
Impact of Lactic Acid Bacteria Fermentation on Phenolic Compounds and Antioxidant Activity of Avocado Leaf Extracts

 ,
,  ,
,  , ,
, ,  and
and 
Abstract
1. Introduction
2. Materials and Methods
2.1. Chemicals and Samples
2.2. Lactic Acid Bacteria Strains and Culture Media
2.3. Preparation of the Inocula
2.4. Fermentation of Avocado Leaves
2.5. Polar Compound Extraction
2.6. Determination of Folin–Ciocalteu Reacting Substances
2.7. Determination of Antioxidant Activity: DPPH and FRAP Assays
2.8. Determination of Polar Compounds by HPLC-ESI-TOF-MS
2.9. Data Processing
3. Results and Discussion
3.1. Screening of Lactic Acid Bacteria in Fermentation of Avocado Leaves
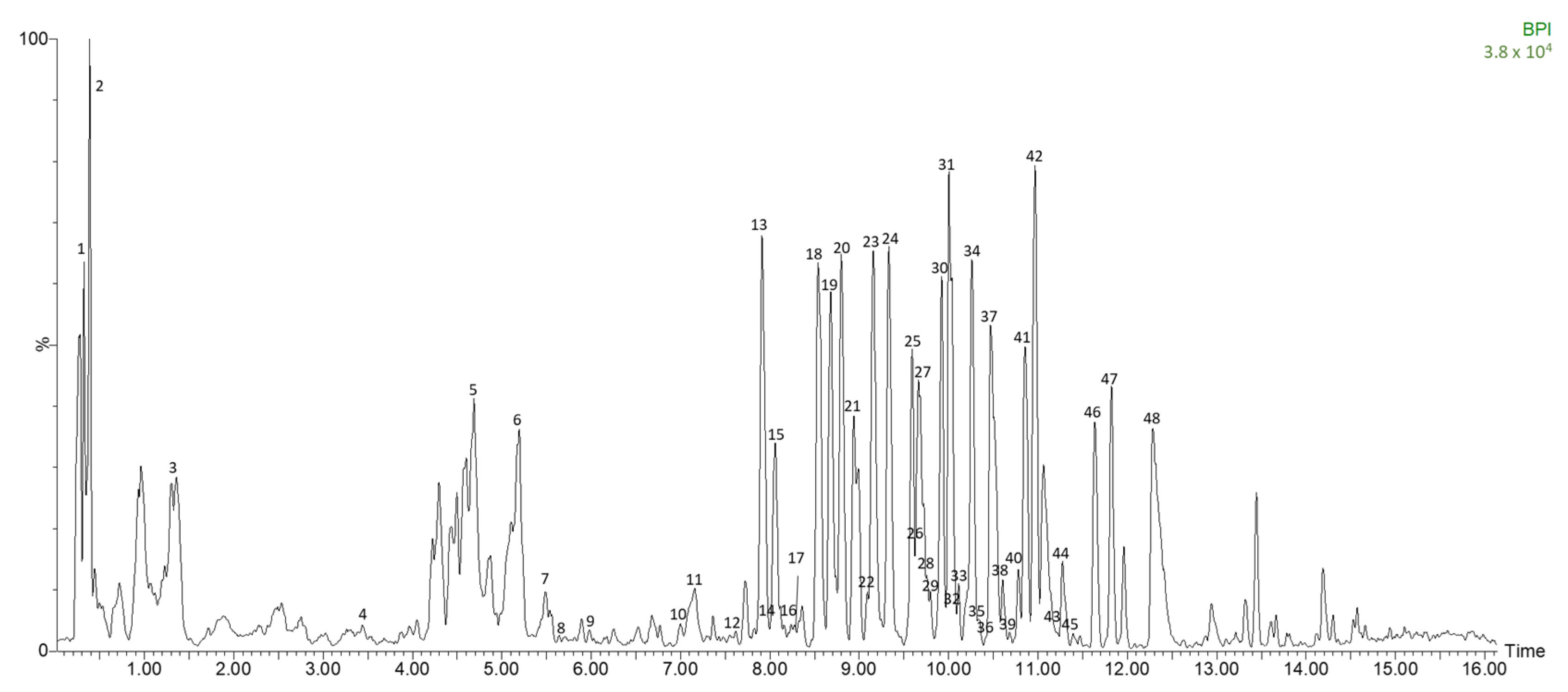
3.2. Identification of Polar Compounds in Fermented Avocado Leaves by HPLC-ESI-TOF-MS
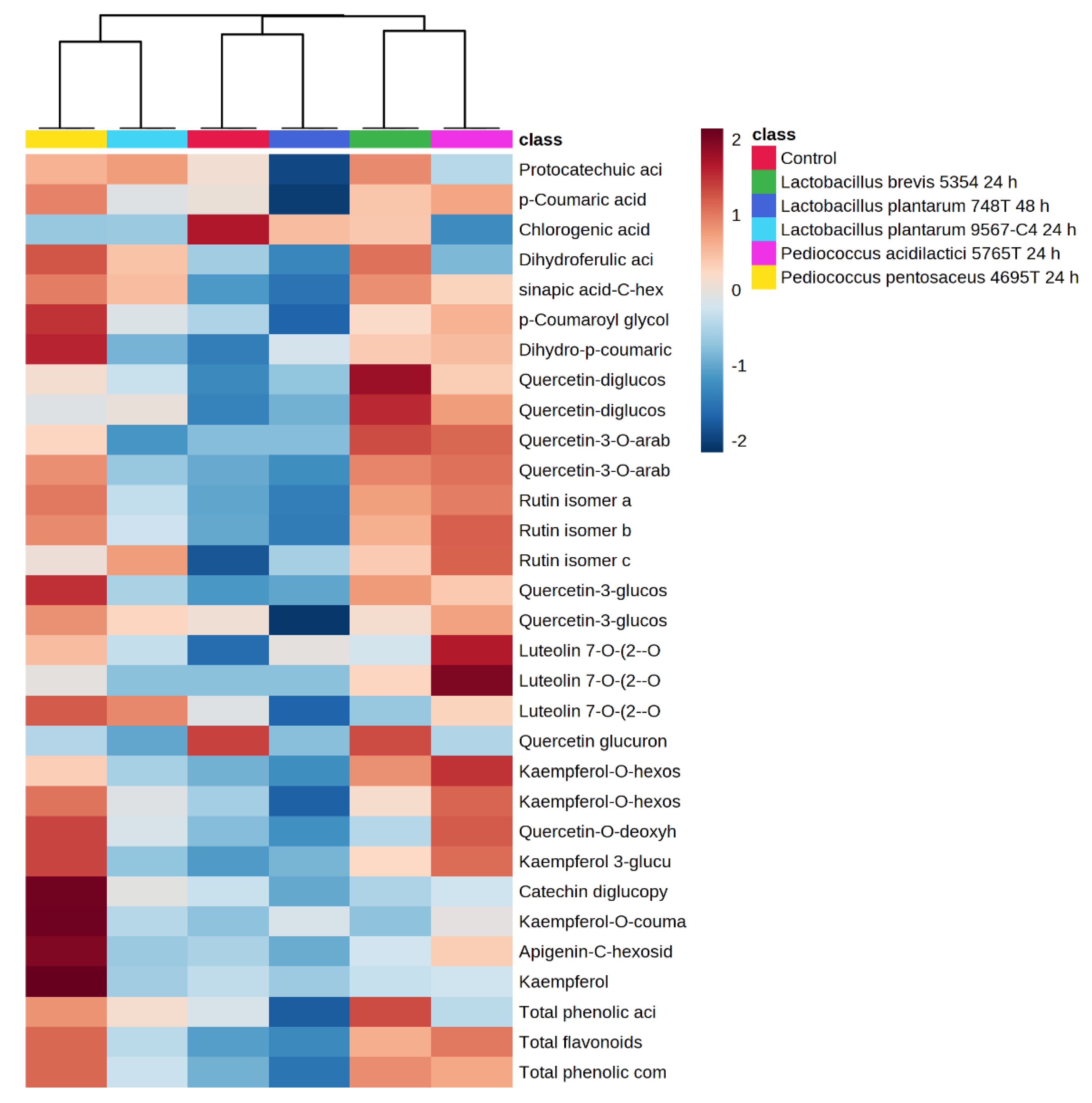
3.3. Quantification of Phenolic Compounds by HPLC-ESI-TOF-MS and Its Biotransformations during Fermentation in Avocado Leaves
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Mora-Sand, A.; Ram, A.; Castillo-Henr, L.; Lopretti-Correa, M.; Vega-Baudrit, J.R. Persea americana Agro-Industrial Waste Biorefinery for Sustainable High-Value-Added Products. Polymers 2021, 13, 1727. [Google Scholar] [CrossRef] [PubMed]
- Sommaruga, R.; Eldridge, H.M. Avocado Production: Water Footprint and Socio-economic Implications. EuroChoices 2021, 20, 48–53. [Google Scholar] [CrossRef]
- Ramírez-Mejía, D.; Levers, C.; Mas, J.F. Spatial patterns and determinants of avocado frontier dynamics in Mexico. Reg. Environ. Chang. 2022, 22, 28. [Google Scholar] [CrossRef] [PubMed]
- Capanoglu, E.; Nemli, E.; Tomas-Barberan, F. Novel Approaches in the Valorization of Agricultural Wastes and Their Applications. J. Agric. Food Chem. 2022, 70, 6787–6804. [Google Scholar] [CrossRef] [PubMed]
- Solís-Salas, L.M.; Sierra-Rivera, C.A.; Cobos-Puc, L.E.; Ascacio-Valdés, J.A.; Silva-Belmares, S.Y. Antibacterial potential by rupture membrane and antioxidant capacity of purified phenolic fractions of Persea americana leaf extract. Antibiotics 2021, 10, 508. [Google Scholar] [CrossRef]
- Nathaniel, O.B.; Selina, A.S.; John, K.M.; Mercy, B.; Sylvester, A.A.; Michael, B.M. Phytoconstituents, antimicrobial and antioxidant properties of the leaves of Persea americana Mill cultivated in Ghana. J. Med. Plants Res. 2015, 9, 933–939. [Google Scholar] [CrossRef]
- Park, S.J.; Nam, Y.H.; Rodriguez, I.; Park, J.H.; Kwak, H.J.; Oh, Y.; Oh, M.; Park, M.S.; Lee, K.W.; Lee, J.S.; et al. Chemical constituents of leaves of Persea americana (avocado) and their protective effects against neomycin-induced hair cell damage. Rev. Bras. Farmacogn. 2019, 29, 739–743. [Google Scholar] [CrossRef]
- Yamassaki, F.T.; Campestrini, L.H.; Zawadzki-Baggio, S.F.; Maurer, J.B.B. Chemical characterization and complement modulating activities of an arabinogalactan-protein-rich fraction from an aqueous extract of avocado leaves. Int. J. Biol. Macromol. 2018, 120, 513–521. [Google Scholar] [CrossRef]
- Rincón-Hernández, C.A.; De La Pérez, J.L.S.; Espinosa-García, F.J. Caracterización química foliar de los árboles de aguacate criollo (Persea americana var. drymifolia) en los bancos de germoplasma de Michoacán, México. Rev. Mex. Biodivers. 2011, 82, 395–412. [Google Scholar] [CrossRef]
- Jimenez, P.; Garcia, P.; Quitral, V.; Vasquez, K.; Parra-Ruiz, C.; Reyes-Farias, M.; Garcia-Diaz, D.F.; Robert, P.; Encina, C.; Soto-Covasich, J. Pulp, Leaf, Peel and Seed of Avocado Fruit: A Review of Bioactive Compounds and Healthy Benefits. Food Rev. Int. 2021, 37, 619–655. [Google Scholar] [CrossRef]
- Martins, S.; Mussatto, S.I.; Martínez-Avila, G.; Montañez-Saenz, J.; Aguilar, C.N.; Teixeira, J.A. Bioactive phenolic compounds: Production and extraction by solid-state fermentation. A review. Biotechnol. Adv. 2011, 29, 365–373. [Google Scholar] [CrossRef] [PubMed]
- Rojas-García, A.; Fuentes, E.; Cádiz-Gurrea, M.d.l.L.; Rodriguez, L.; Villegas-Aguilar, M.D.C.; Palomo, I.; Arráez-Román, D.; Segura-Carretero, A. Biological Evaluation of Avocado Residues as a Potential Source of Bioactive Compounds. Antioxidants 2022, 11, 1049. [Google Scholar] [CrossRef] [PubMed]
- Castro-López, C.; Bautista-Hernández, I.; González-Hernández, M.D.; Martínez-Ávila, G.C.G.; Rojas, R.; Gutiérrez-Díez, A.; Medina-Herrera, N.; Aguirre-Arzola, V.E. Polyphenolic Profile and Antioxidant Activity of Leaf Purified Hydroalcoholic Extracts from Seven Mexican Persea americana Cultivars. Molecules 2019, 24, 173. [Google Scholar] [CrossRef]
- Muñoz, R.; de las Rivas, B.; López de Felipe, F.; Reverón, I.; Santamaría, L.; Esteban-Torres, M.; Curiel, J.A.; Rodríguez, H.; Landete, J.M. Biotransformation of Phenolics by Lactobacillus plantarum in Fermented Foods; Elsevier Inc.: Amsterdam, The Netherlands, 2017; ISBN 9780128023099. [Google Scholar]
- Selma, M.V.; Espín, J.C.; Tomás-Barberán, F.A. Interaction between phenolics and gut microbiota: Role in human health. J. Agric. Food Chem. 2009, 57, 6485–6501. [Google Scholar] [CrossRef] [PubMed]
- Verni, M.; De Mastro, G.; De Cillis, F.; Gobbetti, M.; Rizzello, C.G. Lactic acid bacteria fermentation to exploit the nutritional potential of Mediterranean faba bean local biotypes. Food Res. Int. 2019, 125, 108571. [Google Scholar] [CrossRef] [PubMed]
- Bergillos-Meca, T.; Cabrera-Vique, C.; Artacho, R.; Moreno-Montoro, M.; Navarro-Alarcón, M.; Olalla, M.; Giménez, R.; Seiquer, I.; Ruiz-López, M.D. Does Lactobacillus plantarum or ultrafiltration process improve Ca, Mg, Zn and P bioavailability from fermented goats’ milk? Food Chem. 2015, 187, 314–321. [Google Scholar] [CrossRef] [PubMed]
- Curiel, J.A.; Pinto, D.; Marzani, B.; Filannino, P.; Farris, G.A.; Gobbetti, M.; Rizzello, C.G. Lactic acid fermentation as a tool to enhance the antioxidant properties of Myrtus communis berries. Microb. Cell Fact. 2015, 14, 67. [Google Scholar] [CrossRef]
- Pontonio, E.; Montemurro, M.; Pinto, D.; Marzani, B.; Trani, A.; Ferrara, G.; Mazzeo, A.; Gobbetti, M.; Rizzello, C.G. Lactic acid fermentation of pomegranate juice as a tool to improve antioxidant activity. Front. Microbiol. 2019, 10, 1550. [Google Scholar] [CrossRef]
- Rodríguez, H.; Curiel, J.A.; Landete, J.M.; de las Rivas, B.; de Felipe, F.L.; Gómez-Cordovés, C.; Mancheño, J.M.; Muñoz, R. Food phenolics and lactic acid bacteria. Int. J. Food Microbiol. 2009, 132, 79–90. [Google Scholar] [CrossRef]
- Hatti-Kaul, R.; Chen, L.; Dishisha, T.; El Enshasy, H. Lactic acid bacteria: From starter cultures to producers of chemicals. FEMS Microbiol. Lett. 2018, 365, fny213. [Google Scholar] [CrossRef]
- Wang, Y.; Wu, J.; Lv, M.; Shao, Z.; Hungwe, M.; Wang, J.; Bai, X.; Xie, J.; Wang, Y.; Geng, W. Metabolism Characteristics of Lactic Acid Bacteria and the Expanding Applications in Food Industry. Front. Bioeng. Biotechnol. 2021, 9, 612285. [Google Scholar] [CrossRef] [PubMed]
- Filannino, P.; Di Cagno, R.; Gobbetti, M. Metabolic and functional paths of lactic acid bacteria in plant foods: Get out of the labyrinth. Curr. Opin. Biotechnol. 2018, 49, 64–72. [Google Scholar] [CrossRef] [PubMed]
- Filannino, P.; Bai, Y.; Di Cagno, R.; Gobbetti, M.; Gänzle, M.G. Metabolism of phenolic compounds by Lactobacillus spp. during fermentation of cherry juice and broccoli puree. Food Microbiol. 2015, 46, 272–279. [Google Scholar] [CrossRef] [PubMed]
- Filannino, P.; Gobbetti, M.; De Angelis, M.; Di Cagno, R. Hydroxycinnamic acids used as external acceptors of electrons: An energetic advantage for strictly heterofermentative lactic acid bacteria. Appl. Environ. Microbiol. 2014, 80, 7574–7582. [Google Scholar] [CrossRef]
- Sánchez-Maldonado, A.F.; Schieber, A.; Gänzle, M.G. Structure-function relationships of the antibacterial activity of phenolic acids and their metabolism by lactic acid bacteria. J. Appl. Microbiol. 2011, 111, 1176–1184. [Google Scholar] [CrossRef]
- Reverón, I.; de las Rivas, B.; Muñoz, R.; López de Felipe, F. Genome-wide transcriptomic responses of a human isolate of Lactobacillus plantarum exposed to p-coumaric acid stress. Mol. Nutr. Food Res. 2012, 56, 1848–1859. [Google Scholar] [CrossRef]
- Lamuela-Raventós, R.M. Folin–Ciocalteu method for the measurement of total phenolic content and antioxidant capacity. In Measurement of Antioxidant Activity & Capacity; John Wiley & Sons, Ltd.: Hoboken, NJ, USA, 2017; pp. 107–115. ISBN 9781119135388. [Google Scholar]
- Razola-Díaz, M.d.C.; Guerra-Hernández, E.J.; Rodríguez-Pérez, C.; Gómez-Caravaca, A.M.; García-Villanova, B.; Verardo, V. Optimization of Ultrasound-Assisted Extraction via Sonotrode of Phenolic Compounds from Orange By-Products. Foods 2021, 10, 1120. [Google Scholar] [CrossRef]
- Razola-Díaz, M.d.C.; Gómez-Caravaca, A.M.; Guerra-Hernández, E.J.; Garcia-Villanova, B.; Verardo, V. New Advances in the Phenolic Composition of Tiger Nut (Cyperus esculentus L.) by-Products. Foods 2022, 11, 343. [Google Scholar] [CrossRef]
- Verni, M.; Pontonio, E.; Krona, A.; Jacob, S.; Pinto, D.; Rinaldi, F.; Verardo, V.; Díaz-de-Cerio, E.; Coda, R.; Rizzello, C.G. Bioprocessing of Brewers’ Spent Grain Enhances Its Antioxidant Activity: Characterization of Phenolic Compounds and Bioactive Peptides. Front. Microbiol. 2020, 11, 1831. [Google Scholar] [CrossRef]
- Leonard, W.; Zhang, P.; Ying, D.; Adhikari, B.; Fang, Z. Fermentation transforms the phenolic profiles and bioactivities of plant-based foods. Biotechnol. Adv. 2021, 49, 107763. [Google Scholar] [CrossRef]
- García-Rodríguez, Y.M.; Torres-Gurrola, G.; Meléndez-González, C.; Espinosa-García, F.J. Phenotypic Variations in the Foliar Chemical Profile of Persea americana Mill. cv. Hass. Chem. Biodivers. 2016, 13, 1767–1775. [Google Scholar] [CrossRef] [PubMed]
- Hervert-Hernández, D.; Goñi, I. Dietary polyphenols and human gut microbiota: A review. Food Rev. Int. 2011, 27, 154–169. [Google Scholar] [CrossRef]
- Ghabbour, N.; Lamzira, Z.; Thonart, P.; Cidalia, P.; Markaoui, M.; Asehraou, A. Selection of oleuropein-degrading lactic acid bacteria strains isolated from fermenting moroccan green olives. Grasas Aceites 2011, 62, 84–89. [Google Scholar] [CrossRef]
- Yamassaki, F.T.; Campestrini, L.H.; Faria, S.; Bello, J.; Maurer, B. Avocado leaves: Influence of drying process, thermal incubation, and storage conditions on preservation of polyphenolic compounds and antioxidant activity. Int. J. Food Prop. 2017, 20, 2280–2293. [Google Scholar] [CrossRef]
- Filannino, P.; Tlais, A.Z.A.; Morozova, K.; Cavoski, I.; Scampicchio, M.; Gobbetti, M.; Di Cagno, R. Lactic acid fermentation enriches the profile of biogenic fatty acid derivatives of avocado fruit (Persea americana Mill.). Food Chem. 2020, 317, 126384. [Google Scholar] [CrossRef]
- López-Cobo, A.; Gómez-Caravaca, A.M.; Pasini, F.; Caboni, M.F.; Segura-Carretero, A.; Fernández-Gutiérrez, A. HPLC-DAD-ESI-QTOF-MS and HPLC-FLD-MS as valuable tools for the determination of phenolic and other polar compounds in the edible part and by-products of avocado. LWT—Food Sci. Technol. 2016, 73, 505–513. [Google Scholar] [CrossRef]
- Rothwell, J.A.; Perez-Jimenez, J.; Neveu, V.; Medina-Remón, A.; M’hiri, N.; García-Lobato, P.; Manach, C.; Knox, C.; Eisner, R.; Wishart, D.S.; et al. Phenol-Explorer 3.0: A major update of the Phenol-Explorer database to incorporate data on the effects of food processing on polyphenol content. Database 2013, 2013, bat070. [Google Scholar] [CrossRef]
- Dueñas, M.; Hernández, T.; Estrella, I. Changes in the content of bioactive polyphenolic compounds of lentils by the action of exogenous enzymes. Effect on their antioxidant activity. Food Chem. 2007, 101, 90–97. [Google Scholar] [CrossRef]
- Boskou, G.; Salta, F.N.; Chrysostomou, S.; Mylona, A.; Chiou, A.; Andrikopoulos, N.K. Antioxidant capacity and phenolic profile of table olives from the Greek market. Food Chem. 2006, 94, 558–564. [Google Scholar] [CrossRef]
- Hu, T.; Subbiah, V.; Wu, H.; Amrit, B.K.; Rauf, A.; Alhumaydhi, F.A.; Suleria, H.A.R. Determination and Characterization of Phenolic Compounds from Australia-Grown Sweet Cherries (Prunus avium L.) and Their Potential Antioxidant Properties. ACS Omega 2021, 6, 34687–34699. [Google Scholar] [CrossRef]
- Fan, S.; Qi, Y.; Shi, L.; Giovani, M.; Zaki, N.A.A.; Guo, S.; Suleria, H.A.R. Screening of Phenolic Compounds in Rejected Avocado and Determination of Their Antioxidant Potential. Processes 2022, 10, 1747. [Google Scholar] [CrossRef]
- Mi, J.C.; Howard, L.R.; Prior, R.L.; Clark, J.R. Flavonoid glycosides and antioxidant capacity of various blackberry, blueberry and red grape genotypes determined by high-performance liquid chromatography/mass spectrometry. J. Sci. Food Agric. 2004, 84, 1771–1782. [Google Scholar] [CrossRef]
- Wang, S.Y.; Zheng, W.; Galletta, G.J. Cultural system affects fruit quality and antioxidant capacity in strawberries. J. Agric. Food Chem. 2002, 50, 6534–6542. [Google Scholar] [CrossRef] [PubMed]
- DuPont, M.S.; Mondin, Z.; Williamson, G.; Price, K.R. Effect of variety, processing, and storage on the flavonoid glycoside content and composition of lettuce and endive. J. Agric. Food Chem. 2000, 48, 3957–3964. [Google Scholar] [CrossRef]
- Nakane, R.; Iwashina, T. Flavonol glycosides from the leaves of allium macrostemon. Nat. Prod. Commun. 2015, 10, 1381–1382. [Google Scholar] [CrossRef]
- Xiong, Y.; Zhang, P.; Warner, R.D.; Shen, S.; Johnson, S.; Fang, Z. HPLC-DAD-ESI-QTOF-MS/MS qualitative analysis data and HPLC-DAD quantification data of phenolic compounds of grains from five Australian sorghum genotypes. Data Brief 2020, 33, 106584. [Google Scholar] [CrossRef]
- Figueroa, J.G.; Borrás-Linares, I.; Del Pino-García, R.; Curiel, J.A.; Lozano-Sánchez, J.; Segura-Carretero, A. Functional ingredient from avocado peel: Microwave-assisted extraction, characterization and potential applications for the food industry. Food Chem. 2021, 352, 129300. [Google Scholar] [CrossRef]
- Filannino, P.; Cardinali, G.; Rizzello, C.G.; Buchin, S.; De Angelis, M.; Gobbetti, M.; Di Cagno, R. Metabolic responses of Lactobacillus plantarum strains during fermentation and storage of vegetable and fruit juices. Appl. Environ. Microbiol. 2014, 80, 2206–2215. [Google Scholar] [CrossRef]
- Dueñas, M.; Fernández, D.; Hernández, T.; Estrella, I.; Muñoz, R. Bioactive phenolic compounds of cowpeas (Vigna sinensis L). Modifications by fermentation with natural microflora and with Lactobacillus plantarum ATCC 14917. J. Sci. Food Agric. 2005, 85, 297–304. [Google Scholar] [CrossRef]
- Cavin, J.F.; Andioc, V.; Etiévant, P.X.; Divies, C. Ability of wine lactic acid bacteria to metabolize phenol carboxylic acids. Am. J. Enol. Vitic. 1993, 44, 76–80. [Google Scholar] [CrossRef]
- Landete, J.M.; Rodríguez, H.; Curiel, J.A.; De Las Rivas, B.; Mancheño, J.M.; Muñoz, R. Gene cloning, expression, and characterization of phenolic acid decarboxylase from Lactobacillus brevis RM84. J. Ind. Microbiol. Biotechnol. 2010, 37, 617–624. [Google Scholar] [CrossRef] [PubMed]
- Esteban-Torres, M.; Landete, J.M.; Reverón, I.; Santamaría, L.; de las Rivas, B.; Muñoz, R. A Lactobacillus plantarum esterase active on a broad range of phenolic esters. Appl. Environ. Microbiol. 2015, 81, 3235–3242. [Google Scholar] [CrossRef]
- Lin, S.; Zhu, Q.; Wen, L.; Yang, B.; Jiang, G.; Gao, H.; Chen, F.; Jiang, Y. Production of quercetin, kaempferol and their glycosidic derivatives from the aqueous-organic extracted residue of litchi pericarp with Aspergillus awamori. Food Chem. 2014, 145, 220–227. [Google Scholar] [CrossRef] [PubMed]
- Lee, N.K.; Jeong, J.H.; Oh, J.; Kim, Y.; Ha, Y.S.; Jeong, Y.S. Conversion of Flavonols Kaempferol and Quercetin in Mulberry (Morus alba L.) Leaf Using Plant-Fermenting Lactobacillus plantarum. J. Food Biochem. 2015, 39, 765–770. [Google Scholar] [CrossRef]
- Landete, J.M.; Hernández, T.; Robredo, S.; Dueñas, M.; De Las Rivas, B.; Estrella, I.; Muñoz, R. Effect of soaking and fermentation on content of phenolic compounds of soybean (Glycine max cv. Merit) and mung beans (Vigna radiata [L] Wilczek). Int. J. Food Sci. Nutr. 2015, 66, 203–209. [Google Scholar] [CrossRef] [PubMed]
- Landete, J.M.; Curiel, J.A.; Rodríguez, H.; de las Rivas, B.; Muñoz, R. Aryl glycosidases from Lactobacillus plantarum increase antioxidant activity of phenolic compounds. J. Funct. Foods 2014, 7, 322–329. [Google Scholar] [CrossRef]
- Michlmayr, H.; Kneifel, W. β-Glucosidase activities of lactic acid bacteria: Mechanisms, impact on fermented food and human health. FEMS Microbiol. Lett. 2014, 352, 1–10. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Microorganism | Time (min) | FCRS (mg GAE/g d.w.) | Antioxidant Activity (mg TE/g d.w.) | |
|---|---|---|---|---|
| DPPH | FRAP | |||
| L. mesenteroides 215 | 24 | 17.34 ± 0.30 h | 27.84 ± 0.82 a,b | 57.86 ± 0.65 c,d |
| 48 | 18.94 ± 0.13 g | 30.30 ± 0.06 b–e | 58.40 ± 0.97 d–f | |
| 72 | 20.23 ± 0.42 f | 33.34 ± 1.25 g–i | 72.96 ± 1.23 j | |
| 96 | 18.84 ± 0.33 g | 32.18 ± 1.27 e–g | 70.95 ± 1.21 j | |
| L. brevis 4121T | 24 | 20.06 ± 0.16 f | 35.68 ± 0.52 i,j | 62.31 ± 1.04 e–h |
| 48 | 19.85 ± 0.18 g | 33.17 ± 0.01 f– h | 58.29 ± 0.99 d,e | |
| 72 | 17.83 ± 0.19 h | 30.32 ± 0.70 c–e | 58.53 ± 0.99 d–f | |
| 96 | 19.31 ± 0.18 g | 29.23 ± 0.33 b–d | 60.57 ± 1.02 d–g | |
| L. brevis 5354 | 24 | 29.39 ± 0.60 a | 47.20 ± 1.54 r,s | 91.58 ± 1.53 p,q |
| 48 | 27.62 ± 0.50 b | 43.78 ± 1.27 o–q | 86.34 ± 1.49 m–o | |
| 72 | 25.67 ± 0.42 c,d | 40.29 ± 1.36 l,m | 81.48 ± 1.39 k,l | |
| 96 | 19.87 ± 0.24 g | 31.64 ± 0.89 d–g | 63.43 ± 1.08 g,h | |
| L. plantarum 748T | 24 | 21.98 ± 0.12 e | 25.56 ± 0.46 a | 71.58 ± 1.24 j |
| 48 | 30.72 ± 0.52 a | 44.53 ± 1.00 p,q | 96.61 ± 1.60 r | |
| 72 | 30.07 ± 0.52 a | 43.35 ± 0.92 o–q | 92.01 ± 1.55 p,q | |
| 96 | 26.62 ± 0.40 c | 38.59 ± 1.04 k,l | 85.14 ± 1.45 l–n | |
| L. plantarum 9567 | 24 | 29.09 ± 0.02 a | 49.68 ± 0.78 t–v | 90.38 ± 1.57 o,p,q |
| 48 | 27.65 ± 0.10 b,c | 42.74 ± 0.39 m,n | 93.23 ± 1.54 q,r | |
| 72 | 28.50 ± 0.30 b | 43.59 ± 0.66 o–q | 89.72 ± 1.51 o–q | |
| 96 | 27.08 ± 0.32 c | 40.82 ± 0.75 l–n | 89.10 ± 1.50 n–p | |
| P. acidilactici 5765T | 24 | 29.56 ± 0.37 a | 51.32 ± 0.37 v | 78.50 ± 1.35 k |
| 48 | 25.87 ± 0.08 c,d | 50.01 ± 0.23 u,v | 70.31 ± 1.19 j | |
| 72 | 24.49 ± 0.09 d | 42.85 ± 0.27 n–p | 50.34 ± 0.84 a | |
| 96 | 22.80 ± 0.05 e | 45.45 ± 0.37 q,r | 71.12 ± 1.20 j | |
| P. acidilactici 98 | 24 | 17.77 ± 0.01 h | 28.90 ± 0.07 b,c | 53.59 ± 0.90 a,b |
| 48 | 20.10 ± 0.04 f | 33.09 ± 0.07 f–h | 56.66 ± 0.96 b–d | |
| 72 | 18.35 ± 0.53 g | 32.11 ± 1.28 e–g | 72.28 ± 1.21 j | |
| 96 | 18.58 ± 0.21 g | 31.33 ± 0.36 c–g | 64.51 ± 1.08 g,h | |
| P. pentosaceus 4695T | 24 | 27.70 ± 0.20 b,c | 50.04 ± 0.56 u,v | 93.33 ± 1.57 q,r |
| 48 | 21.49 ± 0.24 e | 48.03 ± 0.55 s–u | 82.79 ± 1.41 l,m | |
| 72 | 21.22 ± 0.16 e | 36.45 ± 0.41 j,k | 65.64 ± 1.11 h,i | |
| 96 | 17.46 ± 0.13 h | 36.63 ± 0.94 j,k | 53.82 ± 0.93 a–c | |
| P. pentosaceus 923 | 24 | 20.21 ± 0.02 f | 42.56 ± 0.74 m–p | 62.30 ± 1.03 e–h |
| 48 | 21.60 ± 0.08 e | 41.38 ± 0.28 m–o | 68.89 ± 1.14 i,j | |
| 72 | 23.41 ± 0.21 d,e | 48.79 ± 0.28 s–u | 78.61 ± 1.35 k | |
| 96 | 23.53 ± 0.59 d | 49.70 ± 0.27 t–v | 83.41 ± 1.41 l,m | |
| L. mesenteroides 219T | 24 | 20.49 ± 0.43 f | 47.47 ± 0.32 r–t | 72.51 ± 1.20 j |
| 48 | 18.94 ± 0.25 g | 43.39 ± 1.09 o–q | 64.97 ± 1.08 h,i | |
| 72 | 20.08 ± 0.01 f | 35.38 ± 0.29 h–j | 62.42 ± 1.04 f–h | |
| 96 | 18.88 ± 0.11 g | 30.79 ± 0.50 c–f | 57.49 ± 1.00 b–d | |
| Control | - | 26.90 ± 0.04 b,c | 53.88 ± 0.58 w | 92.10 ± 1.26 p,q |
| Peak | Time (min) | m/z Experimental | m/z Calculated | Error (ppm) | Score (%) | Molecular Formula | Compound |
|---|---|---|---|---|---|---|---|
| 1 | 0.324 | 191.0554 | 191.0556 | −1.0 | 100 | C7H12O6 | Quinic acid isomer a |
| 2 | 0.394 | 191.0549 | 191.0556 | −3.7 | 100 | C7H12O6 | Quinic acid isomer b |
| 3 | 1.362 | 315.0703 | 315.0716 | −4.1 | 99.97 | C13H16O9 | Protocatechuic acid-4-glucoside |
| 4 | 3.406 | 163.0398 | 163.0395 | 1.8 | 100 | C9H8O3 | p-coumaric acid |
| 5 | 4.688 | 353.0865 | 353.0873 | −2.3 | 99.97 | C16H18O9 | Chlorogenic acid |
| 6 | 5.172 | 371.0977 | 371.0978 | −0.3 | 95.46 | C16H20O10 | Dihydroferulic acid 4-O-glucuronide |
| 7 | 5.478 | 385.1153 | 385.1135 | 4.7 | 90.68 | C17H22O10 | sinapic acid-C-hexoside |
| 8 | 5.644 | 577.1351 | 577.1346 | 0.9 | 99.3 | C30H26O12 | Procyanidin dimer |
| 9 | 6.016 | 221.044 | 221.045 | −4.5 | 100 | C11H10O5 | p-Coumaroyl glycolic acid |
| 10 | 7.138 | 165.0547 | 165.0552 | −3.0 | 100 | C9H10O3 | Dihydro-p-coumaric acid |
| 11 | 7.199 | 865.1981 | 865.198 | 0.1 | 91.8 | C45H38O18 | Procyanidin trimer |
| 12 | 7.568 | 301.0342 | 301.0348 | −2.0 | 96.02 | C15H10O7 | Quercetin |
| 13 | 7.903 | 625.1406 | 625.1405 | 0.2 | 98.96 | C27H30O17 | Quercetin-diglucoside isomer a |
| 14 | 7.973 | 739.1682 | 739.1663 | 2.6 | 90.78 | C39H32O15 | Cinchonain-1a-(4beta->8)-catechin isomer a |
| 15 | 8.048 | 625.1428 | 625.1405 | 3.7 | 99.53 | C27H30O17 | Quercetin-diglucoside isomer b |
| 16 | 8.147 | 451.1015 | 451.1029 | −3.1 | 99.51 | C24H20O9 | Cinchonain |
| 17 | 8.263 | 739.1646 | 739.1663 | −2.3 | 95.25 | C39H32O15 | Cinchonain-1a-(4beta->8)-catechin isomer b |
| 18 | 8.531 | 595.1297 | 595.1299 | −0.3 | 94.55 | C26H28O16 | Quercetin-3-O-arabinosyl-glucoside isomer a |
| 19 | 8.668 | 595.1292 | 595.1299 | −1.17 | 95.67 | C26H28O16 | Quercetin-3-O-arabinosyl-glucoside isomer b |
| 20 | 8.788 | 609.146 | 609.1456 | 0.7 | 99.01 | C27H30O16 | Rutin isomer a |
| 21 | 8.966 | 609.1456 | 609.1456 | 0.0 | 96.23 | C27H30O16 | Rutin isomer b |
| 22 | 9.07 | 595.1286 | 595.1299 | −2.2 | 99.57 | C26H28O16 | Quercetin-3-O-arabinosyl-glucoside isomer c |
| 23 | 9.144 | 463.0862 | 463.0877 | −3.2 | 94.2 | C21H20O12 | Quercetin-3-glucoside isomer a |
| 24 | 9.314 | 463.0866 | 463.0877 | −2.4 | 99.16 | C21H20O12 | Quercetin-3-glucoside isomer b |
| 25 | 9.57 | 579.1331 | 579.135 | −3.3 | 96.32 | C26H28O15 | Luteolin 7-O-(2″-O-pentosyl)-hexoside isomer a |
| 26 | 9.624 | 299.0183 | 299.0192 | −3.0 | 98.09 | C15H8O7 | Emodic acid isomer a |
| 27 | 9.645 | 477.0653 | 477.0669 | −3.4 | 99.22 | C21H18O13 | Quercetin glucuronide |
| 28 | 9.715 | 579.135 | 579.135 | 0.0 | 91.62 | C26H28O15 | Luteolin 7-O-(2″-O-pentosyl)-hexoside isomer b |
| 29 | 9.765 | 565.1204 | 565.1193 | 1.9 | 92.13 | C25H26O15 | Quercetin 3-xilosyl-(1->2)-alpha-L-arabinopyranoside |
| 30 | 9.901 | 447.0918 | 447.0927 | −2.0 | 94.92 | C21H20O11 | Kaempferol-O-hexoside isomer a |
| 31 | 9.992 | 609.1456 | 609.1456 | 0.0 | 99.88 | C27H30O16 | Rutin isomer c |
| 32 | 10.06 | 299.0192 | 299.0192 | 0.0 | 99.87 | C15H8O7 | Emodic acid isomer b |
| 33 | 10.087 | 579.1343 | 579.135 | −1.2 | 92.58 | C26H28O15 | Luteolin 7-O-(2″-O-pentosyl)-hexoside isomer c |
| 34 | 10.244 | 447.0915 | 447.0927 | −2.7 | 93.19 | C21H20O11 | Kaempferol-O-hexoside isomer b |
| 35 | 10.311 | 505.096 | 505.0982 | −4.4 | 96.8 | C23H22O13 | Quercetin 3-O-glucose-6″-acetate |
| 36 | 10.451 | 447.0913 | 447.0927 | −3.1 | 93.55 | C21H20O11 | Quercetin-O-deoxyhesoxide |
| 37 | 10.493 | 461.0706 | 461.0779 | −3.0 | 99.87 | C21H18O12 | Kaempferol 3-glucuronide |
| 38 | 10.567 | 549.124 | 549.1244 | −0.7 | 92.41 | C25H26O14 | Kaempferol 3,4′-dixyloside isomer b |
| 39 | 10.77 | 549.1262 | 549.1244 | 3.3 | 93.06 | C25H26O14 | Kaempferol 3,4′-dixyloside isomer a |
| 40 | 10.845 | 579.136 | 579.135 | 1.7 | 94.44 | C26H28O15 | Luteolin 7-O-(2″-O-pentosyl)-hexoside isomer d |
| 41 | 10.944 | 593.1521 | 593.1506 | 2.5 | 99.34 | C27H30O15 | Catechin diglucopyranoside |
| 42 | 11.022 | 489.1032 | 489.1033 | −0.2 | 93.58 | C23H22O12 | Quercetin 3-O-acetyl-rhamnoside |
| 43 | 11.266 | 531.1507 | 531.1503 | 0.8 | 99.48 | C26H28O12 | Luteolin 7-[6-O-(2-methylbutyryl)-beta-glucoside] isomer a |
| 44 | 11.291 | 489.1024 | 489.1033 | −1.8 | 98.79 | C23H22O12 | Kaempferol 3-O-acetyl-glucoside |
| 45 | 11.374 | 531.15 | 531.1503 | −0.6 | 94.94 | C26H28O12 | Luteolin 7-[6-O-(2-methylbutyryl)-beta-glucoside] isomer b |
| 46 | 11.618 | 431.0989 | 431.0978 | 2.6 | 93.85 | C21H20O10 | Kaempferol-O-coumaroyl |
| 47 | 11.808 | 563.1408 | 563.1401 | 1.2 | 94.47 | C26H28O14 | Apigenin-C-hexoside-C-pentoside |
| 48 | 12.313 | 285.0394 | 285.0399 | −1.8 | 90.15 | C15H10O6 | Kaempferol |
| µg/g d.w. | ||||||
|---|---|---|---|---|---|---|
| Compound | P. acidilactici CECT 5765T | P. pentosaceus CECT 4695T | L. brevis CECT 5354 | L. plantarum CECT 748T | L. plantarum CECT 9567 | Control |
| Protocatechuic acid-4-glucoside | 235.18 ± 10.3 a | 364.60 ± 7.40 c | 401.68 ± 7.01 d | <LOQ | 384.61 ± 7.15 d | 301.93 ± 2.66 b |
| Chlorogenic acid | 37.84 ± 1.25 a | 93.16 ± 0.66 b | 194.14 ± 3.75 c | 201.66 ± 3.66 c | 95.63 ± 2.39 b | 310.69 ± 5.89 d |
| Dihydroferulic acid 4-O-glucuronide | 486.77 ± 6.16 b | 766.37 ± 16.91 d | 741.21 ± 6.43 d | 419.82 ± 14.40 a | 657.75 ± 6.57 c | 516.74 ± 13.19 b |
| Sinapic acid-C-hexoside | 115.99 ± 2.61 c | 132.64 ± 2.93 d | 129.81 ± 4.43 d | 71.78 ± 1.16 a | 120.71 ± 2.40 c | 81.34 ± 1.54 b |
| p-Coumaric acid | 185.67 ± 5.25 d | 200.73 ± 0.05 e | 169.85 ± 2.25 c | <LOQ | 140.29 ± 2.42 a | 147.97 ± 1.55 b |
| p-Coumaroyl glycolic acid | 42.16 ± 1.94 d | 56.81 ± 3.41 e | 35.88 ± 0.36 c | <LOQ | 30.27 ± 0.74 b | 24.05 ± 0.55 a |
| Dihydro-p-coumaric acid | 191.22 ± 4.13 d | 230.99 ± 2.98 e | 186.30 ± 2.94 d | 165.49 ± 0.42 c | 140.21 ± 5.34 b | 121.23 ± 1.94 a |
| Cinchonain | <LOQ | <LOQ | <LOQ | <LOQ | <LOQ | <LOQ |
| Cinchonain-1a-(4beta->8)-catechin isomer a | <LOQ | <LOQ | <LOQ | <LOQ | <LOQ | <LOQ |
| Cinchonain-1a-(4beta->8)-catechin isomer b | <LOQ | <LOQ | <LOQ | <LOQ | <LOQ | <LOQ |
| Procyanidin dimer | <LOQ | <LOQ | <LOQ | <LOQ | <LOQ | <LOQ |
| Procyanidin trimer | <LOQ | <LOQ | <LOQ | <LOQ | <LOQ | <LOQ |
| Catechin diglucopyranoside | 174.76 ± 2.87 a,b | 231.20 ± 14.79 c | 168.59 ± 8.56 a,b | 156.56 ± 2.79 a | 179.05 ± 0.94 b | 173.43 ± 2.46 a,b |
| Quercetin | <LOQ | <LOQ | <LOQ | <LOQ | <LOQ | <LOQ |
| Quercetin-diglucoside isomer a | 396.74 ± 5.37 e | 370.98 ± 11.08 d | 558.21 ± 4.90 f | 277.86 ± 9.81 b | 324.16 ± 5.66 c | 214.69 ± 4.12 a |
| Quercetin-diglucoside isomer b | 217.72 ± 10.52 d | 170.00 ± 3.00 c | 262.31 ± 3.76 e | 124.72 ± 7.42 b | 176.52 ± 1.49 c | 98.53 ± 6.75 a |
| Quercetin-3-O-arabinosyl-glucoside isomer a | 498.00 ± 79.73 d | 394.58 ± 8.20 c | 519.29 ± 8.56 d | 270.03 ± 14.03 b | 223.42 ± 10.62 a | 269.02 ± 0.79 b |
| Quercetin-3-O-arabinosyl-glucoside isomer b | 302.52 ± 10.79 d | 284.07 ± 8.30 d | 291.30 ± 7.83 d | 103.57 ± 8.68 a | 152.27 ± 10.32 c | 126.63 ± 2.70 b |
| Quercetin-3-O-arabinosyl-glucoside isomer c | <LOQ | <LOQ | <LOQ | <LOQ | <LOQ | <LOQ |
| Quercetin-3-glucoside isomer a | 248.31 ± 10.50 c | 317.37 ± 16.93 e | 272.94 ± 2.03 d | 161.98 ± 4.52 ab | 191.52 ± 4.31 | 153.37 ± 1.88 a |
| Quercetin-3-glucoside isomer b | 281.07 ± 16.50 c | 291.79 ± 15.11 c | 225.80 ± 2.41 b | 15.13 ± 2.87 a | 237.81 ± 0.13 b | 221.66 ± 12.70 b |
| Quercetin glucuronide | 244.54 ± 4.62 a | 245.23 ± 9.88 a | 310.17 ± 15.05 b | 234.06 ± 6.78 a | 225.18 ± 3.68 a | 312.69 ± 11.19 b |
| Quercetin 3-apiosyl-(1->2)-alpha-L-arabinopyranoside | <LOQ | <LOQ | <LOQ | <LOQ | <LOQ | <LOQ |
| Quercetin-O-deoxyhesoxide | 223.65 ± 8.65 e | 233.55 ± 2.55 e | 123.24 ± 1.33 c | 75.82 ± 6.74 a | 140.25 ± 3.27 d | 102.90 ± 2.56 b |
| Quercetin 3-O-glucose-6″-acetate | <LOQ | <LOQ | <LOQ | <LOQ | <LOQ | <LOQ |
| Quercetin 3-O-acetyl-rhamnoside | <LOQ | <LOQ | <LOQ | <LOQ | <LOQ | <LOQ |
| Rutin isomer a | 348.26 ± 1.27 e | 350.88 ± 3.37 e | 324.72 ± 5.94 d | 117.66 ± 8.92 a | 219.94 ± 9.95 c | 155.12 ± 10.53 b |
| Rutin isomer b | 281.89 ± 0.47 f | 257.55 ± 5.62 e | 232.75 ± 6.29 d | 54.63 ± 5.25 a | 157.58 ± 7.31 c | 94.67 ± 0.40 b |
| Rutin isomer c | 557.92 ± 1.75 e | 473.91 ± 11.52 c | 498.48 ± 11.19 c | 426.23 ± 10.28 b | 527.58 ± 3.41 d | 332.33 ± 13.44 a |
| Luteolin 7-O-(2″-O-pentosyl)-hexoside isomer a | 184.54 ± 11.24 d | 124.92 ± 9.70 c | 88.77 ± 1.71 b | 97.85 ± 0.37 b | 82.09 ± 2.83 b | 17.24 ± 3.20 a |
| Luteolin 7-O-(2″-O-pentosyl)-hexoside isomer b | <LOQ | <LOQ | <LOQ | <LOQ | <LOQ | <LOQ |
| Luteolin 7-O-(2″-O-pentosyl)-hexoside isomer c | <LOQ | <LOQ | <LOQ | <LOQ | <LOQ | <LOQ |
| Luteolin 7-O-(2″-O-pentosyl)-hexoside isomer d | 146.00 ± 45.85 c | 181.60 ± 14.25 d | 108.99 ± 3.12 b | 69.94 ± 7.67 a | 169.59 ± 1.49 d | 130.81 ± 3.40 c |
| Luteolin 7-[6-O-(2-methylbutyryl)-beta-glucoside] isomer a | <LOQ | <LOQ | <LOQ | <LOQ | <LOQ | <LOQ |
| Luteolin 7-[6-O-(2-methylbutyryl)-beta-glucoside] isomer b | <LOQ | <LOQ | <LOQ | <LOQ | <LOQ | <LOQ |
| Kaempferol | 32.04 ± 3.57 b | 232.24 ± 25.93 c | 26.53 ± 4.71 a,b | <LOQ | 2.82 ± 0.78 a,b | 23.49 ± 0.99 a,b |
| Kaempferol-O-hexoside isomer a | 309.41 ± 8.65 f | 219.59 ± 9.05 d | 258.68 ± 2.99 e | 94.00 ± 6.02 a | 148.73 ± 0.92 c | 120.91 ± 7.55 b |
| Kaempferol-O-hexoside isomer b | 258.40 ± 7.65 c | 250.72 ± 11.78 c | 177.49 ± 2.89 b | <LOQ | 156.92 ± 9.50 b | 116.48 ± 6.57 a |
| Kaempferol 3-glucuronide | 154.26 ± 4.36 e | 171.12 ± 1.78 f | 101.26 ± 0.92 d | 31.31 ± 5.45 b | 40.93 ± 2.45 c | 15.95 ± 2.62 a |
| Kaempferol 3,4′-dixyloside isomer a | <LOQ | <LOQ | <LOQ | <LOQ | <LOQ | <LOQ |
| Kaempferol 3,4′-dixyloside isomer b | <LOQ | <LOQ | <LOQ | <LOQ | <LOQ | <LOQ |
| Kaempferol 3-O-acetyl-glucoside | <LOQ | <LOQ | <LOQ | <LOQ | <LOQ | <LOQ |
| Kaempferol-O-coumaroyl | 38.94 ± 4.85 c | 135.91 ± 3.08 d | 8.34 ± 1.69 a | 34.16 ± 5.92 c | 21.29 ± 7.20 | 8.27 ± 1.39 ab |
| Apigenin-C-hexoside-C-pentoside | 63.37 ± 8.24 c | 137.46 ± 3.42 d | 36.65 ± 4.60 b | <LOQ | 17.34 ± 4.26 a | 22.80 ± 1.98 a |
| Sum of phenolic acids | 1445.14 ± 32.50 b | 1976.73 ± 30.92 e | 2185.23 ± 27.17 f | 886.58 ± 19.65 a | 1686.24 ± 27.02 d | 1557.08 ± 27.32 c |
| Sum of flavonoids | 4937.17 ± 113.27 c,d | 5026.32 ± 190.43 d | 4592.16 ± 105.17 c | 2355.58 ± 113.52 a | 3417.55 ± 90.51 b | 2606.36 ± 97.24 a |
| Sum of phenolic compounds | 6382.32 ± 145.78 d | 7003.05 ± 221.35 e | 6777.39 ± 132.34 d,e | 3242.17 ± 133.17 a | 5103.78 ± 117.53 c | 4163.45 ± 124.56 b |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
De Montijo-Prieto, S.; Razola-Díaz, M.d.C.; Barbieri, F.; Tabanelli, G.; Gardini, F.; Jiménez-Valera, M.; Ruiz-Bravo, A.; Verardo, V.; Gómez-Caravaca, A.M. Impact of Lactic Acid Bacteria Fermentation on Phenolic Compounds and Antioxidant Activity of Avocado Leaf Extracts. Antioxidants 2023, 12, 298. https://doi.org/10.3390/antiox12020298
De Montijo-Prieto S, Razola-Díaz MdC, Barbieri F, Tabanelli G, Gardini F, Jiménez-Valera M, Ruiz-Bravo A, Verardo V, Gómez-Caravaca AM. Impact of Lactic Acid Bacteria Fermentation on Phenolic Compounds and Antioxidant Activity of Avocado Leaf Extracts. Antioxidants. 2023; 12(2):298. https://doi.org/10.3390/antiox12020298
Chicago/Turabian StyleDe Montijo-Prieto, Soumi, María del Carmen Razola-Díaz, Federica Barbieri, Giulia Tabanelli, Fausto Gardini, Maria Jiménez-Valera, Alfonso Ruiz-Bravo, Vito Verardo, and Ana Mª Gómez-Caravaca. 2023. "Impact of Lactic Acid Bacteria Fermentation on Phenolic Compounds and Antioxidant Activity of Avocado Leaf Extracts" Antioxidants 12, no. 2: 298. https://doi.org/10.3390/antiox12020298
APA StyleDe Montijo-Prieto, S., Razola-Díaz, M. d. C., Barbieri, F., Tabanelli, G., Gardini, F., Jiménez-Valera, M., Ruiz-Bravo, A., Verardo, V., & Gómez-Caravaca, A. M. (2023). Impact of Lactic Acid Bacteria Fermentation on Phenolic Compounds and Antioxidant Activity of Avocado Leaf Extracts. Antioxidants, 12(2), 298. https://doi.org/10.3390/antiox12020298