
Brain Iron Homeostasis and Mental Disorders



Abstract
1. Introduction
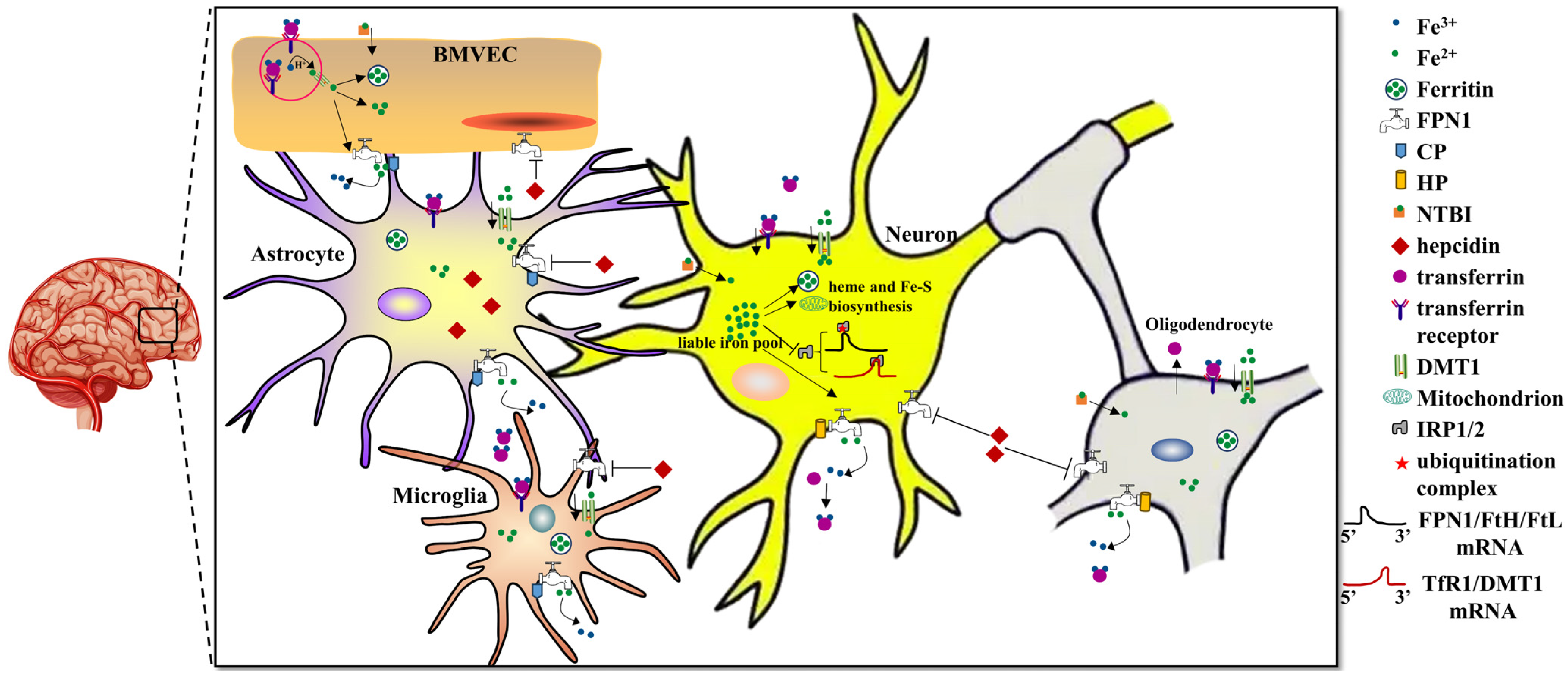
2. Regulation of Brain Iron Homeostasis
2.1. Iron Uptake into the Brain
2.2. Iron Export from the Brain
2.3. Regulation of Iron in the Brain
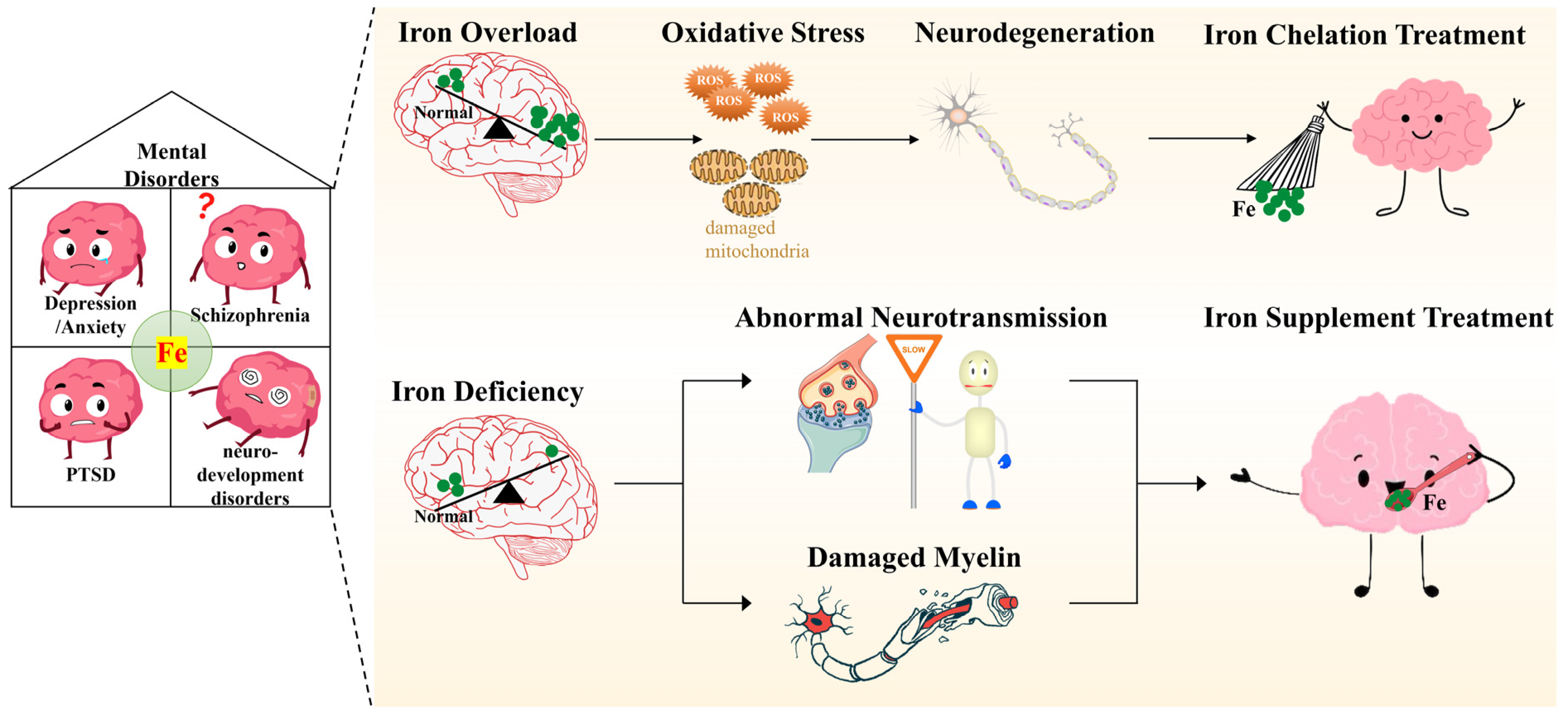
3. Iron and Mental Disorders
3.1. Depression/Anxiety Disorders
3.1.1. Iron Deficiency-Related Depression/Anxiety Disorders
3.1.2. Iron Overload-Related Depression/Anxiety Disorders
3.2. Schizophrenia
3.2.1. Iron Deficiency-Related Schizophrenia
3.2.2. Iron Overload-Related Schizophrenia
3.3. Post-Traumatic Stress Disorder (PTSD)
3.3.1. Iron Deficiency in the Pathology of PTSD
3.3.2. Iron Overload in the Pathology of PTSD and Fear Memory
3.4. Neurodevelopment Disorders
3.4.1. Brain Iron and ADHD
3.4.2. Brain Iron and ASD
4. Potential Mechanisms Underlying the Influence of Iron on Mental Disorders
4.1. Iron and Neurotransmission
4.2. Iron and Oxidative Stress
4.3. Iron and Myelination
5. Limitations of the Presented Studies
6. Conclusions and Prospects
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Solmi, M.; Soardo, L.; Kaur, S.; Azis, M.; Cabras, A.; Censori, M.; Fausti, L.; Besana, F.; Salazar de Pablo, G.; Fusar-Poli, P. Meta-analytic prevalence of comorbid mental disorders in individuals at clinical high risk of psychosis: The case for transdiagnostic assessment. Mol. Psychiatry 2023, 28, 2291–2300. [Google Scholar] [CrossRef] [PubMed]
- Hyman, S.; Chisholm, D.; Kessler, R.; Patel, V.; Whiteford, H. Mental Disorders. Disease Control Priorities Related to Mental, Neurological, Developmental and Substance Abuse Disorders; World Health Organization: Geneva, Switzerland, 2006; pp. 1–20. [Google Scholar]
- Appelbaum, L.G.; Shenasa, M.A.; Stolz, L.; Daskalakis, Z. Synaptic plasticity and mental health: Methods, challenges and opportunities. Neuropsychopharmacology 2023, 48, 113–120. [Google Scholar] [CrossRef]
- Craske, M.G.; Herzallah, M.M.; Nusslock, R.; Patel, V. From neural circuits to communities: An integrative multidisciplinary roadmap for global mental health. Nat. Ment. Health 2023, 1, 12–24. [Google Scholar] [CrossRef]
- Tong, Q.; Cui, X.; Xu, H.; Zhang, X.; Hu, S.; Huang, F.; Xiao, L. D1 receptor-expressing neurons in ventral tegmental area alleviate mouse anxiety-like behaviors via glutamatergic projection to lateral septum. Mol. Psychiatry 2023, 28, 625–638. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.; Pan, X.; Zhou, Y.; Wu, Z.; Ren, K.; Liu, H.; Huang, C.; Yu, Y.; He, T.; Zhang, X.; et al. Lateral septum-lateral hypothalamus circuit dysfunction in comorbid pain and anxiety. Mol. Psychiatry 2023, 28, 1090–1100. [Google Scholar] [CrossRef] [PubMed]
- Breton, J.M.; Barraza, M.; Hu, K.Y.; Frias, S.J.; Long, K.L.; Kaufer, D. Juvenile exposure to acute traumatic stress leads to long-lasting alterations in grey matter myelination in adult female but not male rats. Neurobiol. Stress 2021, 14, 100319. [Google Scholar] [CrossRef]
- Boda, E. Myelin and oligodendrocyte lineage cell dysfunctions: New players in the etiology and treatment of depression and stress-related disorders. Eur. J. Neurosci. 2021, 53, 281–297. [Google Scholar] [CrossRef]
- Guo, Y.; Wu, H.; Dong, D.; Zhou, F.; Li, Z.; Zhao, L.; Long, Z. Stress and the brain: Emotional support mediates the association between myelination in the right supramarginal gyrus and perceived chronic stress. Neurobiol. Stress 2023, 22, 100511. [Google Scholar] [CrossRef]
- Zlomuzica, A.; Plank, L.; Kodzaga, I.; Dere, E. A fatal alliance: Glial connexins, myelin pathology and mental disorders. J. Psychiatr. Res. 2023, 159, 97–115. [Google Scholar] [CrossRef]
- Chakraborty, P.; Dey, A.; Gopalakrishnan, A.V.; Swati, K.; Ojha, S.; Prakash, A.; Kumar, D.; Ambasta, R.K.; Jha, N.K.; Jha, S.K.; et al. Glutamatergic neurotransmission: A potential pharmacotherapeutic target for the treatment of cognitive disorders. Ageing Res. Rev. 2023, 85, 101838. [Google Scholar] [CrossRef]
- Ochi, T.; Vyalova, N.M.; Losenkov, I.S.; Paderina, D.Z.; Pozhidaev, I.V.; Loonen, A.J.; Simutkin, G.G.; Bokhan, N.A.; Wilffert, B.; Ivanova, S.A. Polymorphisms in the adrenergic neurotransmission pathway impact antidepressant response in depressed patients. Neurosci. Appl. 2023, 2, 101016. [Google Scholar] [CrossRef]
- Wang, F.; Yang, X.; Ren, Z.; Chen, C.; Liu, C. Alternative splicing in mouse brains affected by psychological stress is enriched in the signaling, neural transmission and blood-brain barrier pathways. Mol. Psychiatry, 2023; online ahead of print. [Google Scholar]
- Ward, R.J.; Zucca, F.A.; Duyn, J.H.; Crichton, R.R.; Zecca, L. The role of iron in brain ageing and neurodegenerative disorders. Lancet Neurol. 2014, 13, 1045–1060. [Google Scholar] [CrossRef] [PubMed]
- Hsu, C.C.; Senussi, N.H.; Fertrin, K.Y.; Kowdley, K.V. Iron overload disorders. Hepatol. Commun. 2022, 6, 1842–1854. [Google Scholar] [CrossRef] [PubMed]
- Gao, G.; You, L.; Zhang, J.; Chang, Y.Z.; Yu, P. Brain Iron Metabolism, Redox Balance and Neurological Diseases. Antioxidants 2023, 12, 1289. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.D.; Li, H.; Kang, S.; Cui, Y.G.; Zheng, H.; Wang, P.; Han, K.; Yu, P.; Chang, Y.Z. The divergent effects of astrocyte ceruloplasmin on learning and memory function in young and old mice. Cell Death Dis. 2022, 13, 1006. [Google Scholar] [CrossRef] [PubMed]
- Hu, S.; Lin, S.; He, X.; Sun, N. Iron delivery systems for controlled release of iron and enhancement of iron absorption and bioavailability. Crit. Rev. Food Sci. Nutr. 2022, 1–20. [Google Scholar] [CrossRef]
- Murray-Kolb, L.E. Iron and brain functions. Curr. Opin. Clin. Nutr. Metab. Care 2013, 16, 703–707. [Google Scholar] [CrossRef]
- Hua, M.; Shi, D.; Xu, W.; Zhu, L.; Hao, X.; Zhu, B.; Shu, Q.; Lozoff, B.; Geng, F.; Shao, J. Differentiation between fetal and postnatal iron deficiency in altering brain substrates of cognitive control in pre-adolescence. BMC Med. 2023, 21, 167. [Google Scholar] [CrossRef]
- Andrade, C. Antidepressant Prescription in Pregnancy: The Importance of Prenatal Maternal Anemia as a Potential Confound in Studies on Neurodevelopmental and Other Outcomes. J. Clin. Psychiatry 2020, 81, 20f13347. [Google Scholar] [CrossRef]
- Vigani, G.; Murgia, I. Iron-requiring enzymes in the spotlight of oxygen. Trends Plant Sci. 2018, 23, 874–882. [Google Scholar] [CrossRef]
- Todorich, B.; Pasquini, J.M.; Garcia, C.I.; Paez, P.M.; Connor, J.R. Oligodendrocytes and myelination: The role of iron. Glia 2009, 57, 467–478. [Google Scholar] [CrossRef] [PubMed]
- Berthou, C.; Iliou, J.P.; Barba, D. Iron, neuro-bioavailability and depression. eJHaem 2022, 3, 263–275. [Google Scholar] [CrossRef] [PubMed]
- Nave, K.A.; Ehrenreich, H. Myelination and oligodendrocyte functions in psychiatric diseases. JAMA Psychiatry 2014, 71, 582–584. [Google Scholar] [CrossRef] [PubMed]
- Wu, Q.; Hao, Q.; Li, H.; Wang, B.; Wang, P.; Jin, X.; Yu, P.; Gao, G.; Chang, Y.Z. Brain iron deficiency and affected contextual fear memory in mice with conditional Ferroportin1 ablation in the brain. FASEB J. 2021, 35, e21174. [Google Scholar] [CrossRef]
- Qian, Z.M.; Ke, Y. Brain iron transport. Biol. Rev. Camb. Philos. Soc. 2019, 94, 1672–1684. [Google Scholar] [CrossRef]
- Baringer, S.L.; Simpson, I.A.; Connor, J.R. Brain iron acquisition: An overview of homeostatic regulation and disease dysregulation. J. Neurochem. 2023, 165, 625–642. [Google Scholar] [CrossRef]
- Abbott, N.J.; Patabendige, A.A.; Dolman, D.E.; Yusof, S.R.; Begley, D.J. Structure and function of the blood-brain barrier. Neurobiol. Dis. 2010, 37, 13–25. [Google Scholar] [CrossRef]
- Abbott, N.J.; Ronnback, L.; Hansson, E. Astrocyte-endothelial interactions at the blood-brain barrier. Nat. Rev. Neurosci. 2006, 7, 41–53. [Google Scholar] [CrossRef]
- McCarthy, R.C.; Kosman, D.J. Mechanistic analysis of iron accumulation by endothelial cells of the BBB. Biometals Int. J. Role Met. Ions Biol. Biochem. Med. 2012, 25, 665–675. [Google Scholar] [CrossRef]
- Duck, K.A.; Simpson, I.A.; Connor, J.R. Regulatory mechanisms for iron transport across the blood-brain barrier. Biochem. Biophys. Res. Commun. 2017, 494, 70–75. [Google Scholar] [CrossRef]
- Rudisill, S.S.; Martin, B.R.; Mankowski, K.M.; Tessier, C.R. Iron Deficiency Reduces Synapse Formation in the Drosophila Clock Circuit. Biol. Trace Elem. Res. 2019, 189, 241–250. [Google Scholar] [CrossRef] [PubMed]
- Simpson, I.A.; Ponnuru, P.; Klinger, M.E.; Myers, R.L.; Devraj, K.; Coe, C.L.; Lubach, G.R.; Carruthers, A.; Connor, J.R. A novel model for brain iron uptake: Introducing the concept of regulation. J. Cereb. Blood Flow Metab. Off. J. Int. Soc. Cereb. Blood Flow Metab. 2015, 35, 48–57. [Google Scholar] [CrossRef] [PubMed]
- Khan, A.I.; Liu, J.; Dutta, P. Iron transport kinetics through blood-brain barrier endothelial cells. Biochim. Biophys. Acta. Gen. Subj. 2018, 1862, 1168–1179. [Google Scholar] [CrossRef] [PubMed]
- Jefferies, W.A.; Brandon, M.R.; Hunt, S.V.; Williams, A.F.; Gatter, K.C.; Mason, D.Y. Transferrin receptor on endothelium of brain capillaries. Nature 1984, 312, 162–163. [Google Scholar] [CrossRef] [PubMed]
- Bradbury, M.W. Transport of iron in the blood-brain-cerebrospinal fluid system. J. Neurochem. 1997, 69, 443–454. [Google Scholar] [CrossRef]
- Moos, T.; Morgan, E.H. Transferrin and transferrin receptor function in brain barrier systems. Cell. Mol. Neurobiol. 2000, 20, 77–95. [Google Scholar] [CrossRef]
- McCarthy, R.C.; Kosman, D.J. Ferroportin and exocytoplasmic ferroxidase activity are required for brain microvascular endothelial cell iron efflux. J. Biol. Chem. 2013, 288, 17932–17940. [Google Scholar] [CrossRef]
- McCarthy, R.C.; Kosman, D.J. Glial cell ceruloplasmin and hepcidin differentially regulate iron efflux from brain microvascular endothelial cells. PLoS ONE 2014, 9, e89003. [Google Scholar] [CrossRef]
- Faucheux, B.A.; Nillesse, N.; Damier, P.; Spik, G.; Mouatt-Prigent, A.; Pierce, A.; Leveugle, B.; Kubis, N.; Hauw, J.J.; Agid, Y.; et al. Expression of lactoferrin receptors is increased in the mesencephalon of patients with Parkinson disease. Proc. Natl. Acad. Sci. USA 1995, 92, 9603–9607. [Google Scholar] [CrossRef]
- Moroo, I.; Ujiie, M.; Walker, B.L.; Tiong, J.W.; Vitalis, T.Z.; Karkan, D.; Gabathuler, R.; Moise, A.R.; Jefferies, W.A. Identification of a novel route of iron transcytosis across the mammalian blood-brain barrier. Microcirculation 2003, 10, 457–462. [Google Scholar] [CrossRef]
- Malecki, E.A.; Devenyi, A.G.; Beard, J.L.; Connor, J.R. Existing and emerging mechanisms for transport of iron and manganese to the brain. J. Neurosci. Res. 1999, 56, 113–122. [Google Scholar] [CrossRef]
- Fishman, J.B.; Rubin, J.B.; Handrahan, J.V.; Connor, J.R.; Fine, R.E. Receptor-mediated transcytosis of transferrin across the blood-brain barrier. J. Neurosci. Res. 1987, 18, 299–304. [Google Scholar] [CrossRef] [PubMed]
- Qian, Z.M.; Chang, Y.Z.; Zhu, L.; Yang, L.; Du, J.R.; Ho, K.P.; Wang, Q.; Li, L.Z.; Wang, C.Y.; Ge, X.; et al. Development and iron-dependent expression of hephaestin in different brain regions of rats. J. Cell. Biochem. 2007, 102, 1225–1233. [Google Scholar] [CrossRef] [PubMed]
- McCarthy, R.C.; Kosman, D.J. Mechanisms and regulation of iron trafficking across the capillary endothelial cells of the blood-brain barrier. Front. Mol. Neurosci. 2015, 8, 31. [Google Scholar] [CrossRef] [PubMed]
- Andersen, H.H.; Johnsen, K.B.; Moos, T. Iron deposits in the chronically inflamed central nervous system and contributes to neurodegeneration. Cell. Mol. Life Sci. CMLS 2014, 71, 1607–1622. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Feng, R.; Liu, Q.; Feng, J.; Lao, G.; Zhang, M.; Li, J.; Zhang, Y.; Wei, H. APART-QSM: An improved sub-voxel quantitative susceptibility mapping for susceptibility source separation using an iterative data fitting method. Neuroimage 2023, 274, 120148. [Google Scholar] [CrossRef] [PubMed]
- Yu, P.; Chang, Y.Z. Brain Iron Metabolism and Regulation. Adv. Exp. Med. Biol. 2019, 1173, 33–44. [Google Scholar] [CrossRef]
- Zhang, X.; Li, L.; Qi, L.; Fu, Y.; Sun, D.; Chen, S.; Xu, W.; Liu, C.; Zhou, X.; He, G. Distribution pattern of iron deposition in the basal ganglia of different motor subtypes of Parkinson’s disease. Neurosci. Lett. 2023, 807, 137249. [Google Scholar] [CrossRef]
- Zhao, W.; Yang, C.; Tong, R.; Chen, L.; Chen, M.; Gillen, K.M.; Li, G.; Ma, C.; Wang, Y.; Wu, X.; et al. Relationship Between Iron Distribution in Deep Gray Matter Nuclei Measured by Quantitative Susceptibility Mapping and Motor Outcome After Deep Brain Stimulation in Patients With Parkinson’s Disease. J. Magn. Reson. Imaging 2023, 58, 581–590. [Google Scholar] [CrossRef]
- Hosseinpour Mashkani, S.M.; Bishop, D.P.; Raoufi-Rad, N.; Adlard, P.A.; Shimoni, O.; Golzan, S.M. Distribution of Copper, Iron, and Zinc in the Retina, Hippocampus, and Cortex of the Transgenic APP/PS1 Mouse Model of Alzheimer’s Disease. Cells 2023, 12, 1144. [Google Scholar] [CrossRef]
- Juan, S.M.A.; Daglas, M.; Gunn, A.P.; Lago, L.; Adlard, P.A. Characterization of the spatial distribution of metals and profile of metalloprotein complexes in a mouse model of repetitive mild traumatic brain injury. Metallomics 2022, 14, mfac092. [Google Scholar] [CrossRef] [PubMed]
- Ganz, T. Cellular iron: Ferroportin is the only way out. Cell Metab. 2005, 1, 155–157. [Google Scholar] [CrossRef] [PubMed]
- Wu, L.J.; Leenders, A.G.; Cooperman, S.; Meyron-Holtz, E.; Smith, S.; Land, W.; Tsai, R.Y.; Berger, U.V.; Sheng, Z.H.; Rouault, T.A. Expression of the iron transporter ferroportin in synaptic vesicles and the blood-brain barrier. Brain Res. 2004, 1001, 108–117. [Google Scholar] [CrossRef] [PubMed]
- You, L.; Yu, P.P.; Dong, T.; Guo, W.; Chang, S.; Zheng, B.; Ci, Y.; Wang, F.; Yu, P.; Gao, G.; et al. Astrocyte-derived hepcidin controls iron traffic at the blood-brain-barrier via regulating ferroportin 1 of microvascular endothelial cells. Cell Death Dis. 2022, 13, 667. [Google Scholar] [CrossRef]
- Raut, S.B.; Marathe, P.A.; van Eijk, L.; Eri, R.; Ravindran, M.; Benedek, D.M.; Ursano, R.J.; Canales, J.J.; Johnson, L.R. Diverse therapeutic developments for post-traumatic stress disorder (PTSD) indicate common mechanisms of memory modulation. Pharmacol. Ther. 2022, 239, 108195. [Google Scholar] [CrossRef]
- Muckenthaler, M.U.; Galy, B.; Hentze, M.W. Systemic iron homeostasis and the iron-responsive element/iron-regulatory protein (IRE/IRP) regulatory network. Annu. Rev. Nutr. 2008, 28, 197–213. [Google Scholar] [CrossRef]
- Hentze, M.W.; Muckenthaler, M.U.; Galy, B.; Camaschella, C. Two to tango: Regulation of Mammalian iron metabolism. Cell 2010, 142, 24–38. [Google Scholar] [CrossRef]
- Lotan, A.; Luza, S.; Opazo, C.M.; Ayton, S.; Lane, D.J.R.; Mancuso, S.; Pereira, A.; Sundram, S.; Weickert, C.S.; Bousman, C.; et al. Perturbed iron biology in the prefrontal cortex of people with schizophrenia. Mol. Psychiatry 2023, 28, 2058–2070. [Google Scholar] [CrossRef]
- Peng, Y.; Chang, X.; Lang, M. Iron Homeostasis Disorder and Alzheimer’s Disease. Int. J. Mol. Sci. 2021, 22, 12442. [Google Scholar] [CrossRef]
- Heidari, M.; Johnstone, D.M.; Bassett, B.; Graham, R.M.; Chua, A.C.G.; House, M.J.; Collingwood, J.F.; Bettencourt, C.; Houlden, H.; Ryten, M.; et al. Brain iron accumulation affects myelin-related molecular systems implicated in a rare neurogenetic disease family with neuropsychiatric features. Mol. Psychiatry 2016, 21, 1599–1607. [Google Scholar] [CrossRef]
- Xu, M.; Li, Y.; Meng, D.; Zhang, D.; Wang, B.; Xie, J.; Wang, J. 6-Hydroxydopamine Induces Abnormal Iron Sequestration in BV2 Microglia by Activating Iron Regulatory Protein 1 and Inhibiting Hepcidin Release. Biomolecules 2022, 12, 266. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization. Mental Disorders. Available online: https://www.who.int/news-room/fact-sheets/detail/mental-disorders (accessed on 14 May 2022).
- American Psychiatric Association. Diagnostic and Statistical Manual of Mental Disorders, 5th ed.; American Psychiatric Association: Washington, DC, USA, 2013. [Google Scholar]
- Wikipedia. Mental Disorder. Available online: https://en.wikipedia.org/wiki/Mental_disorder#cite_note-WHO2018-10 (accessed on 14 May 2022).
- Kennedy, S.H. Core symptoms of major depressive disorder: Relevance to diagnosis and treatment. Dialogues Clin. Neurosci. 2008, 10, 271–277. [Google Scholar] [CrossRef] [PubMed]
- Briley, P.M.; Webster, L.; Boutry, C.; Cottam, W.J.; Auer, D.P.; Liddle, P.F.; Morriss, R. Resting-state functional connectivity correlates of anxiety co-morbidity in major depressive disorder. Neurosci. Biobehav. Rev. 2022, 138, 104701. [Google Scholar] [CrossRef] [PubMed]
- Hawton, K.; Casanas, I.C.C.; Haw, C.; Saunders, K. Risk factors for suicide in individuals with depression: A systematic review. J. Affect. Disord. 2013, 147, 17–28. [Google Scholar] [CrossRef] [PubMed]
- Porras-Segovia, A.; Nobile, B.; Olie, E.; Gourguechon-Buot, E.; Garcia, E.B.; Gorwood, P.; Abascal-Peiro, S.; Courtet, P. Factors associated with transitioning from suicidal ideation to suicide attempt in the short-term: Two large cohorts of depressed outpatients. J. Affect Disord. 2023, 335, 155–165. [Google Scholar] [CrossRef] [PubMed]
- American Psychiatric Association. Anxiety Disorders: DSM-5® Selections; American Psychiatric Pub: Washington, DC, USA, 2015. [Google Scholar]
- Crocq, M.A. A history of anxiety: From Hippocrates to DSM. Dialogues Clin. Neurosci. 2015, 17, 319–325. [Google Scholar] [CrossRef]
- Hammen, C. Risk Factors for Depression: An Autobiographical Review. Annu. Rev. Clin. Psychol. 2018, 14, 1–28. [Google Scholar] [CrossRef]
- Shah, H.E.; Bhawnani, N.; Ethirajulu, A.; Alkasabera, A.; Onyali, C.B.; Anim-Koranteng, C.; Mostafa, J.A. Iron Deficiency-Induced Changes in the Hippocampus, Corpus Striatum, and Monoamines Levels That Lead to Anxiety, Depression, Sleep Disorders, and Psychotic Disorders. Cureus 2021, 13, e18138. [Google Scholar] [CrossRef]
- Verdon, F.; Burnand, B.; Stubi, C.L.; Bonard, C.; Graff, M.; Michaud, A.; Bischoff, T.; de Vevey, M.; Studer, J.P.; Herzig, L.; et al. Iron supplementation for unexplained fatigue in non-anaemic women: Double blind randomised placebo controlled trial. BMJ 2003, 326, 1124. [Google Scholar] [CrossRef]
- Stewart, R.; Hirani, V. Relationship between depressive symptoms, anemia, and iron status in older residents from a national survey population. Psychosom. Med. 2012, 74, 208–213. [Google Scholar] [CrossRef]
- Yi, S.; Nanri, A.; Poudel-Tandukar, K.; Nonaka, D.; Matsushita, Y.; Hori, A.; Mizoue, T. Association between serum ferritin concentrations and depressive symptoms in Japanese municipal employees. Psychiatry Res. 2011, 189, 368–372. [Google Scholar] [CrossRef] [PubMed]
- Islam, M.R.; Shalahuddin Qusar, M.M.A.; Islam, M.S.; Kabir, M.H.; Mustafizur Rahman, G.K.M.; Hasnat, A. Alterations of serum macro-minerals and trace elements are associated with major depressive disorder: A case-control study. BMC Psychiatry 2018, 18, 94. [Google Scholar] [CrossRef] [PubMed]
- Vahdat Shariatpanaahi, M.; Vahdat Shariatpanaahi, Z.; Moshtaaghi, M.; Shahbaazi, S.H.; Abadi, A. The relationship between depression and serum ferritin level. Eur. J. Clin. Nutr. 2007, 61, 532–535. [Google Scholar] [CrossRef]
- McClung, J.P.; Karl, J.P.; Cable, S.J.; Williams, K.W.; Nindl, B.C.; Young, A.J.; Lieberman, H.R. Randomized, double-blind, placebo-controlled trial of iron supplementation in female soldiers during military training: Effects on iron status, physical performance, and mood. Am. J. Clin. Nutr. 2009, 90, 124–131. [Google Scholar] [CrossRef]
- Abbas, M.; Gandy, K.; Salas, R.; Devaraj, S.; Calarge, C.A. Iron deficiency and internalizing symptom severity in unmedicated adolescents: A pilot study. Psychol. Med. 2021, 53, 2274–2284. [Google Scholar] [CrossRef] [PubMed]
- Lozoff, B.; Jimenez, E.; Hagen, J.; Mollen, E.; Wolf, A.W. Poorer behavioral and developmental outcome more than 10 years after treatment for iron deficiency in infancy. Pediatrics 2000, 105, E51. [Google Scholar] [CrossRef]
- Beard, J.L.; Hendricks, M.K.; Perez, E.M.; Murray-Kolb, L.E.; Berg, A.; Vernon-Feagans, L.; Irlam, J.; Isaacs, W.; Sive, A.; Tomlinson, M. Maternal iron deficiency anemia affects postpartum emotions and cognition. J. Nutr. 2005, 135, 267–272. [Google Scholar] [CrossRef]
- Chang, S.; Wang, L.; Wang, Y.; Brouwer, I.D.; Kok, F.J.; Lozoff, B.; Chen, C. Iron-deficiency anemia in infancy and social emotional development in preschool-aged Chinese children. Pediatrics 2011, 127, e927–e933. [Google Scholar] [CrossRef]
- Coe, C.L.; Lubach, G.R.; Bianco, L.; Beard, J.L. A history of iron deficiency anemia during infancy alters brain monoamine activity later in juvenile monkeys. Dev. Psychobiol. 2009, 51, 301–309. [Google Scholar] [CrossRef]
- Golub, M.S.; Hogrefe, C.E.; Widaman, K.F.; Capitanio, J.P. Iron deficiency anemia and affective response in rhesus monkey infants. Dev. Psychobiol. 2009, 51, 47–59. [Google Scholar] [CrossRef]
- Kukuia, K.K.E.; Torbi, J.; Amoateng, P.; Adutwum-Ofosu, K.K.; Koomson, A.E.; Appiah, F.; Tagoe, T.A.; Mensah, J.A.; Ameyaw, E.O.; Adi-Dako, O.; et al. Gestational iron supplementation reverses depressive-like behavior in post-partum Sprague Dawley rats: Evidence from behavioral and neurohistological studies. IBRO Neurosci. Rep. 2022, 12, 280–296. [Google Scholar] [CrossRef] [PubMed]
- Chen, M.H.; Su, T.P.; Chen, Y.S.; Hsu, J.W.; Huang, K.L.; Chang, W.H.; Chen, T.J.; Bai, Y.M. Association between psychiatric disorders and iron deficiency anemia among children and adolescents: A nationwide population-based study. BMC Psychiatry 2013, 13, 161. [Google Scholar] [CrossRef] [PubMed]
- Beard, J.L.; Erikson, K.M.; Jones, B.C. Neurobehavioral analysis of developmental iron deficiency in rats. Behav. Brain Res. 2002, 134, 517–524. [Google Scholar] [CrossRef] [PubMed]
- Eseh, R.; Zimmerberg, B. Age-dependent effects of gestational and lactational iron deficiency on anxiety behavior in rats. Behav. Brain Res. 2005, 164, 214–221. [Google Scholar] [CrossRef] [PubMed]
- Dobbing, J.; Sands, J. Maternal nutrition and neurological development. Ontogenesis of the Brain. Prague Univ. Karlov. 1974, 2, 167–172. [Google Scholar]
- Texel, S.J.; Camandola, S.; Ladenheim, B.; Rothman, S.M.; Mughal, M.R.; Unger, E.L.; Cadet, J.L.; Mattson, M.P. Ceruloplasmin deficiency results in an anxiety phenotype involving deficits in hippocampal iron, serotonin, and BDNF. J. Neurochem. 2012, 120, 125–134. [Google Scholar] [CrossRef]
- Cutler, P. Iron overload and psychiatric illness. Canadian journal of psychiatry. Rev. Can. Psychiatr. 1994, 39, 8–11. [Google Scholar] [CrossRef]
- Al-Hakeim, H.K.; Najm, A.H.; Al-Dujaili, A.H.; Maes, M. Major Depression in Children with Transfusion-Dependent Thalassemia Is Strongly Associated with the Combined Effects of Blood Transfusion Rate, Iron Overload, and Increased Pro-inflammatory Cytokines. Neurotox. Res. 2020, 38, 228–241. [Google Scholar] [CrossRef]
- Pattanashetti, M.; Mugali, J.; Pattanashetty, N.; Patil, S. A study of severity of depression in thalassemia patients. Int. J. Indian Psychol. 2017, 4, e05. [Google Scholar]
- Yao, S.; Zhong, Y.; Xu, Y.; Qin, J.; Zhang, N.; Zhu, X.; Li, Y. Quantitative Susceptibility Mapping Reveals an Association between Brain Iron Load and Depression Severity. Front. Hum. Neurosci. 2017, 11, 442. [Google Scholar] [CrossRef]
- Wang, F.; Zhang, M.; Li, Y.; Li, Y.; Gong, H.; Li, J.; Zhang, Y.; Zhang, C.; Yan, F.; Sun, B.; et al. Alterations in brain iron deposition with progression of late-life depression measured by magnetic resonance imaging (MRI)-based quantitative susceptibility mapping. Quant Imaging Med. Surg. 2022, 12, 3873–3888. [Google Scholar] [CrossRef] [PubMed]
- Duan, X.; Xie, Y.; Zhu, X.; Chen, L.; Li, F.; Feng, G.; Li, L. Quantitative susceptibility mapping of brain iron deposition in patients with recurrent depression. Psychiatry Investig. 2022, 19, 668. [Google Scholar] [CrossRef] [PubMed]
- Liang, S.; Lu, Y.; Li, Z.; Li, S.; Chen, B.; Zhang, M.; Ji, M.; Gong, W.; Xia, M.; Verkhratsky, A.; et al. Iron Aggravates the Depressive Phenotype of Stressed Mice by Compromising the Glymphatic System. Neurosci. Bull. 2020, 36, 1542–1546. [Google Scholar] [CrossRef]
- Zhang, M.J.; Song, M.L.; Zhang, Y.; Yang, X.M.; Lin, H.S.; Chen, W.C.; Zhong, X.D.; He, C.Y.; Li, T.; Liu, Y.; et al. SNS alleviates depression-like behaviors in CUMS mice by regluating dendritic spines via NCOA4-mediated ferritinophagy. J. Ethnopharmacol. 2023, 312, 116360. [Google Scholar] [CrossRef]
- Weatherall, D.J.; Clegg, J.B. Inherited haemoglobin disorders: An increasing global health problem. Bull. World Health Organ. 2001, 79, 704–712. [Google Scholar]
- Sahu, S.; Agrawal, A.; Shrivastava, J.; Tonk, S. Psychiatric disorders and caregiver burden in children with transfusion dependent beta-thalassaemia and their caregivers. World J. Clin. Pediatr. 2023, 12, 125–132. [Google Scholar] [CrossRef]
- Chen, K.; Zhang, L.; Mao, H.; Chen, K.; Shi, Y.; Meng, X.; Wang, F.; Hu, X.; Fang, X. The impact of iron deposition on the fear circuit of the brain in patients with Parkinson’s disease and anxiety. Front. Aging Neurosci. 2023, 15, 1116516. [Google Scholar] [CrossRef]
- Maaroufi, K.; Ammari, M.; Jeljeli, M.; Roy, V.; Sakly, M.; Abdelmelek, H. Impairment of emotional behavior and spatial learning in adult Wistar rats by ferrous sulfate. Physiol. Behav. 2009, 96, 343–349. [Google Scholar] [CrossRef]
- Chtourou, Y.; Slima, A.B.; Gdoura, R.; Fetoui, H. Naringenin Mitigates Iron-Induced Anxiety-Like Behavioral Impairment, Mitochondrial Dysfunctions, Ectonucleotidases and Acetylcholinesterase Alteration Activities in Rat Hippocampus. Neurochem. Res. 2015, 40, 1563–1575. [Google Scholar] [CrossRef]
- Sobotka, T.J.; Whittaker, P.; Sobotka, J.M.; Brodie, R.E.; Quander, D.Y.; Robl, M.; Bryant, M.; Barton, C.N. Neurobehavioral dysfunctions associated with dietary iron overload. Physiol. Behav. 1996, 59, 213–219. [Google Scholar] [CrossRef]
- Pellegrino, R.M.; Boda, E.; Montarolo, F.; Boero, M.; Mezzanotte, M.; Saglio, G.; Buffo, A.; Roetto, A. Transferrin Receptor 2 Dependent Alterations of Brain Iron Metabolism Affect Anxiety Circuits in the Mouse. Sci. Rep. 2016, 6, 30725. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Zeng, Y.-N.; Yang, P.; Jin, L.-Q.; Xiong, W.-C.; Zhu, M.-Z.; Zhang, J.-Z.; He, X.; Zhu, X.-H. Axonal iron transport in the brain modulates anxiety-related behaviors. Nat. Chem. Biol. 2019, 15, 1214–1222. [Google Scholar] [CrossRef] [PubMed]
- Insel, T.R. Rethinking schizophrenia. Nature 2010, 468, 187–193. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization. Schizophrenia. Available online: https://www.who.int/news-room/fact-sheets/detail/schizophrenia?gclid=Cj0KCQjwhfipBhCqARIsAH9msbnDoRWwwUtm7lZABoRqPR8PXu1VEnpjjohXP_hBixH0RcIpJywe2hMaAgEQEALw_wcB (accessed on 10 January 2023).
- Van Os, J.; Kenis, G.; Rutten, B.P. The environment and schizophrenia. Nature 2010, 468, 203–212. [Google Scholar] [CrossRef] [PubMed]
- Pickard, B. Progress in defining the biological causes of schizophrenia. Expert Rev. Mol. Med. 2011, 13, e25. [Google Scholar] [CrossRef] [PubMed]
- Andreasen, N.C. Understanding the causes of schizophrenia. N. Engl. J. Med. 1999, 340, 645–647. [Google Scholar] [CrossRef]
- Maxwell, A.M.; Rao, R.B. Perinatal iron deficiency as an early risk factor for schizophrenia. Nutr. Neurosci. 2022, 25, 2218–2227. [Google Scholar] [CrossRef]
- Saghazadeh, A.; Mahmoudi, M.; Shahrokhi, S.; Mojarrad, M.; Dastmardi, M.; Mirbeyk, M.; Rezaei, N. Trace elements in schizophrenia: A systematic review and meta-analysis of 39 studies (N = 5151 participants). Nutr. Rev. 2020, 78, 278–303. [Google Scholar] [CrossRef]
- Xu, M.; Guo, Y.; Cheng, J.; Xue, K.; Yang, M.; Song, X.; Feng, Y.; Cheng, J. Brain iron assessment in patients with First-episode schizophrenia using quantitative susceptibility mapping. NeuroImage Clin. 2021, 31, 102736. [Google Scholar] [CrossRef]
- McGrath, J.; Brown, A.; St Clair, D. Prevention and schizophrenia—The role of dietary factors. Schizophr. Bull. 2011, 37, 272–283. [Google Scholar] [CrossRef]
- Sonnenschein, S.F.; Parr, A.C.; Larsen, B.; Calabro, F.J.; Foran, W.; Eack, S.M.; Luna, B.; Sarpal, D.K. Subcortical brain iron deposition in individuals with schizophrenia. J. Psychiatr. Res. 2022, 151, 272–278. [Google Scholar] [CrossRef] [PubMed]
- Ravanfar, P.; Syeda, W.T.; Jayaram, M.; Rushmore, R.J.; Moffat, B.; Lin, A.P.; Lyall, A.E.; Merritt, A.H.; Yaghmaie, N.; Laskaris, L. In Vivo 7-Tesla MRI investigation of brain iron and its metabolic correlates in chronic schizophrenia. Schizophrenia 2022, 8, 86. [Google Scholar] [CrossRef] [PubMed]
- Kornhuber, J.; Lange, K.W.; Kruzik, P.; Rausch, W.-D.; Gabriel, E.; Jellinger, K.; Riederer, P. Iron, copper, zinc, magnesium, and calcium in postmortem brain tissue from schizophrenic patients. Biol. Psychiatry 1994, 36, 31–34. [Google Scholar] [CrossRef] [PubMed]
- Bitanihirwe, B.K.; Woo, T.-U.W. Oxidative stress in schizophrenia: An integrated approach. Neurosci. Biobehav. Rev. 2011, 35, 878–893. [Google Scholar] [CrossRef]
- Kirkpatrick, B.; Miller, B.J. Inflammation and schizophrenia. Schizophr. Bull. 2013, 39, 1174–1179. [Google Scholar] [CrossRef]
- Izquierdo, I.; Furini, C.R.; Myskiw, J.C. Fear Memory. Physiol. Rev. 2016, 96, 695–750. [Google Scholar] [CrossRef]
- Oota-Ishigaki, A.; Takao, K.; Yamada, D.; Sekiguchi, M.; Itoh, M.; Koshidata, Y.; Abe, M.; Natsume, R.; Kaneko, M.; Adachi, T.; et al. Prolonged contextual fear memory in AMPA receptor palmitoylation-deficient mice. Neuropsychopharmacol. Off. Publ. Am. Coll. Neuropsychopharmacol. 2022, 47, 2150–2159. [Google Scholar] [CrossRef]
- Doom, J.R.; Richards, B.; Caballero, G.; Delva, J.; Gahagan, S.; Lozoff, B. Infant Iron Deficiency and Iron Supplementation Predict Adolescent Internalizing, Externalizing, and Social Problems. J. Pediatr. 2018, 195, 199–205.e2. [Google Scholar] [CrossRef]
- McEchron, M.D.; Cheng, A.Y.; Liu, H.; Connor, J.R.; Gilmartin, M.R. Perinatal nutritional iron deficiency permanently impairs hippocampus-dependent trace fear conditioning in rats. Nutr. Neurosci. 2005, 8, 195–206. [Google Scholar] [CrossRef]
- Gewirtz, J.C.; Hamilton, K.L.; Babu, M.A.; Wobken, J.D.; Georgieff, M.K. Effects of gestational iron deficiency on fear conditioning in juvenile and adult rats. Brain Res. 2008, 1237, 195–203. [Google Scholar] [CrossRef][Green Version]
- Guo, S.; Dong, Y.; Cheng, X.; Chen, Z.; Ni, Y.; Zhao, R.; Ma, W. Chronic Psychological Stress Disrupts Iron Metabolism and Enhances Hepatic Mitochondrial Function in Mice. Biol. Trace Elem. Res. 2023, 201, 1761–1771. [Google Scholar] [CrossRef] [PubMed]
- Zhao, M.; Yu, Z.; Zhang, Y.; Huang, X.; Hou, J.; Zhao, Y.; Luo, W.; Chen, L.; Ou, L.; Li, H.; et al. Iron-induced neuronal damage in a rat model of post-traumatic stress disorder. Neuroscience 2016, 330, 90–99. [Google Scholar] [CrossRef] [PubMed]
- Bao, W.D.; Pang, P.; Zhou, X.T.; Hu, F.; Xiong, W.; Chen, K.; Wang, J.; Wang, F.; Xie, D.; Hu, Y.Z.; et al. Loss of ferroportin induces memory impairment by promoting ferroptosis in Alzheimer’s disease. Cell Death Differ. 2021, 28, 1548–1562. [Google Scholar] [CrossRef] [PubMed]
- Schroder, N.; Fredriksson, A.; Vianna, M.R.; Roesler, R.; Izquierdo, I.; Archer, T. Memory deficits in adult rats following postnatal iron administration. Behav. Brain Res. 2001, 124, 77–85. [Google Scholar] [CrossRef]
- Silva, P.F.; Garcia, V.A.; Dornelles Ada, S.; Silva, V.K.; Maurmann, N.; Portal, B.C.; Ferreira, R.D.; Piazza, F.C.; Roesler, R.; Schroder, N. Memory impairment induced by brain iron overload is accompanied by reduced H3K9 acetylation and ameliorated by sodium butyrate. Neuroscience 2012, 200, 42–49. [Google Scholar] [CrossRef]
- Zhong, T.; Qing, Q.J.; Yang, Y.; Zou, W.Y.; Ye, Z.; Yan, J.Q.; Guo, Q.L. Repression of contexual fear memory induced by isoflurane is accompanied by reduction in histone acetylation and rescued by sodium butyrate. Br. J. Anaesth. 2014, 113, 634–643. [Google Scholar] [CrossRef]
- Uberti, V.H.; de Freitas, B.S.; Molz, P.; Bromberg, E.; Schroder, N. Iron Overload Impairs Autophagy: Effects of Rapamycin in Ameliorating Iron-Related Memory Deficits. Mol. Neurobiol. 2020, 57, 1044–1054. [Google Scholar] [CrossRef]
- Wu, J.; Yang, J.J.; Cao, Y.; Li, H.; Zhao, H.; Yang, S.; Li, K. Iron overload contributes to general anaesthesia-induced neurotoxicity and cognitive deficits. J. Neuroinflamm. 2020, 17, 110. [Google Scholar] [CrossRef]
- Zuo, Y.; Li, B.; Xie, J.; Ma, Z.; Thirupathi, A.; Yu, P.; Gao, G.; Zhou, J.; Zhou, C.; Xu, H.; et al. Sevoflurane anesthesia during pregnancy in mice induces cognitive impairment in the offspring by causing iron deficiency and inhibiting myelinogenesis. Neurochem. Int. 2020, 135, 104693. [Google Scholar] [CrossRef]
- Tang, S.; Zhang, G.; Ran, Q.; Nie, L.; Liu, X.; Pan, Z.; He, L. Quantitative susceptibility mapping shows lower brain iron content in children with attention-deficit hyperactivity disorder. Hum. Brain Mapp. 2022, 43, 2495–2502. [Google Scholar] [CrossRef]
- Craig, S.G.; Bondi, B.C.; O’Donnell, K.A.; Pepler, D.J.; Weiss, M.D. ADHD and Exposure to Maltreatment in Children and Youth: A Systematic Review of the Past 10 Years. Curr. Psychiatry Rep. 2020, 22, 79. [Google Scholar] [CrossRef] [PubMed]
- Hoogman, M.; Stolte, M.; Baas, M.; Kroesbergen, E. Creativity and ADHD: A review of behavioral studies, the effect of psychostimulants and neural underpinnings. Neurosci. Biobehav. Rev. 2020, 119, 66–85. [Google Scholar] [CrossRef]
- Chen, Y.; Su, S.; Dai, Y.; Zou, M.; Lin, L.; Qian, L.; Zhou, Q.; Zhang, H.; Liu, M.; Zhao, J.; et al. Quantitative susceptibility mapping reveals brain iron deficiency in children with attention-deficit/hyperactivity disorder: A whole-brain analysis. Eur. Radiol. 2022, 32, 3726–3733. [Google Scholar] [CrossRef] [PubMed]
- Yang, R.; Zhang, Y.; Gao, W.; Lin, N.; Li, R.; Zhao, Z. Blood Levels of Trace Elements in Children with Attention-Deficit Hyperactivity Disorder: Results from a Case-Control Study. Biol. Trace Elem. Res. 2019, 187, 376–382. [Google Scholar] [CrossRef]
- Cortese, S.; Azoulay, R.; Castellanos, F.X.; Chalard, F.; Lecendreux, M.; Chechin, D.; Delorme, R.; Sebag, G.; Sbarbati, A.; Mouren, M.-C.; et al. Brain iron levels in attention-deficit/hyperactivity disorder: A pilot MRI study. World J. Biol. Psychiatry 2012, 13, 223–231. [Google Scholar] [CrossRef] [PubMed]
- Adisetiyo, V.; Jensen, J.H.; Tabesh, A.; Deardorff, R.L.; Fieremans, E.; Di Martino, A.; Gray, K.M.; Castellanos, F.X.; Helpern, J.A. Multimodal MR imaging of brain iron in attention deficit hyperactivity disorder: A noninvasive biomarker that responds to psychostimulant treatment? Radiology 2014, 272, 524–532. [Google Scholar] [CrossRef] [PubMed]
- East, P.L.; Doom, J.R.; Blanco, E.; Burrows, R.; Lozoff, B.; Gahagan, S. Iron Deficiency in Infancy and Sluggish Cognitive Tempo and ADHD Symptoms in Childhood and Adolescence. J. Clin. Child. Adolesc. Psychol. 2023, 52, 259–270. [Google Scholar] [CrossRef] [PubMed]
- Fiani, D.; Engler, S.; Fields, S.; Calarge, C.A. Iron Deficiency in Attention-Deficit Hyperactivity Disorder, Autism Spectrum Disorder, Internalizing and Externalizing Disorders, and Movement Disorders. Child Adolesc. Psychiatr. Clin. N. Am. 2023, 32, 451–467. [Google Scholar] [CrossRef]
- Degremont, A.; Jain, R.; Philippou, E.; Latunde-Dada, G.O. Brain iron concentrations in the pathophysiology of children with attention deficit/hyperactivity disorder: A systematic review. Nutr. Rev. 2020, 79, 615–626. [Google Scholar] [CrossRef]
- Konofal, E.; Lecendreux, M.; Deron, J.; Marchand, M.; Cortese, S.; Zaïm, M.; Mouren, M.C.; Arnulf, I. Effects of iron supplementation on attention deficit hyperactivity disorder in children. Pediatr. Neurol. 2008, 38, 20–26. [Google Scholar] [CrossRef]
- Granero, R.; Pardo-Garrido, A.; Carpio-Toro, I.L.; Ramírez-Coronel, A.A.; Martínez-Suárez, P.C.; Reivan-Ortiz, G.G. The role of iron and zinc in the treatment of adhd among children and adolescents: A systematic review of randomized clinical trials. Nutrients 2021, 13, 4059. [Google Scholar] [CrossRef] [PubMed]
- Lord, C.; Brugha, T.S.; Charman, T.; Cusack, J.; Dumas, G.; Frazier, T.; Jones, E.J.H.; Jones, R.M.; Pickles, A.; State, M.W.; et al. Autism spectrum disorder. Nat. Rev. Dis. Primers 2020, 6, 5. [Google Scholar] [CrossRef] [PubMed]
- Dosman, C.F.; Brian, J.A.; Drmic, I.E.; Senthilselvan, A.; Harford, M.M.; Smith, R.W.; Sharieff, W.; Zlotkin, S.H.; Moldofsky, H.; Roberts, S.W. Children with autism: Effect of iron supplementation on sleep and ferritin. Pediatr. Neurol. 2007, 36, 152–158. [Google Scholar] [CrossRef] [PubMed]
- Herndon, A.C.; DiGuiseppi, C.; Johnson, S.L.; Leiferman, J.; Reynolds, A. Does nutritional intake differ between children with autism spectrum disorders and children with typical development? J. Autism Dev. Disord. 2009, 39, 212–222. [Google Scholar] [CrossRef] [PubMed]
- Millichap, J.G.; Yee, M.M.; Davidson, S.I. Serum ferritin in children with attention-deficit hyperactivity disorder. Pediatr. Neurol. 2006, 34, 200–203. [Google Scholar] [CrossRef] [PubMed]
- Reynolds, A.; Krebs, N.F.; Stewart, P.A.; Austin, H.; Johnson, S.L.; Withrow, N.; Molloy, C.; James, S.J.; Johnson, C.; Clemons, T.; et al. Iron status in children with autism spectrum disorder. Pediatrics 2012, 130 (Suppl. S2), S154–S159. [Google Scholar] [CrossRef]
- Tang, S.; Xu, Y.; Liu, X.; Chen, Z.; Zhou, Y.; Nie, L.; He, L. Quantitative susceptibility mapping shows lower brain iron content in children with autism. Eur. Radiol. 2021, 31, 2073–2083. [Google Scholar] [CrossRef]
- Schmidt, R.J.; Tancredi, D.J.; Krakowiak, P.; Hansen, R.L.; Ozonoff, S. Maternal intake of supplemental iron and risk of autism spectrum disorder. Am. J. Epidemiol. 2014, 180, 890–900. [Google Scholar] [CrossRef]
- Leo, D.; Sorrentino, E.; Volpicelli, F.; Eyman, M.; Greco, D.; Viggiano, D.; di Porzio, U.; Perrone-Capano, C. Altered midbrain dopaminergic neurotransmission during development in an animal model of ADHD. Neurosci. Biobehav. Rev. 2003, 27, 661–669. [Google Scholar] [CrossRef]
- Squillace, M.; Dodero, L.; Federici, M.; Migliarini, S.; Errico, F.; Napolitano, F.; Krashia, P.; Di Maio, A.; Galbusera, A.; Bifone, A.; et al. Dysfunctional dopaminergic neurotransmission in asocial BTBR mice. Transl. Psychiatry 2014, 4, e427. [Google Scholar] [CrossRef]
- Beard, J. Iron deficiency alters brain development and functioning. J. Nutr. 2003, 133, 1468S–1472S. [Google Scholar] [CrossRef] [PubMed]
- Marx, W.; Lane, M.; Hockey, M.; Aslam, H.; Berk, M.; Walder, K.; Borsini, A.; Firth, J.; Pariante, C.M.; Berding, K.; et al. Diet and depression: Exploring the biological mechanisms of action. Mol. Psychiatry 2021, 26, 134–150. [Google Scholar] [CrossRef] [PubMed]
- Beard, J.L.; Connor, J.R. Iron status and neural functioning. Annu. Rev. Nutr. 2003, 23, 41–58. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.; Wessling-Resnick, M. Iron and mechanisms of emotional behavior. J. Nutr. Biochem. 2014, 25, 1101–1107. [Google Scholar] [CrossRef] [PubMed]
- Lozoff, B. Early iron deficiency has brain and behavior effects consistent with dopaminergic dysfunction. J. Nutr. 2011, 141, 740S–746S. [Google Scholar] [CrossRef]
- Felt, B.T.; Beard, J.L.; Schallert, T.; Shao, J.; Aldridge, J.W.; Connor, J.R.; Georgieff, M.K.; Lozoff, B. Persistent neurochemical and behavioral abnormalities in adulthood despite early iron supplementation for perinatal iron deficiency anemia in rats. Behav. Brain Res. 2006, 171, 261–270. [Google Scholar] [CrossRef]
- Beard, J.L.; Felt, B.; Schallert, T.; Burhans, M.; Connor, J.R.; Georgieff, M.K. Moderate iron deficiency in infancy: Biology and behavior in young rats. Behav. Brain Res. 2006, 170, 224–232. [Google Scholar] [CrossRef]
- Frantom, P.A.; Seravalli, J.; Ragsdale, S.W.; Fitzpatrick, P.F. Reduction and oxidation of the active site iron in tyrosine hydroxylase: Kinetics and specificity. Biochemistry 2006, 45, 2372–2379. [Google Scholar] [CrossRef]
- Sarchiapone, M.; Carli, V.; Camardese, G.; Cuomo, C.; Di Giuda, D.; Calcagni, M.L.; Focacci, C.; De Risio, S. Dopamine transporter binding in depressed patients with anhedonia. Psychiatry Res. 2006, 147, 243–248. [Google Scholar] [CrossRef]
- Unger, E.L.; Wiesinger, J.A.; Hao, L.; Beard, J.L. Dopamine D2 receptor expression is altered by changes in cellular iron levels in PC12 cells and rat brain tissue. J. Nutr. 2008, 138, 2487–2494. [Google Scholar] [CrossRef]
- Jellen, L.C.; Lewis, M.M.; Du, G.; Wang, X.; Galvis, M.L.E.; Krzyzanowski, S.; Capan, C.D.; Snyder, A.M.; Connor, J.R.; Kong, L.; et al. Low plasma serotonin linked to higher nigral iron in Parkinson’s disease. Sci. Rep. 2021, 11, 24384. [Google Scholar] [CrossRef] [PubMed]
- Mackler, B.; Person, R.; Miller, L.R.; Inamdar, A.R.; Finch, C.A. Iron deficiency in the rat: Biochemical studies of brain metabolism. Pediatr. Res. 1978, 12, 217–220. [Google Scholar] [CrossRef] [PubMed]
- Shukla, A.; Agarwal, K.N.; Chansuria, J.P.; Taneja, V. Effect of latent iron deficiency on 5-hydroxytryptamine metabolism in rat brain. J. Neurochem. 1989, 52, 730–735. [Google Scholar] [CrossRef] [PubMed]
- Youdim, M.B.; Ben-Shachar, D. Minimal brain damage induced by early iron deficiency: Modified dopaminergic neurotransmission. Isr. J. Med. Sci. 1987, 23, 19–25. [Google Scholar]
- Li, Y.; Kim, J.; Buckett, P.D.; Bohlke, M.; Maher, T.J.; Wessling-Resnick, M. Severe postnatal iron deficiency alters emotional behavior and dopamine levels in the prefrontal cortex of young male rats. J. Nutr. 2011, 141, 2133–2138. [Google Scholar] [CrossRef]
- Kwik-Uribe, C.L.; Golub, M.S.; Keen, C.L. Chronic marginal iron intakes during early development in mice alter brain iron concentrations and behavior despite postnatal iron supplementation. J. Nutr. 2000, 130, 2040–2048. [Google Scholar] [CrossRef]
- Youdim, M.B.; Green, A.R.; Bloomfield, M.R.; Mitchell, B.D.; Heal, D.J.; Grahame-Smith, D.G. The effects of iron deficiency on brain biogenic monoamine biochemistry and function in rats. Neuropharmacology 1980, 19, 259–267. [Google Scholar] [CrossRef]
- Youdim, M.B.; Ben-Shachar, D.; Yehuda, S. Putative biological mechanisms of the effect of iron deficiency on brain biochemistry and behavior. Am. J. Clin. Nutr. 1989, 50, 607–615; discussion 615–607. [Google Scholar] [CrossRef]
- Anderson, J.G.; Fordahl, S.C.; Cooney, P.T.; Weaver, T.L.; Colyer, C.L.; Erikson, K.M. Extracellular norepinephrine, norepinephrine receptor and transporter protein and mRNA levels are differentially altered in the developing rat brain due to dietary iron deficiency and manganese exposure. Brain Res. 2009, 1281, 1–14. [Google Scholar] [CrossRef]
- Kikusui, T.; Nakamura, K.; Kakuma, Y.; Mori, Y. Early weaning augments neuroendocrine stress responses in mice. Behav. Brain Res. 2006, 175, 96–103. [Google Scholar] [CrossRef]
- Kikusui, T.; Ichikawa, S.; Mori, Y. Maternal deprivation by early weaning increases corticosterone and decreases hippocampal BDNF and neurogenesis in mice. Psychoneuroendocrinology 2009, 34, 762–772. [Google Scholar] [CrossRef]
- Wu, L.; Xian, X.; Tan, Z.; Dong, F.; Xu, G.; Zhang, M.; Zhang, F. The Role of Iron Metabolism, Lipid Metabolism, and Redox Homeostasis in Alzheimer’s Disease: From the Perspective of Ferroptosis. Mol. Neurobiol. 2023, 60, 2832–2850. [Google Scholar] [CrossRef] [PubMed]
- Mundra, S.; Tits, J.; Wieland, E.; Angst, U.M. Aerobic and anaerobic oxidation of ferrous ions in near-neutral solutions. Chemosphere 2023, 335, 138955. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.Q.; Zhou, G.N.; Ding, R.R.; Li, Q.; Zhao, H.Q.; Mu, Y. Ferrous ion enhanced Fenton-like degradation of emerging contaminants by sulfidated nanosized zero-valent iron with pH insensitivity. J. Hazard. Mater. 2023, 459, 132229. [Google Scholar] [CrossRef] [PubMed]
- Meneghini, R. Iron Homeostasis, Oxidative Stress, and DNA Damage. Free. Radic. Biol. Med. 1997, 23, 783–792. [Google Scholar] [CrossRef] [PubMed]
- Al-Hakeim, H.K.; Najm, A.H.; Moustafa, S.R.; Maes, M. Construction of an exposure-pathway-phenotype in children with depression due to transfusion-dependent thalassemia: Results of (un)supervised machine learning. J. Affect. Disord. 2021, 282, 644–655. [Google Scholar] [CrossRef]
- Ferreira, A.C.; Santos, T.; Sampaio-Marques, B.; Novais, A.; Mesquita, S.D.; Ludovico, P.; Bernardino, L.; Correia-Neves, M.; Sousa, N.; Palha, J.A.; et al. Lipocalin-2 regulates adult neurogenesis and contextual discriminative behaviours. Mol. Psychiatry 2018, 23, 1031–1039. [Google Scholar] [CrossRef]
- Han, M.; Chang, J.; Kim, J. Loss of divalent metal transporter 1 function promotes brain copper accumulation and increases impulsivity. J. Neurochem. 2016, 138, 918–928. [Google Scholar] [CrossRef]
- Khadrawy, Y.A.; Hosny, E.N.; Magdy, M.; Mohammed, H.S. Antidepressant effects of curcumin-coated iron oxide nanoparticles in a rat model of depression. Eur. J. Pharmacol. 2021, 908, 174384. [Google Scholar] [CrossRef]
- Beard, J.L.; Connor, J.D.; Jones, B.C. Brain iron: Location and function. Prog. Food Nutr. Sci. 1993, 17, 183–221. [Google Scholar]
- Olanow, C.W. An introduction to the free radical hypothesis in Parkinson’s disease. Ann. Neurol. 1992, 32, S2–S9. [Google Scholar] [CrossRef] [PubMed]
- Lam, C.S.; Li, J.J.; Tipoe, G.L.; Youdim, M.B.H.; Fung, M.L. Monoamine oxidase A upregulated by chronic intermittent hypoxia activates indoleamine 2,3-dioxygenase and neurodegeneration. PLoS ONE 2017, 12, e0177940. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Wang, Z.; Cao, J.; Dong, Y.; Chen, Y. Ferroptosis Mechanisms Involved in Hippocampal-Related Diseases. Int. J. Mol. Sci. 2021, 22, 9902. [Google Scholar] [CrossRef]
- Wang, Y.; Wang, S.; Xin, Y.; Zhang, J.; Yang, Z.; Liu, C. Hydrogen sulfide alleviates the anxiety-like and depressive-like behaviors of type 1 diabetic mice via inhibiting inflammation and ferroptosis. Life Sci. 2021, 278, 119551. [Google Scholar] [CrossRef] [PubMed]
- Zucca, F.A.; Segura-Aguilar, J.; Ferrari, E.; Munoz, P.; Paris, I.; Sulzer, D.; Sarna, T.; Casella, L.; Zecca, L. Interactions of iron, dopamine and neuromelanin pathways in brain aging and Parkinson’s disease. Prog. Neurobiol. 2017, 155, 96–119. [Google Scholar] [CrossRef] [PubMed]
- Hakak, Y.; Walker, J.R.; Li, C.; Wong, W.H.; Davis, K.L.; Buxbaum, J.D.; Haroutunian, V.; Fienberg, A.A. Genome-wide expression analysis reveals dysregulation of myelination-related genes in chronic schizophrenia. Proc. Natl. Acad. Sci. USA 2001, 98, 4746–4751. [Google Scholar] [CrossRef]
- Regenold, W.T.; Phatak, P.; Marano, C.M.; Gearhart, L.; Viens, C.H.; Hisley, K.C. Myelin staining of deep white matter in the dorsolateral prefrontal cortex in schizophrenia, bipolar disorder, and unipolar major depression. Psychiatry Res. 2007, 151, 179–188. [Google Scholar] [CrossRef]
- Pajevic, S.; Basser, P.J.; Fields, R.D. Role of myelin plasticity in oscillations and synchrony of neuronal activity. Neuroscience 2014, 276, 135–147. [Google Scholar] [CrossRef]
- Connor, J.R.; Menzies, S.L. Relationship of iron to oligodendrocytes and myelination. Glia 1996, 17, 83–93. [Google Scholar] [CrossRef]
- Yu, G.S.; Steinkirchner, T.M.; Rao, G.A.; Larkin, E.C. Effect of prenatal iron deficiency on myelination in rat pups. Am. J. Pathol. 1986, 125, 620–624. [Google Scholar]
- Schmitt, A.; Simons, M.; Cantuti-Castelvetri, L.; Falkai, P. A new role for oligodendrocytes and myelination in schizophrenia and affective disorders? Eur. Arch. Psychiatry Clin. Neurosci. 2019, 269, 371–372. [Google Scholar] [CrossRef] [PubMed]
- Guo, X.; Jin, X.; Han, K.; Kang, S.; Tian, S.; Lv, X.; Feng, M.; Zheng, H.; Zuo, Y.; Xu, G. Iron promotes neurological function recovery in mice with ischemic stroke through endogenous repair mechanisms. Free. Radic. Biol. Med. 2022, 182, 59–72. [Google Scholar] [CrossRef]
- The Institute for Health Metrics and Evaluation. Global Health Data Exchange (GHDx). Available online: https://vizhub.healthdata.org/gbd-results/ (accessed on 14 May 2022).
- Charlson, F.; van Ommeren, M.; Flaxman, A.; Cornett, J.; Whiteford, H.; Saxena, S. New WHO prevalence estimates of mental disorders in conflict settings: A systematic review and meta-analysis. Lancet 2019, 394, 240–248. [Google Scholar] [CrossRef]
- Moitra, M.; Santomauro, D.; Collins, P.Y.; Vos, T.; Whiteford, H.; Saxena, S.; Ferrari, A.J. The global gap in treatment coverage for major depressive disorder in 84 countries from 2000-2019: A systematic review and Bayesian meta-regression analysis. PLoS Med. 2022, 19, e1003901. [Google Scholar] [CrossRef] [PubMed]
- Farwin, A.; Low, A.; Howard, N.; Yi, H. “My young life, finished already?”: A qualitative study of embedded social stressors and their effects on mental health of low-wage male migrant workers in Singapore. Glob. Health 2023, 19, 47. [Google Scholar] [CrossRef] [PubMed]
- Gordon, J.H.; Tran, K.T.; Visoki, E.; Argabright, S.; DiDomenico, G.E.; Saiegh, E.; Hoffman, K.W.; Erez, G.; Barzilay, R. The Role of Individual Discrimination and Structural Stigma in the Mental Health of Sexual Minority Youth. J. Am. Acad. Child Adolesc. Psychiatry 2023. ahead of print. [Google Scholar] [CrossRef]
- Baron, S.; Cuervo, I.; Shah, D.; Gonzalez, A.; Harari, H.; Flores, D. COVID-19 Infections, Pandemic-Related Social and Economic Impacts, and Changes to Mental and Self-Rated Health Among Latinx Immigrant Housecleaners in New York City: The Safe and Just Cleaners Study. Am. J. Public Health 2023, 113, 893–903. [Google Scholar] [CrossRef] [PubMed]
- Ke, Y.; Ming Qian, Z. Iron misregulation in the brain: A primary cause of neurodegenerative disorders. Lancet Neurol. 2003, 2, 246–253. [Google Scholar] [CrossRef]
- Oshiro, S.; Morioka, M.S.; Kikuchi, M. Dysregulation of iron metabolism in Alzheimer’s disease, Parkinson’s disease, and amyotrophic lateral sclerosis. Adv. Pharmacol. Sci. 2011, 2011, 378278. [Google Scholar] [CrossRef]
- Crapper McLachlan, D.R.; Dalton, A.J.; Kruck, T.P.; Bell, M.Y.; Smith, W.L.; Kalow, W.; Andrews, D.F. Intramuscular desferrioxamine in patients with Alzheimer’s disease. Lancet 1991, 337, 1304–1308. [Google Scholar] [CrossRef]
- Zeng, X.; An, H.; Yu, F.; Wang, K.; Zheng, L.; Zhou, W.; Bao, Y.; Yang, J.; Shen, N.; Huang, D. Benefits of Iron Chelators in the Treatment of Parkinson’s Disease. Neurochem. Res. 2021, 46, 1239–1251. [Google Scholar] [CrossRef]
- Palmer, C.; Roberts, R.L.; Bero, C. Deferoxamine posttreatment reduces ischemic brain injury in neonatal rats. Stroke 1994, 25, 1039–1045. [Google Scholar] [CrossRef] [PubMed]
- Konofal, E.; Cortese, S.; Lecendreux, M.; Arnulf, I.; Mouren, M.C. Effectiveness of iron supplementation in a young child with attention-deficit/hyperactivity disorder. Pediatrics 2005, 116, e732–e734. [Google Scholar] [CrossRef] [PubMed]
- Fisher, J.; Tran, T.; Kriitmaa, K.; Rosenthal, D.; Tran, T. Common perinatal mental disorders in northern Viet Nam: Community prevalence and health care use. Bull. World Health Organ. 2010, 88, 737–745. [Google Scholar] [CrossRef]
- Liu, X.; Lin, C.; Wang, S.; Yu, X.; Jia, Y.; Chen, J. Effects of High Levels of Copper on the Depression-Related Memory Disorders. J. Gerontol. A Biol. Sci. Med. Sci. 2023, 78, 611–618. [Google Scholar] [CrossRef] [PubMed]
- Totten, M.S.; Davenport, T.S.; Edwards, L.F.; Howell, J.M. Trace Minerals and Anxiety: A Review of Zinc, Copper, Iron, and Selenium. Dietetics 2023, 2, 83–103. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Mental Disorders | Participants/Samples | Brain Iron Levels | Methods/Strategies | Key Findings | References |
|---|---|---|---|---|---|
| Depression | Depressed patients | Elevated (putamen, thalamus) | quantitative susceptibility mapping (QSM) | The study indicates the role of excess brain iron in deep gray matter in depression. It also suggests iron may be a potential biomarker for further understanding the pathophysiological mechanism of depression. | [96] |
| Depression | Late-life depressive patients (on antidepressant medication) | Elevated with the progression of depression: medial prefrontal cortex (mPFC), dorsal anterior cingulate cortex (dACC), occipital areas, habenula, brainstem, and cerebellum. | magnetic resonance imaging (MRI)-based QSM | It strengthens the understanding of the progression of brain iron deposition in late-life depression in patients on antidepressant medication and highlight the close relationship between magnetic susceptibility in the medial frontal areas and depression. | [97] |
| Depression | Patients With Recurrent Depression | Elevated (frontal lobes, temporal lobe structures, occipital lobes hippocampal regions, putamen, thalamus, cingulum, and cerebellum) | QSM | Brain iron deposition has been found to be associated with the overall duration of disease onset, rather than the severity of depression. | [98] |
| Depression | Depressive population | Decreased (hypotransferrinemia) | Blood iron detection | The hypotransferrinemia observed in the depressive population could correspond to a new form of brain functional iron deficiency. | [24] |
| Anxiety disorders | PD patients with anxiety | Elevated (ventral mPFC, ventral ACC, precuneus, angular gyrus, middle occipital gyrus, and supplementary motor area (SMA), hippocampus, and substantia nigra) | QSM | Increased iron accumulation in the fear circuit in PD patients with anxiety might contribute to the development of anxiety in PD. | [103] |
| ADHD | ADHD children aged 8–14 years | Decreased estimated brain iron levels in both the right and left thalamus | MRI | Low iron in the thalamus may contribute to ADHD pathophysiology | [142,143] |
| ADHD | ADHD children aged 6–14 years | Deficient iron in bilateral striatums, anterior cingulum, olfactory gyrus, and right lingual gyri | QSM | Brain iron deficiency in these brain regions might be related with ADHD, which might be valuable for further studies. | [140] |
| ASD | children with autism aged 2–3, 3–4, 4–5, and 5–6 years | Decreased iron contents (in caudate nucleus, dentate nucleus, and splenium of the corpus callosum for the 2–3 years group; in the frontal white matter, caudate nucleus, red nucleus, substantia nigra, dentate nucleus, and splenium of the corpus callosum for the 3–4, 4–5, and 5–6 years groups) | MRI enhanced T2*-weighted angiography (ESWAN) sequence scans | The brain iron content of children with autism is lower than that of normal children | [154] |
| Schizophrenia | Patients with first-episode schizophrenia | Decreased iron levels in the bilateral substantia nigra, left red nucleus and left thalamus | QSM effective transverse relaxation rate (R2*) maps | Decreased iron concentration is found in grey matter nuclei of patients with first-episode schizophrenia | [116] |
| Schizophrenia | an adult cohort of individuals with chronic schizophrenia aged 18–65 years | Elevated brain iron (thalamus) | inverse-normalized T2*-weighted contrast (1/nT2*) | Thalamic iron accumulation may act as a potential marker of schizophrenia | [118] |
| Schizophrenia | Post-mortem human brain samples | Elevated brain iron (the prefrontal cortex) | inductively coupled plasma-mass spectrometry (ICP-MS) Western Blots | It provides a pathophysiologic link between perturbed cortical iron biology and schizophrenia and indicates that achievement of optimal cortical iron homeostasis could offer a new therapeutic target | [60] |
| Schizophrenia | individuals with chronic schizophrenia | Increased iron in the putamen | ultra-high field 7 T QSM magnetic resonance spectroscopy (MRS) | Elevated iron levels in the dorsal striatum may be associated with a network-wide impact on iron distribution within other brain regions. | [119] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wu, Q.; Ren, Q.; Meng, J.; Gao, W.-J.; Chang, Y.-Z. Brain Iron Homeostasis and Mental Disorders. Antioxidants 2023, 12, 1997. https://doi.org/10.3390/antiox12111997
Wu Q, Ren Q, Meng J, Gao W-J, Chang Y-Z. Brain Iron Homeostasis and Mental Disorders. Antioxidants. 2023; 12(11):1997. https://doi.org/10.3390/antiox12111997
Chicago/Turabian StyleWu, Qiong, Qiuyang Ren, Jingsi Meng, Wei-Juan Gao, and Yan-Zhong Chang. 2023. "Brain Iron Homeostasis and Mental Disorders" Antioxidants 12, no. 11: 1997. https://doi.org/10.3390/antiox12111997
APA StyleWu, Q., Ren, Q., Meng, J., Gao, W.-J., & Chang, Y.-Z. (2023). Brain Iron Homeostasis and Mental Disorders. Antioxidants, 12(11), 1997. https://doi.org/10.3390/antiox12111997