62, 4075–4080. [Google Scholar]
- Lorsbach, R.B.; Moore, J.; Mathew, S.; Raimondi, S.C.; Mukatira, S.T.; Downing, J.R. TET1, a Member of a Novel Protein Family, Is Fused to MLL in Acute Myeloid Leukemia Containing the t(10;11)(Q22;Q23). Leukemia 2003, 17, 637–641. [Google Scholar] [CrossRef]
- Ito, S.; Shen, L.; Dai, Q.; Wu, S.C.; Collins, L.B.; Swenberg, J.A.; He, C.; Zhang, Y. Tet Proteins Can Convert 5-Methylcytosine to 5-Formylcytosine and 5-Carboxylcytosine. Science 2011, 333, 1300–1303. [Google Scholar] [CrossRef]
- He, Y.-F.; Li, B.-Z.; Li, Z.; Liu, P.; Wang, Y.; Tang, Q.; Ding, J.; Jia, Y.; Chen, Z.; Li, L.; et al. Tet-Mediated Formation of 5-Carboxylcytosine and Its Excision by TDG in Mammalian DNA. Science 2011, 333, 1303–1307. [Google Scholar] [CrossRef]
- Maiti, A.; Drohat, A.C. Thymine DNA Glycosylase Can Rapidly Excise 5-Formylcytosine and 5-Carboxylcytosine: Potential Implications for Active Demethylation of CpG Sites. J. Biol. Chem. 2011, 286, 35334–35338. [Google Scholar] [CrossRef]
- Neri, F.; Incarnato, D.; Krepelova, A.; Rapelli, S.; Pagnani, A.; Zecchina, R.; Parlato, C.; Oliviero, S. Genome-Wide Analysis Identifies a Functional Association of Tet1 and Polycomb Repressive Complex 2 in Mouse Embryonic Stem Cells. Genome Biol. 2013, 14, R91. [Google Scholar] [CrossRef]
- Li, Y.; Zheng, H.; Wang, Q.; Zhou, C.; Wei, L.; Liu, X.; Zhang, W.; Zhang, Y.; Du, Z.; Wang, X.; et al. Genome-Wide Analyses Reveal a Role of Polycomb in Promoting Hypomethylation of DNA Methylation Valleys. Genome Biol. 2018, 19, 18. [Google Scholar] [CrossRef]
- Shi, F.; Qiu, J.; Zhang, S.; Zhao, X.; Feng, D.; Feng, X. Exogenous Melatonin Protects Preimplantation Embryo Development from Decabromodiphenyl Ethane-Induced Circadian Rhythm Disorder and Endogenous Melatonin Reduction. Environ. Pollut. 2022, 292, 118445. [Google Scholar] [CrossRef]
- Chandru, A.; Bate, N.; Vuister, G.W.; Cowley, S.M. Sin3A Recruits Tet1 to the PAH1 Domain via a Highly Conserved Sin3-Interaction Domain. Sci. Rep. 2018, 8, 14689. [Google Scholar] [CrossRef]
- Hrit, J.; Goodrich, L.; Li, C.; Wang, B.-A.; Nie, J.; Cui, X.; Martin, E.A.; Simental, E.; Fernandez, J.; Liu, M.Y.; et al. OGT Binds a Conserved C-Terminal Domain of TET1 to Regulate TET1 Activity and Function in Development. eLife 2018, 7, e34870. [Google Scholar] [CrossRef]
- Yuan, J.; Luo, R.Z.; Fujii, S.; Wang, L.; Hu, W.; Andreeff, M.; Pan, Y.; Kadota, M.; Oshimura, M.; Sahin, A.A.; et al. Aberrant Methylation and Silencing of ARHI, an Imprinted Tumor Suppressor Gene in Which the Function Is Lost in Breast Cancers. Cancer Res. 2003, 63, 4174–4180. [Google Scholar] [PubMed]
- Han, L.-L.; Hou, L.; Zhou, M.-J.; Ma, Z.; Lin, D.-L.; Wu, L.; Ge, Y. Aberrant NDRG1 Methylation Associated with Its Decreased Expression and Clinicopathological Significance in Breast Cancer. J. Biomed. Sci. 2013, 20, 52. [Google Scholar] [CrossRef]
- Xiang, T.-X.; Yuan, Y.; Li, L.-L.; Wang, Z.-H.; Dan, L.-Y.; Chen, Y.; Ren, G.-S.; Tao, Q. Aberrant Promoter CpG Methylation and Its Translational Applications in Breast Cancer. Chin. J. Cancer 2013, 32, 12–20. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Huang, Q.; Zeng, F.; Li, W.; He, Z.; Chen, W.; Zhu, W.; Zhang, B. The Prognostic Value of Global DNA Hypomethylation in Cancer: A Meta-Analysis. PLoS ONE 2014, 9, e106290. [Google Scholar] [CrossRef] [PubMed]
- Hinshelwood, R.A.; Clark, S.J. Breast Cancer Epigenetics: Normal Human Mammary Epithelial Cells as a Model System. J. Mol. Med. 2008, 86, 1315–1328. [Google Scholar] [CrossRef]
- Yi, J.; Gao, R.; Chen, Y.; Yang, Z.; Han, P.; Zhang, H.; Dou, Y.; Liu, W.; Wang, W.; Du, G.; et al. Overexpression of NSUN2 by DNA Hypomethylation Is Associated with Metastatic Progression in Human Breast Cancer. Oncotarget 2017, 8, 20751–20765. [Google Scholar] [CrossRef]
- Soares, J.; Pinto, A.E.; Cunha, C.V.; André, S.; Barão, I.; Sousa, J.M.; Cravo, M. Global DNA Hypomethylation in Breast Carcinoma: Correlation with Prognostic Factors and Tumor Progression. Cancer 1999, 85, 112–118. [Google Scholar] [CrossRef]
- Widschwendter, M.; Jiang, G.; Woods, C.; Müller, H.M.; Fiegl, H.; Goebel, G.; Marth, C.; Müller-Holzner, E.; Zeimet, A.G.; Laird, P.W.; et al. DNA Hypomethylation and Ovarian Cancer Biology. Cancer Res. 2004, 64, 4472–4480. [Google Scholar] [CrossRef]
- Yegnasubramanian, S.; Haffner, M.C.; Zhang, Y.; Gurel, B.; Cornish, T.C.; Wu, Z.; Irizarry, R.A.; Morgan, J.; Hicks, J.; DeWeese, T.L.; et al. DNA Hypomethylation Arises Later in Prostate Cancer Progression than CpG Island Hypermethylation and Contributes to Metastatic Tumor Heterogeneity. Cancer Res. 2008, 68, 8954–8967. [Google Scholar] [CrossRef]
- Barciszewska, A.-M. Global DNA Demethylation as an Epigenetic Marker of Human Brain Metastases. Biosci. Rep. 2018, 38, BSR20180731. [Google Scholar] [CrossRef]
- Feinberg, A.P.; Vogelstein, B. Hypomethylation Distinguishes Genes of Some Human Cancers from Their Normal Counterparts. Nature 1983, 301, 89–92. [Google Scholar] [CrossRef] [PubMed]
- Gama-Sosa, M.A.; Slagel, V.A.; Trewyn, R.W.; Oxenhandler, R.; Kuo, K.C.; Gehrke, C.W.; Ehrlich, M. The 5-Methylcytosine Content of DNA from Human Tumors. Nucleic Acids Res. 1983, 11, 6883–6894. [Google Scholar] [CrossRef] [PubMed]
- Haffner, M.C.; Chaux, A.; Meeker, A.K.; Esopi, D.M.; Gerber, J.; Pellakuru, L.G.; Toubaji, A.; Argani, P.; Iacobuzio-Donahue, C.; Nelson, W.G.; et al. Global 5-Hydroxymethylcytosine Content Is Significantly Reduced in Tissue Stem/Progenitor Cell Compartments and in Human Cancers. Oncotarget 2011, 2, 627–637. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Deng, Q.; Fan, W.; Zeng, Q.; He, H.; Huang, F. Melatonin-Induced Suppression of DNA Methylation Promotes Odontogenic Differentiation in Human Dental Pulp Cells. Bioengineered 2020, 11, 829–840. [Google Scholar] [CrossRef]
- Korkmaz, A.; Reiter, R.J. Epigenetic Regulation: A New Research Area for Melatonin? J. Pineal Res. 2008, 44, 41–44. [Google Scholar] [CrossRef] [PubMed]
- López-Moyado, I.F.; Tsagaratou, A.; Yuita, H.; Seo, H.; Delatte, B.; Heinz, S.; Benner, C.; Rao, A. Paradoxical Association of TET Loss of Function with Genome-Wide DNA Hypomethylation. Proc. Natl. Acad. Sci. USA 2019, 116, 16933–16942. [Google Scholar] [CrossRef]
- Yang, M.; Tao, J.; Wu, H.; Guan, S.; Liu, L.; Zhang, L.; Deng, S.; He, C.; Ji, P.; Liu, J.; et al. Aanat Knockdown and Melatonin Supplementation in Embryo Development: Involvement of Mitochondrial Function and DNA Methylation. Antioxid. Redox Signal. 2019, 30, 2050–2065. [Google Scholar] [CrossRef]
- Zeng, J.; Tao, J.; Xia, L.; Zeng, Z.; Chen, J.; Wang, Z.; Meng, J.; Liu, L. Melatonin Inhibits Vascular Endothelial Cell Pyroptosis by Improving Mitochondrial Function via Up-Regulation and Demethylation of UQCRC1. Biochem. Cell Biol. 2021, 99, 339–347. [Google Scholar] [CrossRef]
- Hsieh, M.-C.; Ho, Y.-C.; Lai, C.-Y.; Chou, D.; Wang, H.-H.; Chen, G.-D.; Lin, T.-B.; Peng, H.-Y. Melatonin Impedes Tet1-Dependent MGluR5 Promoter Demethylation to Relieve Pain. J. Pineal Res. 2017, 63, e12436. [Google Scholar] [CrossRef]
- Wang, X.; Sun, M.; Li, J.; Song, X.; He, H.; Huan, Y. Melatonin Protects against Defects Induced by Enniatin B1 during Porcine Early Embryo Development. Aging 2021, 13, 5553–5570. [Google Scholar] [CrossRef]
- Zhao, X.-M.; Wang, N.; Hao, H.-S.; Li, C.-Y.; Zhao, Y.-H.; Yan, C.-L.; Wang, H.-Y.; Du, W.-H.; Wang, D.; Liu, Y.; et al. Melatonin Improves the Fertilization Capacity and Developmental Ability of Bovine Oocytes by Regulating Cytoplasmic Maturation Events. J. Pineal Res. 2018, 64, e12445. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Bashkenova, N.; Zang, R.; Huang, X.; Wang, J. The Roles of TET Family Proteins in Development and Stem Cells. Development 2020, 147, dev183129. [Google Scholar] [CrossRef] [PubMed]
- Putiri, E.L.; Tiedemann, R.L.; Thompson, J.J.; Liu, C.; Ho, T.; Choi, J.-H.; Robertson, K.D. Distinct and Overlapping Control of 5-Methylcytosine and 5-Hydroxymethylcytosine by the TET Proteins in Human Cancer Cells. Genome Biol. 2014, 15, R81. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Gan, L.; Zhang, T.; Ren, Q.; Sun, C. Melatonin Alleviates Adipose Inflammation through Elevating α-Ketoglutarate and Diverting Adipose-Derived Exosomes to Macrophages in Mice. J. Pineal Res. 2018, 64, e12455. [Google Scholar] [CrossRef] [PubMed]
- Clifton, I.J.; McDonough, M.A.; Ehrismann, D.; Kershaw, N.J.; Granatino, N.; Schofield, C.J. Structural Studies on 2-Oxoglutarate Oxygenases and Related Double-Stranded Beta-Helix Fold Proteins. J. Inorg. Biochem. 2006, 100, 644–669. [Google Scholar] [CrossRef]
- McDonough, M.A.; Loenarz, C.; Chowdhury, R.; Clifton, I.J.; Schofield, C.J. Structural Studies on Human 2-Oxoglutarate Dependent Oxygenases. Curr. Opin. Struct. Biol. 2010, 20, 659–672. [Google Scholar] [CrossRef]
- Reiter, R.J.; Rosales-Corral, S.; Coto-Montes, A.; Boga, J.A.; Tan, D.-X.; Davis, J.M.; Konturek, P.C.; Konturek, S.J.; Brzozowski, T. The Photoperiod, Circadian Regulation and Chronodisruption: The Requisite Interplay between the Suprachiasmatic Nuclei and the Pineal and Gut Melatonin. J. Physiol. Pharmacol. 2011, 62, 269–274. [Google Scholar]
- Cajochen, C.; Münch, M.; Kobialka, S.; Kräuchi, K.; Steiner, R.; Oelhafen, P.; Orgül, S.; Wirz-Justice, A. High Sensitivity of Human Melatonin, Alertness, Thermoregulation, and Heart Rate to Short Wavelength Light. J. Clin. Endocrinol. Metab. 2005, 90, 1311–1316. [Google Scholar] [CrossRef]
- Thapan, K.; Arendt, J.; Skene, D.J. An Action Spectrum for Melatonin Suppression: Evidence for a Novel Non-Rod, Non-Cone Photoreceptor System in Humans. J. Physiol. 2001, 535, 261–267. [Google Scholar] [CrossRef]
- Walker, W.H.; Bumgarner, J.R.; Walton, J.C.; Liu, J.A.; Meléndez-Fernández, O.H.; Nelson, R.J.; DeVries, A.C. Light Pollution and Cancer. Int. J. Mol. Sci. 2020, 21, 9360. [Google Scholar] [CrossRef]
- Schwimmer, H.; Metzer, A.; Pilosof, Y.; Szyf, M.; Machnes, Z.M.; Fares, F.; Harel, O.; Haim, A. Light at Night and Melatonin Have Opposite Effects on Breast Cancer Tumors in Mice Assessed by Growth Rates and Global DNA Methylation. Chronobiol. Int. 2014, 31, 144–150. [Google Scholar] [CrossRef] [PubMed]
- Agbaria, S.; Haim, A.; Fares, F.; Zubidat, A.E. Epigenetic Modification in 4T1 Mouse Breast Cancer Model by Artificial Light at Night and Melatonin—The Role of DNA-Methyltransferase. Chronobiol. Int. 2019, 36, 629–643. [Google Scholar] [CrossRef] [PubMed]
- Zubidat, A.E.; Fares, B.; Fares, F.; Haim, A. Artificial Light at Night of Different Spectral Compositions Differentially Affects Tumor Growth in Mice: Interaction with Melatonin and Epigenetic Pathways. Cancer Control 2018, 25, 1073274818812908. [Google Scholar] [CrossRef]
- Yonis, M.; Haim, A.; Zubidat, A.E. Altered Metabolic and Hormonal Responses in Male Rats Exposed to Acute Bright Light-at-Night Associated with Global DNA Hypo-Methylation. J. Photochem. Photobiol. B 2019, 194, 107–118. [Google Scholar] [CrossRef] [PubMed]
- Korkmaz, A.; Sanchez-Barcelo, E.J.; Tan, D.-X.; Reiter, R.J. Role of Melatonin in the Epigenetic Regulation of Breast Cancer. Breast Cancer Res. Treat. 2009, 115, 13–27. [Google Scholar] [CrossRef] [PubMed]
- Korkmaz, A.; Topal, T.; Tan, D.-X.; Reiter, R.J. Role of Melatonin in Metabolic Regulation. Rev. Endocr. Metab. Disord. 2009, 10, 261–270. [Google Scholar] [CrossRef]
- Kim, M.; Vu, T.-H.; Maas, M.B.; Braun, R.I.; Wolf, M.S.; Roenneberg, T.; Daviglus, M.L.; Reid, K.J.; Zee, P.C. Light at Night in Older Age Is Associated with Obesity, Diabetes, and Hypertension. Sleep 2023, 46, zsac130. [Google Scholar] [CrossRef]
- Kim, J.Y.; Jelinek, J.; Lee, Y.H.; Kim, D.H.; Kang, K.; Ryu, S.H.; Moon, H.R.; Cho, K.; Rha, S.H.; Cha, J.K.; et al. Hypomethylation in MTNR1B: A Novel Epigenetic Marker for Atherosclerosis Profiling Using Stenosis Radiophenotype and Blood Inflammatory Cells. Clin. Epigenet. 2023, 15, 11. [Google Scholar] [CrossRef]
- Al-Naggar, R.A.; Anil, S. Artificial Light at Night and Cancer: Global Study. Asian Pac. J. Cancer Prev. 2016, 17, 4661–4664. [Google Scholar] [CrossRef]
- Gehlert, S.; Clanton, M. Shift Work and Breast Cancer. Int. J. Environ. Res. Public Health 2020, 17, 9544. [Google Scholar] [CrossRef]
- Szkiela, M.; Kusideł, E.; Makowiec-Dąbrowska, T.; Kaleta, D. Night Shift Work—A Risk Factor for Breast Cancer. Int. J. Environ. Res. Public Health 2020, 17, 659. [Google Scholar] [CrossRef] [PubMed]
- Ritonja, J.A.; Aronson, K.J.; Leung, M.; Flaten, L.; Topouza, D.G.; Duan, Q.L.; Durocher, F.; Tranmer, J.E.; Bhatti, P. Investigating the Relationship between Melatonin Patterns and Methylation in Circadian Genes among Day Shift and Night Shift Workers. Occup. Environ. Med. 2022, 79, 673–680. [Google Scholar] [CrossRef] [PubMed]
- Shadan, F.F. Circadian Tempo: A Paradigm for Genome Stability? Med. Hypotheses 2006, 68, 883–891. [Google Scholar] [CrossRef] [PubMed]
- Sultan, A.; Parganiha, A.; Sultan, T.; Choudhary, V.; Pati, A.K. Circadian Clock, Cell Cycle, and Breast Cancer: An Updated Review. Biol. Rhythm. Res. 2017, 48, 353–369. [Google Scholar] [CrossRef]
- Doi, M.; Hirayama, J.; Sassone-Corsi, P. Circadian Regulator CLOCK Is a Histone Acetyltransferase. Cell 2006, 125, 497–508. [Google Scholar] [CrossRef]
- Hirayama, J.; Sahar, S.; Grimaldi, B.; Tamaru, T.; Takamatsu, K.; Nakahata, Y.; Sassone-Corsi, P. CLOCK-Mediated Acetylation of BMAL1 Controls Circadian Function. Nature 2007, 450, 1086–1090. [Google Scholar] [CrossRef]
- Huang, C.-H.; Han, W.; Wu, Y.-Z.; Shen, G.-L. Identification of Aberrantly Methylated Differentially Expressed Genes and Pro-Tumorigenic Role of KIF2C in Melanoma. Front. Genet. 2022, 13, 817656. [Google Scholar] [CrossRef]
- Liu, S.; Cheng, Y.; Wang, S.; Liu, H. Circadian Clock Genes Modulate Immune, Cell Cycle and Apoptosis in the Diagnosis and Prognosis of Pan-Renal Cell Carcinoma. Front. Mol. Biosci. 2021, 8, 747629. [Google Scholar] [CrossRef]
- Van der Watt, P.J.; Roden, L.C.; Davis, K.T.; Parker, M.I.; Leaner, V.D. Circadian Oscillations Persist in Cervical and Esophageal Cancer Cells Displaying Decreased Expression of Tumor-Suppressing Circadian Clock Genes. Mol. Cancer Res. 2020, 18, 1340–1353. [Google Scholar] [CrossRef]
- Zhang, Z.; Liang, Z.; Gao, W.; Yu, S.; Hou, Z.; Li, K.; Zeng, P. Identification of Circadian Clock Genes as Regulators of Immune Infiltration in Hepatocellular Carcinoma. J. Cancer 2022, 13, 3199–3208. [Google Scholar] [CrossRef]
- Reszka, E.; Wieczorek, E.; Przybek, M.; Jabłońska, E.; Kałużny, P.; Bukowska-Damska, A.; Zienolddiny, S.; Pepłońska, B. Circadian Gene Methylation in Rotating-Shift Nurses: A Cross-Sectional Study. Chronobiol. Int. 2018, 35, 111–121. [Google Scholar] [CrossRef] [PubMed]
- Dijk, D.-J.; Archer, S.N. PERIOD3, Circadian Phenotypes, and Sleep Homeostasis. Sleep Med. Rev. 2010, 14, 151–160. [Google Scholar] [CrossRef] [PubMed]
- Sulkava, S.; Ollila, H.M.; Alasaari, J.; Puttonen, S.; Härmä, M.; Viitasalo, K.; Lahtinen, A.; Lindström, J.; Toivola, A.; Sulkava, R.; et al. Common Genetic Variation Near Melatonin Receptor 1A Gene Linked to Job-Related Exhaustion in Shift Workers. Sleep 2017, 40, zsw011. [Google Scholar] [CrossRef] [PubMed]
- Stevenson, T.J.; Prendergast, B.J. Reversible DNA Methylation Regulates Seasonal Photoperiodic Time Measurement. Proc. Natl. Acad. Sci. USA 2013, 110, 16651–16656. [Google Scholar] [CrossRef] [PubMed]
- Lan, M.; Han, J.; Pan, M.-H.; Wan, X.; Pan, Z.-N.; Sun, S.-C. Melatonin Protects against Defects Induced by Deoxynivalenol during Mouse Oocyte Maturation. J. Pineal Res. 2018, 65, e12477. [Google Scholar] [CrossRef]
- Qu, J.; Sun, M.; Wang, X.; Song, X.; He, H.; Huan, Y. Melatonin Enhances the Development of Porcine Cloned Embryos by Improving DNA Methylation Reprogramming. Cell Reprogram. 2020, 22, 156–166. [Google Scholar] [CrossRef]
- Sørensen, A.L.; Timoskainen, S.; West, F.D.; Vekterud, K.; Boquest, A.C.; Ahrlund-Richter, L.; Stice, S.L.; Collas, P. Lineage-Specific Promoter DNA Methylation Patterns Segregate Adult Progenitor Cell Types. Stem. Cells Dev. 2010, 19, 1257–1266. [Google Scholar] [CrossRef]
- Berdasco, M.; Melguizo, C.; Prados, J.; Gómez, A.; Alaminos, M.; Pujana, M.A.; Lopez, M.; Setien, F.; Ortiz, R.; Zafra, I.; et al. DNA Methylation Plasticity of Human Adipose-Derived Stem Cells in Lineage Commitment. Am. J. Pathol. 2012, 181, 2079–2093. [Google Scholar] [CrossRef]
- Gibney, E.R.; Nolan, C.M. Epigenetics and Gene Expression. Heredity 2010, 105, 4–13. [Google Scholar] [CrossRef]
- Rahman, M.M.; Brane, A.C.; Tollefsbol, T.O. MicroRNAs and Epigenetics Strategies to Reverse Breast Cancer. Cells 2019, 8, 1214. [Google Scholar] [CrossRef]
- Tompkins, J.D.; Hall, C.; Chen, V.C.; Li, A.X.; Wu, X.; Hsu, D.; Couture, L.A.; Riggs, A.D. Epigenetic Stability, Adaptability, and Reversibility in Human Embryonic Stem Cells. Proc. Natl. Acad. Sci. USA 2012, 109, 12544–12549. [Google Scholar] [CrossRef] [PubMed]
- Kaur, J.; Daoud, A.; Eblen, S.T. Targeting Chromatin Remodeling for Cancer Therapy. Curr. Mol. Pharmacol. 2019, 12, 215–229. [Google Scholar] [CrossRef] [PubMed]
- Kim, A.; Mo, K.; Kwon, H.; Choe, S.; Park, M.; Kwak, W.; Yoon, H. Epigenetic Regulation in Breast Cancer: Insights on Epidrugs. Epigenomes 2023, 7, 6. [Google Scholar] [CrossRef] [PubMed]
- Jones, P.A.; Taylor, S.M. Cellular Differentiation, Cytidine Analogs and DNA Methylation. Cell 1980, 20, 85–93. [Google Scholar] [CrossRef] [PubMed]
- Stresemann, C.; Lyko, F. Modes of Action of the DNA Methyltransferase Inhibitors Azacytidine and Decitabine. Int. J. Cancer 2008, 123, 8–13. [Google Scholar] [CrossRef] [PubMed]
- Schermelleh, L.; Spada, F.; Easwaran, H.P.; Zolghadr, K.; Margot, J.B.; Cardoso, M.C.; Leonhardt, H. Trapped in Action: Direct Visualization of DNA Methyltransferase Activity in Living Cells. Nat. Methods 2005, 2, 751–756. [Google Scholar] [CrossRef]
- Gnyszka, A.; Jastrzebski, Z.; Flis, S. DNA Methyltransferase Inhibitors and Their Emerging Role in Epigenetic Therapy of Cancer. Anticancer Res. 2013, 33, 2989–2996. [Google Scholar]
- Fenaux, P.; Mufti, G.J.; Hellstrom-Lindberg, E.; Santini, V.; Finelli, C.; Giagounidis, A.; Schoch, R.; Gattermann, N.; Sanz, G.; List, A.; et al. Efficacy of Azacitidine Compared with That of Conventional Care Regimens in the Treatment of Higher-Risk Myelodysplastic Syndromes: A Randomised, Open-Label, Phase III Study. Lancet Oncol. 2009, 10, 223–232. [Google Scholar] [CrossRef]
- Seymour, J.F.; Döhner, H.; Butrym, A.; Wierzbowska, A.; Selleslag, D.; Jang, J.H.; Kumar, R.; Cavenagh, J.; Schuh, A.C.; Candoni, A.; et al. Azacitidine Improves Clinical Outcomes in Older Patients with Acute Myeloid Leukaemia with Myelodysplasia-Related Changes Compared with Conventional Care Regimens. BMC Cancer 2017, 17, 852. [Google Scholar] [CrossRef]
- Platzbecker, U.; Middeke, J.M.; Sockel, K.; Herbst, R.; Wolf, D.; Baldus, C.D.; Oelschlägel, U.; Mütherig, A.; Fransecky, L.; Noppeney, R.; et al. Measurable Residual Disease-Guided Treatment with Azacitidine to Prevent Haematological Relapse in Patients with Myelodysplastic Syndrome and Acute Myeloid Leukaemia (RELAZA2): An Open-Label, Multicentre, Phase 2 Trial. Lancet Oncol. 2018, 19, 1668–1679. [Google Scholar] [CrossRef]
- Seymour, J.F.; Fenaux, P.; Silverman, L.R.; Mufti, G.J.; Hellström-Lindberg, E.; Santini, V.; List, A.F.; Gore, S.D.; Backstrom, J.; McKenzie, D.; et al. Effects of Azacitidine Compared with Conventional Care Regimens in Elderly (≥75 Years) Patients with Higher-Risk Myelodysplastic Syndromes. Crit. Rev. Oncol. Hematol. 2010, 76, 218–227. [Google Scholar] [CrossRef] [PubMed]
- Fenaux, P.; Mufti, G.J.; Hellström-Lindberg, E.; Santini, V.; Gattermann, N.; Germing, U.; Sanz, G.; List, A.F.; Gore, S.; Seymour, J.F.; et al. Azacitidine Prolongs Overall Survival Compared with Conventional Care Regimens in Elderly Patients with Low Bone Marrow Blast Count Acute Myeloid Leukemia. J. Clin. Oncol. 2010, 28, 562–569. [Google Scholar] [CrossRef] [PubMed]
- Kantarjian, H.M.; O’Brien, S.; Shan, J.; Aribi, A.; Garcia-Manero, G.; Jabbour, E.; Ravandi, F.; Cortes, J.; Davisson, J.; Issa, J.-P. Update of the Decitabine Experience in Higher Risk Myelodysplastic Syndrome and Analysis of Prognostic Factors Associated with Outcome. Cancer 2007, 109, 265–273. [Google Scholar] [CrossRef] [PubMed]
- Jabbour, E.; Garcia-Manero, G.; Ravandi, F.; Faderl, S.; O’Brien, S.; Fullmer, A.; Cortes, J.E.; Wierda, W.; Kantarjian, H. Prognostic Factors Associated with Disease Progression and Overall Survival in Patients with Myelodysplastic Syndromes Treated with Decitabine. Clin. Lymphoma Myeloma Leuk. 2013, 13, 131–138. [Google Scholar] [CrossRef]
- Ramos, M.-P.; Wijetunga, N.A.; McLellan, A.S.; Suzuki, M.; Greally, J.M. DNA Demethylation by 5-Aza-2′-Deoxycytidine Is Imprinted, Targeted to Euchromatin, and Has Limited Transcriptional Consequences. Epigenetics Chromatin 2015, 8, 11. [Google Scholar] [CrossRef]
- Giri, A.K.; Aittokallio, T. DNMT Inhibitors Increase Methylation in the Cancer Genome. Front. Pharmacol. 2019, 10, 385. [Google Scholar] [CrossRef]
- Cabezón, M.; Malinverni, R.; Bargay, J.; Xicoy, B.; Marcé, S.; Garrido, A.; Tormo, M.; Arenillas, L.; Coll, R.; Borras, J.; et al. Different Methylation Signatures at Diagnosis in Patients with High-Risk Myelodysplastic Syndromes and Secondary Acute Myeloid Leukemia Predict Azacitidine Response and Longer Survival. Clin. Epigenet. 2021, 13, 9. [Google Scholar] [CrossRef]
- Notari, R.E.; DeYoung, J.L. Kinetics and Mechanisms of Degradation of the Antileukemic Agent 5-Azacytidine in Aqueous Solutions. J. Pharm Sci. 1975, 64, 1148–1157. [Google Scholar] [CrossRef]
- Rogstad, D.K.; Herring, J.L.; Theruvathu, J.A.; Burdzy, A.; Perry, C.C.; Neidigh, J.W.; Sowers, L.C. The Chemical Decomposition of 5-Aza-2′-Deoxycytidine (Decitabine): Kinetic Analyses and Identification of Products by NMR, HPLC, and Mass Spectrometry. Chem. Res. Toxicol. 2009, 22, 1194–1204. [Google Scholar] [CrossRef]
- Agrawal, K.; Das, V.; Vyas, P.; Hajdúch, M. Nucleosidic DNA Demethylating Epigenetic Drugs—A Comprehensive Review from Discovery to Clinic. Pharmacol. Ther. 2018, 188, 45–79. [Google Scholar] [CrossRef]
- Issa, J.-P.J.; Garcia-Manero, G.; Giles, F.J.; Mannari, R.; Thomas, D.; Faderl, S.; Bayar, E.; Lyons, J.; Rosenfeld, C.S.; Cortes, J.; et al. Phase 1 Study of Low-Dose Prolonged Exposure Schedules of the Hypomethylating Agent 5-Aza-2′-Deoxycytidine (Decitabine) in Hematopoietic Malignancies. Blood 2004, 103, 1635–1640. [Google Scholar] [CrossRef] [PubMed]
- Schnekenburger, M.; Grandjenette, C.; Ghelfi, J.; Karius, T.; Foliguet, B.; Dicato, M.; Diederich, M. Sustained Exposure to the DNA Demethylating Agent, 2′-Deoxy-5-Azacytidine, Leads to Apoptotic Cell Death in Chronic Myeloid Leukemia by Promoting Differentiation, Senescence, and Autophagy. Biochem. Pharmacol. 2011, 81, 364–378. [Google Scholar] [CrossRef] [PubMed]
- Hollenbach, P.W.; Nguyen, A.N.; Brady, H.; Williams, M.; Ning, Y.; Richard, N.; Krushel, L.; Aukerman, S.L.; Heise, C.; MacBeth, K.J. A Comparison of Azacitidine and Decitabine Activities in Acute Myeloid Leukemia Cell Lines. PLoS ONE 2010, 5, e9001. [Google Scholar] [CrossRef] [PubMed]
- Zhao, M.; Rudek, M.A.; He, P.; Hartke, C.; Gore, S.; Carducci, M.A.; Baker, S.D. Quantification of 5-Azacytidine in Plasma by Electrospray Tandem Mass Spectrometry Coupled with High-Performance Liquid Chromatography. J. Chromatogr. B Analyt. Technol. Biomed. Life Sci. 2004, 813, 81–88. [Google Scholar] [CrossRef]
- Liu, Z.; Marcucci, G.; Byrd, J.C.; Grever, M.; Xiao, J.; Chan, K.K. Characterization of Decomposition Products and Preclinical and Low Dose Clinical Pharmacokinetics of Decitabine (5-Aza-2′-Deoxycytidine) by a New Liquid Chromatography/Tandem Mass Spectrometry Quantification Method. Rapid Commun. Mass Spectrom. 2006, 20, 1117–1126. [Google Scholar] [CrossRef]
- Gros, C.; Fahy, J.; Halby, L.; Dufau, I.; Erdmann, A.; Gregoire, J.-M.; Ausseil, F.; Vispé, S.; Arimondo, P.B. DNA Methylation Inhibitors in Cancer: Recent and Future Approaches. Biochimie 2012, 94, 2280–2296. [Google Scholar] [CrossRef]
- Bohl, S.R.; Bullinger, L.; Rücker, F.G. Epigenetic Therapy: Azacytidine and Decitabine in Acute Myeloid Leukemia. Expert Rev. Hematol. 2018, 11, 361–371. [Google Scholar] [CrossRef]
- Hu, C.; Liu, X.; Zeng, Y.; Liu, J.; Wu, F. DNA Methyltransferase Inhibitors Combination Therapy for the Treatment of Solid Tumor: Mechanism and Clinical Application. Clin. Epigenet. 2021, 13, 166. [Google Scholar] [CrossRef]
- Seely, D.; Wu, P.; Fritz, H.; Kennedy, D.A.; Tsui, T.; Seely, A.J.E.; Mills, E. Melatonin as Adjuvant Cancer Care with and without Chemotherapy: A Systematic Review and Meta-Analysis of Randomized Trials. Integr. Cancer Ther. 2012, 11, 293–303. [Google Scholar] [CrossRef]
- Pariente, R.; Bejarano, I.; Rodríguez, A.B.; Pariente, J.A.; Espino, J. Melatonin Increases the Effect of 5-Fluorouracil-Based Chemotherapy in Human Colorectal Adenocarcinoma Cells in Vitro. Mol. Cell Biochem. 2018, 440, 43–51. [Google Scholar] [CrossRef]
- Mihanfar, A.; Yousefi, B.; Ghazizadeh Darband, S.; Sadighparvar, S.; Kaviani, M.; Majidinia, M. Melatonin Increases 5-Flurouracil-Mediated Apoptosis of Colorectal Cancer Cells through Enhancing Oxidative Stress and Downregulating Survivin and XIAP. Bioimpacts 2021, 11, 253–261. [Google Scholar] [CrossRef] [PubMed]
- Gao, Y.; Xiao, X.; Zhang, C.; Yu, W.; Guo, W.; Zhang, Z.; Li, Z.; Feng, X.; Hao, J.; Zhang, K.; et al. Melatonin Synergizes the Chemotherapeutic Effect of 5-Fluorouracil in Colon Cancer by Suppressing PI3K/AKT and NF-ΚB/INOS Signaling Pathways. J. Pineal Res. 2017, 62, e12380. [Google Scholar] [CrossRef] [PubMed]
- Liu, D.; Ma, Z.; Di, S.; Yang, Y.; Yang, J.; Xu, L.; Reiter, R.J.; Qiao, S.; Yuan, J. AMPK/PGC1α Activation by Melatonin Attenuates Acute Doxorubicin Cardiotoxicity via Alleviating Mitochondrial Oxidative Damage and Apoptosis. Free Radic. Biol. Med. 2018, 129, 59–72. [Google Scholar] [CrossRef]
- Niu, G.; Yousefi, B.; Qujeq, D.; Marjani, A.; Asadi, J.; Wang, Z.; Mir, S.M. Melatonin and Doxorubicin Co-Delivered via a Functionalized Graphene-Dendrimeric System Enhances Apoptosis of Osteosarcoma Cells. Mater. Sci. Eng. C Mater. Biol. Appl. 2021, 119, 111554. [Google Scholar] [CrossRef] [PubMed]
- Erdogan, C.S.; Al Hassadi, Y.; Aru, B.; Yılmaz, B.; Gemici, B. Combinatorial Effects of Melatonin and Paclitaxel Differ Depending on the Treatment Scheme in Colorectal Cancer in Vitro. Life Sci. 2022, 308, 120927. [Google Scholar] [CrossRef] [PubMed]
- Rohilla, S.; Singh, M.; Priya, S.; Almalki, W.H.; Haniffa, S.M.; Subramaniyan, V.; Fuloria, S.; Fuloria, N.K.; Sekar, M.; Singh, S.K.; et al. Exploring the Mechanical Perspective of a New Anti-Tumor Agent: Melatonin. J. Environ. Pathol. Toxicol. Oncol. 2023, 42, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Pathipaka, R.; Thyagarajan, A.; Sahu, R.P. Melatonin as a Repurposed Drug for Melanoma Treatment. Med. Sci. 2023, 11, 9. [Google Scholar] [CrossRef] [PubMed]
- Hartung, E.E.; Mukhtar, S.Z.; Shah, S.M.; Niles, L.P. 5-Azacytidine Upregulates Melatonin MT1 Receptor Expression in Rat C6 Glioma Cells: Oncostatic Implications. Mol. Biol. Rep. 2020, 47, 4867–4873. [Google Scholar] [CrossRef]
- Nakamura, E.; Kozaki, K.; Tsuda, H.; Suzuki, E.; Pimkhaokham, A.; Yamamoto, G.; Irie, T.; Tachikawa, T.; Amagasa, T.; Inazawa, J.; et al. Frequent Silencing of a Putative Tumor Suppressor Gene Melatonin Receptor 1 A (MTNR1A) in Oral Squamous-Cell Carcinoma. Cancer Sci. 2008, 99, 1390–1400. [Google Scholar] [CrossRef]
- Oprea-Ilies, G.; Haus, E.; Sackett-Lundeen, L.; Liu, Y.; McLendon, L.; Busch, R.; Adams, A.; Cohen, C. Expression of Melatonin Receptors in Triple Negative Breast Cancer (TNBC) in African American and Caucasian Women: Relation to Survival. Breast Cancer Res. Treat. 2013, 137, 677–687. [Google Scholar] [CrossRef]
- Goyal, R.; Gupta, T.; Bal, A.; Sahni, D.; Singh, G. Role of Melatonin in Breast Carcinoma: Correlation of Expression Patterns of Melatonin-1 Receptor with Estrogen, Progesterone, and HER2 Receptors. Appl. Immunohistochem. Mol. Morphol. 2020, 28, 518–523. [Google Scholar] [CrossRef] [PubMed]
- Dauchy, R.T.; Blask, D.E.; Dauchy, E.M.; Davidson, L.K.; Tirrell, P.C.; Greene, M.W.; Tirrell, R.P.; Hill, C.R.; Sauer, L.A. Antineoplastic Effects of Melatonin on a Rare Malignancy of Mesenchymal Origin: Melatonin Receptor-Mediated Inhibition of Signal Transduction, Linoleic Acid Metabolism and Growth in Tissue-Isolated Human Leiomyosarcoma Xenografts. J. Pineal Res. 2009, 47, 32–42. [Google Scholar] [CrossRef] [PubMed]
- Xiang, S.; Dauchy, R.T.; Hoffman, A.E.; Pointer, D.; Frasch, T.; Blask, D.E.; Hill, S.M. Epigenetic Inhibition of the Tumor Suppressor ARHI by Light at Night-Induced Circadian Melatonin Disruption Mediates STAT3-Driven Paclitaxel Resistance in Breast Cancer. J. Pineal Res. 2019, 67, e12586. [Google Scholar] [CrossRef] [PubMed]
- Hill, S.M.; Cheng, C.; Yuan, L.; Mao, L.; Jockers, R.; Dauchy, B.; Blask, D.E. Age-Related Decline in Melatonin and Its MT1 Receptor Are Associated with Decreased Sensitivity to Melatonin and Enhanced Mammary Tumor Growth. Curr. Aging Sci. 2013, 6, 125–133. [Google Scholar] [CrossRef]
- Lesicka, M.; Dmitrzak-Weglarz, M.; Jablonska, E.; Wieczorek, E.; Kapelski, P.; Szczepankiewicz, A.; Pawlak, J.; Reszka, E. Methylation of Melatonin Receptors in Patients with Unipolar and Bipolar Depression. Mech. Ageing Dev. 2023, 211, 111776. [Google Scholar] [CrossRef]
- Lee, C.-C.; Kuo, Y.-C.; Hu, J.-M.; Chang, P.-K.; Sun, C.-A.; Yang, T.; Li, C.-W.; Chen, C.-Y.; Lin, F.-H.; Hsu, C.-H.; et al. MTNR1B Polymorphisms with CDKN2A and MGMT Methylation Status Are Associated with Poor Prognosis of Colorectal Cancer in Taiwan. World J. Gastroenterol. 2021, 27, 5737–5752. [Google Scholar] [CrossRef]
- Lee, S.E.; Kim, S.J.; Yoon, H.-J.; Yu, S.Y.; Yang, H.; Jeong, S.I.; Hwang, S.Y.; Park, C.-S.; Park, Y.S. Genome-Wide Profiling in Melatonin-Exposed Human Breast Cancer Cell Lines Identifies Differentially Methylated Genes Involved in the Anticancer Effect of Melatonin. J. Pineal Res. 2013, 54, 80–88. [Google Scholar] [CrossRef]
- Martín, V.; Sanchez-Sanchez, A.M.; Herrera, F.; Gomez-Manzano, C.; Fueyo, J.; Alvarez-Vega, M.A.; Antolín, I.; Rodriguez, C. Melatonin-Induced Methylation of the ABCG2/BCRP Promoter as a Novel Mechanism to Overcome Multidrug Resistance in Brain Tumour Stem Cells. Br. J. Cancer 2013, 108, 2005–2012. [Google Scholar] [CrossRef]
- Nie, J.; Xiao, P.; Wang, X.; Yang, X.; Xu, H.; Lu, K.; Lu, S.; Liang, X. Melatonin Prevents Deterioration in Quality by Preserving Epigenetic Modifications of Porcine Oocytes after Prolonged Culture. Aging 2018, 10, 3897–3909. [Google Scholar] [CrossRef]
- Ritonja, J.A.; Aronson, K.J.; Flaten, L.; Topouza, D.G.; Duan, Q.L.; Durocher, F.; Tranmer, J.E.; Bhatti, P. Exploring the Impact of Night Shift Work on Methylation of Circadian Genes. Epigenetics 2022, 17, 1259–1268. [Google Scholar] [CrossRef]
- Villicaña, S.; Bell, J.T. Genetic Impacts on DNA Methylation: Research Findings and Future Perspectives. Genome Biol. 2021, 22, 127. [Google Scholar] [CrossRef] [PubMed]
- Miyakuni, K.; Nishida, J.; Koinuma, D.; Nagae, G.; Aburatani, H.; Miyazono, K.; Ehata, S. Genome-Wide Analysis of DNA Methylation Identifies the Apoptosis-Related Gene UQCRH as a Tumor Suppressor in Renal Cancer. Mol. Oncol. 2022, 16, 732–749. [Google Scholar] [CrossRef]
- Ding, K.; Jiang, J.; Chen, L.; Xu, X. Methylenetetrahydrofolate Dehydrogenase 1 Silencing Expedites the Apoptosis of Non-Small Cell Lung Cancer Cells via Modulating DNA Methylation. Med. Sci. Monit. 2018, 24, 7499–7507. [Google Scholar] [CrossRef] [PubMed]
- Dos Santos, P.W.d.S.; Machado, A.R.T.; De Grandis, R.A.; Ribeiro, D.L.; Tuttis, K.; Morselli, M.; Aissa, A.F.; Pellegrini, M.; Antunes, L.M.G. Transcriptome and DNA Methylation Changes Modulated by Sulforaphane Induce Cell Cycle Arrest, Apoptosis, DNA Damage, and Suppression of Proliferation in Human Liver Cancer Cells. Food Chem. Toxicol. 2020, 136, 111047. [Google Scholar] [CrossRef]
- Bernardini, L.; Barbosa, E.; Charão, M.F.; Goethel, G.; Muller, D.; Bau, C.; Steffens, N.A.; Santos Stein, C.; Moresco, R.N.; Garcia, S.C.; et al. Oxidative Damage, Inflammation, Genotoxic Effect, and Global DNA Methylation Caused by Inhalation of Formaldehyde and the Purpose of Melatonin. Toxicol. Res. 2020, 9, 778–789. [Google Scholar] [CrossRef]
- Fang, Y.; Zhang, J.; Li, Y.; Guo, X.; Li, J.; Zhong, R.; Zhang, X. Melatonin-Induced Demethylation of Antioxidant Genes Increases Antioxidant Capacity through RORα in Cumulus Cells of Prepubertal Lambs. Free Radic Biol. Med. 2019, 131, 173–183. [Google Scholar] [CrossRef] [PubMed]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Linowiecka, K.; Slominski, A.T.; Reiter, R.J.; Böhm, M.; Steinbrink, K.; Paus, R.; Kleszczyński, K. Melatonin: A Potential Regulator of DNA Methylation. Antioxidants 2023, 12, 1155. https://doi.org/10.3390/antiox12061155
Linowiecka K, Slominski AT, Reiter RJ, Böhm M, Steinbrink K, Paus R, Kleszczyński K. Melatonin: A Potential Regulator of DNA Methylation. Antioxidants. 2023; 12(6):1155. https://doi.org/10.3390/antiox12061155
Chicago/Turabian StyleLinowiecka, Kinga, Andrzej T. Slominski, Russel J. Reiter, Markus Böhm, Kerstin Steinbrink, Ralf Paus, and Konrad Kleszczyński. 2023. "Melatonin: A Potential Regulator of DNA Methylation" Antioxidants 12, no. 6: 1155. https://doi.org/10.3390/antiox12061155
APA StyleLinowiecka, K., Slominski, A. T., Reiter, R. J., Böhm, M., Steinbrink, K., Paus, R., & Kleszczyński, K. (2023). Melatonin: A Potential Regulator of DNA Methylation. Antioxidants, 12(6), 1155. https://doi.org/10.3390/antiox12061155