
Sarcopenia Is Associated with Changes in Circulating Markers of Antioxidant/Oxidant Balance and Innate Immune Response

 ,
,  , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Population and Design
2.2. Assessment of Sarcopenia
2.3. Laboratory Measurements
2.4. RNA Isolation and Quantitative Real-Time Reverse-Transcription Polymerase Chain Reaction (qRT-PCR)
2.5. Statistical Analysis
3. Results
3.1. Baseline Characteristics
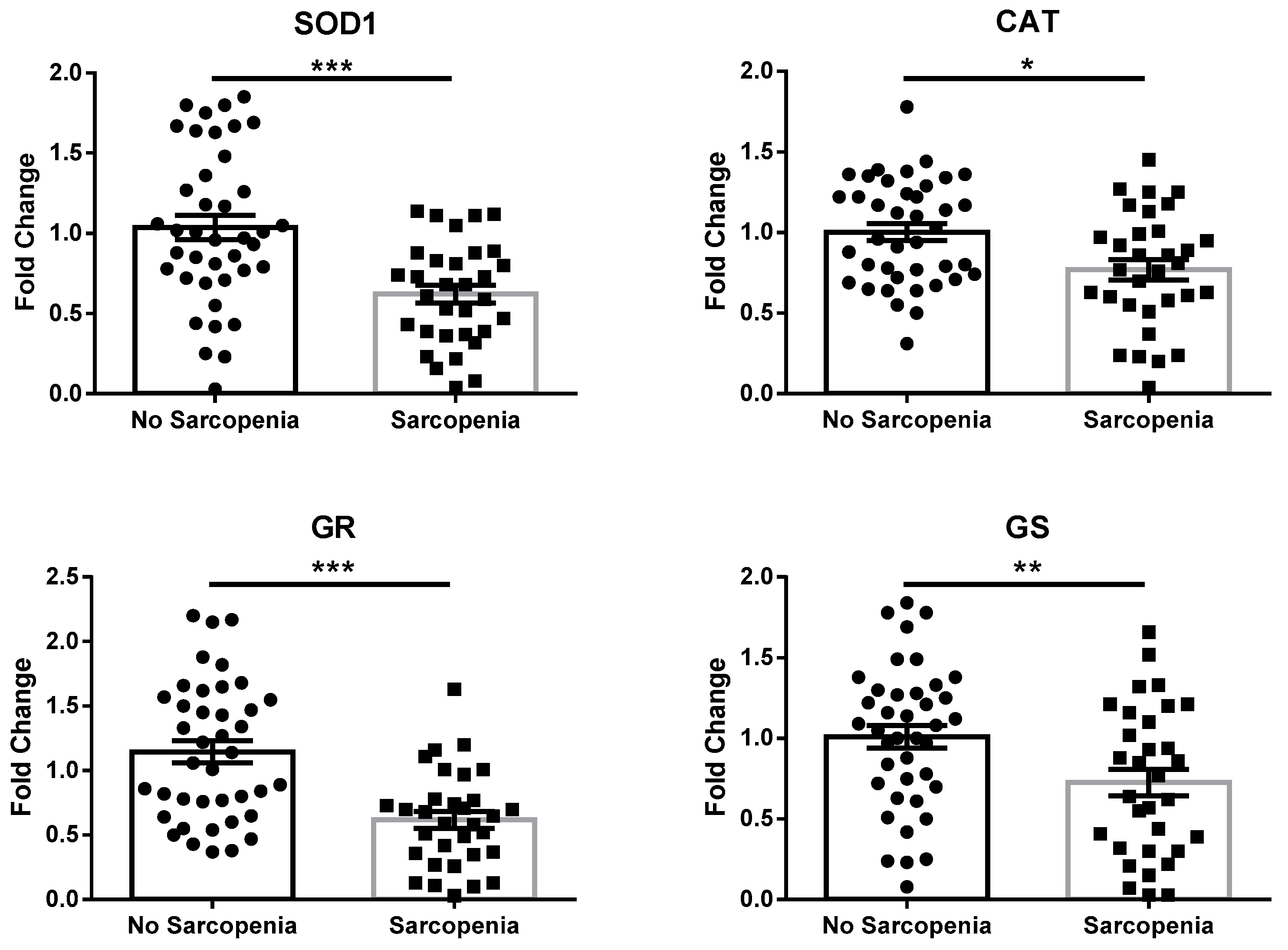
3.2. Circulating Markers of Redox Balance and Immunity Are Altered in Sarcopenia
3.3. Circulating Markers of Redox Homeostasis Are Associated with Innate Immune Response in Sarcopenia
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Iburg, K.M.; Charalampous, P.; Allebeck, P.; Stenberg, E.J.; O’Caoimh, R.; Monasta, L.; Penalvo, J.L.; Pereira, D.M.; Wyper, G.M.A.; Niranjan, V.; et al. Burden of disease among older adults in Europe-trends in mortality and disability, 1990–2019. Eur. J. Public Health 2023, 33, 121–126. [Google Scholar] [CrossRef] [PubMed]
- Cruz-Jentoft, A.J.; Bahat, G.; Bauer, J.; Boirie, Y.; Bruyere, O.; Cederholm, T.; Cooper, C.; Landi, F.; Rolland, Y.; Sayer, A.A.; et al. Sarcopenia: Revised European consensus on definition and diagnosis. Age Ageing 2019, 48, 601. [Google Scholar] [CrossRef] [PubMed]
- Beaudart, C.; Zaaria, M.; Pasleau, F.; Reginster, J.Y.; Bruyere, O. Health Outcomes of Sarcopenia: A Systematic Review and Meta-Analysis. PLoS ONE 2017, 12, e0169548. [Google Scholar] [CrossRef] [PubMed]
- Antuna, E.; Cachan-Vega, C.; Bermejo-Millo, J.C.; Potes, Y.; Caballero, B.; Vega-Naredo, I.; Coto-Montes, A.; Garcia-Gonzalez, C. Inflammaging: Implications in Sarcopenia. Int. J. Mol. Sci. 2022, 23, 15039. [Google Scholar] [CrossRef] [PubMed]
- Liang, Z.; Zhang, T.; Liu, H.; Li, Z.; Peng, L.; Wang, C.; Wang, T. Inflammaging: The ground for sarcopenia? Exp. Gerontol. 2022, 168, 111931. [Google Scholar] [CrossRef]
- Baylis, D.; Bartlett, D.B.; Patel, H.P.; Roberts, H.C. Understanding how we age: Insights into inflammaging. Longev. Health 2013, 2, 8. [Google Scholar] [CrossRef]
- Rong, Y.D.; Bian, A.L.; Hu, H.Y.; Ma, Y.; Zhou, X.Z. Study on relationship between elderly sarcopenia and inflammatory cytokine IL-6, anti-inflammatory cytokine IL-10. BMC Geriatr. 2018, 18, 308. [Google Scholar] [CrossRef] [PubMed]
- Wumaer, A.; Maimaitiwusiman, Z.; Xiao, W.; Xuekelati, S.; Liu, J.; Musha, T.; Wang, H. Plasma tumor necrosis factor-alpha is associated with sarcopenia in elderly individuals residing in agricultural and pastoral areas of Xinjiang, China. Front. Med. 2022, 9, 788178. [Google Scholar] [CrossRef]
- Romano, A.D.; Serviddio, G.; de Mattaheis, A.; Bellanti, F.; Vendemiale, G. Oxidative stress and aging. J. Nephrol. 2010, 23 (Suppl. S15), S29–S36. [Google Scholar]
- Chandrasekaran, A.; Idelchik, M.D.P.S.; Melendez, J.A. Redox control of senescence and age-related disease. Redox Biol. 2017, 11, 91–102. [Google Scholar] [CrossRef] [PubMed]
- Tretter, V.; Hochreiter, B.; Zach, M.L.; Krenn, K.; Klein, K.U. Understanding Cellular Redox Homeostasis: A Challenge for Precision Medicine. Int. J. Mol. Sci. 2021, 23, 106. [Google Scholar] [CrossRef] [PubMed]
- Lian, D.; Chen, M.M.; Wu, H.; Deng, S.; Hu, X. The Role of Oxidative Stress in Skeletal Muscle Myogenesis and Muscle Disease. Antioxidants 2022, 11, 755. [Google Scholar] [CrossRef] [PubMed]
- Bellanti, F.; Romano, A.D.; Lo Buglio, A.; Castriotta, V.; Guglielmi, G.; Greco, A.; Serviddio, G.; Vendemiale, G. Oxidative stress is increased in sarcopenia and associated with cardiovascular disease risk in sarcopenic obesity. Maturitas 2018, 109, 6–12. [Google Scholar] [CrossRef]
- Pigna, E.; Greco, E.; Morozzi, G.; Grottelli, S.; Rotini, A.; Minelli, A.; Fulle, S.; Adamo, S.; Mancinelli, R.; Bellezza, I.; et al. Denervation does not Induce Muscle Atrophy Through Oxidative Stress. Eur. J. Transl. Myol. 2017, 27, 6406. [Google Scholar] [CrossRef]
- Bellanti, F.; Lo Buglio, A.; Vendemiale, G. Redox Homeostasis and Immune Alterations in Coronavirus Disease-19. Biology 2022, 11, 159. [Google Scholar] [CrossRef] [PubMed]
- Morris, G.; Gevezova, M.; Sarafian, V.; Maes, M. Redox regulation of the immune response. Cell Mol. Immunol. 2022, 19, 1079–1101. [Google Scholar] [CrossRef]
- Meng, S.J.; Yu, L.J. Oxidative stress, molecular inflammation and sarcopenia. Int. J. Mol. Sci. 2010, 11, 1509–1526. [Google Scholar] [CrossRef]
- Chen, M.; Wang, Y.; Deng, S.; Lian, Z.; Yu, K. Skeletal muscle oxidative stress and inflammation in aging: Focus on antioxidant and anti-inflammatory therapy. Front. Cell Dev. Biol. 2022, 10, 964130. [Google Scholar] [CrossRef]
- Dodds, R.M.; Syddall, H.E.; Cooper, R.; Benzeval, M.; Deary, I.J.; Dennison, E.M.; Der, G.; Gale, C.R.; Inskip, H.M.; Jagger, C.; et al. Grip strength across the life course: Normative data from twelve British studies. PLoS ONE 2014, 9, e113637. [Google Scholar] [CrossRef]
- Gould, H.; Brennan, S.L.; Kotowicz, M.A.; Nicholson, G.C.; Pasco, J.A. Total and appendicular lean mass reference ranges for Australian men and women: The Geelong osteoporosis study. Calcif. Tissue Int. 2014, 94, 363–372. [Google Scholar] [CrossRef]
- Cruz-Jentoft, A.J.; Baeyens, J.P.; Bauer, J.M.; Boirie, Y.; Cederholm, T.; Landi, F.; Martin, F.C.; Michel, J.P.; Rolland, Y.; Schneider, S.M.; et al. Sarcopenia: European consensus on definition and diagnosis: Report of the European Working Group on Sarcopenia in Older People. Age Ageing 2010, 39, 412–423. [Google Scholar] [CrossRef]
- Serviddio, G.; Romano, A.D.; Greco, A.; Rollo, T.; Bellanti, F.; Altomare, E.; Vendemiale, G. Frailty syndrome is associated with altered circulating redox balance and increased markers of oxidative stress. Int. J. Immunopathol. Pharmacol. 2009, 22, 819–827. [Google Scholar] [CrossRef] [PubMed]
- Tsuchida, M.; Miura, T.; Mizutani, K.; Aibara, K. Fluorescent substances in mouse and human sera as a parameter of in vivo lipid peroxidation. Biochim. Biophys. Acta 1985, 834, 196–204. [Google Scholar]
- Molloy, R.M.; Mc Connell, R.I.; Lamont, J.V.; FitzGerald, S.P. Automation of biochip array technology for quality results. Clin. Chem. Lab. Med. 2005, 43, 1303–1313. [Google Scholar] [CrossRef] [PubMed]
- Kregel, K.C.; Zhang, H.J. An integrated view of oxidative stress in aging: Basic mechanisms, functional effects, and pathological considerations. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2007, 292, R18–R36. [Google Scholar] [CrossRef]
- Moylan, J.S.; Reid, M.B. Oxidative stress, chronic disease, and muscle wasting. Muscle Nerve 2007, 35, 411–429. [Google Scholar] [CrossRef] [PubMed]
- Siu, P.M.; Pistilli, E.E.; Alway, S.E. Age-dependent increase in oxidative stress in gastrocnemius muscle with unloading. J. Appl. Physiol. 2008, 105, 1695–1705. [Google Scholar] [CrossRef] [PubMed]
- Muller, F.L.; Song, W.; Jang, Y.C.; Liu, Y.; Sabia, M.; Richardson, A.; Van, R.H. Denervation-induced skeletal muscle atrophy is associated with increased mitochondrial ROS production. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2007, 293, R1159–R1168. [Google Scholar] [CrossRef]
- Pollock, N.; Staunton, C.A.; Vasilaki, A.; McArdle, A.; Jackson, M.J. Denervated muscle fibers induce mitochondrial peroxide generation in neighboring innervated fibers: Role in muscle aging. Free Radic. Biol. Med. 2017, 112, 84–92. [Google Scholar] [CrossRef]
- Schafer, F.Q.; Buettner, G.R. Redox environment of the cell as viewed through the redox state of the glutathione disulfide/glutathione couple. Free Radic. Biol. Med. 2001, 30, 1191–1212. [Google Scholar] [CrossRef]
- Berndt, C.; Lillig, C.H.; Flohe, L. Redox regulation by glutathione needs enzymes. Front. Pharmacol. 2014, 5, 168. [Google Scholar] [CrossRef]
- Meister, A.; Tate, S.S. Glutathione and related gamma-glutamyl compounds: Biosynthesis and utilization. Annu. Rev. Biochem. 1976, 45, 559–604. [Google Scholar] [CrossRef]
- Couto, N.; Wood, J.; Barber, J. The role of glutathione reductase and related enzymes on cellular redox homoeostasis network. Free Radic. Biol. Med. 2016, 95, 27–42. [Google Scholar] [CrossRef] [PubMed]
- Mosoni, L.; Breuille, D.; Buffiere, C.; Obled, C.; Mirand, P.P. Age-related changes in glutathione availability and skeletal muscle carbonyl content in healthy rats. Exp. Gerontol. 2004, 39, 203–210. [Google Scholar] [CrossRef] [PubMed]
- Sanchez-Castellano, C.; Martin-Aragon, S.; Bermejo-Bescos, P.; Vaquero-Pinto, N.; Miret-Corchado, C.; Merello de Miguel, A.; Cruz-Jentoft, A.J. Biomarkers of sarcopenia in very old patients with hip fracture. J. Cachexia Sarcopenia Muscle 2020, 11, 478–486. [Google Scholar] [CrossRef] [PubMed]
- Rolfe, D.F.S.; Brown, G.C. Cellular energy utilization and molecular origin of standard metabolic rate in mammals. Physiol. Rev. 1997, 77, 731–758. [Google Scholar] [CrossRef] [PubMed]
- Chan, K.M.; Decker, E.A. Endogenous skeletal muscle antioxidants. Crit. Rev. Food Sci. Nutr. 1994, 34, 403–426. [Google Scholar] [CrossRef] [PubMed]
- Deepa, S.S.; Van, R.H.; Brooks, S.V.; Faulkner, J.A.; Larkin, L.; McArdle, A.; Jackson, M.J.; Vasilaki, A.; Richardson, A. Accelerated sarcopenia in Cu/Zn superoxide dismutase knockout mice. Free Radic. Biol. Med. 2019, 132, 19–23. [Google Scholar] [CrossRef]
- Umanskaya, A.; Santulli, G.; Xie, W.; Andersson, D.C.; Reiken, S.R.; Marks, A.R. Genetically enhancing mitochondrial antioxidant activity improves muscle function in aging. Proc. Natl. Acad. Sci. USA 2014, 111, 15250–15255. [Google Scholar] [CrossRef] [PubMed]
- Bano, G.; Trevisan, C.; Carraro, S.; Solmi, M.; Luchini, C.; Stubbs, B.; Manzato, E.; Sergi, G.; Veronese, N. Inflammation and sarcopenia: A systematic review and meta-analysis. Maturitas 2017, 96, 10–15. [Google Scholar] [CrossRef]
- Franceschi, C.; Garagnani, P.; Parini, P.; Giuliani, C.; Santoro, A. Inflammaging: A new immune-metabolic viewpoint for age-related diseases. Nat. Rev. Endocrinol. 2018, 14, 576–590. [Google Scholar] [CrossRef]
- Asoudeh, F.; Dashti, F.; Raeesi, S.; Heshmat, R.; Bidkhori, M.; Jalilian, Z.; Hashemi, R. Inflammatory cytokines and sarcopenia in Iranian adults-results from SARIR study. Sci. Rep. 2022, 12, 5471. [Google Scholar] [CrossRef]
- da Costa Teixeira, L.A.; Avelar, N.C.P.; Peixoto, M.F.D.; Parentoni, A.N.; Santos, J.M.D.; Pereira, F.S.M.; Danielewicz, A.L.; Leopoldino, A.A.O.; Costa, S.P.; Arrieiro, A.N.; et al. Inflammatory biomarkers at different stages of Sarcopenia in older women. Sci. Rep. 2023, 13, 10367. [Google Scholar] [CrossRef] [PubMed]
- Pawelec, G. Age and immunity: What is “immunosenescence”? Exp. Gerontol. 2018, 105, 4–9. [Google Scholar] [CrossRef] [PubMed]
- Palla, A.R.; Ravichandran, M.; Wang, Y.X.; Alexandrova, L.; Yang, A.V.; Kraft, P.; Holbrook, C.A.; Schurch, C.M.; Ho, A.T.V.; Blau, H.M. Inhibition of prostaglandin-degrading enzyme 15-PGDH rejuvenates aged muscle mass and strength. Science 2021, 371, eabc8059. [Google Scholar] [CrossRef]
- Jia, H.; Huang, W.; Liu, C.; Tang, S.; Zhang, J.; Chen, C.; Tian, Y.; Zhong, W. Immunosenescence is a therapeutic target for frailty in older adults: A narrative review. Ann. Transl. Med. 2022, 10, 1142. [Google Scholar] [CrossRef]
- Wilson, D.; Jackson, T.; Sapey, E.; Lord, J.M. Frailty and sarcopenia: The potential role of an aged immune system. Ageing Res. Rev. 2017, 36, 1–10. [Google Scholar] [CrossRef]
- Barbe-Tuana, F.; Funchal, G.; Schmitz, C.R.R.; Maurmann, R.M.; Bauer, M.E. The interplay between immunosenescence and age-related diseases. Semin. Immunopathol. 2020, 42, 545–557. [Google Scholar] [CrossRef] [PubMed]
- Song, M.; Graubard, B.I.; Rabkin, C.S.; Engels, E.A. Neutrophil-to-lymphocyte ratio and mortality in the United States general population. Sci. Rep. 2021, 11, 464. [Google Scholar] [CrossRef] [PubMed]
- Lacy, P.; Stow, J.L. Cytokine release from innate immune cells: Association with diverse membrane trafficking pathways. Blood 2011, 118, 9–18. [Google Scholar] [CrossRef] [PubMed]
- Deshmane, S.L.; Kremlev, S.; Amini, S.; Sawaya, B.E. Monocyte chemoattractant protein-1 (MCP-1): An overview. J. Interferon Cytokine Res. 2009, 29, 313–326. [Google Scholar] [CrossRef] [PubMed]
- Turkowski, K.; Brandenburg, S.; Mueller, A.; Kremenetskaia, I.; Bungert, A.D.; Blank, A.; Felsenstein, M.; Vajkoczy, P. VEGF as a modulator of the innate immune response in glioblastoma. Glia 2018, 66, 161–174. [Google Scholar] [CrossRef] [PubMed]
- Lingappan, K. NF-kappaB in Oxidative Stress. Curr. Opin. Toxicol. 2018, 7, 81–86. [Google Scholar] [CrossRef] [PubMed]
- Petermann-Rocha, F.; Balntzi, V.; Gray, S.R.; Lara, J.; Ho, F.K.; Pell, J.P.; Celis-Morales, C. Global prevalence of sarcopenia and severe sarcopenia: A systematic review and meta-analysis. J. Cachexia Sarcopenia Muscle 2022, 13, 86–99. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
| Variable | No Sarcopenia n = 40 (55.6%) | Sarcopenia n = 32 (44.4%) | p Value |
|---|---|---|---|
| Age, years | 74.8 (±5.1) | 76.4 (±4.5) | 0.168 |
| F Sex, n (%) | 14 (45.0%) | 10 (31.2%) | 0.737 |
| Smoking, n (%) | 10 (25.0%) | 12 (37.5%) | 0.252 |
| Alcohol consumer, n (%) | 21 (52.5%) | 17 (53.1%) | 0.958 |
| Low physical activity, n (%) | 8 (20%) | 9 (28.1%) | 0.420 |
| Diabetes mellitus, n (%) | 22 (55.0%) | 19 (59.3%) | 0.709 |
| Hyperlipidemia, n (%) | 20 (50%) | 20 (62.5%) | 0.289 |
| Hypertension, n (%) | 28 (70.0%) | 20 (62.5%) | 0.502 |
| Chronic liver disease, n (%) | 1 (2.5%) | 2 (6.2%) | 0.429 |
| Chronic kidney disease, n (%) | 5 (12.5%) | 8 (25%) | 0.171 |
| Co-morbidities > 3, n (%) | 6 (15%) | 9 (28.1%) | 0.173 |
| Pharmacotherapy > 6, n (%) | 6 (15%) | 10 (31.2%) | 0.099 |
| Weight, kg | 70.3 (±7.4) | 71.9 (±6.4) | 0.456 |
| BMI, kg/m2 | 28.6 (±2.8) | 26.1 (±1.9) | 0.024 |
| Waist circumference, cm | 108.2 (±15.4) | 100.4 (±11.9) | 0.021 |
| Tricipital fold, cm | 20.4 (±9.6) | 18.2 (±7.9) | 0.300 |
| Hemoglobin, g/dL | 12.4 (±2.1) | 9.5 (±2.2) | <0.001 |
| Glucose, mg/dL | 109.4 [72.0, 134.2] | 115.7 [75.7, 143.1] | 0.707 |
| HbA1c, % | 6.67 [5.97, 8.40] | 6.16 [5.52, 7.84] | 0.358 |
| Insulin, μUI/mL | 6.1 [1.1, 12.8] | 18.0 [4.1, 36.7] | <0.001 |
| HOMA-IR index | 1.78 (±0.18) | 5.44 (±0.87) | <0.001 |
| Total cholesterol, mg/dL | 157.7 [110.6, 203.8] | 132.4 [100.7, 177.5] | 0.013 |
| LDL-cholesterol, mg/dL | 99.2 [56.8, 135.0] | 82.2 [52.8, 117.2] | 0.063 |
| HDL-cholesterol, mg/dL | 49.3 [26.0, 72.1] | 37.1 [19.6, 59.5] | 0.020 |
| Triglycerides, mg/dL | 107.6 (±43.0) | 96.9 (±60.1) | 0.408 |
| Creatinine, mg/dL | 1.29 [0.44, 1.67] | 1.68 [1.26, 2.31] | 0.020 |
| Albumin, g/dL | 3.12 (±0.62) | 2.8 (±0.5) | 0.450 |
| 25-(OH)-vitamin D, ng/mL | 29.1 [10.1, 51.8] | 15.1 [5.5, 39.8] | 0.012 |
| Variable | No Sarcopenia n = 40 (55.6%) | Sarcopenia n = 32 (44.4%) | p Value |
|---|---|---|---|
| Handgrip strength, kg | 26.8 ± 9.1 | 16.2 ± 9.2 | <0.001 |
| ASM/height2, kg/m2 | 9.4 ± 2.3 | 5.2 ± 1.9 | <0.001 |
| Gait speed, m/s | 0.9 [0.4, 1.5] | 0.4 [0.1, 0.9] | <0.001 |
| ADL, score | 6 [5, 6] | 4 [1, 6] | <0.001 |
| IADL, score | 5 [4, 8] | 2 [1, 5] | <0.001 |
| MNA, score | 22.6 ± 6.2 | 17.9 ± 5.9 | 0.002 |
| GDS-SF, score | 3 [1, 6] | 5 [3, 7] | 0.001 |
| MMSE, score | 24.1 ± 4.9 | 16.7 ± 8.4 | <0.001 |
| Variable | No Sarcopenia n = 40 (55.6%) | Sarcopenia n = 32 (44.4%) | p Value |
|---|---|---|---|
| GSH, μM | 60.6 (±25.1) | 36.4 (±24.5) | <0.001 |
| GSSG, μM | 5.14 (±3.21) | 6.43 (±3.17) | 0.093 |
| GSSG/GSH, % | 8.42 (±4.75) | 17.7 (±10.6) | <0.001 |
| HNE–protein adducts (AUF) | 25.4 (±19.8) | 49.6 (±28.8) | <0.001 |
| MDA–protein adducts (AUF) | 58.9 (±45.4) | 89.9 (±53.2) | 0.009 |
| Variable | No Sarcopenia n = 40 (55.6%) | Sarcopenia n = 32 (44.4%) | p Value |
|---|---|---|---|
| ESR, mm/h | 10.4 [4.3, 21.2] | 23.8 [2.1, 89.4] | 0.032 |
| CRP, mg/L | 4.98 [1.12, 12.6] | 21.5 [3.46, 64.8] | 0.019 |
| Fibrinogen, mg/dL | 208.4 (±184.5) | 417.3 (±224.0) | <0.001 |
| Ferritin, ng/mL | 314.6 (±118.3) | 524.1 (±334.7) | <0.001 |
| α2-globulins, g/dL | 5.46 (±1.89) | 9.64 (±6.82) | <0.001 |
| γ-globulins, g/dL | 0.72 (±0.55) | 0.91 (±0.80) | 0.237 |
| WBC, n/mm3 | 6762 [4412, 14,131] | 8291 [4016, 15,322] | 0.383 |
| Neutrophils, n/mm3 | 3814 [1964, 7852] | 5829 [2366, 9983] | 0.025 |
| Lymphocytes, n/mm3 | 1098 (612, 3056] | 1791 [534, 3827] | 0.057 |
| Monocytes, n/mm3 | 0.77 [0.12–1.34] | 0.53 [0.08–1.21] | 0.129 |
| NLR | 2.19 (±1.94) | 3.25 (±2.52) | 0.047 |
| No Sarcopenia n = 40 (55.6%) | Sarcopenia n = 32 (44.4%) | p Value | |
|---|---|---|---|
| IL-1α, pg/mL | 0.96 [0.50, 1.95] | 0.73 [0.50, 1.58] | 0.206 |
| IL-1β, pg/mL | 0.95 [0.44, 1.96] | 0.82 [0.46, 1.84] | 0.503 |
| IL-2, pg/mL | 1.15 (±0.65) | 1.21 (±0.61) | 0.630 |
| IL-4, pg/mL | 1.34 [0.10, 2.64] | 0.10 [0.00, 0.10] | 0.002 |
| IL-6, pg/mL | 15.6 (±12.9) | 25.7 (±26.2) | 0.036 |
| IL-8, pg/mL | 529 [128, 1457] | 269 [46, 804] | 0.101 |
| IL-10, pg/mL | 1.00 (±0.82) | 1.41 (±1.17) | 0.085 |
| TNF, pg/mL | 3.31 (±2.07) | 4.53 (±1.59) | 0.008 |
| IFN-γ, pg/mL | 0.28 (±0.33) | 0.35 (±0.39) | 0.555 |
| VEGF, pg/mL | 292 (±115) | 417 (±182) | <0.001 |
| EGF, pg/mL | 10.0 (±14.4) | 14.9 (±13.8) | 0.148 |
| MCP-1, pg/mL | 306 (±129) | 414 (±163) | 0.002 |
| Predictor (per SD Increase) | Change (95% C.I.) in GSSG/GSH | p Value | R2 |
|---|---|---|---|
| Handgrip strength | +2.3 (+0.51, +4.5) | 0.04 | 0.30 |
| ASM/height2 | +1.9 (−0.6, +4.3) | 0.12 | 0.02 |
| Gait speed | +0.9 (−1.8, +3.6) | 0.49 | 0.00 |
| Neutrophils | +1.5 (+0.2, +2.8) | 0.07 | 0.01 |
| NLR | +4.8 (+1.2, +6.8) | 0.002 | 0.42 |
| IL-1α | +2.1 (+0.1, +4.3) | 0.06 | 0.03 |
| IL-6 | +2.9 (+0.6, +5.2) | 0.02 | 0.15 |
| TNF | +1.3 (+0.2, +2.7) | 0.09 | 0.01 |
| VEGF | +0.5 (−2.7, +1.8) | 0.39 | 0.00 |
| MCP-1 | +0.3 (−0.2, +2.8) | 0.48 | 0.00 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bellanti, F.; Lo Buglio, A.; Quiete, S.; Dobrakowski, M.; Kasperczyk, A.; Kasperczyk, S.; Vendemiale, G. Sarcopenia Is Associated with Changes in Circulating Markers of Antioxidant/Oxidant Balance and Innate Immune Response. Antioxidants 2023, 12, 1992. https://doi.org/10.3390/antiox12111992
Bellanti F, Lo Buglio A, Quiete S, Dobrakowski M, Kasperczyk A, Kasperczyk S, Vendemiale G. Sarcopenia Is Associated with Changes in Circulating Markers of Antioxidant/Oxidant Balance and Innate Immune Response. Antioxidants. 2023; 12(11):1992. https://doi.org/10.3390/antiox12111992
Chicago/Turabian StyleBellanti, Francesco, Aurelio Lo Buglio, Stefano Quiete, Michał Dobrakowski, Aleksandra Kasperczyk, Sławomir Kasperczyk, and Gianluigi Vendemiale. 2023. "Sarcopenia Is Associated with Changes in Circulating Markers of Antioxidant/Oxidant Balance and Innate Immune Response" Antioxidants 12, no. 11: 1992. https://doi.org/10.3390/antiox12111992
APA StyleBellanti, F., Lo Buglio, A., Quiete, S., Dobrakowski, M., Kasperczyk, A., Kasperczyk, S., & Vendemiale, G. (2023). Sarcopenia Is Associated with Changes in Circulating Markers of Antioxidant/Oxidant Balance and Innate Immune Response. Antioxidants, 12(11), 1992. https://doi.org/10.3390/antiox12111992