
Metabolomic Analysis Reveals Domestication-Driven Reshaping of Polyphenolic Antioxidants in Soybean Seeds
Abstract
1. Introduction
2. Materials and Methods
2.1. Reagents and Plant Materials
2.2. Metabolite Extraction
2.3. UHPLC-MS/MS Analysis
2.4. Mass Spectrometric Data Processing
2.5. Structural Annotation and Identification of Metabolites
2.6. Assessment of Total Antioxidant Capacity
2.6.1. The FRAP Assay
2.6.2. The ABTS Assay
2.7. Statistical Analysis
3. Results
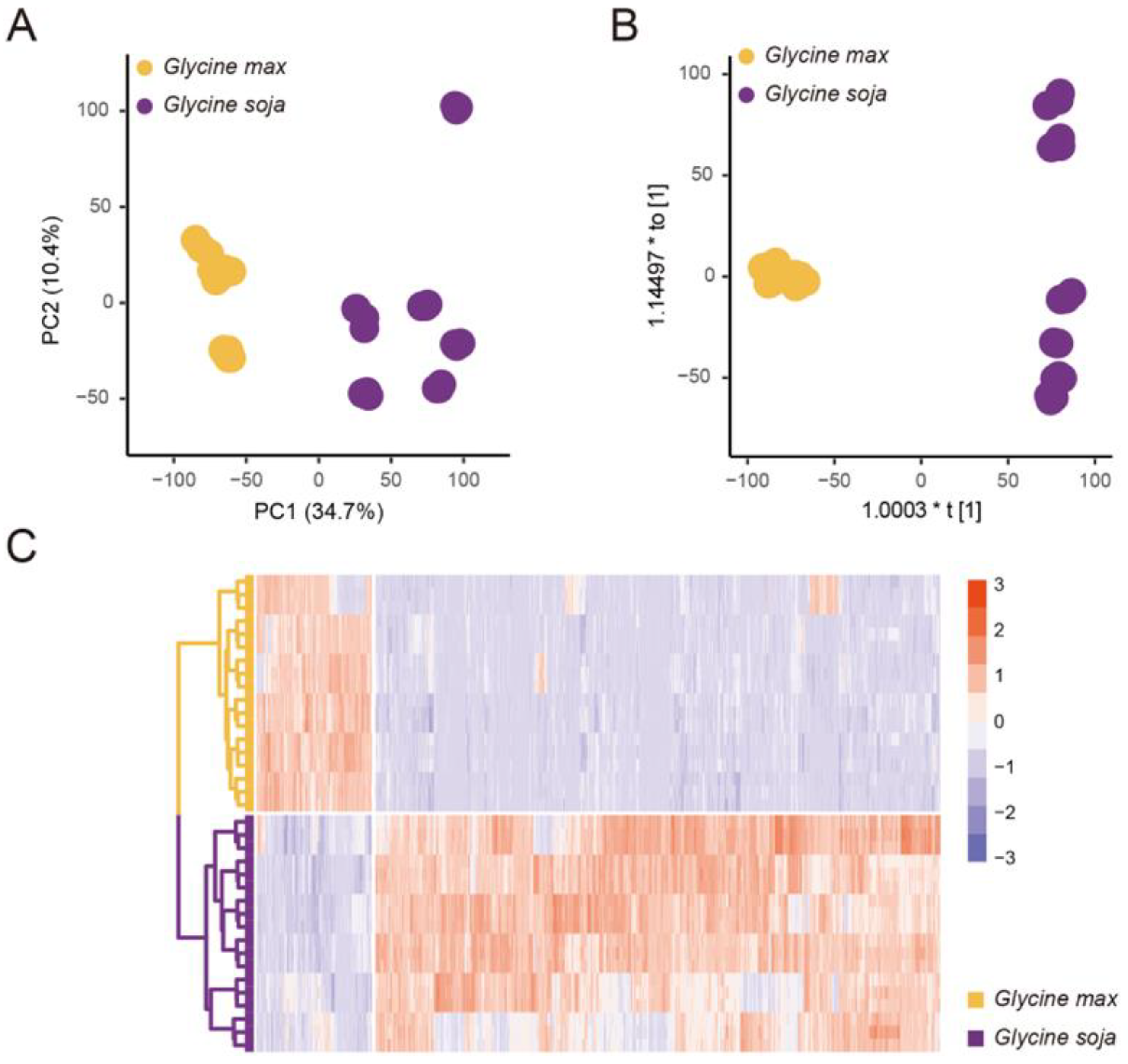
3.1. Comparative Analysis Revealed the Metabolome Variances between Wild and Cultivated Soybeans
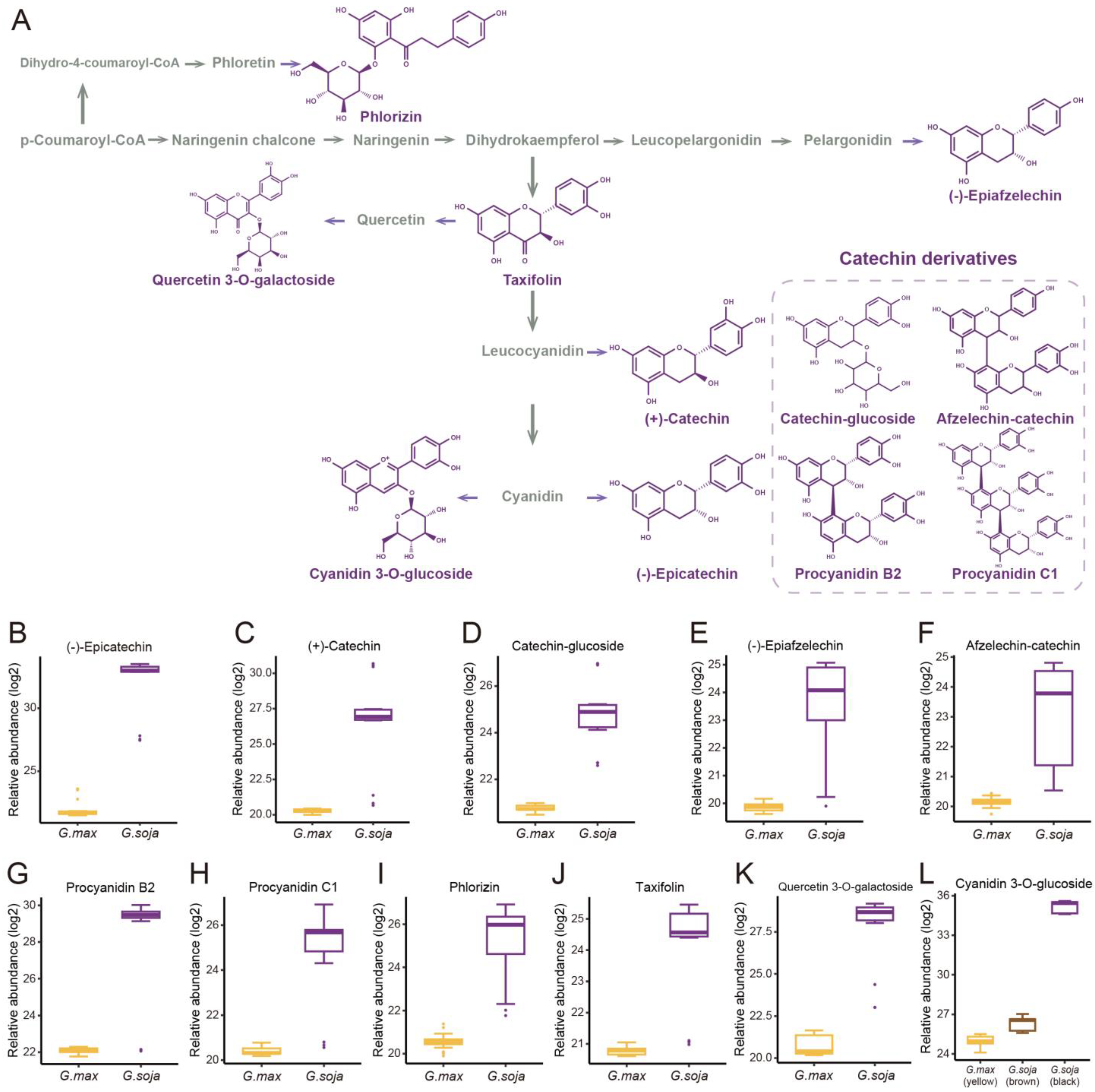
3.2. Dissection of the Key Featured Metabolites of Soybean Domestication
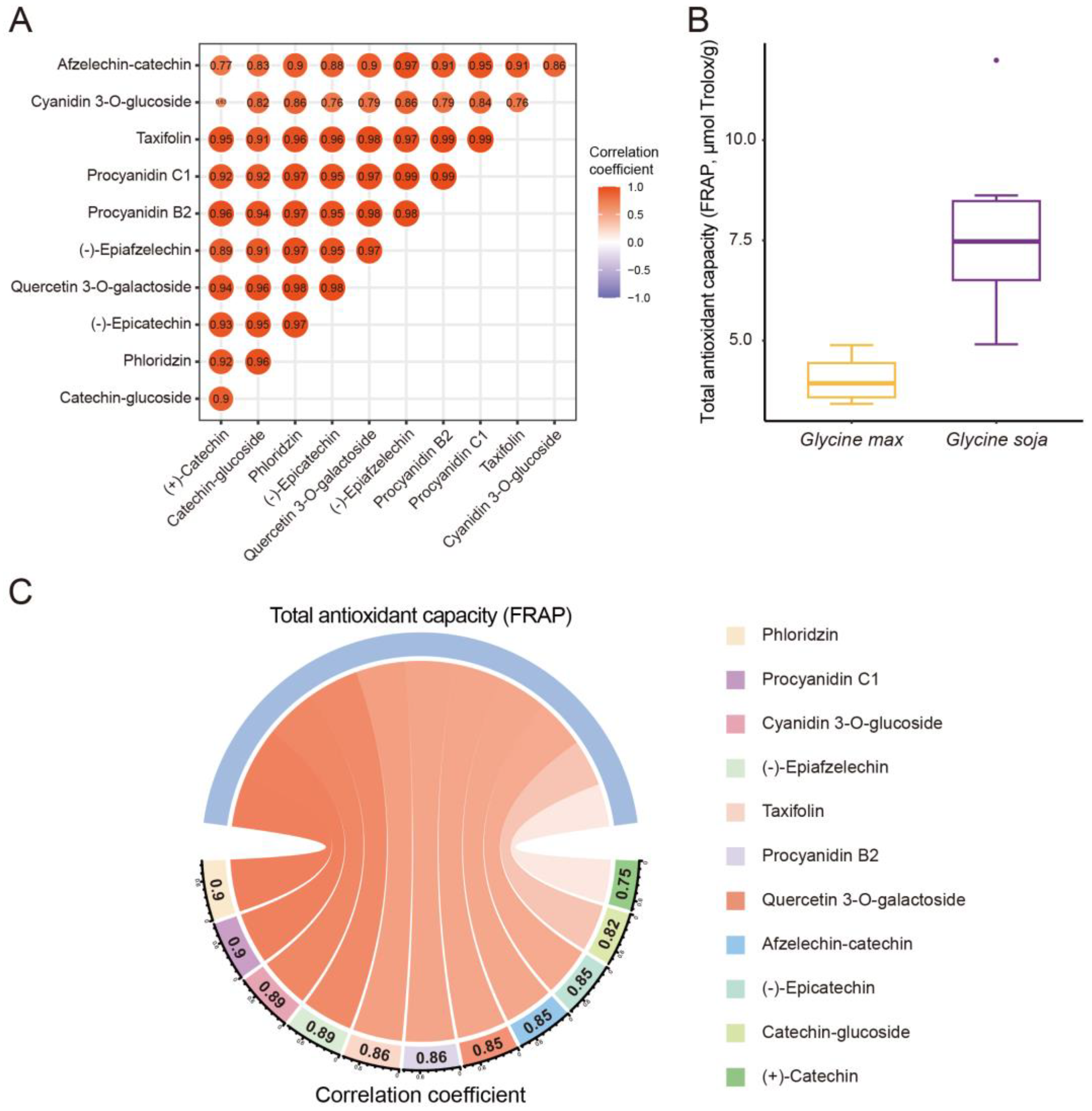
3.3. Correlation Analysis between Domestication-Related Polyphenols and Antioxidant Activities of Soybeans
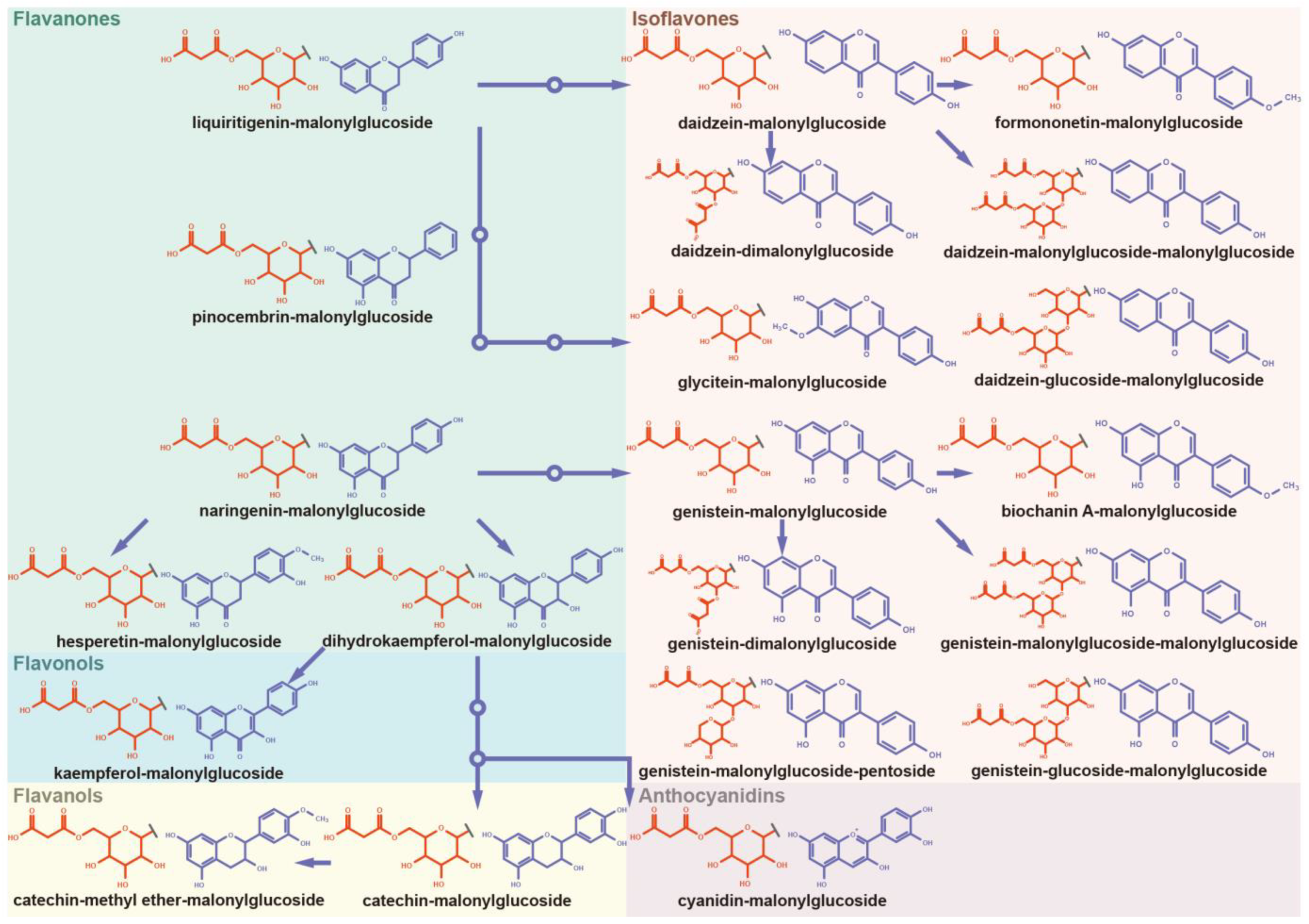
3.4. Panoramic Malonylation Modificome of Flavonoids and Isoflavonoids
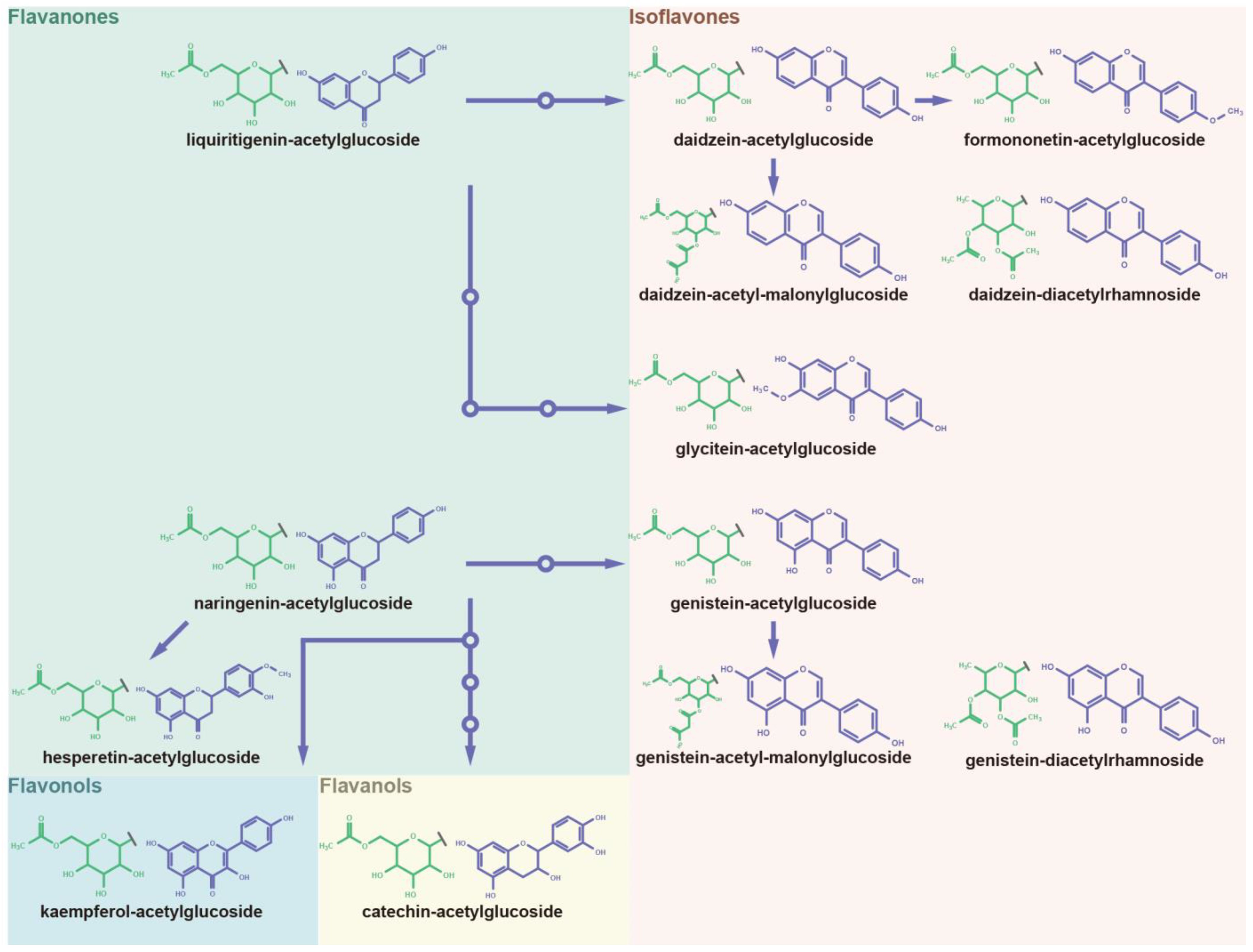
3.5. Panoramic Acetylation Modificome of Flavonoids and Isoflavonoids
4. Discussion
4.1. Domestication-Related Polyphenols, the Promising Targets for Nutrition Fortification
4.2. Panoramic Insights into Malonylome and Acetylome of Polyphenols
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Scharff, L.B.; Saltenis, V.L.R.; Jensen, P.E.; Baekelandt, A.; Burgess, A.J.; Burow, M.; Ceriotti, A.; Cohan, J.P.; Geu-Flores, F.; Halkier, B.A.; et al. Prospects to improve the nutritional quality of crops. Food Energy Secur. 2021, 11, e327. [Google Scholar] [CrossRef]
- Hirschi, K.D. Nutrient biofortification of food crops. Annu. Rev. Nutr. 2009, 29, 401–421. [Google Scholar] [CrossRef] [PubMed]
- Horvat, D.; Šimić, G.; Drezner, G.; Lalić, A.; Ledenčan, T.; Tucak, M.; Plavšić, H.; Andrić, L.; Zdunić, Z. Phenolic Acid Profiles and Antioxidant Activity of Major Cereal Crops. Antioxidants 2020, 9, 527. [Google Scholar] [CrossRef] [PubMed]
- Asensi-Fabado, M.A.; Munne-Bosch, S. Vitamins in plants: Occurrence, biosynthesis and antioxidant function. Trends Plant Sci. 2010, 15, 582–592. [Google Scholar] [CrossRef] [PubMed]
- Vezza, T.; Canet, F.; de Marañón, A.M.; Bañuls, C.; Rocha, M.; Víctor, V.M. Phytosterols: Nutritional Health Players in the Management of Obesity and Its Related Disorders. Antioxidants 2020, 9, 1266. [Google Scholar] [CrossRef]
- Yan, Z.; Zhong, Y.; Duan, Y.; Chen, Q.; Li, F. Antioxidant mechanism of tea polyphenols and its impact on health benefits. Anim. Nutr. 2020, 6, 115–123. [Google Scholar] [CrossRef]
- Alseekh, S.; Scossa, F.; Wen, W.; Luo, J.; Yan, J.; Beleggia, R.; Klee, H.J.; Huang, S.; Papa, R.; Fernie, A.R. Domestication of Crop Metabolomes: Desired and Unintended Consequences. Trends Plant Sci. 2021, 26, 650–661. [Google Scholar] [CrossRef]
- Isanga, J.; Zhang, G.-N. Soybean Bioactive Components and their Implications to Health—A Review. Food Rev. Int. 2008, 24, 252–276. [Google Scholar] [CrossRef]
- Wei, T.-T.; Chandy, M.; Nishiga, M.; Zhang, A.; Kumar, K.K.; Thomas, D.; Manhas, A.; Rhee, S.; Justesen, J.M.; Chen, I.Y.; et al. Cannabinoid receptor 1 antagonist genistein attenuates marijuana-induced vascular inflammation. Cell 2022, 185, 1676–1693. [Google Scholar] [CrossRef]
- Guang, C.; Chen, J.; Sang, S.; Cheng, S. Biological functionality of soyasaponins and soyasapogenols. J. Agric. Food Chem. 2014, 62, 8247–8255. [Google Scholar] [CrossRef]
- Alam, M.A.; Islam, P.; Subhan, N.; Rahman, M.M.; Khan, F.; Burrows, G.E.; Nahar, L.; Sarker, S.D. Potential health benefits of anthocyanins in oxidative stress related disorders. Phytochem. Rev. 2021, 20, 705–749. [Google Scholar] [CrossRef]
- Zhang, R.F.; Zhang, F.X.; Zhang, M.W.; Wei, Z.C.; Yang, C.Y.; Zhang, Y.; Tang, X.J.; Deng, Y.Y.; Chi, J.W. Phenolic Composition and Antioxidant Activity in Seed Coats of 60 Chinese Black Soybean (Glycine max L. Merr.) Varieties. J. Agric. Food Chem. 2011, 59, 5935–5944. [Google Scholar] [CrossRef]
- Gao, R.; Han, T.; Xun, H.; Zeng, X.; Li, P.; Li, Y.; Wang, Y.; Shao, Y.; Cheng, X.; Feng, X.; et al. MYB transcription factors GmMYBA2 and GmMYBR function in a feedback loop to control pigmentation of seed coat in soybean. J. Exp. Bot. 2021, 72, 4401–4418. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Du, H.; Li, P.; Shen, Y.; Peng, H.; Liu, S.; Zhou, G.-A.; Zhang, H.; Liu, Z.; Shi, M.; et al. Pan-Genome of Wild and Cultivated Soybeans. Cell 2020, 182, 162–176.e113. [Google Scholar] [CrossRef] [PubMed]
- Fang, J. Bioavailability of anthocyanins. Drug Metab. Rev. 2014, 46, 508–520. [Google Scholar] [CrossRef] [PubMed]
- Shen, N.; Wang, T.; Gan, Q.; Liu, S.; Wang, L.; Jin, B. Plant flavonoids: Classification, distribution, biosynthesis, and antioxidant activity. Food Chem. 2022, 383, 132531. [Google Scholar] [CrossRef]
- Grzesik, M.; Naparlo, K.; Bartosz, G.; Sadowska-Bartosz, I. Antioxidant properties of catechins: Comparison with other antioxidants. Food Chem. 2018, 241, 480–492. [Google Scholar] [CrossRef]
- Mendes, A.P.; Borges, R.S.; Neto, A.M.; de Macedo, L.G.; da Silva, A.B. The basic antioxidant structure for flavonoid derivatives. J. Mol. Model 2012, 18, 4073–4080. [Google Scholar] [CrossRef]
- Surai, P.F. Silymarin as a Natural Antioxidant: An Overview of the Current Evidence and Perspectives. Antioxidants 2015, 4, 204–247. [Google Scholar] [CrossRef]
- Liang, Y.; Chen, G.; Zhang, F.; Yang, X.; Chen, Y.; Duan, Y.; Yu, M.; Zhang, S.; Han, J. Procyanidin B2 Reduces Vascular Calcification through Inactivation of ERK1/2-RUNX2 Pathway. Antioxidants 2021, 10, 916. [Google Scholar] [CrossRef]
- Jokioja, J.; Yang, B.; Linderborg, K.M. Acylated anthocyanins: A review on their bioavailability and effects on postprandial carbohydrate metabolism and inflammation. Compr. Rev. Food Sci. Food Saf. 2021, 20, 5570–5615. [Google Scholar] [CrossRef]
- Raab, T.; Barron, D.; Vera, F.A.; Crespy, V.; Oliveira, M.; Williamson, G. Catechin glucosides: Occurrence, synthesis, and stability. J. Agric. Food Chem. 2010, 58, 2138–2149. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Zhou, H.; Xiao, N.; Wu, X.; Shan, Y.; Chen, L.; Wang, C.; Wang, Z.; Huang, J.; Li, A.; et al. Expanding the Coverage of Metabolic Landscape in Cultivated Rice with Integrated Computational Approaches. Genom. Proteom. Bioinform. 2022, 20, 702–714. [Google Scholar] [CrossRef]
- Wang, F.; Liigand, J.; Tian, S.; Arndt, D.; Greiner, R.; Wishart, D.S. CFM-ID 4.0: More Accurate ESI-MS/MS Spectral Prediction and Compound Identification. Anal. Chem. 2021, 93, 11692–11700. [Google Scholar] [CrossRef]
- Pulido, R.; Bravo, L.; Saura-Calixto, F. Antioxidant activity of dietary polyphenols as determined by a modified ferric reducing/antioxidant power assay. J. Agric. Food Chem. 2000, 48, 3396–3402. [Google Scholar] [CrossRef] [PubMed]
- Miller, N.J.; Rice-Evans, C.A. Factors influencing the antioxidant activity determined by the ABTS.+ radical cation assay. Free. Radic. Res. 1997, 26, 195–199. [Google Scholar] [CrossRef] [PubMed]
- Li, H.-J.; Deinzer, M.L. Tandem Mass Spectrometry for Sequencing Proanthocyanidins. Anal. Chem. 2007, 79, 1739–1748. [Google Scholar] [CrossRef]
- Gosch, C.; Halbwirth, H.; Stich, K. Phloridzin: Biosynthesis, distribution and physiological relevance in plants. Phytochemistry 2010, 71, 838–843. [Google Scholar] [CrossRef] [PubMed]
- Kausar, H.; Ambrin, G.; Okla, M.K.; Soufan, W.; Al-Ghamdi, A.A.; Ahmad, A. Metabolic Flux Analysis of Catechin Biosynthesis Pathways Using Nanosensor. Antioxidants 2020, 9, 288. [Google Scholar] [CrossRef]
- Chang, W.T.; Huang, W.C.; Liou, C.J. Evaluation of the anti-inflammatory effects of phloretin and phlorizin in lipopolysaccharide-stimulated mouse macrophages. Food Chem. 2012, 134, 972–979. [Google Scholar] [CrossRef]
- Topal, F.; Nar, M.; Gocer, H.; Kalin, P.; Kocyigit, U.M.; Gulcin, I.; Alwasel, S.H. Antioxidant activity of taxifolin: An activity-structure relationship. J. Enzyme. Inhib. Med. Chem. 2016, 31, 674–683. [Google Scholar] [CrossRef]
- Sunil, C.; Xu, B. An insight into the health-promoting effects of taxifolin (dihydroquercetin). Phytochemistry 2019, 166, 112066. [Google Scholar] [CrossRef] [PubMed]
- Orzelska-Gorka, J.; Szewczyk, K.; Gawronska-Grzywacz, M.; Kedzierska, E.; Glowacka, E.; Herbet, M.; Dudka, J.; Biala, G. Monoaminergic system is implicated in the antidepressant-like effect of hyperoside and protocatechuic acid isolated from Impatiens glandulifera Royle in mice. Neurochem. Int. 2019, 128, 206–214. [Google Scholar] [CrossRef] [PubMed]
- Huang, J.; Zhou, L.; Chen, J.; Chen, T.; Lei, B.; Zheng, N.; Wan, X.; Xu, J.; Wang, T. Hyperoside Attenuate Inflammation in HT22 Cells via Upregulating SIRT1 to Activities Wnt/β-Catenin and Sonic Hedgehog Pathways. Neural. Plast. 2021, 2021, 8706400. [Google Scholar] [CrossRef] [PubMed]
- Xing, H.; Fu, R.; Cheng, C.; Cai, Y.; Wang, X.; Deng, D.; Gong, X.; Chen, J. Hyperoside Protected Against Oxidative Stress-Induced Liver Injury via the PHLPP2-AKT-GSK-3beta Signaling Pathway In Vivo and In Vitro. Front. Pharmacol. 2020, 11, 1065. [Google Scholar] [CrossRef] [PubMed]
- Tan, J.; Li, Y.; Hou, D.X.; Wu, S. The Effects and Mechanisms of Cyanidin-3-Glucoside and Its Phenolic Metabolites in Maintaining Intestinal Integrity. Antioxidants 2019, 8, 479. [Google Scholar] [CrossRef]
- Bernatoniene, J.; Kopustinskiene, D.M. The Role of Catechins in Cellular Responses to Oxidative Stress. Molecules 2018, 23, 965. [Google Scholar] [CrossRef] [PubMed]
- Hori, K.; Wada, M.; Yahara, S.; Watanabe, T.; Devkota, H.P. Antioxidant phenolic compounds from the rhizomes of Astilbe rivularis. Nat. Prod. Res. 2018, 32, 453–456. [Google Scholar] [CrossRef]
- Wong, K.C.; Cao, S.; Dong, X.; Law, M.C.; Chan, T.H.; Wong, M.S. (−)-Epiafzelechin Protects against Ovariectomy-induced Bone Loss in Adult Mice and Modulate Osteoblastic and Osteoclastic Functions In Vitro. Nutrients 2017, 9, 530. [Google Scholar] [CrossRef]
- Min, K.R.; Hwang, B.Y.; Lim, H.S.; Kang, B.S.; Oh, G.J.; Lee, J.; Kang, S.H.; Lee, K.S.; Ro, J.S.; Kim, Y. (−)-Epiafzelechin: Cyclooxygenase-1 inhibitor and anti-inflammatory agent from aerial parts of Celastrus orbiculatus. Planta. Medica 1999, 65, 460–462. [Google Scholar] [CrossRef]
- Xu, Q.; Fu, Q.; Li, Z.; Liu, H.; Wang, Y.; Lin, X.; He, R.; Zhang, X.; Ju, Z.; Campisi, J.; et al. The flavonoid procyanidin C1 has senotherapeutic activity and increases lifespan in mice. Nat. Metab. 2021, 3, 1706–1726. [Google Scholar] [CrossRef]
- Chen, L.; Teng, H.; Xie, Z.; Cao, H.; Cheang, W.S.; Skalicka-Woniak, K.; Georgiev, M.I.; Xiao, J. Modifications of dietary flavonoids towards improved bioactivity: An update on structure-activity relationship. Crit. Rev. Food Sci. Nutr. 2018, 58, 513–527. [Google Scholar] [CrossRef]
- Ahmad, M.Z.; Zhang, Y.; Zeng, X.; Li, P.; Wang, X.; Benedito, V.A.; Zhao, J.; Gifford, M. Isoflavone malonyl-CoA acyltransferase GmMaT2 is involved in nodulation of soybean by modifying synthesis and secretion of isoflavones. J. Exp. Bot. 2021, 72, 1349–1369. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, M.Z.; Li, P.; Wang, J.; Rehman, N.U.; Zhao, J. Isoflavone Malonyltransferases GmIMaT1 and GmIMaT3 Differently Modify Isoflavone Glucosides in Soybean (Glycine max) under Various Stresses. Front. Plant Sci. 2017, 8, 735. [Google Scholar] [CrossRef] [PubMed]
- Fang, S.; Qu, Q.; Zheng, Y.; Zhong, H.; Shan, C.; Wang, F.; Li, C.; Peng, G. Structural characterization and identification of flavonoid aglycones in three Glycyrrhiza species by liquid chromatography with photodiode array detection and quadrupole time-of-flight mass spectrometry. J. Sep. Sci. 2016, 39, 2068–2078. [Google Scholar] [CrossRef]
- Iijima, Y.; Suda, K.; Suzuki, T.; Aoki, K.; Shibata, D. Metabolite Profiling of Chalcones and Flavanones in Tomato Fruit. J. Jpn. Soc. Hortic. Sci. 2008, 77, 94–102. [Google Scholar] [CrossRef]
- Pellati, F.; Orlandini, G.; Pinetti, D.; Benvenuti, S. HPLC-DAD and HPLC-ESI-MS/MS methods for metabolite profiling of propolis extracts. J. Pharm. Biomed. Anal. 2011, 55, 934–948. [Google Scholar] [CrossRef]
- Zheng, G.D.; Zhou, P.; Yang, H.; Li, Y.S.; Li, P.; Liu, E.H. Rapid resolution liquid chromatography-electrospray ionisation tandem mass spectrometry method for identification of chemical constituents in Citri Reticulatae Pericarpium. Food Chem. 2013, 136, 604–611. [Google Scholar] [CrossRef]
- Abad-Garcia, B.; Garmon-Lobato, S.; Berrueta, L.A.; Gallo, B.; Vicente, F. A fragmentation study of dihydroquercetin using triple quadrupole mass spectrometry and its application for identification of dihydroflavonols in Citrus juices. Rapid Commun. Mass Spectrom. 2009, 23, 2785–2792. [Google Scholar] [CrossRef]
- Ma, C.; Lv, H.; Zhang, X.; Chen, Z.; Shi, J.; Lu, M.; Lin, Z. Identification of regioisomers of methylated kaempferol and quercetin by ultra high performance liquid chromatography quadrupole time-of-flight (UHPLC-QTOF) tandem mass spectrometry combined with diagnostic fragmentation pattern analysis. Anal. Chim. Acta 2013, 795, 15–24. [Google Scholar] [CrossRef]
- Barnes, J.S.; Schug, K.A. Structural characterization of cyanidin-3,5-diglucoside and pelargonidin-3,5-diglucoside anthocyanins: Multi-dimensional fragmentation pathways using high performance liquid chromatography-electrospray ionization-ion trap-time of flight mass spectrometry. Int. J. Mass Spectrom. 2011, 308, 71–80. [Google Scholar] [CrossRef]
- Nakata, R.; Yoshinaga, N.; Teraishi, M.; Okumoto, Y.; Huffaker, A.; Schmelz, E.A.; Mori, N. A fragmentation study of isoflavones by IT-TOF-MS using biosynthesized isotopes. Biosci. Biotechnol. Biochem. 2018, 82, 1309–1315. [Google Scholar] [CrossRef]
- Zhang, B.; Hettiarachchy, N.; Chen, P.; Horax, R.; Cornelious, B.; Zhu, D. Influence of the application of three different elicitors on soybean plants on the concentrations of several isoflavones in soybean seeds. J. Agric. Food Chem. 2006, 54, 5548–5554. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Z.; Jiang, Y.; Wang, Z.; Gou, Z.; Lyu, J.; Li, W.; Yu, Y.; Shu, L.; Zhao, Y.; Ma, Y.; et al. Resequencing 302 wild and cultivated accessions identifies genes related to domestication and improvement in soybean. Nat. Biotechnol. 2015, 33, 408–414. [Google Scholar] [CrossRef]
- Lu, S.; Dong, L.; Fang, C.; Liu, S.; Kong, L.; Cheng, Q.; Chen, L.; Su, T.; Nan, H.; Zhang, D.; et al. Stepwise selection on homeologous PRR genes controlling flowering and maturity during soybean domestication. Nat. Genet. 2020, 52, 428–436. [Google Scholar] [CrossRef]
- Goettel, W.; Zhang, H.; Li, Y.; Qiao, Z.; Jiang, H.; Hou, D.; Song, Q.; Pantalone, V.R.; Song, B.H.; Yu, D.; et al. POWR1 is a domestication gene pleiotropically regulating seed quality and yield in soybean. Nat. Commun. 2022, 13, 3051. [Google Scholar] [CrossRef]
- Singh, B.; Singh, J.P.; Kaur, A.; Singh, N. Phenolic composition and antioxidant potential of grain legume seeds: A review. Food Res. Int. 2017, 101, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Chen, P.X.; Tang, Y.; Marcone, M.F.; Pauls, P.K.; Zhang, B.; Liu, R.; Tsao, R. Characterization of free, conjugated and bound phenolics and lipophilic antioxidants in regular- and non-darkening cranberry beans (Phaseolus vulgaris L.). Food Chem. 2015, 185, 298–308. [Google Scholar] [CrossRef]
- Aguilera, Y.; Duenas, M.; Estrella, I.; Hernandez, T.; Benitez, V.; Esteban, R.M.; Martin-Cabrejas, M.A. Evaluation of phenolic profile and antioxidant properties of Pardina lentil as affected by industrial dehydration. J. Agric. Food Chem. 2010, 58, 10101–10108. [Google Scholar] [CrossRef] [PubMed]
- Zhou, S.; Sekizaki, H.; Yang, Z.; Sawa, S.; Pan, J. Phenolics in the seed coat of wild soybean (Glycine soja) and their significance for seed hardness and seed germination. J. Agric. Food Chem. 2010, 58, 10972–10978. [Google Scholar] [CrossRef] [PubMed]
- Yun, D.Y.; Kang, Y.G.; Kim, M.; Kim, D.; Kim, E.H.; Hong, Y.S. Metabotyping of different soybean genotypes and distinct metabolism in their seeds and leaves. Food Chem. 2020, 330, 127198. [Google Scholar] [CrossRef] [PubMed]
- Hyeon, H.; Xu, J.L.; Kim, J.K.; Choi, Y. Comparative metabolic profiling of cultivated and wild black soybeans reveals distinct metabolic alterations associated with their domestication. Food Res. Int. 2020, 134, 109290. [Google Scholar] [CrossRef]
- Yun, D.Y.; Kang, Y.G.; Yun, B.; Kim, E.H.; Kim, M.; Park, J.S.; Lee, J.H.; Hong, Y.S. Distinctive Metabolism of Flavonoid between Cultivated and Semiwild Soybean Unveiled through Metabolomics Approach. J. Agric. Food Chem. 2016, 64, 5773–5783. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Alseekh, S.; Fernie, A.R.; Luo, J. The Structure and Function of Major Plant Metabolite Modifications. Mol. Plant 2019, 12, 899–919. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name | Structure | Classification | RT (min) | Exact Mass | Characteristic Fragment Ions | Reference |
|---|---|---|---|---|---|---|
| cyanidin 3-O-glucoside |  | anthocyanins | 5.25 | 449.1084 | [M–Glc]+: 287.0556, 0,2A+: 149.0239, 0,2B+: 137.0239, 0,3A+: 121.029. | Chemical standard |
| (+)-catechin |  | flavanols | 5.51 | 290.079 | [M+H]+: 291.0869, 1,4B+: 165.0552, 1,4B+–H2O: 147.0446, 1,3A+: 139.0395, 1,2B+: 123.0446. | Chemical standard |
| catechin-glucoside |  | flavanols | 5.69 | 452.1319 | [M+H–Glc]+: 291.0869, 1,4B+: 165.0552, 1,2A+–H2O: 151.0395, 1,4B+–H2O: 147.0446, 1,3A+: 139.0395, 1,2B+: 123.0446. | Fragmentation pattern in [27]. |
| procyanidin B2 |  | proanthocyanidins | 6.03 | 578.1424 | [M+H]+: 579.1503, 427.1029, 409.0923, 301.0712, 291.0869, 289.0712, 287.0556, 165.0552, 151.0395, 147.0446, 139.0395, 127.0395, 123.0446. | Chemical standard |
| (−)-epicatechin |  | flavanols | 6.32 | 290.079 | [M+H]+: 291.0869, 1,4B+: 165.0552, 1,4B+–H2O: 147.0446, 1,3A+: 139.0395, 1,2B+: 123.0446. | Chemical standard |
| procyanidin C1 |  | proanthocyanidins | 6.55 | 866.2058 | [M+H]+: 867.2136, 589.1346, 579.1503, 451.1029, 427.1029, 409.0923, 301.0712, 291.0869, 289.0712, 287.0556, 165.0552, 151.0395, 147.0446, 139.0395, 127.0395, 123.0446. | Chemical standard |
| afzelechin-catechin |  | proanthocyanidins | 6.61 | 562.1475 | [M+H]+: 563.1553, 411.108, 291.0869, 287.0556, 285.0763, 273.0763, 165.0552, 151.0395, 147.0446, 139.0395, 123.0446, 107.0497. | Fragmentation pattern in [27]. |
| (−)-epiafzelechin |  | flavanols | 7.00 | 274.0841 | [M+H]+: 275.0919, 1,4B+: 149.0603, 1,3A+: 139.0395, 1,2B+: 107.0497. | Chemical standard |
| quercetin 3-O-galactoside |  | flavonols | 7.39 | 464.0955 | [M+H–Gal]+: 303.0505, [M+H–Gal–H2O]+: 285.0399, 0,2A+: 165.0188, 1,3A+: 153.0188, 0,2B+: 137.0239. | Chemical standard |
| taxifolin |  | dihydroflavonols | 7.74 | 304.0583 | [M–H]−: 303.0505, [M–H–H2O]−: 285.0399, 1,4B−: 177.0188, 1,3A−: 151.0031, 1,4A−: 125.0239. | Chemical standard |
| phlorizin |  | dihydrochalcones | 8.55 | 436.1369 | [M–H–Glc]−: 273.0763, [M–H–Glc–C7H6O]−: 167.0344, [M–H–Glc–C7H6O–C2H2O]−: 125.0239, [M–H–Glc–C7H6O–CO2]−: 123.0446. | Chemical standard |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, X.; Li, S.; Wang, J.; Chen, G.; Tao, X.; Xu, S. Metabolomic Analysis Reveals Domestication-Driven Reshaping of Polyphenolic Antioxidants in Soybean Seeds. Antioxidants 2023, 12, 912. https://doi.org/10.3390/antiox12040912
Li X, Li S, Wang J, Chen G, Tao X, Xu S. Metabolomic Analysis Reveals Domestication-Driven Reshaping of Polyphenolic Antioxidants in Soybean Seeds. Antioxidants. 2023; 12(4):912. https://doi.org/10.3390/antiox12040912
Chicago/Turabian StyleLi, Xuetong, Sujuan Li, Jian Wang, Guang Chen, Xiaoyuan Tao, and Shengchun Xu. 2023. "Metabolomic Analysis Reveals Domestication-Driven Reshaping of Polyphenolic Antioxidants in Soybean Seeds" Antioxidants 12, no. 4: 912. https://doi.org/10.3390/antiox12040912
APA StyleLi, X., Li, S., Wang, J., Chen, G., Tao, X., & Xu, S. (2023). Metabolomic Analysis Reveals Domestication-Driven Reshaping of Polyphenolic Antioxidants in Soybean Seeds. Antioxidants, 12(4), 912. https://doi.org/10.3390/antiox12040912